
Abstract
Understanding the interactions between the structures and functions underlying regime shifts in dryland social-ecological systems (SESs) and how they respond to climate change is critical for predicting and managing the future of these ecosystems. Due to the high spatiotemporal variability and sensitivity of drylands ecosystem to natural and anthropogenic disturbances, it is challenging to predict the state shifts of dryland SESs. This theme delves into the mechanisms and geographical heterogeneity of resilience and the maintenance of the stability of dryland SESs that involve threshold behaviors. We emphasized the importance of considering both biotic and abiotic factors to identify the factors that drive the evolution of ecosystem structures and functions in drylands. The research frontier involves understanding how ecohydrological and socioeconomic processes drive the evolution of dryland SESs in a geographically diverse and scale-dependent context, developing comprehensive indicators, models, and multivariable approaches, and the development of effective management strategies that can maintain the sustainability of dryland SESs in the face of ongoing global environmental changes.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The structure and function of dryland ecosystems, as well as their interactions with the social and economic systems of the inhabitants, are key determinants of the stability, resilience and sustainability of dryland social-ecological systems (SESs) (Fig. 3.1). However, the dryland ecosystems are highly heterogenous in space and time and are subjected to a range of natural and anthropogenic disturbances that can pushing ecosystems to tipping points and even cause regime shifts. Understanding the biotic and abiotic mechanisms underlying the structure and function of dryland ecosystems is essential for promoting sustainable development, enhancing the delivery of ecosystem services, and building resilience to global environmental change. In this chapter, we explore the structure, functions, and interactions of dryland ecosystems, as well as the challenges and opportunities for their management and conservation.

The interrelations among structure, function, and stability in dryland ecosystems
Drylands are characterized by a patterned ecosystem structure, spatial heterogeneity in functional attributes, and extensive structure–function interactions (Berdugo et al. 2020; Buxton et al. 2022; Maestre et al. 2021; Mayor et al. 2019; Meloni et al. 2019; Saco et al. 2018; Stavi et al. 2021). In drylands, the extreme climatic conditions and rapid ecosystem processes make the ecosystem highly sensitive to drivers. Both ecosystem structure and function are prone to fluctuations and changes (D’Odorico et al. 2013; Bestelmeyer et al. 2015). Globally, aridification caused by climate change has significantly affected the structure and function of dryland ecosystems, resulting in systemic and abrupt changes in multiple ecosystem attributes (Berdugo et al. 2020; Maestre et al. 2021). As drylands continue to expand, more than 20% of the terrestrial surface projected to cross one or several of these thresholds by 2100 (Berdugo et al. 2020). Understanding the spatial heterogeneity of vegetation assemblages and biodiversity along environmental gradients, particularly those of water availability and aridity, is essential. Moreover, the spatial organization and evolutionary trajectory of natural dryland ecosystems along environmental gradients must be addressed (Berdugo et al. 2020). Given the importance of dryland ecosystems for human well-being, there is growing interest in understanding their structure, functioning, and interactions with human societies, as well as in developing strategies to conserve and sustainably manage these ecosystems.
2 Dryland Ecosystem Structure and Functions
2.1 Dryland Ecosystem Structure
Dryland ecosystem structure refers to the abiotic and biotic structures and their spatial patterns in arid and semiarid regions. It includes physical and chemical components of the environment, such as spatially heterogenous soil type, topography, and climate, as well as varying biotic components, such as vegetation, animals, and microorganisms (Schowalter 2011). Dryland ecosystems habitat various organisms, including mammals, birds, reptiles, and insects etc. They play important roles in nutrient cycling, pollination, seed dispersal, and other ecosystem processes. The soil in dryland ecosystems is typically low in organic matter and nutrients and can be prone to erosion and desertification. The physical and chemical properties of the soil greatly influence plant growth and ecosystem functioning. Dryland ecosystems are characterized by a variety of plant communities adapt to the unique climatic and environmental conditions of the area. The diversity and composition of vegetation can significantly affect ecosystem functions, such as nutrient cycling, carbon sequestration, and soil quality. Water is a limiting resource in dryland ecosystems. Its availability and distribution greatly influences the structure and function of ecosystems (Hoover et al. 2021; Zhou et al. 2021). Some dryland ecosystems rely on seasonal precipitation and intermittent water sources.
The structure of dryland ecosystems fluctuates or change rapidly (D’Odorico et al. 2013; Bestelmeyer et al. 2015). Dryland ecosystems have evolved over millions of years in response to various environmental factors including climate, topography, soil characteristics, and biotic interactions (García-Palacios et al. 2018; Bestelmeyer et al. 2015). There is great spatial heterogeneity and large variety of plant and animal communities coexist in close proximity. This heterogeneity is driven by the patchy distribution of resources, such as water and nutrients, and the variable climatic conditions that shape the landscape. Consequently, dryland ecosystems feature with a high degree of adaptability and resilience (Maestre et al. 2011). Organisms inhabiting these ecosystems have evolved various strategies to cope with drought, heat, and other environmental stressors. For example, many desert plants have deep roots that allow them to tap into water stored deep in the soil, whereas others have evolved mechanisms to store water within their tissues (Liu et al. 2013).
2.2 Dryland Ecosystem Functions
Sustaining the productivity, soil erosion control and mitigating water, carbon and nutrient cycling are important functions of dryland ecosystems. They support the livelihoods of millions of people who depend on them for food, fiber, and fuel. Dryland ecosystem functions are highly interconnected, and changes in one function can have a cascading effect on others. Changes in vegetation cover can affect the water balance of the system, redistribution of materials, which in turn affects ecosystem productivity and nutrient cycling (Turnbull et al. 2012; Maestre et al. 2021; Mayor et al. 2019). Similarly, changes in nutrient availability can affect the diversity and productivity of vegetation, and affects carbon sequestration and water balance.
Functioning of dryland ecosystems is highly sensitive to environmental changes (Tietjen et al. 2010; Maestre et al. 2021; Moreno-Jiménez et al. 2019). Climate change and land-use change alters the ecosystem processes and finally the ecosystem services (Smiraglia et al. 2016). Natural and anthropogenic disturbances (Eldridge and Greene 1994, Bochet et al. 2021) have important role in shaping the dynamic of dryland ecosystem functions (Zika and Erb 2009; Abel et al. 2021). For instance, water and nutrient cycling processes in drylands are highly responsive to environmental changes. Changes in precipitation patterns and temperature regimes affect the productivity and composition of vegetation, which in turn affect carbon and nutrient cycling. Disturbance events such as fires have long-lasting impacts on carbon and nutrient cycling through altering vegetation cover and soil properties. Nitrogen is often a limiting nutrient in drylands. Nitrogen fixation by biological soil crusts has been found to be a crucial process in maintaining the nitrogen balance of dryland ecosystems (Belnap and Lange 2003). Furthermore, inputs of anthropogenic nitrogen from sources such as fertilizer application and atmospheric deposition can have significant impacts on the functioning of dryland ecosystems (Yahdjian et al. 2011). Grazing is fundamental livelihood for residents in dryland over the world. It mitigates dryland ecosystem structure and functions in various avenues (Eldridge and Greene 1994). It affects vegetation cover, species composition, and nitrogen fixation of biological soil crust in dryland steppe (Liu et al. 2009). Overgrazing can led to the reduction of hydrological function (Vandandorj et al. 2017), enhances soil loss and plant invasion (Belnap et al. 2009), which negatively impact the water cycling and productivity of dryland ecosystems at various scales. Understanding the functioning of dryland ecosystems and how they are affected by environmental change is critical for sustainable management. Further researches is needed to address the remaining knowledge gaps in dryland ecosystem functioning and develop effective management strategies that balance conservation and development goals in dryland regions.
2.3 Structure-Functions Interactions in Dryland Ecosystems
Understanding the structure–function interactions in dryland ecosystems is fundamental for enhancing ecosystem services supply and reducing the negative impact of environmental change. There is necessity to incorporate structure–function interactions into dryland ecosystem management strategies and call for future research to explore the potential trade-offs among management goals. Ecosystem structure is influenced by ecosystem functions, and can be defined as the minimal or parsimonious pattern of organization required for a function to operate (Müller 1997). Understanding the interactions between ecosystem structure and function at multiple spatial scales has significantly increased our understanding of how terrestrial ecosystem functions respond to global environmental change (Maestre et al. 2021; Turnbull and Wainwright 2019). Among the structure–function interactions, interaction between biodiversity and ecological functions is a critical question in ecology (Peterson et al. 1998). Biodiversity, particularly species richness, has been shown to positively affect ecosystem functions at all spatial scales and trophic levels (Maestre et al. 2021). Biodiversity has a significant impact on ecosystem functions at different hierarchical scales. Therefore, it is crucial to consider biodiversity in ecosystem management (Oliver et al. 2015). A focus on resilience rather than the short-term ecosystem functions and delivery of services, along with the consideration of specific underlying mechanisms, can bridge the research areas of biodiversity-ecosystem function and ecological resilience. This can ultimately aid the development of evidence-based yet flexible ecosystem management strategies. Perennial plant species richness has been found to be crucial for ecosystem function in dryland systems (Midgley 2012), which has global relevance, particularly for developing and least-developed countries facing desertification trends. However, a comprehensive understanding of how biodiversity across trophic levels, in conjunction with abiotic drivers, determines ecosystem function is still needed. For instance, earlier experimental studies and approaches undertaken in temperate grasslands have yet to address this multilayered question.
