
Abstract
The breeding of crop plants is a highly effective means of increasing agricultural productivity in a sustainable and environmentally safe way. Prebreeding and population improvement not only capture essential genetic resources and move desired traits along variety development pipelines but also help assure the creation of broad and dynamic gene pools to meet future, unanticipated needs. To efficiently meet multiple breeding objectives requires both interdisciplinary collaboration and a grasp of a wide range of scientific knowledge and expertise. This chapter addresses a range of topics that define and govern potato breeding, drawing from the experiences of both international and regional potato breeding programs, to orient readers to the interlinked components of population improvement and variety development. Using a case study approach to discuss breeding objectives together with respective implications for breeding needs, methods, and awareness-raising approaches for impact, we detail some key research and achievements contributing to current state of the art. Major populations under improvement at the International Potato Center along with breeding objectives and trait levels selected are described in terms of the agroecologies or uses they address in developing country national programs; these are contrasted with a discussion of the Cornell University program that is oriented to the northeastern US. A sample stage gate process, accelerated multi-trait selection schemes, heritability and heterosis exploitation, genomic selection, data management, and end user consultations are introduced in the contexts of these two programs. The topic of this chapter is supported and augmented with further details on subjects closely related to potato breeding, provided in chapters contributed to this volume by Ortiz and Mihovilovich, Ghislain and Douches, Burgos et al., and Ellis et al. The authors hope that the content serves to orient researchers and managers in countries with different degrees of development to plan and succeed in impactful potato improvement programs.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Biofortification
- Population improvement
- Polyploid crop breeding
- True hybrid potato breeding
- Genome-wide association study
- Product Profiles
- Variety development
1 Implications of Genetics, Genepools, and Biology for Potato Breeding
1.1 Key Features
Commercial potato of world importance is a heterozygous, autotetraploid, clonal crop (2n = 4x = 48). Modern varieties are the products of extensive breeding between different cultivar groups and wild species. Potato varieties grown outside of South America since the end of the sixteenth century, as well as landrace (indigenous) cultivars grown in lowland Chile and in the high Andes are referred to as Solanum tuberosum, within which several groups are recognized (Spooner et al. 2014).
The genetic resources available for potato improvement comprise a polyploid series (2n = 2x = 24 to 2n = 6x = 72) with genetic features that facilitate gene transfer across ploidy levels. Farmers’ landrace varieties dominate potato production in the Andean center of origin and diversity; as cultivars they offer tremendous trait diversity in readily useable form. Landrace cultivars, improved varieties and wild potato species in Solanum section Petota comprise GPs (gene pools) 1 and 2, according to the gene pool concept of Harlan and de Wet (1971) and are relatively straightforward to use in breeding. The wild tuber-bearing potatoes (section Petota) include some 200 species (see Spooner et al. 2014 for a review of taxonomic treatments). These typically produce only small tubers, which often contain high levels of toxic glycoalkaloids, and many require short days for tuberization. Domestication of wild potatoes for use as food likely involved selection for increased tuber size and reduced glycoalkaloid content, probably simultaneously in multiple locations in Andean and coastal regions of South America (Ugent et al. 1982, 1987).
Unreduced gametes, self-incompatibility, and inter-specific reproductive barriers played key roles in polyploidization and the maintenance of species boundaries during the evolution and domestication of potato, and knowledge of their genetics is useful in germplasm enhancement. Diploid potatoes are out-crossing due to a system of gametophytic self-incompatibility (Pushkarnath 1942; De Nettancourt 1977), which prevents inbreeding and thereby promotes intraspecific genetic variation. Tetraploid potato, on the other hand, is self-compatible. The breakdown of the gametophytic self-incompatibility system that operates in diploids is a common phenomenon in angiosperm polyploids (Frankel and Galun 1977; Levin 1983), but the molecular mechanism is not known (Comai 2005). Selfing results in severe inbreeding depression in most potatoes. Self-incompatibility at the diploid level, and inbreeding depression in both diploids and tetraploids, make it difficult to eliminate unfavorable alleles or drive favorable alleles to fixation. Several genes governing reproductive isolation, crossability, and ploidy are further discussed in the chapter contributed to this volume by Ortiz and Mihovilovic.
Potato varieties are maintained by clonal (vegetative, asexual) reproduction. Potato tubers are modified stems and comprise the vegetative “seed” used to propagate a variety. In potatoes with an even number of chromosomes (2x, 4x, 6x) it is possible to produce sexual (botanical) seed which provides for the generation of new genotypes. Once variation has been created in the form of sexual seed, any seedling has the potential to become a new variety via clonal propagation. Although most potatoes produce a large number of botanical seeds per fruit (~200) as well as many fruits per plant, clonal propagation results in a low propagation coefficient (five to tenfold per generation). Clonal breeding facilitates intentional and un-intentional exposure of candidate varieties to pathogens. This helps breeders eliminate undesirable genotypes, but also necessitates steps to reduce exposure to detrimental viruses that are transmitted through vegetative (tuber) seed. In common with other root and tuber crops the limited ability to phenotype and potato for desirable morphological or developmental features is challenged by the underground location of the harvested product.
1.2 Genome Constitution and Variation
Maximum heterozygosity has been considered essential for performance of tetraploid potato, with inbreeding leading to reduced vigor and yield, flower bud abortion, lack of flower bud formation, and sterility (De Jong and Rowe 1971; Mendiburu and Peloquin 1977). However, empirical research with hybrid families has suggested that poor performance may be due to the expression of recessive alleles (De Jong and Rowe 1971) or that the presence of certain alleles may be more important for high yield (Bonierbale et al. 1993). Genomic studies have revealed that tri-allelic and tetra-allelic single nucleotide polymorphisms (SNPs) are rare in potato cultivars (Hirsch et al. 2013; 2014), though these might be expected in outstanding clones if yield or vigor were associated with maximum heterozygosity. Nevertheless, when the products of individual genes are amplified by PCR (polymerase chain reaction) it is not uncommon to find three alleles at a locus.
Xu et al. (2011) proposed heterozygosity as the key feature enabling the frequent occurrence of gene presence/absence variants and other potentially deleterious mutations in the genome of the heterozygous diploid potato clone “RH.” Next-generation (short read length) sequencing of 807 genes from 83 potato cultivars revealed a tremendous amount of genetic variation in potato. On average, there is one variant (SNP or indel) every 24 base pair (bp) in exons, and one variant every 15 bp in introns (Uitdewilligen et al. 2013). The average minor allele frequency of a variant is low, though, at 0.14, and 61% of variants have minor allele frequencies less than 0.05 (Uitdewilligen et al. 2013). Given the sequence variation, and keeping in mind that tetraploid potato can contain up to four alleles at each locus, it is easy to understand why so much phenotypic variation results every time two heterozygous potato clones are crossed. Considerable effects due to dominance and epistasis are possible, and experience with such out-crossing polyploids has shown that the genetic variation due to dominance and epistatic effects is large compared to that seen in diploid crops (Gruneberg et al. 2009). This suggests that heterosis largely determines the performance of out-crossing, clonally propagated crops, although its basis in potato has not been fully elucidated.
1.3 The Cost of Increasing Genetic Variation
Most plant breeding training programs place heavy emphasis on the need to increase genetic variation. There is value in this, of course, as wild species contain many useful traits, genes/alleles not present in modern cultivars. But there is also a considerable cost that is not appreciated by those unfamiliar with potato breeding. Wild potato species contain countless alleles that are undesirable for potato production, and when a cultivated potato is crossed with a wild accession, oftentimes the offspring is quite poorly adapted. Many further generations of crossing are needed to eliminate the undesirable alleles, while keeping the desired ones. When embarking on such a venture it is important to realize upfront that the process typically takes decades. It may be possible to use molecular markers to speed the process up, by selecting against the donor genome (and for the gene of interest) after each cycle of crossing, but the process will still be much slower than adding a gene to cultivated potato by Agrobacterium-mediated transformation, or by editing an existing cultivated allele to a desired wild species allele with CRISPR-Cas9 (https://www.yourgenome.org/facts/what-is-crispr-cas9), once the respective target genes are known. It is worth noting that the “adaptation gap” between wild species germplasm and cultivated germplasm is growing over time, as breeders continually work to improve cultivated germplasm by increasing the frequency of desirable alleles and decreasing the frequency of undesirable ones. Despite the challenges, support for germplasm enhancement programs that strategically bridge this gap and enable the continuous influx of valuable genes from crop wild relatives through improved populations and into varieties is critical to meeting the world’s growing need for food in the face of climate change.
1.4 Genetic Enhancement
Potato breeding strategies frequently include research to efficiently access traits from beyond the variety-ready germplasm base. Such trait research or pre-breeding is expected to result in new materials, methods, tools, knowledge, and approaches to support the breeding process rather than in finished products or varieties. Genetic and biochemical research often provides insights and tools that enable gains in traits toward new breeding objectives or improved program results. Pre-breeding is conducted in parallel with mainstream breeding activities, such as by developing support populations that are upgraded for trait levels or improved for agronomic traits so that new types of diversity can be introduced from un-adapted to adapted germplasm without impeding advance toward established breeding goals.
Before embarking on breeding for new traits or using uncharacterized germplasm, it is important to assess and consider positive and negative correlations among traits. Trait correlation influences the success of cross combinations and can determine breeding progress when multiple traits are concerned. Negative trait associations can be critical in achieving breeding progress and influence the choice of parents as well as the selection approach that will be most successful. Embarking on medium- to long-term population development does not result in new varieties after a single recombination and selection cycle, but should result in better parents that will help meet the medium- to long-term objectives of breeding programs.
Support populations are useful for enhancing diversity from un-adapted germplasm to avoid introducing undesirable features into advanced breeding populations. In the case of potato, wild and landrace relatives often carry undesired agro-morphological traits like deep eyes, small tuber size and late maturity, a requirement for short days, or high glycoalkaloid content, in addition to untapped resistance to biotic and abiotic factors and nutritional traits. Those undesired traits may be eliminated from hybrid populations by backcrossing to improved types, or source populations may be enhanced for agronomic traits before desired traits are transferred to improved genepools. The large majority of potato’s genetic resources are diploid, and breeding at this level results in faster genetic gains than breeding at the tetraploid level. Thus, particularly for multigenic traits, pre-breeding in diploid source germplasm before incorporating new traits into tetraploid breeding populations can be very effective.
Introgressing novel traits from distant wild species such as those in the tertiary gene pool (GP3) may require the use of bridge species to circumvent interspecific reproductive barriers, and unreduced gametes to transfer traits across ploidy levels. When interspecific crossing is possible, backcross schemes are modified from those used for inbred crops, such that a different genotype of the recipient germplasm (adapted type) is used in each cycle of crossing with the trait donor or selected hybrid to avoid inbreeding depression. This is illustrated by Gaiero et al. (2017) who introgressed partial resistance to bacterial wilt, caused by Ralstonia solanacearum, from the sexually incompatible GP3 species S. commersonii by bridge crosses with 2x S. tuberosum Phureja Group and successive backcrosses with different S. tuberosum Group Tuberosum genotypes. The resulting advanced backcross progenies are now being used by several breeding programs due to the acute need for resistance to bacterial wilt.
1.5 Case Study 1: Genetic Enhancement and Incorporation of Iron Content from Diploid into Tetraploid Cultivated Potatoes
With support from HarvestPlus (www.harvestplus.org) advocating the breeding of staple crops for micronutrient density, CIP has sought to increase the content of iron and zinc in potato through an inter-ploid breeding strategy. Recurrent selection in a base population of landrace potatoes took advantage of greater response to selection at the diploid level, and resulted in a population of diploid potato reaching 35 ppm iron and 30 ppm zinc from a baseline of 20 and 16 ppm, respectively. Elevated iron and zinc contents achieved in the source germplasm were incorporated into advanced, tetraploid populations via unreduced gametes. The resulting tetraploid potato population had iron and zinc concentrations twofold higher than baseline levels, which approaches the breeding targets for human populations with deficiencies of these minerals and high potato intake (Bonierbale et al. 2007; Section 6.3.4). Further work remains to be done to identify clones adapted to target agro-ecologies that maintain as favorable a package of traits as possible, including new levels of iron or zinc concentrations in resilient, consumer-accepted table potatoes.
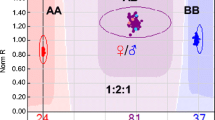
Analyzing biofortified populations under recurrent selection in diploid potato germplasm, Paget et al. (2014) found moderate to high and positive correlation between iron (Fe) and zinc (Zn) contents from cycle I (r2 = 0.45) and from cycle II (r2 = 0.72), indicating that evaluation and selection for one of these traits will result in concomitant increase in the other. Negative genetic correlations were found between dry matter and Fe, Zn, Ca, and vitamin C contents (genetic correlation close to zero for vitamin C in Cycle 1) when analyzed on a dry-weight basis. In contrast, the same genetic correlation estimates were positive (but small) when analyzed on a fresh-weight basis (Table 6.1).
The genetic correlations in this example are strong enough that under multi-trait selection the breeding population is improved simultaneously for iron and zinc concentrations (Fig. 6.1a, b), whereas the average tuber weight of the population decreases, and the number of tubers per plant increases (Fig. 6.1c, d). A reduction in tuber size was encountered during population improvement at the diploid level even though it was considered in selection. This loss was mitigated, however, by returning to the tetraploid level via 4x–2x crosses by which the gains in the mineral contents realized at the 2x level were transferred to advanced populations.


Box plots of ranges of trait variation through three cycles of recurrent selection in diploid potato germplasm. (a) Iron concentration; (b) zinc concentration; (c) average tuber weight (reflecting size); (d) Tuber number per plant
2 Principles of Potato Breeding Methods and Approaches
The principal advantage of breeding clonally propagated crops is that each clonal variety is fixed and simple to maintain. Genetic purity is less of an issue in vegetatively—than in sexually—propagated crops. One substantial disadvantage of vegetative propagation though is that diseases are easily transmitted across clonal generations during propagation; another is that potato planting material is bulky and perishable by nature and the production of healthy material is expensive.
The single most challenging aspect of potato breeding is the identification of superior individuals that combine as many high priority traits, and as few weaknesses, as possible from a given cohort of F1 progeny in a reasonable time frame. Additional important challenges include the improvement of support populations, and the selection or construction of parents. The parents used for crossing are highly heterozygous—exceptions are inbreeding lines generated by self-fertilization or doubled-monoploid production. Each potato seed that results from a heterozygous × heterozygous cross differs, at many loci, from any other seed from the same cross. The resulting heterozygous genotypes are subject to selection after being fixed (stabilized in genetic terms) by clonal propagation.