In dryland landscape, surface cover pattern is of critical importance for the structure–function interaction. The vegetation structure participates in regulating the water balance of ecosystems by mediating the interception and infiltration of rainfall as well as the redistribution of water within the ecosystem (Doerr et al. 2000). Besides, soil structure, which regulates nutrient cycling and carbon sequestration in dryland ecosystems (Lal 2015) is important in sustainable management strategies. Soil organic carbon accumulation in drylands is influenced by a range of factors including climate, vegetation cover, and soil texture (Lal 2004). Restoring the vegetation cover in degraded drylands can improve carbon and nutrient cycling, increase soil water retention, and prevent soil erosion. The conservation of biological soil crusts also help maintain soil stability (Guo et al. 2008; Rodriguez-Caballero et al. 2013) and nutrient cycling (Belnap et al. 2016)in drylands.
The ecosystem structure of drylands interacts with functions through multiple feedbacks, particularly hydrological feedback (D’Odorico and Bhattachan, 2012). As showed by Fu et al. (2021) in Mediterranean drylands, the land-use changes have significant impacts on soil carbon and microbial diversity, which have implications for ecosystem functions, such as nutrient cycling and water regulation. It was illustrated that the composition and diversity of plant communities led to changes in ecosystem functions such as carbon and nutrient cycling, which reshapes soil properties, such as organic matter content and soil texture that affect plant growth and nutrient availability, and finally the ecosystem productivity and resilience to drought (Maestre et al. 2012).
3 Structure-Function Interactions Driven by Global Change
3.1 Structure–Function Interactions Along Aridity Gradients
Complex relationships exist between vegetation structure, ecosystem functioning, and environmental factors along aridity gradients (Maestre et al. 2015; Hu et al. 2021; Migliavacca et al. 2021). Studies focusing on the vegetation response to climate change and the identification of the aridity threshold have highlighted the importance of monitoring subtype dynamics and transition zones for predicting and mitigating the impacts of climate change on dryland ecosystems. For example, the relationship between vegetation cover and primary productivity is nonlinear and varies depending on the amount and timing of rainfall (Zhao et al. 2021). A aridity threshold of 0.54 has been identified as a critical point beyond which plant productivity undergoes abrupt changes, leading to systematic changes (Berdugo et al. 2020). Aridity has been found to reduce vegetation growth in drylands worldwide, although patterns vary among ecosystems and climate zones (Xu et al. 2018). Regional differences in ecosystem responses to climate change can be attributed to the varied responses of drylands to aridity gradients (Huang et al. 2016). The evolutionary trajectory of vegetation activity in response to variations in aridity in drylands plays an important role in predicting future ecosystem functions under global climate change. The expansion of drylands has negative consequences on carbon sequestration and increases the risk of ecosystem degradation (Huang et al. 2016). However, the response of vegetation to changes in aridity can vary across different subtypes and is regulated by the structure–function interactions within ecosystems. Abrupt changes in each subtype may result in a shift in drylands (Zhao et al. 2021). Drying and wetting trends are common climatic phenomena observed over long time (Zaitchik et al. 2023; Sheffield et al. 2012; He et al. 2019), and have a significant impact on plant growth and vegetation cover in drylands worldwide (Feng and Fu 2013; Huang et al. 2016). Small changes in the aridity may have a significant impact on the vegetation status because the response of vegetation to wetting or drying differ greatly in dryland ecosystems. However, the heterogeneity in the magnitudes and areas of drying and wetting trends may cause misinterpretation of the terrestrial and atmospheric feedbacks, leading to an inaccurate assessment of the “dry gets drier and wet gets wetter” paradigm (Greve et al. 2014; Huang et al. 2016).
The trajectory of vegetation trends in response to temporal changes in aridity was highly dependent on the spatial aridity gradient. Changes in spatial aridity can result in distinct vegetation responses to drying and wetting owing to differences in the capacity for resistance to drought among different subtypes (Xu et al. 2018). Resistance capacity is highly dependent on spatial aridity, with increased spatial aridity index (AI) improving the ability of plants to withstand drought. In drylands, vegetation activity is constrained by water availability (Zhao et al. 2020) and is positively correlated with AI (Huang et al. 2016). Normalized difference vegetation index (NDVI) trends shows regional differentiation along the aridity gradient, with lower vegetation cover and productivity in drier regions, resulting in lower NDVI trends than those in wetter regions (Xu et al. 2019).
Box 3.1 Soil nutrient changes in the northern dryland of China—a long-term observation of the Chinese Ecosystem Research Network (CERN)
Soil comprises the largest carbon and nitrogen pool in terrestrial ecosystems, particularly in drylands dominated by grasslands and deserts. More than 90% of carbon and nitrogen are stored in the soil, making it an essential indicator of ecosystem health (Sharrow and Ismail 2004). The balance of soil nutrient elements is reflected in the soil carbon and nitrogen content, and their ecological stoichiometric characteristics, which are primarily influenced by regional water and heat conditions. The total amount of soil carbon and nitrogen varies greatly due to soil formation factors and human activities, resulting in varying soil carbon–nitrogen ratios across temporal and spatial scales (Wang and Yu 2008). The soil carbon–nitrogen ratio serves as an effective method for evaluating soil quality (Walker and Adam 1958) and ecosystem health (Yang et al. 2017), representing the integration of ecosystem function variability. Therefore, long-term field observations of soil nutrient content are of significant importance for understanding the carbon sink potential of the ecosystem and predicting its response to future climate change.
Long-term monitoring data from the Chinese Ecosystem Research Network (CERN) stations have revealed the spatial patterns and temporal evolution of soil carbon and nitrogen characteristics from 2004 to 2018, including their response to climate change (temperature and precipitation). The content of soil organic carbon and total nitrogen in China's dryland ecosystems exhibited strong spatial differentiation, with an increasing trend from west to east, but the carbon–nitrogen ratio remained relatively stable. Between 2004 and 2018, the carbon and nitrogen content in semi-arid drylands showed an increasing trend, leading to improved soil quality, whereas arid drylands did not experience significant changes. Carbon–nitrogen content and ratio were significantly positively correlated with precipitation and negatively correlated with temperature, indicating the significant influence of hydrothermal control. By continuously monitoring soil carbon and nitrogen characteristics and their temporal and spatial changes in climate sensitivity, it is possible to provide a scientific basis for accurately assessing and predicting soil quality and ecosystem health.

Carbon–nitrogen characteristics of soil in ecological stations in drylands. CLD, Cele Desert station; LZD, Linze Desert station; ESD, Erdos Desert station; NMD, Naiman Desert station

Variation of soil organic carbon content, total nitrogen content, and carbon–nitrogen ratio (dash lines represents the regression with significant trends at 0.05 level); relationship of soil organic carbon, nitrogen, and carbon–nitrogen ratio with climate
The results demonstrate that soil carbon and total nitrogen content increase from west to east with decreasing aridity (Figs. 3.2 and 3.3). From 2004 to 2018, soil carbon significantly increased in semi-arid drylands, including ESD and NMD, while no significant variation was observed in arid drylands, including CLD and LZD. This implies a carbon sequestration potential in semi-arid drylands under climate change. However, the variation in soil nitrogen content did not present an obvious pattern along the aridity gradient, suggesting a potential risk of the impact of soil nitrogen limitation on ecosystem structure and function change. Soil carbon and nitrogen content are significantly controlled by hydrothermal conditions, showing positive and negative correlations with precipitation and temperature, respectively. Future research should explore the impact of the interaction of precipitation and temperature on soil carbon and nitrogen content and the differences in temporo-spatial patterns along the aridity gradient. Furthermore, studying soil-vegetation and soil-microbe activities is necessary to explore the climate-carbon feedback under global change. Investigating the biochemical cycle of the soil–vegetation–atmosphere continuum and exploring the evolution characteristics of the ecosystem and its response to climate change is essential to increase the terrestrial ecosystem carbon sink and ecological health.
3.2 Responses and Feedback of Dryland Ecosystem to Climate Change
Recent climate change research largely confirmed the impacts on ecosystems and provides a greater mechanistic understanding and geographic specificity for those impacts (Grimm et al. 2013). Pervasive climate change impacts on ecosystems affect their productivity or ability to process chemical elements. The combined impacts of wildfires and insect outbreaks decrease forest productivity, mostly in the arid and semi-arid West. Forests in wetter regions are more productive owing to warming. Shifts in species ranges are so extensive that by 2100, they may alter the biome composition across 5–20% of the US land area. The accelerated loss of nutrients from terrestrial ecosystems to receiving waters is caused by winter warming and intensification of the hydrological cycle. Ecosystem feedback, especially that associated with the release of carbon dioxide and methane from wetlands and the thawing of permafrost soils, magnifies the rate of climate change (Grimm et al. 2013).
Global environmental changes rapidly alter the structure–function relationships in ecosystems. Changes in precipitation patterns, nutrient inputs and losses, plant photosynthesis rates, and extreme climatic events reshape vegetation cover patterns and can cause unexpected, abrupt, or catastrophic shifts in ecosystems, resulting in the loss or gain of ecological resources (García-Palacios et al. 2018; Mayor et al. 2019; Ursino 2019). Alterations in precipitation patterns can lead to changes in soil moisture and nutrient availability, which can affect microbial activity and nutrient cycling (Huang et al. 2015). These changes can affect primary productivity, carbon storage, and soil fertility (Jobbágy and Jackson, 2000; Schlesinger et al. 1990). Despite the high vulnerability, sensitivity, and fluctuations of dryland ecosystems, there is a lack of verification and predictive power for catastrophic responses to changing environmental conditions, particularly in large-scale ecosystems (Rietkerk et al. 2004; Ursino 2019).
Climate change has been identified as a significant driver of vegetation activity, with reciprocal impacts on land surface cover changes (Schimel et al. 2000; Nemani et al. 2003). The effects of climate change include both interannual and long-term trends that have substantial impacts on vegetation growth (Ryo et al. 2019). In water-limited ecosystems, increased precipitation can lead to enhanced vegetation growth, which, in turn, creates biophysical feedback to the climate system, resulting in increased evapotranspiration, surface cooling, and precipitation (Davin and de Noblet Ducoudre 2010; Yu et al. 2020). In the Patagonian rangelands, along with climate, vegetation structure is also an essential factor in shaping ecosystem functioning (Gaitán et al. 2015). Maintaining and enhancing vegetation cover, particularly grasses, is crucial for mitigating the adverse effects of climate change on ecosystem functioning.
In recent decades, global greening and deforestation have caused significant changes in vegetation dynamics with implications for climate feedback mechanisms (Piao et al. 2015; Chen et al. 2019a; 2019b; Davin and de Noblet-Ducoudre 2010; Strassburg et al. 2012; Seymour and Harris 2019). As global warming surpasses the optimal temperature for vegetation growth, there is a limited safe operating space for these ecosystems (Huang et al. 2019; Xu et al. 2013). Furthermore, dominant climatic factors vary spatially and temporally, leading to potential differences in vegetation activity with climate change and spatial variations. To develop better predictive models for the trajectory of dryland ecosystems under future climate scenarios, research efforts must focus on improving our understanding of these variations (García-Ruiz et al. 2011). Cold and high-latitude ecosystems have experienced a 16.4% decline in vegetated land area limited by temperature owing to rapid warming (Keenan and Riley 2018). In Australia, the precipitation threshold for water limitation of vegetation cover declined significantly from 1982 to 2010 as the vegetation adapted (Ukkola et al. 2015). Deforestation results in surface warming, whereas surface greening leads to cooling effects (Davin and de Noblet-Ducoudre 2010; Yu et al. 2020). Reforestation in Europe has resulted in divergent responses to summer temperature changes (Davin et al. 2020).
Box 3.2 Climate change and vegetation dynamics in the Euro-Asia Transect
The Euro-Asia Transect (EAT) is a crucial component of regional ecological security and the global carbon cycle. Encompasses the Mongolia Plateau, Loess Plateau, Central Asia, and Mediterranean (Fig. 3.4). The majority of the EAT is dryland and has a delicate ecosystem consisting of dry and desert grasslands that are sensitive to climate change. Vegetation cover is critical for preserving environmental stability, providing food and livelihood security, and improving soil quality, among other benefits (Ravi et al. 2010; Xu et al. 2017). However, global warming and extreme climate events have increased the vulnerability of vegetation activity and prompted the degradation of the fragile ecosystems, negatively impacting the sustainability and human well-being in the EAT. Therefore, investigating the contribution of climate change, particularly the interaction of precipitation with temperature on vegetation dynamics, is essential for ecosystem management and predicting the future of global drylands.

Location of the Euro-Asia transect drylands. Drylands are divided by aridity index into four subregions: hyperarid, arid, semiarid, and dry subhumid
Contribution of climate change to the variability of Normalized Differential Vegetation Index as the proxy of vegetation dynamics in the Euro-Asia Transact (EAT) are quantified using the GeoDetector Model (Fig. 3.5). Results demonstrated that precipitation (PRE) contributed much more than temperature (TEM) to the EAT vegetation dynamics, and the interaction of PRE with TEM had an enhanced positive effect on the NDVI variability compared to the sum of PRE and TEM contribution. Regions with PRE contribution below 10% account for more than half (53%) of the EAT dryland, and that between 10 and 30% account for 40% of the studied area. Patterns of TEM contribution is similar to that of PRE, regions below 30% account for about 98% of the EAT dryland. Importantly, the interaction of PRE with TEM exerted a significant breakthrough impact on the vegetation index variability. Regions of the interaction contribution of PRE with TEM that are above 30% account for 46% of the EAT. Moreover, those that are between 10 and 30% account for 51% of the area. Furthermore, 86% of the EAT dryland showed the enhanced positive effect of the interaction of PPT with TEM which is even larger than that of the sum of PRE and TEM contribution, remarkably improving the interpretation of climate change impacts on vegetation dynamics.

Contribution of climate change to vegetation dynamics in the Euro-Asia transact drylands. a Contribution of PRE to NDVI variability; b Contribution of TEM to NDVI variability; c Contribution of the interaction between PRE and TEM to NDVI variability; d Difference between contribution of PRE-TEM interaction and the sum of PRE and TEM to NDVI variability
Recent studies reveal that the present ‘hot model’ is poor in performance in climate simulations and exaggerates the earth and ecosystem impacts (Hausfather et al. 2022; Voosen 2022). The EAT dryland occupies a large area as a widely inland ecosystem. It is substantially relevant because of the biochemical cycles and geo-physical feedback between the terrestrial and atmospheric domains through carbon-climate responses. Quantifying the contribution or the association between climate change and vegetation dynamics is important to understand the mechanism of the terrestrial carbon sink effect through plant physio-ecological process and surface physical-feedback. Multi-site comprehensive and comparative studies based on long-term field monitoring would provide confident parameter modules for the ecological processes and earth models to improve the accuracy of carbon-climate interaction simulations and predictions.
Anthropogenic disturbances, such as land-use change, overgrazing, and desertification, affect the impacts of climate change on drylands, highlighting the need for improved monitoring and modeling tools to accurately capture ecosystem dynamics in response to climate change (Verwijmeren et al. 2013; He et al. 2011; Richardson et al. 2019). Understanding the complex interplay between the structure and function of drylands, as well as their responses to global changes, is critical for the sustainable management and conservation of dryland ecosystems. Interdisciplinary research and multiscale approaches along with the integration of new technologies and methods are necessary to achieve this goal.
3.3 The Geographical Diversity of the EvolutionTrajectory of Dryland
The trajectory of dryland evolution is influenced by various geographic factors, including climate, topography, soil characteristics, and biotic interactions. These factors vary widely across regions, resulting in diverse patterns of vegetation cover, nutrient cycling, and ecosystem functions (Maestre et al. 2021). In addition, the interactions between vegetation and soil properties can influence the ability of the ecosystem to resist and recover from disturbances, such as drought, fire, and grazing. Furthermore, the effects of climate change on dryland ecosystems can vary across spatial scales, from individual plant responses to regional changes in precipitation patterns (Schlaepfer et al. 2017). This variability can also be influenced by interactions between different components of the ecosystem, such as plant–herbivore interactions, feedback between vegetation and soil processes, and the effects of climate on biotic communities (Schwinning and Sala 2004). Because of the effects of climate change on drylands vary across biophysical and socioeconomic contexts (Doerr et al. 2000; Maestre et al. 2016), as well as the specific adaptive capacity and resilience of each system (Bardgett et al. 2021), the geographically diverse structures and functions of different drylands bring high uncertainty in the trajectory of local dryland changes under climate change (Berdugo et al. 2022).
The geographically diverse human activities also participated in making the diverse trajectory of dryland ecosystem dynamic. Overgrazing, land-use change, and drought, which are geographically diverse, can also affect the trajectory of dryland evolution (Belnap et al. 2009, Eldridge and Greene 1994). Grazing can lead to soil erosion, vegetation loss, and changes in nutrient cycling, contributs to desertification and land degradation (Osem et al. 2013). However, well-managed grazing systems that take into account the ecological processes of dryland ecosystems can positively affect biodiversity, soil health, and carbon sequestration (Teague et al. 2016). Agriculture can also have significant impacts on dryland ecosystems, particularly through the use of irrigation, which can lead to soil salinization and waterlogging, and the use of agrochemicals, which can negatively impact soil health and biodiversity (Lal 2015; Cherlet et al. 2018). However, sustainable agricultural practices such as conservation agriculture and agroforestry can contribute to the restoration of dryland ecosystems by enhancing soil health, biodiversity, and carbon sequestration (González-Sánchez et al. 2016; Mbow et al. 2014). Urbanization can also have significant impacts on dryland ecosystems, including habitat loss, fragmentation, and changes in microclimate (Grimm et al. 2008; Seto et al. 2012). Green infrastructure, such as parks and urban forests, can contribute to the restoration of dryland ecosystems by providing habitats for wildlife, improving air and water quality, and reducing the urban heat island effect (Nowak et al. 2006).
Predicting the trajectory of local dryland ecosystem changes under climate change can be challenging because of the high uncertainty associated with the interactions among these diverse factors (Verwijmeren et al. 2013). Since the trajectory of dryland evolution is complex and influenced by multiple factors, including climate change, land-use changes, and social and cultural dynamics, the effective management and restoration of dryland ecosystems require interdisciplinary and integrated approaches that incorporate the ecological, social, and economic aspects of these systems. By combining scientific knowledge with local knowledge and engaging local communities and stakeholders, sustainable and effective strategies for the conservation and restoration of dryland ecosystems can be developed.