Polyploidy, heterozygosity, and heterosis make the identification of good parents particularly challenging. At present, the performance of a parent can only be determined after the fact, that is, by looking at its progeny. Advances in genomic selection may make it possible to identify good parents in advance. Heterozygous tetraploid potato genotypes harbor great allelic diversity and interactions that are responsible for their performance as clones. As parental clones are not inbred, a genotype can never be reconstituted after sexual crossing or self-pollination. A practical consequence of this for potato breeding is that many, many, traits segregate in the progeny of any parent or parental combination. Directional breeding results in incremental changes in gene frequencies among progenies.
In the course of potato breeding an “F1 clone hybrid” is generally crossed with another “F1 clone hybrid,” and the progeny is heterogeneous and has an extreme large segregation variance. A good parent generates a large genetic variation around a high family mean for a given trait. Once heterogeneous and heterozygous progenies are generated by crossing two potato clones, selection of clones within a given pool of genetic variation for variety development is conceptually, if not technically, straightforward. All the genetic advantages of clonally propagated crops can be used for variety development, and the genotype that will finally be released is among the progeny immediately after the initial crossing.
Inter-group crosses are important in population improvement of clonally propagated crops. Gruneberg et al. (2009) have suggested that this aspect of clone breeding is often neglected and may be the reason for low breeding progress in many clonally propagated crops when compared to the improvement of sexually reproducing ones. Complementary germplasm groups have not been identified in potato but strategies to maximize heterozygosity have been proposed to increase yield in tetraploid potatoes (e.g., Chase 1963). However, it is unlikely that the direct relationship of maximum heterozygosity and yield will extend to crosses involving un-adapted germplasm (Bonierbale et al. 1993). Thresholds for heterosis were suggested upon finding of increased yield and vigor in two-way hybrids (cultivated × wild species) with respect to crosses within the cultivated genepool, but no additional increments in hybrids involving cultivated potato and two wild species (3-way hybrids) (Sanford and Hanneman 1982). Thus, it is unlikely that heterotic groups could be established in potato germplasm on the basis of genetic distance measures or maximum heterozygosity alone.
2.1 Population Improvement
Medium- to long-term genetic gain can be achieved over sequential cycles of crossing and selection. Recurrent selection is defined as reselection generation after generation, with inter mating of selected plants to produce the recombinant population for the next cycle of selection. The goal of recurrent selection is to improve the mean performance of a population of plants; a secondary goal, but nevertheless also important, is to maintain as much genetic variability as possible. Open recurrent selection is a method for improving the mean performance of a population while maintaining and increasing genetic variability by periodically introducing new sources of traits under selection. In genetic enhancement a few cycles of recombination facilitate the breakdown of linkage blocks so that desired traits from unadapted germplasm can be carried forward and undesirable ones left behind, minimizing the effects of linkage drag thus shortening the time required to introgress or incorporate new diversity into advanced populations. Depending on the stage of a population under improvement, selections may be considered for use as varieties, or as parents that will contribute to further gains toward complex breeding objectives. Figure 6.2 depicts simultaneous and integrated practices of recurrent selection, genetic enhancement, and variety selection in a potato breeding program.


Schematic representation of components of a potato breeding program. (Courtesy of Elisa Mihovilovich)
In polyploid potato, more than one allele per locus can be transferred to the next generation in gametes, and thus, in contrast to diploids, the genetic variation due to dominance influences the response to selection in population improvement as long as the population is not in equilibrium. In tetraploid potato, a population is usually not in equilibrium after recombining parental material in controlled crossings, and 1/3 of the dominance variance is exploitable for selection progress when selection takes place on the female and male side [for further details see Wricke and Weber (1986) and Gallais (2004). The exploitation of the dominance variance in population improvement, in combination with faster genetic gains in diploids than in tetraploids, and the inheritance of 2n gametes as described by Ortiz (1998) provide great diversity and flexibility for potato improvement.
2.2 Crossing Parents
The choice of parents is an important step in any breeding program, especially for medium- and long-term breeding progress. The identification and number of crosses to perform at a given breeding stage is a factor of the knowledge available on potential parents (i.e., trait constitution and breeding value), the breeding objective, and facilities and resources available for crossing and selection. As mentioned above, the polyploid and highly heterozygous nature of potato mean that dominance and epistatic effects contribute considerably to clone performance, and for this reason, little is known about the value of a parent or specific cross combination until it has been tested.
Parental value can be assessed strategically through the conduct of progeny tests in appropriate breeding designs, the evaluation of pedigrees, or empirically through observation of selection ratios. Even when hundreds of crosses are made, it is often observed that the best clones trace back to very few crosses. Thus, it is desirable to predict which genotypes will be the best parents, since if this is known, efforts can be concentrated on the generation and the evaluation of the most promising combinations. Regardless of how much information is available to support the choice of parents, a practical approach is to sow around 200 seeds of every cross to be evaluated, and then sow more seed or emphasize the parents of those combinations that performed well in future years.
Parental value is a function of the genetic constitution of a trait donor. Complex traits are often comprised of several components, and defining those that contribute reliably to influence trait values is a prerequisite to successful identification of parents. Accurate evaluation of progeny is the most effective means to identify superior progenitors of inherited characteristics. Sprague and Tatum (1941) introduced the concepts of general combining ability (GCA) and specific combining ability (SCA) to distinguish between the average performance of parents in crosses (GCA) and the deviation of individual crosses from the average of crosses (SCA).
Studies of combining ability make it possible to identify parents (those with good GCA) that will perform well in most crosses, and also facilitate the development of superior hybrids through the use of parents with high SCA. Genetic analyses to measure combining ability further allow breeders to estimate genetic effects and parameters such as trait variance, covariance, correlations among traits, heritability, and the relative importance of additive and dominance variation. GCA represents mainly additive and additive × additive types of genetic variance. Thus, when a potato clone is selected as having good GCA, it means that the algebraic sum of the additive and additive × additive epistasis effects it passed on to its offspring produced a favorable result in excess of the average of all the offspring tested. GCA has a conceptual implication that each line being evaluated is tested against a large random sample of some specified population (Plaisted et al. 1962). In actual tests for GCA, however, the testers (usually males) often represent only a limited sample of a population.
2.3 Mating Designs
The term “mating design ” refers to the mating of parents in a systematic plan of crosses to determine genetic parameters and/or parental value. These procedures are particularly useful for the identification of plants that will be the most effective parents in pre-breeding or recurrent selection programs. However, systematic mattings can be difficult to achieve because of sterility and incompatibility encountered during crossing. Mating designs have been classified according to the number of factors to be analyzed, the parents, and modalities of combination. Some mating designs are used more extensively than others, but each has its advantages and disadvantages depending on the reference population under consideration and the information desired. Following are descriptions of some of the most important designs for identifying superior progenitors to help assure genetic gains in a potato breeding program. Figure 6.3a–c, illustrates some of these mating designs.


Schematic representation of different mating designs in which parent lines are indicated with numbers, and the cross combinations between them are represented with checkmarks in shaded cells. (a) Partial dialel, (b) Line × tester, and (c) Design II
Diallel Analysis is the best way to determine the combining ability of parents. It consists of the analysis of a set of crosses produced involving “n” lines in all possible combinations, a so-called diallel cross. The diallel mating design has been used extensively in potato germplasm enhancement and can be very useful if properly analyzed and interpreted. The analysis of diallels provides information on GCA and SCA of parents and their crosses and makes it possible to determine if reciprocal crosses give equivalent results.
The most commonly used methods of diallel designs are those proposed by:
-
Griffing (1956), by which general and specific combining abilities are estimated;
-
Gardner and Heberhart (1966), by which the variety and heterosis are evaluated; and
-
Hayman (1954), in which information regarding additive and dominance effects for a characteristic and the genetic values of the parents is used.
One of the most used is Griffing’s Method 2, which estimates GCA and SCA) relating mainly to additive and non-additive gene effects (dominance and epistasis).
2.3.1 Partial Diallel
Analyses involving “n” parents in all possible combinations become unmanageable as the number of lines (n) increases. On the other hand, if only a small number of parents are tested, the estimates of combining ability tend to have a large sampling error. These difficulties led to the development of sampling crosses produced by large numbers of parents, without affecting the efficiency of the diallel technique. In a normal diallel, each line is involved in (n − 1) crosses. Kempthorne and Curnow (1961) presented the concept of the partial diallel design in which only a random sample of crosses, say of size’s, is analyzed where “s” is less than n −1.
2.3.2 Line × Tester Design
-
The Line × Tester Design also provides information about the GCA and SCA of parents, and is also helpful in estimating various types of gene effects. The crossing plan of this design is as follows:
-
“l” lines are crossed to “t” testers so that l × t full-sib progenies are produced;
-
These progenies, with or without parents (i.e., testers and lines), are then tested in a replicated trial using a suitable experimental design, say randomized block design.
-
If there are three testers and seven lines, there are 7 × 3 = 21 crosses. For evaluation, the 21 crosses along with 10 parents, for a total of 31 entries, might be tested in a randomized complete block design with four replications. For this case, uniform planting material must be produced to enable inclusion of parental clones and progeny in the same trial.
2.3.3 Design II
In mating Design II (or Factorial Design), described by Comstock and Robinson (1948), the genetic information is similar to that obtained with Diallel Analysis.
Different sets of parents are used as males and females. If a set of eight parents is included in the design II, 16 crosses will be obtained. This design is advantageous when not all clones to be tested are male or female fertile.
2.4 Breeding Values
The estimated breeding value (EBV) of an individual can be calculated on the basis of pedigrees and performance in the course of a breeding program, and does not rely on the conduct of mating designs. Using appropriate statistical analysis, breeding value predicts how useful each individual would be as a parent; it expresses the ability of a parent to pass on superior trait levels to its offspring and is used for ranking breeding performance of an individual as a parent relative to the population average. The use of information on the individual and all relatives greatly increases the accuracy of selection in a breeding program (Lynch and Walsh 1998).
The calculation of breeding value goes beyond the typical estimation of genotypic or parental values with models based on fixed effects, by enabling the estimation of random effects of a mixed model. Mixed linear models are able to model different covariance structures and thus provide an improved representation of the underlying random and error components of variance (Oakey et al. 2007). The application of mixed models to estimate breeding values uses pedigree information to model and exploit genetic correlation among relatives and applies flexible variance–covariance structures for genotype-by-environment interaction to accurately predict performance (Piepho et al. 2008). The improved accuracy afforded by the use of all data in a breeding program with mixed models allows the analysis of repeated measures, unbalanced design experiments, spatial data, and multi-environment trials.
Breeding value has been successfully applied in several crops (sugarcane, eucalyptus, soybean, maize, and even potato). Slater et al. (2014) illustrate selection in potato based on breeding values. They conclude that using best linear unbiased prediction (BLUP) and pedigree to estimate breeding values can result in increased genetic gains for low heritability traits in auto tetraploid potato. Theory and applicability of breeding values in quantitative trait improvement are illustrated in Bernardo (2002) Relevant statistical packages that fit linear mixed models to large data sets using the Residual Maximum Likelihood (REML), approach, such as the Asreml-R reference manual (Butler et al. 2009) provide powerful software for the use of breeding values in plant breeding.
In animal breeding, EBVs are the basis for marketing breeding parents and they provide breeders with critical information for selection decisions. In applying EBVs, it is important to achieve a balance between the different groups of traits and to place emphasis on those traits that are important to the objective population, markets, and environment. It is not feasible, nor always desirable to seek high EBVs for all traits in a single progenitor. In fact, a comprehensive range of EBVs has the advantage that it is possible to avoid extremes in particular traits and select for animals/genotypes with balanced overall performance. The method is particularly useful in non-inbred populations and potato breeders should pay careful attention to this analytical approach.
2.5 Early Versus Late Generation Selection
In early generations it is possible to select for highly heritable traits for which accurate assessment of a genotype can be carried out on one or a few plants, but for more genetically complex traits, and for traits where interplant competition is an issue, it is necessary to evaluate clones in multi-plant plots planted as blocks under homogenous field conditions. The aim is an unbiased comparison of genotypes within blocks. The number of plants per plot and the number of replications or blocks depends on the breeding stage. The low propagation coefficient of potato (about ten depending on the propagation method used) limits the amount of planting material available at each stage of selection and is one reason that potato breeding is relatively slow (at least 8–10 years and usually more, from crossing until variety release). Large numbers of genotypes can be assessed for simple traits in early stages when small amounts of seeds are available; while more complex traits are assessed in later stages when larger quantities of seeds are needed for replicated trials, though less genetic diversity is represented.
Multistage selection can be managed in subsequent steps from early to later stages, or with indices that use a weight assigned to each trait. In practice, some characters are selected sequentially especially where there is clearly a lowest acceptable value (tuber size, shape, and color as well as pest and disease resistance), while others are selected simultaneously by aggregating characters into an index (often an intuitively formed index such as score values of overall performance.
2.6 Case Study 2: The Use of a Selection Index in Potato Breeding
An early-generation (seedling stage) selection method was applied at CIP for the identification of families and individuals that tuberize well under long days and high temperatures. Five groups of families generated by intercrossing CIP’s best advanced parents from two populations and long day-adapted varieties underwent a greenhouse test during summer, applying two photoperiods: natural short days of 12 h and simulated long days using lamps to extend the photoperiod to 16 h. Four plant morphology prototypes were identified according to patterns of above- and below-ground growth including branching, stolon, and tuber formation (Fig. 6.4). Individual plants of each family were evaluated at harvest taking into account parameters including breeder’s preference, a tuberization score based on the four patterns, tuber uniformity and physiological disorders including sprouting tubers, knobbiness, chain tubers, and cracking.


Plant morphology and growth patterns observed among families in early generation screening for adaptation to warm, long day environments. (a) Typical morphology under 12:12 h light:dark regime; (b) Typical morphology under 16:8 h light:dark regime; (c) Above-ground morphology of representative plants with growth patterns I, II, III, and IV; Below-ground morphology of representative plants with growth patterns I, II, III, and IV
A selection index (SI) was built assigning a weight to each of these parameters, three for breeders’ preference (BP), two for tuberization score (TS) and tuber uniformity (TU), and one for physiological disorders: sprouting (Sp), knobbiness (Kn), chained tubers (Ch), and cracks (Cr)
Families with a high frequency of progenies exceeding an estimated selection index of 54 were selected. Seven of these selected families and three unselected were used in validation studies under field conditions.
2.7 Stability and Adaptation
Breeding programs typically breed for several locations rather than just one. Hence field evaluations that underpin selection must simulate a range of environments. For this reason, within limits of the propagation coefficient, clones are tested in plots within homogenous blocks at several locations over several years. The objective of multilocation trials is to assess promising genotypes with respect to narrow and broad adaptation. In practical potato breeding, “broad adaptation,” the ability to perform well in a range of environments, appears essential for a clone to have any meaningful commercial success.
The wide range of quality preferences and the numerous pests and diseases addressed by potato breeding programs dictates the exposures required for selection. Decentralized selection is generally required by a program with multiple target environments and is best designed to address sets of traits that can logistically be assessed together in target environments or sites that represent them well. Along the course of selection, a combination of on-station and on-farm trials are usually performed.
National programs requesting potato breeding materials from CIP are provided with germplasm from different selection stages depending on their capacity to evaluate clones and the relative suitability of advanced germplasm to their production targets. Advanced clones and parents are available as in vitro plantlets. The genotypes provided meet multiple selection criteria for the given target environment, as assessed in similar testing sites. Upon receipt they are propagated and subject to multilocational trials before use as parents in national programs or testing as varieties.
2.8 Case Study 3: Breeding and Variety Development in Bangladesh
Breeding for stable yields and multiple disease resistance at CIP takes advantage of germplasm collections, broad-based advanced populations, and environmental diversity in Peru. One such population is the advanced lowland tropics virus resistant population, which is adapted to dry arid regions where virus pressure is high. With support from GIZ/BMZ and follow-up support from USAID, CIP provided 35 potato breeding lines of this population as in vitro plants to the Tuber Crop Research Centre of the Bangladesh Agricultural Research Institute in 2009. After local evaluation over a period of 7 years, a salt tolerant variety (BARI Alu-72) and a heat tolerant variety (BARI Alu-73) adapted to Bangladesh were released.
When distant environments can only be partially simulated in CIP’s breeding program, samples of true seed families (TSF) from good parents can be distributed. TSF are available for national programs with the capacity to carry out early generation selection as well as variety identification trials. One advantage in working with TSF is the considerable reduction in time-to-release compared to that required when receiving advanced clones. Conducting selection with true seed in target environments can save 6 years or more, i.e., time that would otherwise be required for CIP breeders to conduct selection from the same TSF at their trial sites and recommend elite clones as in vitro plants.
When national programs test CIP germplasm as TSF, valuable information is returned to CIP regarding family performance (i.e., frequency of selected clones in each family), and this serves to refine the concept of adaptation and best bet materials for that location. National programs that invest in identifying parents based on local performance, and then utilize these parents for crossing and recurrent selection, benefit from knowing which breeding materials will serve those best and are likely to experience significantly faster genetic gains over the medium to long term.
2.9 Case Study 4: Collaborative Breeding in Vietnam
Advanced germplasm from CIP’s lowland tropic virus resistant (LTVR) and late blight heat tolerant (LBHT) breeding populations developed in Peru were dispatched to Vietnam in the form of in vitro plants from 2005 to 2010. The materials were used in two ways: (i) for direct variety testing, and (ii) in cross-breeding to improve adaptation to local conditions.
Results of the collaboration demonstrated both the direct utility of clones from Peru in Vietnam—through the identification of two outstanding clones recommended for variety testing, and the advantage of local breeding, yielding, after three rapid cycles of recurrent selection, a new generation of elite clones with higher yields than the elite clones provided from Peru. The yield increase from local breeding and selection was realized in a considerably less time (9 years) than would have been required for the development, introduction, and testing of a new cycle of elite clones from CIP (at least 12 years) (Fig. 6.5a, b).


Collaborative approach for potato breeding in Vietnam. (a) Two avenues for decentralized evaluation and improvement; (b) Frequency distributions for yield of selected potato clones in three cycles of recurrent selection conducted in Vietnam
There are several advantages of testing early generation potato progenies at more than one location. Information from contrasting environments can be combined if the breeder tests a clone at two or more locations; those few clones that are broadly adapted can be identified early in the selection process. In addition, if one location experiences low disease pressure, daughter tubers from that site can be used to provide clean planting material for the next generation of selection. The healthy seed plot provides for an assessment of performance in the absence of major production constraints, while various pressures can be applied at additional sites. Alternatively, a protected environment (e.g., quarantine screen house) can be used to maintain a healthy copy of each selected genotype, but unlike the maintenance of a healthy seed plot, this typically does not allow for significant production of seed to support evaluation along the breeding cycle.
Rapid and real-time exchange of information on selection percentage between breeders and selectors helps inform breeders on the future crossing plans most likely to meet the requestor’s needs. This simple data can confirm the breeders’ concept of GCA and suitability of test environments with respect to the targeted ones. Breeders can use the information returned from selectors regarding percentage of selection in controlled cross families to exploit heterosis through cross combinations. Selection of parents and cross prediction can probably be improved by systematic collection of data on the percent of clones selected in families by the assignment of parents with good GCA or mutual SCA to complementary gene pools. This would benefit both CIP’s and the national programs’ breeding efforts, if it resulted in increased frequency of high-yielding clones with needed traits, or potentially, in yield jumps.
2.10 Importance and Relation of GCA and Heterosis in Potato
Heterosis was examined in a new tetraploid potato hybrid population obtained by crossing members of two advanced breeding populations developed at CIP for adaptation to the highlands (B3) and subtropical lowlands (LTVR). Significant positive heterotic values ranged from 18 to 60% for mid-parent heterosis and from 16 to 162% for best parent heterosis or heterobeltiosis. Positive heterotic values were found for tuber yield and tuber number in each environment. Significant GxE interaction on heterosis showed a differential effect of the environment on the magnitude of heterosis expression.
The expression of heterobeltiosis within B3 and LTVR confirms the effectiveness of the population breeding method on maintaining a broad genetic base (Mendoza and Sawyer 1985). Mid-parent heterosis can be the result of the combination of good levels of GCA of parents and some level of SCA of the cross. A reciprocal recurrent selection scheme (RRS) has been initiated to benefit from intercrossing between best parents of two complementary populations, B3 and LTVR (RRS), also known as recurrent reciprocal half sib selection, is a form of recurrent selection used to improve both GCA and SCA of a population for a character or characters.
3 Potato Breeding Procedures Overview
3.1 Hybridization
Heterozygous parents are recombined in controlled biparental combinations or by use of bulk pollen to create new variation in the form of sexual seed. The crossing block is established after selecting the parents to be used as females and as males, and the “planting on a brick method” or modifications of it can be practiced to encourage flowering and fruit set over a period of about 10 weeks. In the “brick method,” potato tubers are placed on bricks partially buried in the soil. The tubers sprout and stems are allowed to grow. After some time the soil around the base of the full-size plant is washed away with a spray of water, which exposes emerging stolons and small tubers, but does not disrupt the root system that has penetrated in and around the brick. Thereafter, stolons and tuber that emerge from the base of the plant are removed. Potato has a complete flower with five anthers and a stigma that becomes receptive as pollen is shed from open flowers. Emasculation is practiced to prevent selfing when genetic studies will be performed, when parental value will be assessed in progeny tests and for best control of pedigrees (record-keeping).
Crossing at CIP is enhanced by flowering induction. Under long day conditions most potato clones flower to some extent. Typically, S. tuberosum Phureja Group flowers under both long and short-day photoperiods. Clones of Group tuberosum, however, usually will not flower under short days. In addition, there are many clones, especially those of early maturity, that rarely flower under any conditions or that flower sparsely and over a very short time period. For these reasons it is often desirable to induce flowering. This can be done by either of two methods: “grafting” or “planting on a brick.” In the grafting method shoots of potato are grafted onto the stems of tomato plants. In both methods the idea is to prevent potato from developing beyond the vegetative flowering phase to the tuberization-senescense phase. In addition to these mechanical methods, other traditional flowering-enhancing practices may be followed such as long-day lighting, sprays of 40 ppm gibberellic acid at 4-day intervals, and heavy nitrogen fertilization. Temperature and humidity are also important. Ideal temperatures are 20 °C day and 16 °C night. Humidity should be 80% or higher. Procedures and crossing techniques for potato breeding as practiced at CIP are documented at https://research.cip.cgiar.org/potatoknowledge/proceduretechniques.php. A modification of “planting on a brick” using peat pots is shown. The open source software CIPCROSS (see Section 6.3.3) can be used to document all aspects of the crossing block, including pedigrees, storage, and inventory using bar coding for accuracy and efficiency.
3.2 Selection Schemes
Selection of potato varieties from botanical seed is conducted in several steps. Figure 6.6 shows how the crossing of two parents might be followed by five sequential steps in time (one selection step conducted with seedling plants and four subsequent selection steps conducted with cloned plants from tubers of the seedling genotypes). This scheme is an oversimplification since many crosses or families are developed simultaneously and many seeds of each family are subject to evaluation and selection at the same time, while here only three seedlings in each of 11 families are shown.


Multilocation Clonal selection Scheme. (Modified from Gruneberg et al. 2009)
In principle, if the true seed plant could be cloned in large quantities and the population could be assessed with adequate accuracy, it is theoretically possible to select the “best” genotype in the first year, since maximum variation is present at the start of each selection cycle and no genetic changes are realized during it. In practice though, the individual genotypes selected in early stages of each clonal selection cycle are subject to propagation via tubers to permit adequate plot size and replication for later-stage assessments.
The overall selection scheme should consider the vulnerability of clonal planting material to disease. At least some tubers of every clone should be planted in a disease-free location, or time and resources should be allocated for the eventual elimination of pathogens from successful clones before they can be distributed among geographic regions.
Many breeding programs use only one location at the early selection stage, preferring to evaluate in favorable conditions such as on station, while also producing healthy seed for use in later selection stages. This location should be as free from virus pressure as possible to help keep seed clean over the years. For example, the New York breeding program makes all evaluations for the first 2 clonal years in a single location, going to a second location (for an unreplicated evaluation of yield) in the third clonal generation, and multiple locations (for replicated yield trials) starting in the fourth clonal year. Typical numbers of genotypes per cycle in this program are: 20,000 seedlings in year 1, 18,000 four hill plots in year 2, 1500 twenty-hill plots in year 3, and 250 hundred-hill plots in year 4.
Nevertheless, the first clonal generation can be evaluated at more than one location if sufficient clonal seed can be produced. For simultaneous evaluation at up to three locations in the first clonal generation, true seed families (TSF) are first converted to tuber families (TF) so that at least four copies of each genotype from each family are produced. Methods for evaluating TF include (1) planting three to five tubers of each genotype in a single row at a single location or (2) equivalent samples of each tuber family are evaluated in two or more locations, using one tuber per genotype and location. Multilocational testing of first clonal generation tuber families can be done with, or without the identification and labeling of individual genotypes. Detailed procedures for the production of tuber families followed at CIP are illustrated at: https://research.cip.cgiar.org/potatoknowledge/tuberfamilies.php
3.3 Data Management and Analytical Tools
Standardized information on the performance of progenies and selected clones across environments is necessary in order for breeders to efficiently make decisions about selection and variety release. The methods of data generation and processing that are utilized in plant breeding have radically changed in recent years. With the advancement of new high throughput technologies, data have grown in terms of quantity as well as complexity. However, the significance of the information that is hidden in newly generated experimental data can often be deciphered only by linking it to other data, collected previously and/or by others. Collaboration that makes it possible to connect disparate data sources and analyze them in meaningful ways with other researchers requires robust but practical data management solutions. CIP has developed an on line Global Trial Data Management System: https://research.cip.cgiar.org/gtdms/. Three key components are CIPCROSS, HIDAP, and Field Book Registry.
CIPCROSS is a botanical seed inventory tracking system for clonal crop breeding that enables tracking of breeding materials from crossing blocks and botanical seed inventories through to seed distribution. CIPCROSS is open source software, available online, and comprises two main tools: (a) The Crossing Management System (CIPCROSS Tool v1.1 for Pocket PC) used the main development platforms Visual Basic 2008 for Windows Mobile OS and Microsoft SQL Server Compact 3.5 for Windows Mobile software. This tool facilitates the barcode labeling of parents and crosses, storing the information in a database and generating automated reports and (b) The Botanical Seed Inventory System (CIPSIS Tool v1.1) was designed as an easy-to-use web application. It uses PHP, Yii Booster, Bootstrap, CSS, Java Script, HTML and MySQL database programming languages. This tool facilitates the recording and accessing of information on the location of botanical seed in the storage facility, documentation of germination tests, updates on seed stocks, information about crosses and breeder’s name. The system helps breeders develop, integrate, and organize their information in a database, avoid typing mistakes and saving work time (24 person–hours are saved per 14,250 labels). The Roots & Tubers Base centralized data repository facilitates access to the information with queries and filters for advanced searches (Fig. 6.7—Users can download these tools through the web page of “The Global Trial data Management System from CIP” https://research.cip.cgiar.org/gtdms/)


CIPCROSS workflow: (a) The Crossing Management System (CIPCROSS Tool v1.1 for Pocket PC); (b) The Botanical Seed Inventory System (CIPSIS Tool v1.1) and c) Global Roots & Tubers Base as a centralize data repository; (c) Global Roots & Tubers Base as a centralize data repository
Data collection in potato breeding requires capacity to use and share standard protocols for the selection of clones from seedlings through to observational trials and on to preliminary and advanced yield trials, each accompanied by field books and structures for data collection, analysis, and reporting.
The highly integrated data analysis platform called HIDAP was developed by CIP’s breeding program and Research Informatics Unit to facilitate and unify data collection, quality control, and data analysis for clonally propagated crops. HIDAP provides a single platform for use by potato and sweetpotato breeders. It supports compliance with Open Access, open standards such as the potato and sweetpotato crop ontologies and linkages with relevant corporate and community databases such as CIP’s BioMart (https://research.cip.cgiar.org/gtdms/biomart) and SweetPotatoBase (www.sweetpotatobase.org). HIDAP builds on the statistical platform R. This includes the R shiny tools, the knitr package, the Agricolae package also developed by CIP (https://cran.r-project.org/web/packages/agricolae/agricolae.pdf), and more than 100 other R packages. The R shiny package enables implementation of interactive web pages that are usable online and offline. The knitr package enables the creation of reproducible reports. Numerous statistical analyses can be performed using R and R functions developed at CIP. The software is available for download at https://research.cip.cgiar.org/gtdms/hidap/.
HIDAP is connected to the institutional pedigree and corporate database at CIP facilitating the tracking of clones and families generated through breeding. This connectivity enables verification and maintenance of the identity of clones across the different selection stages and tracing of pedigrees in selection and breeding. The HIDAP network enables researchers to share field books with colleagues, regional breeding programs, and/or partners. To use this network, users must register and create a login account. Once logged in, the user can share, download, and receive field books for different selection stages in a user-friendly interface. A download count helps to keep track of users and uses of this tool.