4 Stability and Resilience of Dryland Ecosystem and Implications for Restoration
4.1 Ecosystem Stability and Resilience in Drylands
Ecosystem stability and resilience are intrinsically determined by the interactions between ecosystem structure and function (D’Odorico et al. 2013; Maestre et al. 2021). The presence of alternative stable states can have unexpected and serious consequences due to anthropogenic environmental changes and natural perturbations (Schröder et al. 2005). Dryland ecosystems exhibit slow changes in structure but rapid responses to external drivers and feedback (Bestelmeyer et al. 2015; Saco et al. 2018). The critical role of ecohydrological feedback in driving state changes in dryland ecosystems has been previously highlighted (Saco et al. 2018). Multiple types of feedbacks, particularly ecohydrological feedback, couple dryland ecosystem structures with their functions (D’Odorico et al. 2013). The relationships between structure and function at various spatial scales are indicative of how terrestrial ecosystems respond to global change (Maestre et al. 2021) and are intrinsic determinants of ecosystem state change (Mayor et al. 2013; Saco et al. 2018). Descriptions of ecosystem stability depend on the scale of observation, including physical and temporal scales and subjectively chosen indicators. Resilience refers to the ability of an ecosystem to recover from disturbances and maintain its functions over time (Folke et al. 2004). It describes the ability of an ecosystem to resist changes in response to environmental stressors (Holling 1973). Descriptions of ecosystem stability also depend on the scale of observations, with different indicators and processes becoming relevant at different spatial and temporal scales (Huston 2014). Aridity gradients have a significant effect on ecosystem stability (Maestre et al. 2012). Dryland ecosystems may experience frequent disturbances, such as droughts and fires, which can disrupt their functions and reduce their stability (Reynolds et al. 2007). However, these ecosystems may also exhibit unique adaptations to water scarcity, such as efficient water use and high carbon storage in soils (Maestre et al. 2021). The resilience to different magnitudes of drought and different ecosystems to environmental stressors should be addressed. Comprehensive indicators that cover both “slow” and “fast” variables are needed for describing the resilience of dryland ecosystems. Spatio-temporal variations in resilience, coupled with water input pulses, should be included.
Structure–function interaction plays fundamental role in determining the stability and resilience of the dryland ecosystems. The physical structure of dryland ecosystems, including soil composition, vegetation cover, and hydrological patterns, plays a crucial role in determining ecosystem functions, such as carbon sequestration, nutrient cycling, and water retention. These ecological functions affect the structure of the ecosystem, creating a feedback loop between structure and function. One of the central questions regarding the interactions between ecosystem structure and function is how biodiversity relates to ecological functions (Peterson et al. 1998). Increasingly diverse ecosystems have a greater probability of including species with disproportionate positive or negative effects on ecosystem functioning (Naeem et al. 2009). Mounting evidence indicates that biodiversity enhances multifunctionality (Chen et al. 2018; D’Odorico et al. 2013; Maestre et al. 2021) and increases the stability and resilience of ecosystems in changing environments (Folke et al. 1996). However, ecosystem structure and function in drylands rely heavily on water availability and feedback to water redistribution and cycling by changing the physical environment (Saco et al. 2020; Urgeghe et al. 2010) or the physiological capacity for water transpiration (Fisher et al. 2011). Among all the structural features, the spatial and temporal patterns of vegetation cover in drylands are crucial for ecosystem functioning, and mitigates ecosystem stability and resilience. Restoring vegetation cover in a degraded rangeland ecosystem improved the soil structure and water infiltration, leading to enhanced plant growth and carbon sequestration (Doerr et al. 2000; Valone et al. 2002). Structure–function interactions in a degraded grassland ecosystem showed that increasing plant diversity through the restoration of native plant species enhanced ecosystem functions, such as soil carbon sequestration and nutrient cycling (Zhang et al. 2020). Increasing soil organic matter through the incorporation of cover crops in a degraded dryland ecosystem in the Great Plains of the United States improved the soil structure and water retention, leading to enhanced carbon sequestration and increased resilience to drought (Blanco-Canqui et al. 2020). Thus remote sensing derived information on vegetation cover, biomass, and productivity are important indicators of ecosystem stability (Zhang et al. 2013).
Soil carbon stability and greenness are two key indicators adopted to assess ecosystem stability along the aridity gradient. Soil carbon stability is an important measure of ecosystem health as it reflects the amount of carbon stored in soils over long periods and contributes to global carbon cycling (Köchy et al. 2015). Greenness, on the other hand, is a measure of the amount of vegetation cover and photosynthetic activity in an ecosystem, which can be indicative of its overall productivity and health (Zhu et al. 2015). By examining how these indicators vary across an aridity gradient, we gain a better understanding of the factors that contribute to ecosystem stability and how they respond to environmental change. The diversity and abundance of plant and animal species provide important information regarding the ecosystem’s functions and services (Díaz et al. 2016). Additionally, the functional traits of species, such as their water-use efficiency and tolerance to drought, can help explain the patterns of ecosystem stability along the aridity gradient (Siefert et al. 2015). By examining ecosystem stability along the aridity gradient at different spatial and temporal scales and using a range of indicators, we can gain a comprehensive understanding of how these ecosystems respond to environmental changes and the factors that contribute to their stability.
Dryland ecosystems can exhibit high resilience to climate change. However, this resilience may vary across spatial scales and may be influenced by factors such as land-use change and management practices (Fu et al. 2021). In addition to changes in vegetation, climate change has led to alterations in nutrient cycling, soil moisture, and other ecosystem processes in drylands. These changes have been shown to have cascading effects on entire ecosystems, ranging from microbial communities to larger mammals (Eldridge et al. 2016). Owing to their unique combinations of stressors, regional ecosystems may differ considerably in their normal ranges of primary and secondary productivity, species composition, diversity, and nutrient cycling, making the patterns of their responses to stressors highly variable and unpredictable (Rapport and Whitford 1999). Therefore, understanding the mechanisms and geographical heterogeneity of resilience and sustaining the stability of dryland ecosystems are critical scientific tasks that should be included in dryland ecosystem research.
4.2 Mechanisms of Maintaining Resilience and Stability
Dryland ecosystems are characterized by limited water resources. Thus, ecohydrology plays a crucial role in connecting ecosystem structures and functions. The ecohydrological status of a dryland ecosystem determines its resilience and stability based on water availability (Fig. 3.6). This suggests that, in areas with high water availability, water inputs can be increased through water management or more efficient water use through improved ecosystem structure and function. Because water availability is critical in dryland environments, the relationships between ecosystem structure and multifunctionality in drylands are unique compared to other ecosystems. Interactions relevant to ecohydrological behaviors should be focused on, such as water redistribution among vegetation patches, the effect of vegetation organization on runoff redistribution, and rainfall portioning. The organization of vegetation at different spatial scales influence the redistribution of water and nutrients, ultimately affecting ecosystem productivity and resilience. Temporal variability in water availability can lead to shifts in plant community composition and function, highlighting the need for long-term monitoring and evaluation of ecosystem responses to environmental change in drylands.

Ecohydrology is essential in understanding the interaction among structure, function, and resilience of dryland ecosystems, and their coupling to the social system
Plant traits, such as root length, hydraulic conductivity, and stomatal conductance, strongly influence water-use efficiency and drought tolerance of dryland plant communities (Díaz et al. 2016; Pillar et al. 2018). Dryland ecosystems exhibit complex patterns of connectivity and feedback between vegetation and hydrological processes that have important implications for the resilience and stability of these systems (Feng et al. 2016). Zhou et al. (2016) investigated the impact of vegetation organization on runoff generation in semiarid grasslands, whereas Zhang et al. (2020) examined the effects of shrub introduction on soil properties and implications for revegetation in a dryland.
Restoration activities that focus on enhancing soil health and water storage capacity can also help build resilience to impacts of climate change such as drought and desertification (Hoover et al. 2021). This highlights the importance of understanding the ecohydrological behavior of dryland ecosystems to develop effective management and restoration practices.
Understanding the interactions between the structure and function of dryland ecosystems at multiple spatial scales is essential to manage these ecosystems and mitigating the effects of global environmental change (Maestre et al. 2021; Turnbull and Wainwright 2019). Vegetation plays a critical role in regulating dryland ecosystem processes such as carbon and nutrient cycling, water availability, and soil erosion control. The spatial arrangement of vegetation cover, including the distribution of vegetation patches and gaps, can have a significant impact on ecosystem processes such as water infiltration and runoff (Doerr et al. 2000; Wang et al. 2015). Nitrogen-fixing plants or deep-rooted plants can enhance ecosystem processes and resilience to environmental stressors such as drought or nutrient limitation (Maestre et al. 2021; Ochoa-Hueso et al. 2019). The effectiveness of vegetation in regulating ecosystem processes may depend on the spatial scale of observations and the characteristics of the landscape context (Maestre et al. 2012; Zhang et al. 2016). Vegetation cover can affect soil microbial communities and nutrient cycling, which, in turn, can affect plant growth and productivity (Delgado-Baquerizo et al. 2016). Management strategies promoting the maintenance of healthy and resilient dryland ecosystems should be developed.
4.3 Ecological Restoration of Dryland for SES Sustainability
Dryland SESs are complex systems that involve interactions between natural resources, ecosystems, human activities, and institutions. They are highly sensitive to climate change. Restoration of degraded drylands has gained increasing recognition as a potential approach for enhancing ecosystem resilience and stability. Restoration practices that increase plant functional diversity can enhance ecosystem resistance and resilience to drought, nutrient limitation, and other stressors (Irob et al. 2023; Lepš et al. 2018; Hallett et al. 2017). Restoring soil organic matter can improve soil water-holding capacity and nutrient availability, as well as promote soil aggregation and microbial activity (Zhao et al. 2023). Plant diversity can enhance ecosystem resilience to climate variability and disturbance and increase soil nutrient cycling and carbon storage (Maestre et al. 2012; Soliveres et al. 2014).