The Field Book Registry (https://research.cip.cgiar.org/cipfieldbookregistry/) facilitates updating field books in real time and viewing their status in the database. The data generated in CIP’s potato breeding program is stored in the “Global Roots & Tubers Base” utilizing the free BioMart software https://research.cip.cgiar.org/gtdms/biomart/. This database has been structured for storage of phenotypic, genotypic, pedigree, geographical, and environmental data. Through the metadata and the search function using filters, the user can retrieve data from the experiments conducted by CIP or partner programs using CIP materials. The availability of the data is managed in conjunction with Dataverse following CGIAR open access guidelines (Fig. 6.8).


Flowchart of CIP’s breeding information system: Standardized evaluation protocols and ontologies implement templates to collect and analyze data with HIDAP software; Subsequent data storage through the field book register allows incorporation of breeding information into the Roots & Tubers Base in which phenotypic and genotypic data are integrated and made available for further use
The use of the database and software tools enables analysis of phenotypic and genotypic data. A key goal is the identification of effective models that predict phenotypic traits and outcomes, elucidating important biomarkers and generating important insights into the genetic underpinnings of heritability.
4 Setting Objectives for Potato Breeding
Successful breeding relies fundamentally on having a clear set of prioritized breeding objectives. Key enabling information and tools for setting breeding objectives include: knowledge of the opinions and needs of stakeholder groups, knowledge of the production constraints and cropping systems of target populations and environments, and standardized means to measure and describe traits. Knowledge of the strengths and weaknesses of predominant varieties provides an important baseline for setting breeding objectives.
Having a good understanding of desired end-user traits requires ongoing, interdisciplinary communication and collaboration. This helps increase the chances that a new variety meets actual needs and is thus adopted, as well as minimizing the chance that new varieties (and the traits that define them, their management and marketing or use) might marginalize or disadvantage social or gender groups such as poor farmers or women, who often have access to different resources than men do. There is also a need to consider social and economic dimensions and client demography such as market access, youth, and urban/rural factors. A CGIAR system-wide initiative is currently underway to assess and encourage breeding that responds to social diversity including often-neglected or marginalized groups with emphasis on gender (http://www.rtb.cgiar.org/gender-breeding-initiative/?lang=en).
Potato breeding is characterized by having a large number of objectives, which include traits specific to the intended market and use of the crop (end-user traits), as well as traits related to productivity and protection against local diseases and insect pests (grower traits).
4.1 Targeting of New Potato Varieties
Building on CIP’s experiences of a global breeding program that developed two agro-ecologically adapted populations (one for highland tropics and one for subtropical lowlands), increased attention is now given to prioritizing packages of traits for which relative values are determined with stakeholders in a more decentralized manner. This involves the cross-disciplinary estimation of trait values at global, regional, and local scales, the development of corresponding variety or product profiles with stakeholder involvement, and the setting of breeding priorities to meet those via selection decisions.
4.2 Case Study 5: RTB Priority Setting
An example of assessing trait values can be seen in the priority setting exercise conducted by the Root , Tubers and Bananas (RTB) Program of the CGIAR (http://www.rtb.cgiar.org/resources/impact-assessments/) Kleinwechter et al. (2014). Through a global survey carried out from 2012 to 2013, the RTB program sought to rank production constraints of each RTB crop (i.e., cassava, yam, banana, sweetpotato, and potato) and research options to alleviate them. This exercise was led by agricultural economists specialized in impact assessment. It provided an overview of problems affecting the potato sector and potential solutions including, but not limited to, breeding. The potato survey (Hareau et al. 2014) provided respondents with a list of 91 research options, organized around the areas of crop improvement, crop and resource management, seed management, genetic resources, value chains, postharvest utilization, and marketing, as well as socioeconomic research and extension.
Five of the top ten ranked research options for potato were directly related to breeding. Breeding for late blight resistance, drought tolerance, earliness, and high yield ranked second, third, fourth, and eighth of the 91 options provided, while the generic “germplasm enhancement” ranked seventh. An ex-ante assessment of the selected potato technologies revealed significant differences in terms of net present value (NPV) and internal rate of return (IRR) on investments across the different research options. Late blight- and virus-resistant varieties had the largest expected net benefits ($US 4.7 billion and $US 3.9 billion), and high rates of return of 87% and 104%, respectively, in the high-adoption scenario.
4.3 Case Study 6: Adjusting and Ranking Priority Traits in New York (NY) State
Staff of the breeding program in New York State U. S. meet many times each year with stakeholders, in both formal and informal settings, to continually discuss what the program’s priorities should be. The frequent interaction helps stakeholders feel comfortable expressing their views, and allows the program to detect changes in industry priorities more quickly, as breeding goals always change with time.
Two examples can illustrate how stakeholder feedback has changed the NY program over the past 5 years. (1) NY growers recently made repeated requests to develop earlier maturing potatoes, to solve a problem the NY breeding program had unwittingly helped create. NY breeders have always put a high priority on yield, and selection for yield tends to select for later maturity. Unfortunately, a suite of late maturing varieties also shortened the timeframe in which NY growers could harvest their crop. The NY program now pays much more attention to maturity, and is prepared to select potatoes with lower yield if they mature early. (2) Potato chip factories recently began to ask for smaller chipping potatoes, as more and more of their product is now sold in small bags. In response, the program now selects smaller potatoes than it (or regional chipping factories) would have been willing to process in the not-so-distant past.
One approach that the NY breeding program has found useful when prioritizing traits is to compare what stakeholders ask for (“what stakeholders say”) with the attributes of widely grown varieties (“what stakeholders do”). When there is an apparent disparity between words and action, there is an opportunity for deeper understanding. Fifteen years ago growers in NY kept asking for new varieties with resistance to common scab. What made the request unusual is that growers already had several resistant varieties to choose from, and that the variety they grew most each year was highly susceptible. What the NY program eventually realized is that the popular scab susceptible variety had two quality attributes—outstanding fry color out of cold storage and high specific gravity—that were far more important to the chip factories than the scab resistance growers kept asking for. As a result, NY re-ranked selection criteria, placing fry color and specific gravity above resistance to common scab.
4.4 Product Profiles
A product profile establishes a set of targeted attributes that a new plant variety or animal breed is expected to meet for successful release in a given market segment. Attributes must be understood as traits reaching a specified level; this level being defined either in absolute or relative terms (Ragot et al. 2018). Thus, a product profile may list yield (25 tons/ha or more; or 15% over variety V1 across a range of soil fertility conditions), tolerance to potato leaf roll virus (same as or better than variety V2), or dry matter content (no less than 18%, or no less than variety V3). The development of product profiles may best be done in collaboration with, for example, pathologists, agronomists, or nutritionists, as well as user communities who can contribute specific knowledge, tools, and approaches for setting trait levels, which in turn become the breeding targets within a profile.
4.5 Tools and Metrics
CIP’s potato breeding program targets low input conditions by relying heavily on endemic disease pressure, poor soils and a series of intentional exposures to stress for screening and selection. The product profiles incorporate quantitative breeding objectives for productivity, protection, and utilization traits with emphasis on resilience, the setting of quantitative breeding objectives relies on knowledge of baseline variety characteristics and available genetic resources, the expected effects of changes in trait levels, and means to measure gains toward them.
Metrics for disease resistance: One special tool used by CIP is an interval scale for expressing potato resistance to late blight that indicates resistance levels required for satisfactory control of the disease in agro-ecologies with varying degrees of pathogen pressure. The use of this interval scale facilitates setting quantitative resistance breeding objectives in a robust manner within a breeding program’s trait improvement framework. The late blight susceptibility scale of Yuen and Forbes (2009) enables assessment of resistance levels with a reduced coefficient of variation among trials as compared to other semi-quantitative metrics like AUDPC or rAUDPC. The scale uses reference cultivars in regression analysis and helps breeders to measure and describe resistance of genotypes independent of environment or inoculum level, which can vary from site to site and year to year. This approach gives breeders a simple numerical metric for quantitative traits that is useful for setting baselines, breeding goals, and calculating genetic gains.
Application of the susceptibility scale to setting breeding targets is based on the understanding that a variety with level 5 (more susceptible) would be sufficiently tolerant to provide the same level of protection in low-pressure agro-ecologies as a variety with level 2 (less susceptible) would provide in a agro-ecology with high disease pressure. The use of this scale is illustrated in Field assessment of resistance in potato to Phytophthora infestans at https://research.cip.cgiar.org/potatoknowledge/lateblight.ph.
Metrics for nutritional traits: To help reduce the health burden of iron and zinc deficiencies, quantitative targets for levels of these two elements in potato (a biofortification breeding goal) were set through collaboration between potato breeders at CIP and nutritionists of Harvest Plus. Consideration was given to (1) nutritional status of the target population, (2) dietary increments of iron and zinc known to be effective from other approaches (i.e., food fortification or nutrient supplementation), and (3) features affecting feasibility of nutrition impact such as consumption (potato intake), heritability, bioavailability, and retention of these minerals in potato prepared for use as food.
Nutritionists consider that dietary increments of 0.4 and 0.2 mg/day of iron and zinc, respectively, can have a positive biological effect on the health status of populations at risk of micronutrient malnutrition associated with deficiencies in these minerals. The provision of these increments through biofortified varieties is feasible when mineral concentration of a variety, consumption levels, bioavailability, and retention in the diet are sufficient. This food-based approach to reducing micronutrient malnutrition relies on variety change but not on modification of consumption patterns, i.e., high versus low potato intake. Logically, a population that consumes twice as much potato as another one would ingest twice as much iron from that source. And the percent of the Estimated Average Requirement (EAR) of the nutrients it provides would be double.
Quantification of these features for a given varietal and dietary context assumes conservative estimates of 10% and 25% bioavailability of iron and zinc, respectively, from potato, and that minerals are not lost in cooking. Figure 6.9 illustrates how different levels of iron and zinc concentrations in potato (referred to as biofortification levels: Base line, 1, 2, and 3, where base line represents current potato varieties) contribute to the EAR of iron and zinc for women of fertile age who consume 100, 200, or 400 g of potato per day. For reference, women in parts of Rwanda consume over 500 g of potato per day (Personal communication, Harvest Plus), while women in parts of Peru, may consume 800. The iron and zinc levels included in potato product profiles for populations of the Andes or the central African highlands are 45 mg/kg Fe and 35 mg/kg Zn (just over biofortification level 2 in Fig. 6.9) which can be expected to provide 50% of the EAR of both minerals for women consuming 400 g potato a day. This is a considerable increment over the baseline, but has been assessed as feasible by CIP’s breeding program after evaluating genetic diversity, estimating heritability, and realizing significant gains (subsection 6.1.5) in cultivated potato germplasm. Achievement of such genetic gains would only provide 12–15% of the EAR for women who consume 100 g of potato per day or less, as in much of the subtropical lowlands of Asia. The inclusion of a biofortification breeding target to increase Fe or Zn intake from potato by 0.4 or 0.3 mg/day, or reach 50% EAR for this market segment, would require greater genetic gains than presently estimated to be feasible by the interploid breeding strategy undertaken by CIP. Nevertheless, high iron and zinc potatoes have been requested by the national programs of both India and Bangladesh. In such cases, a food systems intervention seeking overall increments of micronutrients from co-staple crops should be considered.


Influence of Fe and Zn concentration and intake of potato on the estimated average requirement for women of fertile age. (a) Definition of baseline (non-biofortified) and incremental concentrations (biofortification levels 1, 2, and 3) of iron and zinc in potato tubers. (b) Percent of EAR for iron for women of fertile age met by consuming 100, 200, or 400 g/day of potatoes with concentration levels 0, 1, 2, and 3
Biofortification targets between level 2 and 3 in Fig. 6.9a have been set as part of the product profiles for table potatoes oriented to tropical highland agro-ecologies and populations with significant levels of anemia, who also consume potato as a main food or staple crop. The value of nutritional traits can be assessed with the disability adjusted life years (DALY metric). DALY is ordinarily used to assess the impact of public health burdens like human disease or illness. It extends the concept of potential years of life lost due to premature death to include equivalent years of “healthy” life lost by virtue of being in a state of poor health or disability. In so doing, mortality and morbidity are combined into a single, common metric (Meenakshi et al. 2007).
4.6 Setting of Breeding Priorities
Once product profiles are defined, breeding programs can proceed to refine trait lists and allocate resources to address those that are most feasible. While goals and priorities are important for breeding programs and the institutions that host and sponsor them, the list of traits a breeder can work with is limited, and in this light, he/she will select a subset of those in the profile for the day-to-day efforts of the program. Breeders may also be aware of a few “must have” traits that do not come up in stakeholder consultation, and since the inclusion of any trait in a breeding effort (whether to change or even maintain trait levels) requires investment, and thus has a cost, these should be made explicit and, ideally, added to the product profile.
Experience and knowledge of existing germplasm are applied to identify the set of attributes to be addressed in breeding. For each attribute, a quantified description of the desired result (maximize, reach specified level, maintain a certain minimum level, etc.) and a unique rank or priority is assigned. Ranking represents the relative effort required to develop the set of traits that will enable the new variety to respond to the opportunity or constraint identified in the targeting exercise and taken on by the breeding program. Relative effort required to address each objective are determined by the proximity of the program’s genetic materials to the needed level of each trait, and the genetic control and relationships among various traits.
4.7 Selection Decisions in Potato Breeding
Because potato is highly heterozygous, a large number of traits segregate every time a cross is made. Every offspring receives a unique combination of desirable alleles at some loci and undesirable alleles at others. No offspring is ever “perfect.” An issue of considerable practical importance, therefore, is deciding which phenotypes (clones; genotypes) are worth keeping, and which should be discarded at each stage of the selection scheme.
Selection decisions are the decisions a breeder makes to use, advance, or discard a selection unit (plant, progeny, progenitor, clone, experimental variety), guided by the breeding priorities, using specific decision-making tools (e.g., assessments of the targeted traits, estimates of breeding value, presence/absence of specific markers, selection indices, combining ability).
In early stages of selection, when the number of plants of each genotype is small, it is important to select stringently only for highly heritable traits. Meaningful assessments of yield, on the other hand, can only be performed in later stages. As selection progresses, and the individuals not meeting breeders’ criteria have been eliminated, it becomes especially important to keep product profiles in mind, i.e., to select not just on the basis of individual traits, but on the suite of traits that collectively define a product, with attention to specific traits at specific stages of the selection scheme.
4.8 Case Study 7: Profile and Selection Decisions for Chipping Potato for Northeast USA
As nations develop there tends to be a shift in potato consumption, away from fresh and more towards processed potatoes, primarily as chips or French fries. Varieties intended for fries or chips ideally have higher levels of starch, so that they absorb less oil when fried. In addition, if potatoes need to be stored in the cold for a long time—typically to ensure a year-round supply of raw product—then it is useful if they are resistant to cold-induced sweetening, as the presence of high levels of glucose and fructose will cause potato slices to turn dark brown when fried.
To improve resistance to cold-sweetening, the New York program has adopted a low-tech approach that could be adopted anywhere. The essence of the approach is this: store potatoes at a temperature where only a small percent will fry to a light color. Intercross those that do, and after 5 years or so, once the population has reached a point where many progeny fry well, lower the cold storage temperature by 0.5–1.0 °C the following year. Intercross the few that fry well under the new, colder regime, and repeat the process. Recurrent selection has proven very effective at improving fry color in the New York program; many clones now fry well out of 3–9 months of storage at 6 °C. Ideally the breeding program will fry from slightly colder storages than those currently in use by regional industry, to help drive down the temperatures that industry can use over time. In general, the colder the storage, the longer that potatoes can be stored.
The two most important attributes in chipping potatoes are starch content (strongly correlated with specific gravity) and fry color, followed by tuber shape and size. About 70% of the New York breeding program effort is dedicated to developing new chipping varieties.
The New York program practices very little selection on seedling tubers, as the performance of plants grown in pots correlates poorly with performance of plants grown in the field. When the first field generation is harvested, the principal selection criteria are for tuber yield, tuber shape, and size (all visually assessed). The ideal chipping potato is round and the size of a baseball. Clones with low yield, or shapes and sizes too far from baseballs, are not selected.
The New York breeding program chips (out of cold storage) every clone that survives 1 or more years of selection in the field, discarding all that do not chip as well as, or better than, the industry standard chipping variety “Snowden.” Absolute fry color can vary from season to season, although the ranking of fry color between clones does not vary greatly.
In the USA a specific gravity of 1.080 is deemed the minimum for processing. Nevertheless, because starch content can vary considerably from season to season (warm nights result in lower starch content than cool nights, all other things being equal), in New York the level of starch is always evaluated relative to that in the widely grown chipping variety, “Atlantic.” The specific gravity of Atlantic is typically above 1.090 in New York, but in some seasons it is less. The New York breeding program will not continue to evaluate a chipping clone whose gravity averages 0.010 or more less than Atlantic, and prefers clones to average 0.005 or fewer points less. New York begins to evaluate specific gravity after a clone’s second year in the field.
Replicated, multisite yield trials begin in the fourth field year, where clones that yield less than 90% of Atlantic are discarded. Clones that exhibit considerably more internal or external defects than “Atlantic” or “Snowden” in yield trials are also discarded, regardless of their yield, fry color, or specific gravity. Clones that survive several years of field trials are then evaluated on commercial farms, and processed in commercial chipping plants, where growers and chipping plants make the final decisions about which clones merit release as new varieties.
4.9 Selection Decisions: Marker-Assisted Selection
4.9.1 How the New York Program Uses Molecular Markers
Even though current NY breeder WDJ was trained as a molecular geneticist, the New York breeding program does not make extensive use of marker assisted selection, primarily because there are not yet many publicly available markers linked to traits of high priority.
One of the two markers currently used in New York is 57R (Finkers-Tomczak et al. 2011), tightly linked to the H1 gene, which confers resistance to race Ro1 of Globodera rostochiensis. Developing potatoes resistant to race Ro1 has been the highest priority of the New York breeding program for the past four decades; almost every cross made in NY has at least one Ro1-resistant parent. All offspring that survive 2 years of visual selection for appearance and yield in the field (about 200 clones, out of 20,000 initially planted) are screened with 57R over the winter. The vast majority of clones lacking the marker are discarded, the only exceptions being those that look especially promising for other high value traits (e.g., excellent chip color combined with high specific gravity).
RYSC3 (Kasai et al. 2000) is the other marker currently used in New York. This marker is tightly linked to Ryadg, which confers immunity to potato virus Y (PVY), and like 57R, is used to screen clones that have survived 2 years of selection in the field. At present the presence/absence of the RYSC3 marker is just one of many data points NY uses to make a decision about each clone, with other data points including chip color, specific gravity, resistance to common scab, and visual assessments of yield and tuber appearance.
Although markers tightly linked to potato virus X (Gebhardt et al. 2006), wart (Gebhardt et al. 2006), verticillium wilt (Bae et al. 2008), Columbia root-knot nematode (Zhang et al. 2007), the pale cyst nematode (Sattarzadeh et al. 2006), and late blight resistance genes (e.g., Colton et al. 2006) have been reported, none of these traits are important enough in New York to influence selection decisions, and thus none of these markers are used routinely.
4.10 Breeding Objectives at CIP
Priority setting exercises (Fuglie 2007; Hareau et al. 2014), complemented by evolving requests from national programs for elite germplasm and candidate varieties, both contributed to CIP’s profiles for new potato varieties tailored to the contrasting agroecologies, cropping, and food systems its program addresses. Two key products conceived in a recent exercise were (1) “Agile Potato” for intensification and diversification of cereal-based systems of Asia and, (2) mid-elevation tropical highlands late blight resistant potato.
CIP’s major breeding populations are oriented to develop table and processing varieties for sustainable potato production in tropical highland and lowland areas faced with either of two basic sets of biotic and abiotic constraints: those encountered in the tropical and subtropical highlands, where late blight, cyst nematodes and frost are limiting, and farmers are increasingly faced with drought; and warmer, drier production areas, that are challenged by viruses, bacterial wilt, drought, root knot nematode and other pests, Increasing requests to support production in mid-elevation tropics of Sub Saharan Africa and Asia as well as temperate regions of Asia have led to the inclusion of heat tolerance and long day adaptation into our breeding objectives, requiring the introduction of new germplasm, screening methods, and trial sites. Present emphasis is on: earliness to enable potato’s full potential to contribute to incomes and productive cropping systems; heat and drought tolerance to enable expansion to mid-elevation regions nearer to population centers, at the same time helping farmers adapt to climate change, and; nutritional value with specific attention to iron and zinc biofortification. CIP aims to reach new thresholds for these traits, while maintaining and broadening levels of disease resistance achieved in the past, and also responding to regional preferences for culinary characteristics and market opportunities.
4.10.1 Case Study 8: Agile Potato for Asia
4.10.1.1 Product Profile
The “Agile Potato” project seeks to develop potato varieties for fresh and processing use in short crop production windows of Asia. The product profile includes 70–90 day maturity; resistance to multiple viruses (PVY and PLRV, with PVX resistance desirable); stress tolerance (heat, drought, and salinity); resistance to late blight; and market-oriented quality (high dry matter and chip processing). The expectation is that varieties with short crop duration that resist disease, require less-frequent seed replacement, thrive in warm seasons, and store well under rustic conditions, can readily be inserted into diverse cropping patterns dominated by rice or wheat. Agile potato varieties would fit into cropping windows currently left fallow in the cereal-based systems of subtropical lowland and temperate regions of Asia. They should help increase system productivity through intensification as well as help meet growing demand for processed potatoes, thus contributing to income generation and employment. Breeding to meet this profile is backed up by trait research to enhance and accelerate breeding methods, and inter-disciplinary research to promote fast-track systems for variety identification and release. With this profile, CIP seeks to develop strategies for sustainable intensification of farming systems, raise awareness and expand consumer demand for potato while diversifying cereal-based diets with potato as a nutritious co-staple or vegetable food. A gamut of research on ecological and socioeconomic consequences of intensification is also planned in order to identify appropriate indicators of sustainability and monitor effects of benefits to intended sectors, including the poor whose livelihoods would be affected by the proposed intervention.
The target is to select 70–80 day heat-tolerant potatoes with competitive yields and more than 18% dry matter and resistance to potato viruses for subtropical lowlands of Southwest Asia. Commercial varieties and elite selections from previous cycles of the lowland tropics virus resistant population (LTVR) are comparators for performance in representative or target selection sites. In the following figures we show a group of selected early clones that performed significantly better than past generations and local varieties for yield and quality at 70 days after planting in warm environments, yielding in a range of 20–40 tons/ha, compared to the best local varieties such as Unica and Reiche (Fig. 6.10) and adequate dry matter content over 18% under the abiotic stress conditions of lowlands (Fig. 6.11).


Scatter plot showing yield of selected clones in a new generation of LTVR population that surpass levels of varieties and clones from previous generation grown under hot conditions of San Ramon, Junin, Peru, and harvested at 70 days after planting


Variation for dry matter content of potato selections in LTVR population across environments. Three hundred LTVR clones were grown in ten hill plots in six locations in Peru and evaluated for dry matter content within 10 days of harvest
Dry matter (DM) content is an important component to be taken into account in the breeding process especially when selection is focused on lowland tropical areas. In addition to the loss of fresh weight yield of tubers associated with high temperatures, the DM content of tubers are also reduced by 1% as the air temperature rises by 1 °C over a temperature variation average of 15–25 °C. This is due to the rapid decline of photosynthesis (Midmore and Rhoades 1988). This tendency of decrease in DM of clones in the LTVR population can be appreciated in Fig. 6.11, showing decreasing DM contents with decreasing altitude and thus increasing temperature of the evaluation sites. However, breeding by recurrent selection for yield as well as DM content in this population has maintained this important trait at between 11 and 19% even in the warmest season, and 15–21% in the high humid jungle site of San Ramon, Peru. This means that genetic diversity is sufficient to permit selection for heat tolerance and genotypes with good yield and adequate DM content can be identified in LTVR in spite of the critical negative correlation between temperature and DM in potato.
Crossing with long day adapted parents and evaluation under simulated long day conditions in Peru permitted the identification of LTVR families and advanced clones with adaptation to higher latitudes, for example of Vietnam where day lengths are intermediate, and Uzbekistan and Tajikistan where they are long combined with high summer temperatures. The same screening approach also helped provide pressure for improving the earliness (70–80 days) of LTVR at low latitudes. The compensatory effect of temperature on day length provided by warm mid-latitude conditions of southern Peru has also been key in breeding for early maturity, which can also be referred to as a relaxed requirement for short days. The identification of a moderate frequency of genotypes as well as superior parents with good performance under higher latitudes suggests that the LTVR breeding strategy is likely to be successful in developing the agile potato for both of its target environments—subtropical lowland and temperate low and mid-elevation regions of Asia.
4.10.1.2 Source Population LTVR
From the 1980s through the late 1990s, CIP endeavored to strengthen its lowland tropics (LT) population with multiple virus resistance developing the lowland tropics virus resistant population (LTVR). Development of the broad-based LTVR population used an open-recurrent selection strategy to incorporate foreign elite and advanced clones into its lowland tropics adapted breeding population. Lowland tropical environments were characterized as having day lengths of 10–14 h, minimum night-time temperatures of 18–20 °C, mean maximum temperatures greater than 25 °C and mean annual soil temperatures at 50 cm depth of 22 °C or more. LTVR population combines the heat tolerance and early bulking ability of S. tuberosum germplasm bred under the summer conditions of the northern Hemisphere (Cubillos and Plaisted 1976) on one hand, with virus resistance from native Andigenum Group germplasm and long day adapted Andigenum Group germplasm, namely Neotuberosum, on the other (Plaisted 1987; Mendoza 1990). The Neotuberosum and Andigenum Group germplasm contributed major genes for extreme resistance to potato virus Y (PVY) and potato virus X (PVX) (Muñoz et al. 1975; Gálvez et al. 1992). Early generation screening was routinely performed to discard PVX and PVY susceptible individuals in any trial in which LTVR families were evaluated.
Testing sites included CIP’s experimental stations in Peru (see Section 6.4.6) which are relatively dry, humid, hot, and hot and humid lowland environments. Experiments comparing the yield stability of diverse clones across these locations revealed that hybrids from S. tuberosum × Neotuberosum crosses performed significantly better than those from S. tuberosum × S. tuberosum or S. tuberosum × Andigenum Group. It is possible that the higher level of heterozygosity together with prior selection of the divergent parental materials for adaptation accounted for the better performance of crosses between S. tuberosum and Neotuberosum clones (Mendoza and Estrada 1979). These authors also reported that S. tuberosum × neotuberosum hybrids are adapted to hot and humid conditions and able to produce tubers at 60–70 days after planting, albeit under short days. These hybrids made a strong contribution to the LTVR population, and thus may account in part for its adaptation to warm conditions. The subsequent cycles of genotypic recurrent selection led to the selection of a number of multiplex PVY and/or PVX extreme resistant progenitors (Mendoza et al. 1996) some of which have since been shown to also possess good GCA for yield and dry matter content. Multiplex refers to the genetic constitution of a polyploid individual having more than one copy of an allele, in this case RRrr or RRRr, assuring the transmission of resistance conferred by a major gene to nearly 100% of its cross progeny. In parallel to population improvement and parental line development, high yielding clones with excellent tuber quality were selected during each recurrent selection cycle.
A second stage of the breeding process aimed to incorporate foreign elite and advanced clones by means of an open-recurrent selection strategy. Assessment of genetic parameters in a North Carolina Design II estimated heritability for resistance to PLRV infection to be 0.54–0.69. Higher estimates were obtained when greenhouse-reared infective aphids were used to conduct controlled inoculations with sprouted tubers before tuber families were planted in the field, suggesting that controlled inoculations minimize errors or escapes because of a more even distribution of inoculum. Additive genetic effects were shown to contribute the most to resistance variability (74%), followed by dominance effects (10%) and additive × environment (location) effects (5%), suggesting that breeding advances could be expected in further cycles of recurrent selection (Salas 2002).
4.10.1.3 Multilocation Testing and Selection Decisions
A stage plan can be used to communicate selection decisions made in early and late stages of breeding. The stage-gate process portrays stepwise decisions with thresholds for each characteristic that govern decisions to keep or discard individuals in the course of a given breeding cycle. In this process, the “gates” describe the collection of thresholds or criteria that must be met if a clone is to pass from one stage of selection to another along the clonal selection scheme outlined in Fig. 6.12, presents a stage plan for selection in the LTVR population to meet the Agile Potato profile.