Restoration practices that increase plant functional diversity include a range of techniques such as planting diverse mixtures of native species, using seed mixes that incorporate a variety of functional traits, and promoting the natural regeneration of vegetation. In arid and semi-arid ecosystems, restoration projects that incorporate a mix of deep- and shallow-rooted plant species can enhance soil water availability and reduce erosion (Maestre et al. 2021). In degraded grasslands, restoration efforts that promote a diverse mix of grasses, forbs, and legumes can enhance nutrient cycling and improve soil fertility (Hobbs and Norton 1996). Martínez-Vilalta and Lloret (2016) found that the functional diversity of vegetation was positively correlated with the resistance of Mediterranean shrublands to drought and that restoration practices increased functional diversity and improved their resistance. Increased functional diversity also enhance ecosystem resilience by promoting faster recovery after disturbances (Lavorel and Grigulis, 2012). By promoting plant functional diversity, restoration efforts can help build more resilient and productive ecosystems that can better cope with environmental stressors and provide valuable ecosystem services to human communities.
Developing new approaches to evaluate and monitor the success of restoration efforts is critical for promoting effective restoration practices. New approaches for monitoring and evaluating the success of dryland restoration efforts have emerged, including the use of remote sensing and machine learning to assess changes in vegetation cover, soil properties, and other indicators of ecosystem health (Wang et al. 2018). Remote-sensing technologies are used to monitor vegetation dynamics and ecosystem processes at different spatial and temporal scales (Brandt et al. 2018; Wang et al. 2023a). Detecting changes in vegetation cover, biomass, productivity, soil moisture, and other important variables can indicate the effectiveness of restoration efforts. Social-ecological monitoring frameworks have been developed to assess the impact of restoration on both the ecological and social components of dryland ecosystems (Verwijmeren et al. 2013). Such integrated approaches can help ensure that restoration efforts are not only ecologically effective but also socially just and sustainable. These approaches have the potential to provide more accurate and timely information on the effectiveness of restoration practices and support adaptive management and decision-making.
4.4 Ecosystem Management and Structure–Function Inter Action in Drylands
The management of SESs has been scrutinized because of the rising degradation of ecosystems caused by human activities, which poses a threat to human well-being (Clark and Dickson 2003; Folke, 2006; Ostrom 2009). Drylands face the challenge of low water availability, which limits both the material support services of ecosystems and human livelihoods. To address these issues, scientists, governments, and organizations have launched numerous international programs aimed at developing transdisciplinary research-based strategies for problem-solving and SES management (Carpenter et al. 2012). Recent research has shown the potential of nature-based solutions to combat desertification and enhance the resilience of dryland ecosystems to climate change (Bekele et al. 2021; Seddon et al. 2021). These approaches include agroforestry, soil conservation measures, and water-harvesting techniques, which can improve soil quality, enhance biodiversity, and provide multiple benefits to local communities. By combining traditional and modern knowledge systems, context-specific solutions that build on the strengths and resources of local communities can be developed. Other recent studies have highlighted the importance of understanding the socio-ecological dynamics of dryland ecosystems and the potential for collaborative governance approaches to support effective management and restoration efforts (Bawa et al. 2021; Fu et al. 2021; Schlüter et al. 2019). These approaches prioritize the engagement of local communities and stakeholders, the incorporation of multiple knowledge systems, and the building of social networks and partnerships to support ongoing learning and adaptation.
Interactions between various components of dryland SESs can be nonlinear and exhibit thresholds, making it difficult to anticipate abrupt changes and tipping points that have significant consequences for ecosystem services and human well-being (Folke et al. 2010; Scheffer et al. 2012; Allen et al. 2016; Berdugo et al. 2022). Impacts of climate change on dryland SESs differ substantially and depend on factors such as the availability and distribution of water resources, land-use patterns, soil quality, biodiversity, socioeconomic conditions, and governance arrangements, which vary greatly across different dryland regions (Intergovernmental Panel on Climate Change (IPCC) 2019; Millennium Ecosystem Assessment 2005). Furthermore, human activities, such as land-use change and water management, can have significant impacts on the structure and function of dryland ecosystems, further complicating the predictions of their trajectory under climate change (Bastin et al. 2017). Efforts have been made to develop effective management and conservation strategies for the species. For example, global assessments, such as the Intergovernmental Panel on Climate Change (IPCC) and the Global Drylands Initiative (GDI), have identified key drivers of dryland degradation and potential pathways for restoration and adaptation (Intergovernmental Panel on Climate Change (IPCC) 2019; Global Drylands Initiative (GDI) 2021). In addition, advances in remote sensing and modeling techniques have allowed for more accurate mapping and monitoring of dryland ecosystems at regional and global scales (Brandt et al. 2018). The potential feedback between climate change and human activities, such as land-use change and water management, can create additional uncertainties and complexities that are difficult to capture in predictive models (Dearing et al. 2015; Wang et al. 2023a). Emerging technologies such as remote sensing and big data analysis offer new opportunities for monitoring and understanding the dynamics of dryland ecosystems, including the impacts of climate change and human activities (Wang et al. 2023b; Omuto et al. 2010). These technologies can support more accurate and timely monitoring of ecosystem health and inform the design and evaluation of management and restoration strategies.
As explored before, the study of SESs is an expanding scientific field (Colding and Barthel 2019), and it is important to establish a shared analytical framework and consistent standards to compare effective management strategies. An assessment strategy is required to ensure the practical application of effective SES management strategies. However, quantifying the complexity of social, economic, and ecological factors remains challenging due to the lack of long-term social survey data and accurate mathematical models of SESs. In dryland areas, the structure and function of the SESs are closely tied to water availability and are highly vulnerable to natural and human-induced disturbances. There are three associated themes need to be addressed to restore and manage dryland ecosystem for SES sustainability: (i) the spatial and temporal pattern of the evolution of SESs; (ii) the response and feedback of SESs under climate change and the implementation of management strategies; (iii) the assessment of the interaction of natural and societal measurements and their mechanisms. Together, these aim to explore the nexus trajectory of the nature and society coupling mechanism and determine early warning indicators for identifying SES regime shifts (Fu et al. 2021), which will help provide a basis for SES management strategies and human well-being. Further work is also needed to draw on other disciplines to develop appropriate indicators for the simultaneous resilience of multiple ecosystem functions. Efforts towards ecosystem restoration in drylands should strive to increase resistance to environmental stressors while promoting long-term stability. This can be achieved through a combination of measures, such as planting drought-resistant species, improving soil health, reducing erosion, and managing human activities. The successful restoration of dryland ecosystems requires an integrated approach that addresses both the ecological and social dimensions of the system.
Box 3.3 Restoring dryland ecosystems through a social-ecological framework: the smart grassland management system and community-based agriculture reorganization in Inner Mongolia, China
The subregions of global drylands exhibit variations in ecosystem structure and functions, land degradation, and human dependence. Nature-based solutions provide the basic principles for adaptive and sustainable management of Social-Ecological Systems. To cope with future climate change and promote the achievement of the United Nations Sustainable Development Goals, countries worldwide are taking practical actions by adjusting livelihood strategies and ensuring ecosystem stability. In Inner Mongolia, China, activities such as afforestation for carbon sequestration, smart grassland management, and community-based agriculture have been implemented, resulting in significant improvements in carbon sequestration, reduction of land degradation, and enhancement of people’s livelihoods.
In 2010, the Horinger Ecological Restoration Project was launched in Inner Mongolia, China by the Laoniu Foundation and The Nature Conservancy. The project aimed to enhance carbon sequestration capacity and ecosystem services through ecological restoration and livelihood optimization, and to explore trade-offs between natural ecological protection and utilization, and technical engineering for system restoration. The project includes afforestation, gully management, slope restoration, community-based agriculture reorganization, and smart grassland management (Figs. 3.7 and 3.8). The forestry carbon sequestration project in Shengle International Ecological Demonstration Zone in Inner Mongolia was certified with a gold certificate in climate, community, and biodiversity in 2013 under the Clean Development Mechanism (CDM). The carbon storage of this planted forest is expected to reach 220,000 tons of CO2 by 2041, of which 160,000 tons have already been traded and subscribed by Warner Disney.

Restoration of grassland ecosystem structure and function can be achieved through a combination of engineering and biointegration techniques for gully management and smart grassland management. This involves implementing measures such as enclosure and controlled grazing to improve surface vegetation cover and biomass, which in turn helps to maintain the health and stability of the grassland ecosystem

Applying technology to increase and stabilize agricultural yields, and developing feeding and animal husbandry practices to reduce damage to the ecosystem
The project aims to plan ecological restoration, protection, and sustainable development and prioritize protection to comprehensively restore and manage the natural ecosystem and artificial production system for carbon sink enhancement. In conjunction with regional development and the needs of residents, the project promotes systematic ecological restoration and social development cooperation. Activities such as compensation, environmental education, and technical training are carried out to promote scientific farming, develop water-saving techniques, climate-smart agriculture, and explore the tradeoff between food production and economic prosperity in local communities. The project implements precise and intensive management of seasonal grazing and establishes an information platform for comprehensive evaluation of the grassland. Based on the balance of grass and livestock, a seasonal grassland management system is established to determine grazing area and time according to the sustainable utilization potential of the grassland and to promote the balanced development of animal husbandry and grassland production in society.