Stage plan for selection in the LTVR population to meet the Agile Potato profile
Yield and processing quality traits of LTVR clones generated during open-recurrent selection were evaluated in replicated experiments in a range of lowland environments of Peru. Dry matter content was consistently higher in the cool arid lowlands of the coast under irrigation (average = 21%) than in the warm humid area of the central jungle (average = 19%), highlighting the detrimental effect of high temperatures on this trait. As expected, significant G × E interaction was encountered for dry matter content, but also for glucose content and chip color. High yielding clones with good table or processing quality combining virus resistances and heat tolerance have been released by national programs and public institutions in target countries. Varieties of LTVR origin include: Tacna, Costanera, Unica, Reiche, Maria Bonita, and Maria Tambeña released in Peru; Kinga, Meva, Kinigi, Muziranzara, Muruta, Yayla Kizi, Baseko, Enfula, Chamak, Dheera, and IRA-92 in Africa; and Raniag, which was named in the Philippines. Evidence of the wide adaptation of bred materials from this population is provided by the success of the variety “Tacna” in the lowland subtropics of China where it is known as “Jizhangshu 8” and planted on an estimated 133,215 ha in 2015 from which 70,533 ha in Hebei, 26,000 ha in Shanxi, 22,667 in Inner Mongolia, and further 3000 has in the increasingly drought-prone northwest province of Gansu with more than 3000 ha (Gatto et al. 2018). Additional evidence of broad adaptation comes from the variety Unica that is now grown in at least six countries.
4.10.1.4 Breeding Priorities
Following CIP’s multidisciplinary conceptualization of the agile potato, the potato breeding program refreshed its strategy by setting priorities for combining the needed traits in subgroups of the LTVR population. The inclusion of resistance to PVY in the LTRV population and selection for highland tropical as well as subtropical lowland conditions to enable seed production has had a cost in terms of earliness even after three to four cycles of recurrent selection. The infusion of Tuberosum germplasm and selection under heat and mid-long days, for plant type and stability from early generations is strategy to improve earliness in the LD (long day) and HT (heat tolerant) groups. In this interest, tuber quality becomes an issue and attention is needed to keeping dry matter high and glycoalkaloids, which provide a bitter flavor to potato and can be toxic, low.
Often unrecognized and rarely controlled, the accumulation of viruses in seed and ware tubers is perhaps the most serious constraint to economical potato production in the tropics. Breeding for resistance to the world’s most important potato virus PVY is possible due to the availability of major genes (Ryadg, Rysto) that confer extreme resistance, and for which selectable markers are available. On the other hand, developing effective levels of resistance to PLRV is one of the more challenging components of the Agile Potato profile. Most sources of resistance to PLRV are quantitatively inherited and provide partial resistance to infection and/or accumulation of the virus. Despite moderate heritability, the frequency of resistant clones in advanced populations has been low. Thus, a relatively high priority is given to incorporating a new, highly heritable source of high levels of resistance to PLRV, designated Rladg recently identified in Andigenum Group germplasm into advanced populations that already carry extreme resistance to PVY at high frequency.
New sources of PLRV resistance, namely, accessions LOP-868, HUA-332 and OCH-7643, confirmed following mass selection in S. tuberosum Andigenum Group germplasm held at CIP, were shown to have exceptionally high GCA for this trait (Mihovilovich et al. 2007). Further characterization of these sources has been the basis of supporting trait research, the benefits of which are expected to boost levels and frequencies of PLRV resistance in the LTVR and other potato populations. Using a population of dihaploid individuals derived from LOP-868, Rladg was mapped to potato chromosome V, with high heritability demonstrated to be due to its multiple resistance mechanisms and a duplex allelic conformation of a major dominant gene in the donor accession (Velásquez et al. 2007). A PCR based marker (RGASC850) located 2 cM from the gene has been developed to facilitate the introgression and tracking of this major gene in advanced breeding populations (Mihovilovich et al. 2014). This marker has been incorporated into a multiplex PCR system for assisted selection of PLRV and PVY resistance using previously available information on Ryadg from Kasai et al. (2000).
Table 6.2 presents the attributes, profile and breeding priorities contributing to the Agile Potato program and outlines some of the key research that supports the objective. Breeding priorities represent the relative effort dedicated to each attribute of the profile, given present status of the LTVR population.
4.10.1.5 Multi Trait Selection for Increased Earliness and Tolerance to Abiotic Stress
Most recently, emphasis in improving CIP’s LTVR population toward the agile potato has targeted increased earliness and tolerance to abiotic stress. A new earlier maturing and abiotic stress tolerant generation that combined superior LTVR progenitors with tuberosum varieties was generated. Accelerated selection based on multilocation trials and simultaneous screening for abiotic stress (heat, drought and simulated long days) in divergent and stressful environments as well as screening for PVY resistance was implemented in Peru. The scheme requires only 4 years of selection and evaluation from the true seed progeny generation until the identification of advanced clones (see Fig. 6.6).
4.10.1.6 Evaluation and Selection Sites
CIPs proximity to significant agroecological diversity in Peru is an asset for the identification of patterns and trends of potato genotypes’ response to environments. Potato is grown in coastal, high jungle and highland ecologies ranging from sea level to 3500 masl, providing wide variation in weather patterns including temperature, humidity and rainfall. Most environments have short daylengths of around 11–12 h but the southern extreme at 17°S, provide daylength of about 14 h, which considering high summer temperatures approach threshold conditions for long day adaptation. Three experiment stations (San Ramon, Huancayo and La Molina) and additional sites accessed by local agreements (Table 6.3 provide evaluation conditions representative of the tropical lowland and highland potato environments targeted by agile potato. Indeed, potato seed for the subtropical lowlands is often produced at higher, vector-free, locations, requiring adaptation of the same variety to both highland and lowland conditions. Moving potato populations through the biotic and abiotic stresses present among these sites, and particularly the combination of temperature and day length, permit the identification of potato genotypes with wide adaptability and stability for yield.
4.10.1.7 Trait Research in Support of the Agile Potato
Proximal sensing for efficient development of the agile potato: Remote sensing technologies are increasingly being used to assess crops by breeders and physiologists. These technologies have the potential to simplify evaluation of large breeding populations and improve selection efficiency over traditional field measurements. Different vegetative indices combined together may, for example, improve yield prediction (Rodriguez Junior et al. 2014). CIP’s work in this area has aimed to explore genetic variation for the canopy temperature differential (CTD) and Normalized Difference Vegetation Index (NDVI) measurements and their relations with tuber yield components using 27 segregating families of the LTVR population under water stress, toward identifying families with tolerance to drought at early stages of selection. A high negative correlation (−0.66) of NDVI with Harvest index-fresh weight was observed, as were a high positive correlation (0.74) with plant wilting, a moderately high (0.50) correlation with plant vigor under drought conditions, and a significant negative correlation (−0.5) of CTD with tuber yield (tons/ha). Remote sensing techniques may provide a way to save resources and accelerate genetic gains through the early identification of families with tolerance to abiotic stress in a breeding program.
Trait research for understanding of complex, underground development: In breeding for a short vegetative cycle, CIP emphasizes bulking-based maturity, as opposed to foliar maturity or senescence. Tuber bulking results from two basic processes, tuber initiation and tuber growth (Radley et al. 1961). Like Andigenum Group potatoes, most varieties and clones from CIP’s program yield significantly before physiological maturity, and development and harvest date cannot be assessed on foliar senescence alone. Indeed, if CIP-bred varieties are harvested when foliage has senesced, they will generally be well beyond the optimum time for harvest. Timing and duration of bulking vary with location, environmental factors, and genetics. Early tuber initiation and growth are necessary for acceptable production in areas or varieties in which potatoes are often harvested prior to physiological maturity.
To help provide early-maturing, resilient potatoes to farmers in medium and high latitudes of Asia, a panel of 162 CIP breeding lines was characterized for bulking behavior under four environmental conditions in field plots in La Molina, Peru. The environmental conditions for this study were: simulated long (16 h), and short (12 h) day lengths, in each of two seasons—cool winter and hot summer. The traits evaluated in the four field plots include yield (number and weight of tubers on two harvest dates) and dry matter content, in addition to which tuber induction was assessed by the cutting method of Ewing (1992). Tuber bulking ratio was calculated as the number of medium-sized and large tubers over the total number of tubers (tiny + small + medium + large) multiplied by the ratio of tuberized plants. Phenotypic assessment according to CIP’s Protocol for tuber bulking maturity assessment of elite and advanced potato clones (https://research.cip.cgiar.org/potatoknowledge/bulking.php) enabled the classification of genotypes by maturity through cluster analysis according to Evanno and Regnaut (2005), and thereby, the identification of clones for which tuberization and bulking were more or less influenced by heat and photoperiod, including a small set of selections classified as day neutral.
Genotyping of the panel with version 1 of the SolCAP infinium array (8303 single nucleotide polymorphism (SNP) markers) enabled the identification of genomic regions and SNP markers influencing potato tuberization and bulking related traits affected by photoperiod and high temperatures. Prerequisites for this genome-wide association study (GWAS) included the calculation of the population membership matrix and SNP-based kinship relationship matrix and the estimation of linkage disequilibrium (LD decay to r2 = 0.2 over 0.3 Mb) for the panel. GWAS-associated SNP jointly explained up to 60% of the phenotypic variation for the various traits, Therefore, MAS could be efficiently used for pre-screening of seedlings before further field evaluation, and contribute to shift the mean of the selection population for adaptation to warm, long day environments.
The same information was also used in a pilot study of genomic selection (GS). The underlying concept in GS is to model the entire complement of quantitative trait loci (QTL) effects across the genome to produce a genomic estimated breeding value (GEBV) from which progenitors or clones can be selected after genotyping. This pilot study tested several models proposed by Endelman (2011) to assess the feasibility of applying GS to improve selection of parental clones for early tuberization and bulking under long day-length. Prediction accuracies of ≥0.3 can be regarded as sufficiently high to merit implementation of GS (Heffner et al. 2010). Sufficiently high values were attained for tuberization induction, stolon length, bulking ratio at 75 and 90 days, and marketable tuber number at 75 days. As all markers are simultaneously fitted to capturing most of the trait heritability, GS can be expected to help reduce the length of selection cycles aimed at improving traits for which accurate phenotyping can be performed.
5 Prospects for True Hybrid Potato Breeding
Several countries and institutions have conducted research proposing the production of potato by sexual or botanical seed. Since the vegetative (tuber) planting material of clonally propagated potatoes is typically called seed potato, the term true potato seed (TPS) was introduced for tetraploid potato varieties produced from botanical seed. TPS aimed to overcome difficulties associated with production of vegetative seed including slow multiplication rate, accumulation of pathogens, and the bulky and perishable nature of tuber seed (Kidane-Mariam et al. 1985; Golmirzaie et al. 1994; Almekinders et al. 2009). As potato is a cross-fertilizing crop developed from heterozygous parents that do not “breed true,” TPS is heterogeneous and thus variable for a large number of characteristics. While this variability is the source of variation for a clonal variety program, it is generally undesirable in varieties per se.
True hybrid potato breeding , on the other hand, aims at developing uniform true seed (TS) hybrid varieties from complementary superior inbred stocks developed at the diploid level. It would combine the advantages of TPS, (large numbers of disease-free propagules), diploid breeding (simplified genetics), and homogeneity (by way of combining inbred instead of heterozygous parents) in a new potato breeding system akin to that of maize.
Preliminary reports of productive diploid hybrids (de Vries et al. 2016), and the advantages of “re-inventing potato” as an inbred line-based crop (Jansky et al. 2016) make this effort one of the most intriguing developments in potato breeding over the past decade. Diploid inbreeding-tolerant potatoes would allow for considerably more systematic incorporation of new genes and traits, such as by backcrossing to fixed genotypes, as well as the possibility of substantial yield gains by crossing between well-defined heterotic groups. Working with diploid potatoes would also greatly facilitate the use of desirable recessive alleles. In principle, the low multiplication rate of clonally propagated potato can be circumvented by moving to diploid hybrids, where true seed from inbred × inbred crosses can produce vast numbers of genetically identical offspring in a short period of time.
Several steps need to be taken before diploid potato inbred line breeding might become routine. Most important is eliminating the self-incompatibility system, so that inbred lines can be readily developed. Recently, the dominant self-incompatibilty locus inhibitor allele (Sli) identified in certain self-compatible (SC) variants of the diploid wild potato Solanum chacoense (Hosaka and Hanneman 1998) has been used to overcome self-incompatibility (SI) and develop SC inbreds directly from the original source or by introgression to SI 2x cultivated germplasm (Phumichai and Hosaka 2006; Jansky et al. 2014). Additional research may be needed in order to manage SC as a trait in potato breeding, to improve the vigor of potato seedlings and to assure male fertility in diploid male parents.
Investment in inbreeding seems to be greater so far in the private than in the public sector. Nevertheless, some public breeding programs including CIP’s have begun to seek some of the advantages that this novel strategy can offer to breeding and genetics for potato improvement. CIP has undertaken research to develop (1) a self-compatible) 2x breeding platform for broadening the genetic base of potatoes by hybridization across genepools, and (2) inbred genetic stocks for gene discovery and trait capture. Initial stocks and experience gained in the process of this research will contribute to longer term goals of developing complementary superior diploid inbred parental lines that will produce uniform, heterotic true seed 2x varieties when combined by crossing.
6 Variety Testing and Seed Links
6.1 How the Breeding Program Is Integrated with Seed Production in the State of New York in the United States
In addition to a public sector breeding program, the state of New York also maintains a public sector nuclear seed farm (the Uihlein Farm, in Lake Placid NY), which produces clean seed for commercial seed growers in the region. Each year the NY breeding program identifies up to five promising selections—typically clones that have been evaluated for at least 6 years in the field—and sends them to the Uihlein Farm. The Uihlein Farm first tests each clone for viruses, removing any that are present, and then, after a few years, depending on the multiplication scheme chosen, offers minitubers or limited-generation field grown tubers of these selections for sale.
Over time the breeding program has learned that it is important not to discuss promising clones with local industry until seed will soon be, or already is, available. If a clone elicits excitement, and growers ask for seed yet find none, they quickly lose interest, and it is very difficult to rekindle interest once seed does become available. When a clone is discussed before seed is available from the Uihlein Farm, the breeding program strives to provide up to 1000 kg of seed for commercial ware and chipstock growers to evaluate on their farms in the interim.
The NY seed farm bears some risk in this arrangement, as the breeding program typically discovers, over the course of additional years of evaluation, serious weaknesses in some of the clones being multiplied, rendering the clone unmarketable. On the other hand, the NY seed farm also benefits when it can be the first supplier of a new variety.
6.2 Regional Trials for Variety Assessment
Procedures, interfaces and responsibilities for trialing elite potato clones for variety release vary in degree of organization or formality across and sometimes within Countries.
As the aim of plant breeding is to produce new, improved varieties, breeders should be aware of and anticipate meeting the several steps required for variety recognition, release and registration.
Depending on program structure and target territory) and the complexity of the selection criteria, potato variety breeding typically involves a minimum of 6–7 years (and often more) of crossing and selection work, followed by a year or 2 of early testing to identify superior clones. The next step is the entry of candidate varieties into official variety registration recommendation trials. Programs in which breeding is done in the territory of release are more straight-forward in this respect than international programs in which additional time is required for quarantine exchange and bulking of seed in a new location. Depending on access to infrastructure and communication about objectives, this can take 2–5 years. The public and private sector differ in this respect since while public programs generally provide elite materials as true seed families or a small number of in vitro plants for research and breeding that may lead to local selection, breeding companies provide a reasonable quantity of seed of known varieties along with support and incentives for their testing and release.
6.3 Case Study 9: Regional Trials for Potato Variety Development in the United States
Northeast Regional Potato Trials—trialing system is a network for testing elite clones as candidate varieties. At present public-sector potato breeding in the US is divided into four regions—east, central, west and south. Each region has a regional trialing network, in which the agronomic performance of advanced clones is evaluated across a wide geographical range. The trial serves both to identify those relatively uncommon clones with broad adaptation, which are most likely to become commercially acceptable, and to identify varieties that may serve the needs of narrow geographic areas that have no breeding program of their own. Taking the eastern US as an example: the system consists of four public sector breeding programs—2 based in Maine, one in North Carolina, and 1 in New York—and 11 evaluation sites, 2 in NY and 1 each in Maine, North Carolina, Pennsylvania, Florida, Virginia, Ohio, Maryland and occasionally the Canadian provinces of New Brunswick and Prince Edward Island. Evaluators at each site rate clones using standardized scoring scales for yield, specific gravity, shape, internal and external defects, maturity, texture of tuber skin, and overall appearance, and upload the data to a common web portal (potatoes.ncsu.edu/NE.html). The originating breeding programs use the data to help make selection decisions, while regional evaluators use it to help inform local growers about which new varieties will perform best in their region. Breeders and evaluators also meet once a year to discuss, in person, the performance of clones in the trial the preceding year, and to make incremental adjustments in how the trial data is collected, analyzed and reported. Over the past decade separate national programs in the US, focused on chip and french fry variety development, have built upon the standardized framework of the eastern region trial network, with that data housed at potatoesusa.mediusag.com.
6.4 How Does the New York Program Decide When to Release a Variety?
In many countries a new potato variety cannot be legally grown until it has been formally evaluated in registration trials, found to be distinct, uniform and stable (DUS), and shown to have “value for cultivation and use.” Decisions on all of these characters are made by an independent regulatory body, not the originating breeder.
In contrast, in the United States there is no legal requirement for comparable tests before a potato variety can be grown. (Although there is a requirement for DUS data, if a US breeder wants to procure plant breeders rights.) While some US universities have panels of breeders that evaluate candidate varieties, and can deny release of a variety by a colleague, others, like Cornell University in New York, have no formal potato variety release process at all.
Is either of these approaches preferable? Coauthor Walter de Jong is comfortable to operate in an unregulated environment. In his view the uniformity and stability tests are essentially meaningless for a clonally propagated crop. In addition, while he recognizes that evaluating distinctness is important to prevent anyone from re-releasing varieties developed by others, as long as two highly heterozygous autotetraploids have been crossed, all offspring will be distinct, as so many loci segregate. In other words, it is trivial to create a distinct potato. Variety release decisions in New York are made, instead, by gauging stakeholder interest. Prior to formal release, seed growers are permitted to grow candidate quantities under an evaluation license. When a candidate variety reaches a threshold, typically 10 ha or more of seed across all seed growers, and the seed growers ask for the variety to receive a formal (nonexperimental) name, it is released. One specific situation where the New York program will not release a variety, even if there is industry interest, is if tuber glycoalkaloids are too high (>20 mg/100 g tuber fresh weight).
6.5 Case Study 10: All India Coordinated Crop Improvement Project and Release of Kufri-Lima from LTVR Population
In India, state and national systems for verity testing and release coexist, and to some degree depend on each other, and these policies serve as a model for other countries in south Asia. Formal testing of a candidate variety by the All India Coordinated Crop Improvement Projects (AICCIP) , and the subsequent recommendation of the variety for release by the AICCIP are required before variety can be released by the Central Sub-Committee that permits notification and thereby enables seed production and trade. In the case of potato, a minimum of 3 years of multilocational trials and assessment for Value for Cultivation and Use (VCU) take advantage of 16 locations across the country. These locations use standardized design and data templates and report on performance as well as meteorological conditions. Experiments are designed for Rabi (winter) season for irrigated areas such as Central, Northern, and Eastern regions (Indo-Gangetic Plains). Plateau regions (700–1200 masl) and hilly regions (1500–3000 masl) are represented in the multilocational trial series.
A new variety should be suitable for specified agro-climatic and soil conditions, have an ability to withstand typical stress conditions, and have tolerance/resistance to pests and diseases. It should also show distinct advantages over the existing equivalent released varieties, a process facilitated by standard formats for data collection. The breeders or Principal Investigators and Zonal Co-ordinators attend the meeting to provide wider information on the variety. The Director of Crop Development Programme is invited to provide information on the response of farmers to promotion and demonstration trials if they were conducted. The release proposal proforma requires the breeder to ensure availability of enough seed stock for seed multiplication on at least 10 ha.
An interesting feature of the All India trials project is that it reports on potato varieties as components of cropping systems, and not only on potato as a commodity. The pertinent proforma includes information on the productivity and profitability of potato and each of different crops in intercropping systems. Long-term experiments address issues of nutrient cycling and crop management that offer valuable methodology and possibly valuable data sets and recommendations for CIP’s concern for the resilience of its Agile Potato proposition, proposing intensification of cereal-based systems with attention to sustainability.
Recent emphasis of collaboration between CIP and the Central Potato Research Institute (CPRI) of India have been on identifying early-maturing, heat tolerant, virus-resistant varieties suitable for intensifying and diversifying rice-based production systems. CIP introduces elite clones as in vitro plants as well as true seed from crosses between parents with good GCA for features of performance in subtropical lowland environments for used in breeding or direct identification of varieties for the vast Indo-Gangetic Plains and hilly regions of southwest Asia. The release of Kufri-Lima in October 2017, bred as CIP397065.28, is an exciting result of this collaboration. Potato virus are a serious problem for farmers in India, particularly in Gujarat, Rajasthan, Madhya Pradesh, Chhattisgarh, Odisha, West Bengal, and East Uttar Pradesh, where temperatures are higher in the winter season. Crop losses due to viruses range between 20 and 50%, and farmers from these areas traditionally need to import seed from Northern India (Punjab). “Farmers usually have to wait until temperatures drop to plant potatoes, but because of its tolerance to heat, Kufri Lima can be planted a full 20–30 days earlier than other local varieties.” Earlier planting means earlier harvests, giving Kufri Lima farmers the ability to sell their potatoes at a premium price before other varieties hit the market. Farmers can expect to be paid 40–50% higher prices than those who harvest potatoes during the normal season. Heat tolerance and early maturity mean that Kufri Lima farmers can invest their earnings into a second winter harvest helping them to improve the overall productivity of the cropping system, while virus resistance may enable seed production closer to home. https://www.potatopro.com/companies/central-potato-research-institute-cpri.
Kufri-Lima was introduced to CPRI as CIP397065.28 in 2006 after initial selection in Peru. It was evaluated for adaptation, quality, and resistance to degeneration along with several other selections from the LTVR population with the support of ICAR (International Council of Agricultural Research), Central Potato Research Institute, GIZ/BMZ and USAID.
6.6 Variety Release and Registration
Keen awareness of timelines, procedures and actors involved in decisions on the release of varieties from elite clones and of policies and practices that permit or enable seed production is essential for the uptake of new varieties and subsequent benefit of their component and composite traits. Country specific systems for variety testing, release and dissemination of seed to farmers are governed by sets of national laws, scientific guidelines, norms, and standard practices which together can be termed “Regulatory Frameworks.”
Registration is often required for seed to be multiplied and marketed in a country or territory or for exportation outside the territory. Release is intended to make a newly developed variety available to the public for general cultivation in the regions for which it is adapted. Even though they are not directly responsible, breeders should be aware of the designated service responsible for recommending the registration of new varieties in their target locations, as well as the Basic information required for application. Additional efficiencies can be realized when breeding programs are aware of and contribute to the availability of Basic material required for application and variety release. Knowledge of the agency with which a reference sample has to be deposited, up to date reference to the format for information required and the quantity of seed to be available at time of release vary be country or state. Depending on the territory in question, the designated service responsible for approving the registration of new varieties is usually a ministry or department of agriculture, which may have crop specific rules and regulations for variety development. In India and Bangladesh, for example, potato is among the crops for which a special category called “notified” is required for varieties to enter seed production schemes.
Additional policies, processes and agencies are those responsible for demonstration or promotion trials for pre-released and released varieties where required; maintenance of certification standards; and the production and storage of foundation seed and provision of seed of new varieties. Policy regulating seed trade is of less direct concern, though also important to breeding programs. An excellent reference for procedures and policy for variety testing and release can be found at http://www.coraf.org/wasp2013/wp-content/uploads/2013/07/ECOWAS_VAR_REGIST_MANUAL_SEP_081.pdf.
6.7 Regional Frameworks for Variety Release
Beyond national seed regulatory systems, several regional frameworks are being developed or implemented across countries. Advantages of regional frameworks include improved access by farmers to seed of improved varieties at affordable prices, avoidance of repetitive national testing, making seed trade easier, faster, and less expensive for introduction of new varieties, timely availability of quantity and high quality of seed and choice of variety needed. Regional frameworks encourage individual countries to adopt national seed legislation and establish national variety catalogues that contribute to growers’ awareness of new varieties. Regional registration can result in a larger market share for varieties and reduced time and cost for national variety release when second and third country releases are simplified by admissibility of available performance trial data.
Fast track options for variety registration at regional level rely on policies and communication practices that recognize prior registration in one member country and one season testing as sufficient for variety release in a second one. Similarly relevant are federal and state or provincial procedures that complement each other, enabling the benefits of new varieties to be realized across states or countries. Mechanisms are also needed whereby release proposal on the basis of data from state and farmers’ field trials could be used for the zonal release of a variety, across an agro-ecological zone that covers more than one state or one country (Tripp et al. 1997). These same authors conclude that adjustment to seed regulatory frameworks is necessary because of significant changes in national seed systems, including: reductions in budget for public agricultural research; the failure of many seed parastatals; increasing concern about plant genetic diversity; pressure for the establishment of plant variety protection; the increasing contributions of commercial seed enterprises; and the emergence of innovative local level variety development and seed production initiatives.
A report of test cases in Africa supported by Syngenta documented the inclusion of three new potato varieties in the COMESA (Common Market for Eastern & Southern Africa) regional catalogue in 2016 (https://www.syngentafoundation.org/sites/g/files/zhg576/f/seeds_policy_regional_variety_release_test_cases_317.pdf). Notably, these are the first non-maize varieties to be included in the region’s list.
7 Concluding Remarks
Potato breeding is at a crossroad. Gene editing and large scale genotyping are at hand, and true hybrid breeding appears feasible, even for this complex polyploid and traditionally clonally propagated crop. Together with pre- and post-Mendelian methods, these advances promise to help accelerate genetic gain to nourish a growing population, help preserve the environment, and confront economic and climate constraints. Sustained breeding and supporting trait research will assure potato’s role in a more comprehensive and integrated agriculture in which quality and resilience, in addition to the historical emphasis on yield, are major goals. Community efforts including the development of cross-species tools and research aimed at understanding the principles underlying crop performance will be needed to meet growing demands on agriculture. The programs illustrated here use crop ontologies to support understanding of traits and attempt to exploit heterosis, inter-ploidy breeding, and accelerated selection schemes to reach their goals. Beyond this, research on root systems, G × E interaction and photosynthetic efficiency merit attention. The lengthy 12–20 years required to develop and release a new potato variety, coupled with climate variability, call for intensified research on selection strategies and choice of breeding method. The use of marker-assisted and genomic selection, the possibilities offered by inbreeding coupled with gametophyte selection and hybrid development, “big data” and efficient real-time phenotypic data collection and analysis are most compelling at present. Finally, collaboration across sites and disciplines and the utilization of statistical models for assessing genotypic adaptation and breeding value are critical to the efficient development and deployment of better potato varieties.