5 Mechanism for Regime Shifts in Dryland SESs
5.1 Overview of the Regime Shift and Its Impact on SESs
Regime shifts are associated with critical transitions in ecosystem functioning and structure, which can occur in ecosystems due to changes in environmental conditions or management that cross a threshold, leading to large, sudden, and often undesirable changes in the system (Carpenter et al. 1999; Scheffer et al. 2015). According to Scheffer et al. (2001), regime shifts can occur when a complex system is pushed past a tipping point, where the system suddenly reorganizes into a new state with different ecosystem functions and structures. Bestelmeyer et al. (2015) suggested that regime shifts in dryland SESs can occur because of slow or rapid responses to changes in external drivers and feedback within SESs. It is important to address context-specific socio-ecological feedback in drylands that involves threshold behaviors to prevent or mitigate the negative impacts of regime shifts.
Regime shifts can be triggered by a variety of factors such as climate change, land-use change, and changes in biotic interactions (Scheffer et al. 2015). In drylands, regime shifts likely occur due to the vulnerability to land degradation, which can lead to decreased ecosystem resilience and increased susceptibility to droughts and other disturbances (Doerr et al. 2000). Furthermore, dryland SESs are often characterized by complex feedback between social and ecological processes, which can make predicting and managing regime shifts challenging (Folke et al. 2004). Prolonged droughts or changes in precipitation patterns can cause a decline in vegetation cover, leading to soil erosion, increased water runoff, and reduced soil fertility. These changes can create a positive feedback loop in which the loss of vegetation cover leads to further degradation of land and a shift towards a new ecological state with different dynamics and feedback. Socioeconomic factors such as population growth, urbanization, and globalization can also contribute to regime shifts by creating new demands for natural resources and changing the way in which they are managed. By addressing the underlying drivers of regime shifts, such as climate change, land use, and socioeconomic conditions, it may be possible to reduce the risk of ecological degradation and promote more sustainable and resilient dryland ecosystems.
The responses and feedback of SESs to regime shifts are complex and context-specific. In some cases, the negative impacts of regime shifts can trigger further changes in the social and ecological components of a system, leading to a spiraling decline (Cumming et al. 2005). In other cases, regime shifts can lead to positive transformations, such as the emergence of new social norms, institutions, and practices that promote sustainability and resilience (Folke et al. 2010). The dynamics and drivers of regime shifts in SESs need to be paid more attention. Recent studies have highlighted the importance of cross-scale interactions and teleconnections in shaping the vulnerability and resilience of dryland ecosystems (D’Odorico et al. 2013; Wang-Erlandsson et al. 2018). Other studies have emphasized the role of social networks and learning processes in enabling adaptive governance and collective action in the face of regime shifts (Bodin and Prell, 2011; Ernstson et al. 2010). A better understanding of the social dynamics of dryland SESs is necessary for the effective management of regime shifts. Social dynamics, such as resource competition, conflict resolution, and trust among stakeholders, can strongly influence the responses and feedback of SESs to regime shifts (Bowker et al. 2012). Hence, stakeholder engagement and participation in the management of dryland SESs are crucial to ensure the long-term sustainability and resilience of these systems. The response and feedback of SESs to regime shifts requires a comprehensive and multidisciplinary approach to understand and manage effectively.
5.2 Approach and Indicators for Early Warning of Regime Shifts
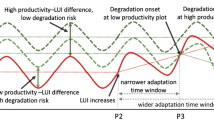
Vulnerable dryland ecosystems readily fluctuate at large magnitude because of their intrinsic biological and physical structures and interactions, as well as the combined impacts of climate change and human activities. Consequently, the structure, functions, and interactions of fragile dryland ecosystems may change significantly (D’Odorico et al. 2013), leading to shifts among alternative stable states. When a critical threshold is surpassed, ecosystems can undergo catastrophic changes and reorganize into different states (Angeler and Allen, 2016; Turnbull and Wainwright, 2019). However, the mechanisms underlying these interactions remain controversial and poorly understood (Loreau and Mazancourt, 2013). Early warning for regime shifts in drylands can help prevent or mitigate the negative impacts of such shifts ecosystems and communities that depend on them.
Early warning of a state shift is still an issue that needs to be addressed. Recently, many early warning signals have been extracted from the spatial patterns of vegetation assembly at the local scale (Berdugo et al. 2017; Bestelmeyer et al. 2013; Nijp et al. 2019; Saco et al. 2020). Single aspects of regime shifts are often addressed because it is easier to handle the relationship between variables (Corrado et al. 2014; Zurlini et al. 2014). However, predictions based on a single indicator cannot describe the whole story of state shifts in dryland ecosystems, owing to great fluctuations, high sensitivity, and vulnerability to natural and/or anthropogenic disturbances. Additionally, increased variance and autocorrelation are potential early warning indicators that are readily used (Kéfi et al. 2014). However, they fail to predict nonlinear changes (Burthe et al. 2016), although nonlinearity (McGuire and McDonnell, 2010; Sarah 2004), feedback (Saco et al. 2018; Turnbull et al. 2012), and behavioral thresholds (Eby et al. 2017; Schwinning et al. 2004; Zehe et al. 2007) are common in dryland ecosystems. These characteristics of dryland ecosystems lead to a low predictability of state shifts caused by changes in ecosystem structure, and the difficulty in determining the tipping point for state shifts does not mention alternative stable states. Thus, understanding how state shifts because of changes in ecosystem structure, and what the tipping point is, requires an understanding of the biotic and abiotic mechanisms underlying state shifts and stability from a more holistic perspective. Furthermore, comprehensive indicators and models should be developed and multivariate approaches are necessary.
To develop effective early warning systems, it is important to consider indicators that can provide insights into the potential for a regime shift to occur. The AI can serve as an important early warning signal of ecosystem shifts in drylands. Vegetation productivity responds to moderate drying and wetting trends with increased greening; however, excessive drying and wetting can impede an increase in the NDVI (Berdugo et al. 2020; Zhao et al. 2020). The relationship between vegetation dynamics and climate change is nonlinear and complex. One reason for greening is the strong acclimation of grasslands, which cover most drylands, to climate change, allowing the development of high resilience and the ability of plants to recover from adverse conditions (Marcolla et al. 2011; Reichstein et al. 2013). However, grasslands are highly sensitive to drying, and drought events can significantly decrease terrestrial carbon sequestration (Xu et al. 2019). It is important to note that the effect of a single drought event on vegetation productivity may not be significant if it contradicts long-term trends owing to the nested hierarchical structure of complexity (Ryo et al. 2019). In addition, wetting promotes increased carbon sequestration in water-constrained drylands. However, NDVI does not increase indefinitely, as shifts in the dominance of other environmental factors can affect plant growth (Keenan and Riley 2018; Zhao et al. 2020). By utilizing the AI threshold and analyzing long-term trends, researchers can develop a better understanding of the relationship between vegetation dynamics and climate change, ultimately helping to protect vital ecosystems (Berdugo et al. 2020; Zhao et al. 2020). It is important to note that the effect of a single drought event on vegetation productivity may not be significant if it contradicts long-term trends owing to the nested hierarchical structure of complexity (Ryo et al. 2019). As aridity increases in drylands, the availability of soil water decreases, limiting photosynthesis and the ability of ecosystems to sequester carbon (Peng et al. 2013; Doughty et al. 2015; Frank et al. 2015; Xu et al. 2019). However, in wetter years, drylands can quickly become significant carbon sinks because of the less constrained availability of soil water, which leads to increased vegetation productivity (Poulter et al. 2014). Recent research suggests that “vegetation decline”, which is observed in satellite data, is a key feature in the initial stage of ecosystem transition (Berdugo et al. 2020).
5.3 Prediction Models in Sustainable SESs
Because the regime shifts of dryland ecosystems involve changing spatial organization of vegetation, ecohydrological processes, soil loss, and their interactions (Peters et al. 2015; Hoover et al. 2021; Grünzweig et al. 2022), it is necessary to develop a comprehensive model describing how regime shifts occur. Understanding these complex relationships is critical for predicting ecosystem shifts in the drylands and developing effective conservation strategies. The biotic and abiotic mechanisms (Mayor et al. 2013, 2019) underlying the state shifts, particularly the ecohydrological determinants of dryland ecosystem regime shifts (Hoover et al. 2021), need to be explored. There is growing recognition of the need to address context-specific socio-ecological feedback in drylands that involves threshold behaviors. Prediction models should account for the impacts of climate change and their potential effects on the resilience and sustainability of SESs. Dryland vegetation distribution is determined by the spatial pattern of precipitation, and the controlling drivers vary based on climatology, terrain, and ecological regions. Soil moisture-atmosphere feedback dominates land carbon uptake variability, and biogeochemical and biogeophysical feedback is important for predicting ecosystem carbon cycles (Yuan et al. 2019; Humphrey et al. 2021; Windisch et al. 2021). The association between climate change and vegetation dynamics is modulated by water availability, which can be tracked by monitoring the soil moisture (Zhou et al. 2021; Miralles-Wilhelm, 2022; Erofeeva, 2021). Changes in soil moisture-precipitation feedback and soil moisture-carbon coupling link precipitation and vegetation growth. The asymmetrical and nonlinear relationship between precipitation and aboveground net primary productivity significantly affects the global carbon cycle (Quan et al. 2019; Moon et al. 2019; Maurer et al. 2020). Variations in soil moisture could signify changes in this association, which affects vegetation-climate coupling and dryland carbon sequestration (Zhao et al. 2021; Martínez-Fernandez et al. 2021; Zhou et al. 2021; Humphrey et al. 2021). Moderate soil moisture can maintain reasonable surface-atmosphere feedback, strengthening the association (Özkan and Gökbulak 2017). Tracing changes in soil moisture is crucial for predicting the impact of climate change on future vegetation (Fig. 3.9) and can guide the development of climate-vegetation association models.

Schematic diagram of the effect of soil moisture on carbon sinks in drylands through the traceability effect. The effective association with sufficiently high correlation coefficients indicates the coupling of vegetation growth and precipitation. Blue arrows indicate the determinant effects. Orange lines indicate the interactions among elements. The purple arrow indicates the traceability effects of soil moisture on the PRE-VI association. PRE, precipitation. VI, vegetation index (Zhao et al. 2022)
The development of prediction models for sustainable SESs in drylands is a rapidly evolving interdisciplinary field. The integration of remote sensing, multi-criteria assessments, resilience, and participatory modeling approaches is critical for addressing the complex and context-specific feedback that underlies the sustainability and resilience of these systems. A multicriteria assessment framework for evaluating the sustainability of dryland ecosystems that incorporates social, economic, and environmental indicators has been provided for policy interventions to promote sustainable land use (Wang et al. 2023b). Machine learning techniques for predicting the future states of SESs based on historical data are used. For example, using data on water flow, temperature, and other environmental variables to accurately predict water quality levels up to several days in advance, which could help inform management decisions to maintain sustainable water resources (Islam et al. 2021). Integrated model that combines social and ecological factors was developed to predict the impacts of land-use change on ecosystem services in the Yucatan Peninsula of Mexico, such as carbon sequestration, water regulation, and biodiversity (Mendoza-Ponce et al. 2018). Participatory modeling approach was used to develop a predictive model for sustainable agriculture or rangeland ecosystems that incorporated local knowledge and expertise and was able to predict the impacts of different practices to promote sustainability (Gorripati et al. 2023). These models should be multidisciplinary and transdisciplinary and should account for the complexity, variability, and uncertainty of these systems. Trade-offs and synergies are often present in complex SESs, and prediction models should account for these interactions to avoid unintended consequences and optimize the sustainability and resilience of these systems. Since the dryland SESs are water-limited, the incorporation of impacts of climate change and the uncertainty into prediction models, along with a multidisciplinary and transdisciplinary approach, is critical for developing sustainable SES models for drylands.
6 Summary and Perspectives
Dryland SESs are complex and highly sensitive to both natural and anthropogenic disturbances, making them vulnerable to regime shifts that can have negative effects on the delivery of ecosystem services. To understand the mechanisms underlying regime shifts and the maintenance of stability and resilience in dryland SESs, it is essential to consider the interactions between the structure and function of these systems at multiple spatial scales. Hydrological feedback is particularly important in drylands, and climate change adds another layer of uncertainty to predicting the trajectory of local dryland SES changes. Therefore, related researches have significant implications for the management and conservation of dryland SESs, ultimately helping maintain the provision of ESs and promote sustainable development in these regions.
We highlight the importance of considering context-specific socio-ecological feedback that involves threshold behaviors in drylands. This feedback can be responsible for slow or rapid responses to changes in external drivers and feedback within SESs, ultimately leading to irreversible or persistent regime shifts. Thus, developing comprehensive indicators and models and introducing multivariate approaches are crucial for predicting the possibility of future regime shifts in dryland SESs. Understanding the biotic and abiotic mechanisms underlying regime shifts in dryland SESs and their stability from both holistic and context-specific perspectives is essential for identifying how dryland SESs change in specific contexts. This understanding can help address questions about tipping points and regime changes that may significantly affect the delivery of ESs in geographically different dryland SESs.
Future research priorities should include understanding the mechanisms and geographical heterogeneity of resilience and stability, developing comprehensive indicators and models, and introducing multivariable approaches to predict the possibility of regime shifts in dryland SESs, which identified in this theme, are critical for unraveling the complexities of dryland SESs and their interactions, predicting the possibility of regime shifts, and developing strategies to maintain their stability and resilience. The multidisciplinary nature of these research highlights the need for collaboration across various fields, including ecology, hydrology, economics, and the social sciences, to achieve a comprehensive understanding of dryland SESs and inform management decisions that promote sustainable development.
References
Abel C, Horion S, Tagesson T et al (2021) The human–environment nexus and vegetation–rainfall sensitivity in tropical drylands. Nat Sustain 4:25–32
Allen CR, Angeler DG, Cumming GS et al (2016) Quantifying spatial resilience. J Appl Ecol 53:625–635
Angeler DG, Allen CR (2016) EDITORIAL: quantifying resilience. J Appl Ecol 53:617–624
Bardgett RD, Bullock JM, Lavorel S et al (2021) Combatting global grassland degradation. Nat Rev Earth Environ 2(10):720–735
Bastin J-F, Berrahmouni N, Grainger A et al (2017) The extent of forest in dryland biomes. Science 356:635
Bawa KS, Sengupta A, Chavan V et al (2021) Securing biodiversity, securing our future: a national mission on biodiversity and human well-being for India. Biol Cons 253:108867
Bekele AE, Drabik D, Dries L et al (2021) Large-scale land investments, household displacement, and the effect on land degradation in semiarid agro-pastoral areas of Ethiopia. Land Degrad Dev 32:777–791
Belnap J, Lange OL (eds) (2003) Biological soil crusts: structure, function, and management. Springer, Berlin
Belnap J, Reynolds RL, Reheis MC et al (2009) Sediment losses and gains across a gradient of livestock grazing and plant invasion in a cool, semi-arid grassland, Colorado Plateau, USA. Aeol Res 1:27–43
Belnap J, Weber B, Büdel B (2016) Biological soil crusts as an organizing principle in drylands. In: Weber B, Büdel B, Belnap J (eds) Biological soil crusts: an organizing principle in drylands. Springer International Publishing, Cham, pp 3–13
Berdugo M, Kéfi S, Soliveres S et al (2017) Plant spatial patterns identify alternative ecosystem multifunctionality states in global drylands. Nat Ecol Evol 1:0003
Berdugo M, Delgado-Baquerizo M, Soliveres S et al (2020) Global ecosystem thresholds driven by aridity. Science 367:787
Berdugo M, Gaitán J, Delgado-Baquerizo M et al (2022) Prevalence and drivers of abrupt vegetation shifts in global drylands. Proc Natl Acad Sci USA 119:e2123393119
Bestelmeyer BT, Duniway MC, James DK et al (2013) A test of critical thresholds and their indicators in a desertification-prone ecosystem: more resilience than we thought. Ecol Lett 16:339–345
Bestelmeyer BT, Okin GS, Duniway MC et al (2015) Desertification, land use, and the transformation of global drylands. Front Ecol Environ 13(1):28–36
Blanco-Canqui H, Laird DA, Heaton E et al (2020) Soil carbon increased by twice the amount of biochar carbon applied after six years: field evidence of negative priming. GCB Bioenergy 12:240–251
Bochet E, Molina MJ, Monleón V et al (2021) Interactions of past human disturbance and aridity trigger abrupt shifts in the functional state of Mediterranean holm oak woodlands. CATENA 206:105514
Bodin Ö, Prell C (2011) Social networks and natural resource management: uncovering the social fabric of environmental governance. Cambridge University Press, Cambridge
Bowker MA, Miller ME, Belote RT (2012) Assessment of rangeland ecosystem conditions, salt creek watershed and dugout ranch, Southeastern Utah. US Geological Survey Open-File Report 2012–1061. http://pubs.usgs.gov/of/2012/1061/. Accessed 21 Sept 2022
Brandt M, Wigneron J-P, Chave J et al (2018) Satellite passive microwaves reveal recent climate-induced carbon losses in African drylands. Nat Ecol Evol 2:827–835
Burthe SJ, Henrys PA, Mackay EB et al (2016) Do early warning indicators consistently predict nonlinear change in long-term ecological data? J Appl Ecol 53:666–676
Buxton JE, Abrams JF, Boulton CA et al (2022) Quantitatively monitoring the resilience of patterned vegetation in the Sahel. Glob Change Biol 28:571–587
Carpenter SR, Brock WAB, Hanson PC (1999) Ecological and social dynamics in simple models of ecosystem management. Conserv Ecol 3:4
Carpenter SR, Folke C, Norström A et al (2012) Program on ecosystem change and society: an international research strategy for integrated social–ecological systems. Curr Opin Environ Sustain 4:134–138
Chen S, Wang W, Xu W et al (2018) Plant diversity enhances productivity and soil carbon storage. Proc Natl Acad Sci USA 115:4027–4032
Chen C, He B, Yuan WP et al (2019a) Increasing interannual variability of global vegetation greenness. Environ Res Lett 14(12):124005
Chen C, Park T, Wang X et al (2019b) China and India lead in greening of the world through land-use management. Nat Sustain 2:122–129
Cherlet M, Hutchinson C, Reynolds J et al (2018) World atlas of desertification. Publications Office of the European Union, Luxembourg