References
Almekinders CJM, Chujoy E, Thiele G (2009) The use of true potato seed as pro-poor technology: the efforts of an international agricultural research institute to innovating potato production. Potato Res 52:275–293
Bae J, Halterman D, Jansky S (2008) Development of a molecular marker associated with Verticillium wilt resistance in diploid interspecific potato hybrids. Mol Breed 22:61–69
Bernardo R (2002) Breeding for quantitative traits in plants. Stemma Press, Woodbury
Bonierbale M, Plaisted RL, Tanksley S (1993) A test of the maximum heterozygosity hypothesis using molecular markers in tetraploid potatoes. Theor Appl Genet 86:481–491
Bonierbale M, Amoros W, Burgos G, Salas E, Suarez H (2007) Prospects for enhancing the nutritional value of potato by plant breeding. African Potato Association conference proceedings, Alexandria, Egypt, vol 7, pp 26–46
Butler DG, Cullis BR, Gilmour AR, Gogel BJ (2009) ASReml-R reference manual. v. 3. Queensland Department of Primary Industries & Fisheries, Brisbane
Chase SS (1963) Analytic breeding in Solanum tuberosum L.—a scheme utilizing parthenotes and other diploid stocks. Can J Genet Cytol 5:359–363
Colton L, Groza H, Wielgus S, Jiang J (2006) Marker-assisted selection for the broad-spectrum potato late blight resistance conferred by gene RB derived from a wild potato species. Crop Sci 46:589–594
Comai L (2005) The advantages and disadvantages of being polyploid. Nat Rev Genet 6:836–846
Comstock RE, Robinson HF (1948) The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4:254–266
Cubillos A, Plaisted R (1976) Heterosis for yield in hibrids between S. tuberosum spp. Tuberosum and S. tuberosum spp. Andigena. Am Potato J 53:143–150
De Jong H, Rowe PR (1971) Inbreeding in cultivated diploid potatoes. Potato Res 14:74–83
De Nettancourt D (1977) Incompatibility in angiosperms. Springer, Berlin
Endelman JB (2011) Ridge regression and other kernels for genomic selection with R package rrBLUP. Plant Genome 4:250–255
Evanno G, Regnaut S (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620. https://doi.org/10.1111/j.1365-294X.2005.02553.x
Ewing EE (1992) Critical photoperiods for tuberization: a screening technique with potato cuttings. Am Potato J 55:43–53
Finkers-Tomczak A, Bakker E, de Boer J, van der Vossen E, Achenbach U, Golas T, Suryaningrat S, Smant G, Bakker J, Goverse A (2011) Comparative sequence analysis of the potato cyst nematode resistance to locus H1 reveals a major lack of co-linearity between three haplotypes in potato (Solanum tuberosum ssp.). Theor Appl Genet 122:595–608
Frankel R, Galun E (1977) Pollination mechanisms, reproduction, and plant breeding. Springer, New York
Fuglie K (2007) Priorities for potato research in developing countries: results of a survey. Am J Potato Res 84:353–365
Gaiero P, Mazzella C, Vilaró F, Speranza P, de Jong H (2017) Pairing analysis and in situ hibridisation reveal autopolyploid-like behaviour in Solanum commersonii × S. tuberosum (potato) interspecific hybrids. Euphytica 213:137. https://doi.org/10.1007/s10681-017-1922-4
Gallais A (2004) Quantitative genetics and selection theory in autopolyploid plants. INRA, Paris
Gálvez R, Mendoza HA, Fernández-Northcote EN (1992) Herencia de la inmunidad al virus Y de la papa (PVY) en clones derivados de Solanum tuberosum spp. Andigena. Fitopatología 27:8–15
Gardner CO, Heberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22:439–452
Gatto M, Hareau G, Pradel W, Suarez V, Qin J (2018) Release and adoption of improved potato varieties in Southeast and South Asia. International Potato Center (CIP) Lima, Peru, 42p. Social Sciences Working Paper No. 2018-2. ISBN 978-92-9060-501-0
Gebhardt C, Bellin D, Henselewski H, Lehmann W, Schwarzfischer J, Valkonen J (2006) Marker-assisted combination of major genes for pathogen resistance in potato. Theor Appl Genet 112:1458–1464
Golmirzaie A, Malagamba P, Pallais N (1994) Breeding potatoes based on true seed propagation. In: Bradshaw J, MacKay G (eds) Potato genetics. CAB International, Wallingford, pp 499–512
Griffing B (1956) Concepts of general and specific combining ability to relation to diallel crossing system. Anst J Biol Sci 9:463–493
Grüneberg WJ, Mwanga R, Andrade M, Espinoza J (2009) Chapter 5: Selection methods Part 5: Breeding clonally propagated crops. In: Ceccarelli S, Guimarães EP, Weltzien E (eds) Plant breeding and farmer participation. FAO, Rome, pp 275–322
Hareau G, Kleinwechter U, Pradel W, Suarez V, Okello J, Vikraman S (2014) Strategic assessment of research priorities for potato. CGIAR Research Program on Roots, Tubers and Bananas (RTB), Lima (Peru). RTB Working Paper 2014-8. Available online at: www.rtb.cgiar.org
Harlan IR, de Wet IMI (1971) A rational classification of cultivated plants. Taxon 20(4):509–517
Hayman BI (1954) The theory and analysis of diallel crosses. Genetics 39:789–809
Heffner EL, Lorenz AJ, Jannink J l, Sorrells ME (2010) Plant breeding with genomic selection: Gain per unit time and cost. Crop Sci 50(5):1681–1690
Hirsch C, Hisrch CD, Felcher K, Coombs J, Zarka D, VanDeynze A, DeJong W, Veilleux RE, Jansky S, Bethke P, Douches DS, Buell CR (2013) Retrospective view of North American potato (Solanum tuberosum L.) breeding in the 20th and 21st centuries. G3 Genes Genomes. Genetics 3:1003–1013
Hirsch CD, Hamilton J, Childs K, Cepela J, Crisovan E, Vaillancourt B, Hirsch CN, Habermann M, Neal B, Buell CR (2014) Spud DB: a resource for mining sequences, genotypes, and phenotypes to accelerate potato breeding. Plant Genome 7(1):1–12
Hosaka K, Hanneman RE (1998) Genetics of self-compatibility in a self-incompatible wild diploid potato species Solanum chacoense. 1. Detection of an S locus inhibitor (Sli) gene. Euphytica 99:191–197. https://doi.org/10.1023/A:1018353613431
Jansky SH, Chung YS, Kittipadukal P (2014) M6: a diploid potato inbred line for use in breeding and genetics research. J Plant Regist 8:195–199. https://doi.org/10.3198/jpr2013.05.0024crg
Jansky SH, Charkowski AO, Douches DS, Gusmini G, Richael C, Bethke PC, Spooner DM, Novy RG, De Jong H, De Jong WS, Bamberg JB, Thompson AL, Bizimungu B, Holm DG, Brown CR, Haynes KG, Sathuvalli VR, Veilleux RE, Miller JC, Bradeen JM, Jiang JM (2016) Reinventing potato as a diploid inbred line–based crop. Crop Sci 56:1–11
Kasai K, Morikawa Y, Sorri VA, Valkonen JP, Gebhardt C, Watanabe KN (2000) Development of SCAR markers to the PVY resistance gene Ryadg based on a common feature of plant disease resistance genes. Genome 43:1–8
Kempthorne O, Curnow RN (1961) The partial diallel cross. Biometrics 17:229–250
Kidane-Mariam HM, Mendoza H, Wissar R (1985) Performances of true potato seed families derived from intermating tetraploid parental lines. Am Potato J 62:643–652. https://doi.org/10.1007/BF02853473
Kleinwechter U, Hareau G, Suarez V (2014) Prioritization of options for potato research for development—results from a global expert survey. CGIAR Research Program on Roots, Tubers and Bananas (RTB), Lima (Peru). RTB Working Paper 2014-7. Available online at: www.rtb.cgiar.org\
Levin DA (1983) Polyploidy and novelty in flowering plants. Am Nat 122:1–25
Lynch M, Walsh B (1998) Genetics and analysis of quantitative traits. Sinauer Associates, Sunderland
Meenakshi JV, Johnson N, Manyong VM, De Groote H, Javelosa J, Yanggen D, Naher F, Gonzalez C, Garcia J, Meng E (2007). How cost-effective is biofortification in combating micronutrient malnutrition? An ex-ante assessment HarvestPlus Working Paper No.2/. HarvestPlus, Washington, DC. www.harvestplus.org
Mendiburu AO, Peloquin SJ (1977) Bilateral sexual polyploidization in potatoes. Euphytica 26:573–583
Mendoza HA (1990) Mejoramiento poblacional. Una estrategia para la utilización del germoplasma de papa. En: Mejoramiento de papa en el Cono Sur, pp 47–62
Mendoza HA, Estrada NR (1979) Breeding potatoes for tolerance to stress: heat and frost. In: Mussel H, Staples RR (eds) Stress physiology in crop plants. Wiley, New York, pp 227–262
Mendoza HA, Sawyer RL (1985) The breeding program at the International Potato Center (CIP). In: Russell GE (ed). Progress in plant breeding, 1. London, Butterworths, 325 p
Mendoza HA, Mihovilovich EJ, Saguma F (1996) Identification of triplex (YYY) potato virus Y (PVY) immune progenitor derived from Solanum tuberosum spp. Andigena. Am Potato J 73(1):13–19. ISSN 0003-0589
Midmore DJ, Rhoades RE (1988) Applications of agrometeorology to the production of potato in the warm tropics. In: Rijks D, Stigter CJ (eds) Agrometeorology of the potato crop. International Society for Horticultural Science (ISHS); International Potato Center (CIP); World Meteorological Organization (WMO); European Association for Potato Research; Royal Netherlands Meteorological Institute, Wageningen, pp 103–136. Acta Horticulturae. no. 214
Mihovilovich E, Alarcón L, Pérez AL, Alvarado J, Arellano C, Bonierbale M (2007) High levels of heritable resistance to potato leafroll virus (PLRV) in Solanum tuberosum subsp. Andigena. Crop Sci 47(3):1091–1103
Mihovilovich E, Aponte M, Linqvist-Kreuze H, Bonierbale M (2014) An RGA-derived SCAR marker linked to PLRV resistance from Solanum tuberosum ssp. Andigena. Plant Mol Biol Report 32:117–128. https://doi.org/10.1007/s11105-013-0629-5
Muñoz FJ, Plaisted RL, Thurston HD (1975) Resistance to potato virus Y in S. tuberosum spp. Andigena. Am Portao J 52:107–115
Oakey H, Verbyla AP, Cullis BR, Wei XM, Pitchford WS (2007) Joint modeling of additive and non-additive (genetic line) effects in multi-environment trials. Theor Appl Genet 114:1319–1332. https://doi.org/10.1007/s00122-007-0515-3
Ortiz R (1998) Potato breeding via ploidy manipulations. Plant Breed Rev 16:15–85
Paget M, Amoros W, Salas E, Eyzaguirre R, Alspach P, Apiolaza L, Noble A, Bonierbale M (2014) Genetic evaluation of micronutrient traits in diploid potato from a base population of Andean Landrace Cultivars. Crop Sci 54:1949–1959
Phumichai C, Hosaka K (2006) Cryptic improvement for fertility by continuous selfing of diploid potatoes using Sli gene. Euphytica 149:251–258. https://doi.org/10.1007/s10681-005-9072-5
Piepho H-P, Möhring J, Melchinger AE, Büchse A (2008) Blup for phenotypic selection in plant breeding. Euphytica 161:209–222
Plaisted RL (1987) Advances and limitations in the utilization of Neotuberosum in potato breeding. In: Jellis GJ, Richardson DE (eds) The production of new potato varieties. Cambridge University Press, Cambridge, pp 186–196
Plaisted RL, Sanford L, Federer WT, Kehr AE, Peterson LC (1962) Specific and general combining ability for yield in potatoes. Am Potato J 39:185–197
Pushkarnath P (1942) Studies on sterility in potatoes. 1. The genetics of self- and cross-incompatibilities. Indian J Genet and Pl Br 2:11–36
Radley RW, Taha MA, Bremner PM (1961) Tuber bulking in the potato crop. Nature 191:782–783
Ragot M, Bonierbale M, Weltzien E (2018) From market demand to breeding decisions—a framework. RTB Working Paper.
Rodriguez Junior FA, Ortiz-Monasterio I, Zarco-Tejada PJ, Ammar K, Gérard B (2014) Using precision agriculture and remote sensing techniques to improve genotype selection in a breeding program. Conference Paper.International Conference on Precision Agriculture. http://quantalab.ias.csic.es/pdf/Rodrigues%20Jr.%20et%20al.%202014.pdf
Salas E (2002) Estudio de la variabilidad genética de la resistencia al Virus del Enrollamiento de la papa (PLRV) en una población tetraploide. Tesis para optar el Grado de Magister Scientiae. Universidad Nacional Agraria—La Molina. Lima, Peru
Sanford JC, Hanneman RE (1982) Intermating of potato haploids and spontaneous sexual polyploidization—effects on heterozygosity. Am J Potato Res 59:407–414
Sattarzadeh A, Achenback U, Lubeck J, Strahwald J, Tacke E, Hofferbvert H, Rothsteyn T, Gebhardt C (2006) Single nucleotide polymorphism (SNP) genotyping as basis for developing a PCR-based marker highly diagnostic for potato varieties with high resistance to Globodera pallida pathotype Pa2/3. Mol Breed 18:301–312
Slater AT, Wilson GM, Cogan NO, Forster JW, Hayes BJ (2014) Improving the analysis of low heritability complex traits for enhanced genetic gain in potato. Theor Appl Genet 127(4):809–820
Spooner DM, Ghislain M, Simon R, Jansky SH, Gavrilenko T (2014) Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot Rev 80:283–383. https://doi.org/10.1007/s12229-014-9146. —Toward a Rational Classification of Cultivated Plants—Jstor https://www.jstor.org/stable/1218252
Sprague GF, Tatum A (1941) General vs. specific combining ability in single crosses of corn. J Am Soc Agron 34:923–932
Tripp R, Louwaars NP, Van Der Burg WJ, Virk DS, Witcombe JR (1997) Alternatives for Seed regulatory reform: an analysis of variety testing, variety regulation and seed quality control, Agricultural Research and Extension Network Paper, no. 69. http://hdl.handle.net/10535/4581
Ugent DL, Pozorski S, Pozorski T (1982) Archaeological potato tuber remains from the Casma Valley of Peru. Econ Bot 36:182–192
Ugent DL, Dillehay T, Ramirez C (1987) Potato remains from a Late Pleistocene settlement in southcentral Chile. Econ Bot 41:17–27
Uitdewilligen JGAML, Wolters A-MA, D’hoop BB, Borm TJA, Visser RGF, van Eck HJ (2013) A next-generation sequencing method for genotyping-by-sequencing of highly heterozygous autotetraploid potato. PLoS One 8(5):e62355. https://doi.org/10.1371/journal.pone.0062355
Velásquez AC, Mihovilovich E, Bonierbale M (2007) Genetic characterization and mapping of major gene resistance to potato leafroll virus in Solanum tuberosum ssp. Andigena. Theor Appl Genet 114(6):1051–1058
De Vries MI, ter Maat M, Pim Lindhout P (2016) The potential of hybrid potato for East-Africa. Open Agric. 1: 151–156
Wricke G, Weber WE (1986) Quantitative genetics and selection in plant breeding. de Gruyter, Berlin
Xu X et al (2011) Genome sequence and analysis of the tuber crop potato. Nature 475(7355):189–195
Yuen J, Forbes G (2009) Estimating the level of susceptibility to Phytophthora infestans in potato genotypes. The American Phytopathological Society. Phytopathology 99(6):2
Zhang LH, Mojtahedi H, Kuang H, Baker B, Brown CR (2007) Marker-assisted selection of Columbia root-knot nematode resistance introgressed from Solanum bulbocastanum. Crop Sci 47:2021–2026
Acknowledgements
We gratefully acknowledge Elisa Mihovilovich and Thiago Mendes for valuable discussions toward this chapter.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2020 The Author(s)
About this chapter
Cite this chapter
Bonierbale, M.W., Amoros, W.R., Salas, E., de Jong, W. (2020). Potato Breeding. In: Campos, H., Ortiz, O. (eds) The Potato Crop. Springer, Cham. https://doi.org/10.1007/978-3-030-28683-5_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-28683-5_6
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-28682-8
Online ISBN: 978-3-030-28683-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)