Clark WC, Dickson NM (2003) Sustainability science: the emerging research program. Proc Natl Acad Sci USA 100(14):8059–8061
Colding J, Barthel S (2019) Exploring the social-ecological systems discourse 20 years later. Ecol Soc 24(1):2
Corrado R, Cherubini AM, Pennetta C (2014) Early warning signals of desertification transitions in semiarid ecosystems. Phys Rev E 90(6):062705
Cumming GS, Barnes G, Perz S et al (2005) An exploratory framework for the empirical measurement of resilience. Ecosystems 8:975–987
Davin EL, Rechid D, Breil M et al (2020) Biogeophysical impacts of forestation in Europe: first results from the LUCAS (Land Use and Climate Across Scales) regional climate model intercomparison. Earth Syst Dynam 11:183–200
D’Odorico P, Bhattachan A (2012) Hydrologic variability in dryland regions: impacts on ecosystem dynamics and food security. Philos Trans R Soc B Biol Sci 367:3145–3157
D’Odorico P, Bhattachan A, Davis KF et al (2013) Global desertification: drivers and feedbacks. Adv Water Resour 51:326–344
Davin EL, de Noblet-Ducoudré N (2010) Climatic impact of global-scale deforestation: radiative versus nonradiative processes. J Clim 23:97–112
Dearing J, Acma B, Bub S et al (2015) Social-ecological systems in the Anthropocene: the need for integrating social and biophysical records at regional scales. Anthropocene Rev 2:220–246
Delgado-Baquerizo M, Maestre FT, Reich PB et al (2016) Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat Commun 7:10541
Díaz S, Kattge J, Cornelissen J et al (2016) The global spectrum of plant form and function. Nature 529:167–171
Doerr SH, Shakesby RA, Walsh RPD (2000) Soil water repellency: its causes, characteristics and hydro-geomorphological significance. Earth Sci Rev 51:33–65
Doughty CE, Metcalfe DB, Girardin CAJ et al (2015) Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 519:78–82
Eby S, Agrawal A, Majumder S et al (2017) Alternative stable states and spatial indicators of critical slowing down along a spatial gradient in a savanna ecosystem. Glob Ecol Biogeogr 26(6):638–649
Eldridge DJ, Greene RSB (1994) Microbiotic soil crusts-a review of their roles in soil and ecological processes in the rangelands of Australia. Soil Res 32(3):389–415
Eldridge DJ, Poore AGB, Ruiz-Colmenero M et al (2016) Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecol Appl 26:1273–1283
Ernstson H, van der Leeuw SE, Redman CL et al (2010) Urban transitions: on urban resilience and human-dominated ecosystems. Ambio 39:531–545
Erofeeva EA (2021) Plant hormesis and shelford’s tolerance law curve. J For Res 32(5):1789–1802
Feng S, Fu Q (2013) Expansion of global drylands under a warming climate. Atmos Chem Phys 13:10081–10094
Feng X, Fu B, Piao S et al (2016) Revegetation in China’s Loess Plateau is approaching sustainable water resource limits. Nat Clim Chang 6:1019–1022
Fisher JB, Whittaker RJ, Malhi Y (2011) ET come home: potential evapotranspiration in geographical ecology. Glob Ecol Biogeogr 20(1):1–18
Folke C (2006) Resilience: the emergence of a perspective for social–ecological systems analyses. Glob Environ Chang 16:253–267
Folke C, Holling CS, Perrings C (1996) Biological diversity, ecosystems, and the human scale. Ecol Appl 6:1018–1024
Folke C, Carpenter S, Walker B et al (2004) Regime shifts, resilience, and biodiversity in ecosystem management. Annu Rev Ecol Evol Syst 35:557–581
Folke C, Carpenter SR, Walker B et al (2010) Resilience thinking: integrating resilience, adaptability and transformability. Ecol Soc 15:20
Frank D, Reichstein M, Bahn M et al (2015) Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Glob Change Biol 21:2861–2880
Fu B, Stafford-Smith M, Wang Y et al (2021) The Global-DEP conceptual framework—research on dryland ecosystems to promote sustainability. Curr Opin Environ Sustain 48:17–28
Gaitán JJ, Oliva GE, Bran DE, et al (2015) Vegetation structure is as important as climate for explaining ecosystem function across Patagonian rangelands. J Ecol 102:1419–1428
García-Palacios P, Gross N, Gaitán J et al (2018) Climate mediates the biodiversity–ecosystem stability relationship globally. Proc Natl Acad Sci USA 115:8400–8405
García-Ruiz JM, López-Moreno JI, Vicente-Serrano SM et al (2011) Mediterranean water resources in a global change scenario. Earth Sci Rev 105(3–4):121–139
Global Drylands Initiative (GDI) (2021) Restoring drylands: from degradation to regeneration for sustainable development. GDI
González-Sánchez EJ, Kassam A, Basch G et al (2016) Conservation agriculture and its contribution to the achievement of agri-environmental and economic challenges in Europe. AIMS Agric Food 1(4):387–408
Gorripati R, Thakur M, Kolagani N (2023) Promoting climate resilient sustainable agriculture through participatory system dynamics with crop-water-income dynamics. Water Resour Manage 37:3935–3951
Greve P, Orlowsky B, Mueller B et al (2014) Global assessment of trends in wetting and drying over land. Nat Geosci 7:716–721
Grimm NB, Faeth SH, Golubiewski NE et al (2008) Global change and the ecology of cities. Science 319:756–760
Grimm NB, Chapin FS III, Bierwagen B et al (2013) The impacts of climate change on ecosystem structure and function. Front Ecol Environ 11:474–482
Grünzweig JM, De Boeck HJ, Rey A et al (2022) Dryland mechanisms could widely control ecosystem functioning in a drier and warmer world. Nat Ecol Evol 6:1064–1076
Guo Y, Zhao H, Zuo X et al (2008) Biological soil crust development and its topsoil properties in the process of dune stabilization, Inner Mongolia, China. Environ Geol 54:653–662
Hallett LM, Stein C, Suding KN (2017) Functional diversity increases ecological stability in a grazed grassland. Oecologia 183:831–840
Hausfather Z, Marvel K, Schmidt GA et al (2022) Climate simulations: recognize the ‘hot model’ problem. Nature 605:26–29
He N, Zhang Y, Yu Q et al (2011) Grazing intensity impacts soil carbon and nitrogen storage of continental steppe. Ecosphere 2(1):1–10
He B, Wang S, Guo L et al (2019) Aridity change and its correlation with greening over drylands. Agric for Meteorol 278:107663
Hobbs RJ, Norton DA (1996) Towards a conceptual framework for restoration ecology. Restor Ecol 4(2):93–110
Holling CS (1973) Resilience and stability of ecological systems. Annu Rev Ecol Syst 4:1–23
Hoover DL, Pfennigwerth AA, Duniway MC (2021) Drought resistance and resilience: The role of soil moisture–plant interactions and legacies in a dryland ecosystem. J Ecol 109:3280–3294
Hu W, Ran J, Dong L et al (2021) Aridity-driven shift in biodiversity–soil multifunctionality relationships. Nat Commun 12:5350
Huang G, Li Y, Su YG (2015) Effects of increasing precipitation on soil microbial community composition and soil respiration in a temperate desert, Northwestern China. Soil Biol Biochem 83:52–56
Huang J, Yu H, Guan X et al (2016) Accelerated dryland expansion under climate change. Nat Clim Chang 6:166–171
Huang M, Piao S, Ciais P et al (2019) Air temperature optima of vegetation productivity across global biomes. Nat Ecol Evol 3:772–779
Humphrey V, Berg A, Ciais P et al (2021) Soil moisture-atmosphere feedback dominates land carbon uptake variability. Nature 592(7852):65–69
Huston MA (2014) Disturbance, productivity, and species diversity: empiricism vs logic in ecological theory. Ecology 95:2382–2396
Intergovernmental Panel on Climate Change (IPCC) (2019) Climate change and land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems. https://www.ipcc.ch/srccl/
Irob K, Blaum N, Weiss-Aparicio A et al (2023) Savanna resilience to droughts increases with the proportion of browsing wild herbivores and plant functional diversity. J Appl Ecol 60(2):251–262
Islam ARMT, Talukdar S, Mahato S et al (2021) Flood susceptibility modelling using advanced ensemble machine learning models. Geosci Front 12(3):101075
Jobbágy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol Appl 10:423–436
Keenan TF, Riley WJ (2018) Greening of the land surface in the world’s cold regions consistent with recent warming. Nat Clim Chang 8:825–828
Kéfi S, Guttal V, Brock WA et al (2014) Early warning signals of ecological transitions: methods for spatial patterns. PLoS ONE 9:e92097
Köchy M, Hiederer R, Freibauer A (2015) Global distribution of soil organic carbon—Part 1: masses and frequency distributions of SOC stocks for the tropics, permafrost regions, wetlands, and the world. Soil 1:351–365
Lal R (2004) Soil carbon sequestration to mitigate climate change. Geoderma 123:1–22
Lal R (2015) Restoring soil quality to mitigate soil degradation. Sustainability 7:5875–5895
Lavorel S, Grigulis K (2012) How fundamental plant functional trait relationships scale-up to trade-offs and synergies in ecosystem services. J Ecol 100:128–140
Lepš J, Májeková M, Vítová A et al (2018) Stabilizing effects in temporal fluctuations: management, traits, and species richness in high-diversity communities. Ecology 99(2):360–371
Liu HJ, Han XG, Li LH et al (2009) Grazing density effects on cover, species composition, and nitrogen fixation of biological soil crust in an Inner Mongolia Steppe Rangeland. Ecol Manag 62:321–327
Liu F, Archer SR, Gelwick F et al (2013) Woody plant encroachment into grasslands: spatial patterns of functional group distribution and community development. PLoS ONE 8:e84364
Loreau M, Mazancourt CD (2013) Biodiversity and ecosystem stability: a synthesis of underlying mechanisms. Ecol Lett 16:106
Maestre FT, Bowker MA, Cantón Y et al (2011) Ecology and functional roles of biological soil crusts in semi-arid ecosystems of Spain. J Arid Environ 75:1282–1291
Maestre FT, Quero JL, Gotelli NJ et al (2012) Plant species richness and ecosystem multifunctionality in global drylands. Science 335:214–218
Maestre FT, Delgado-Baquerizo M, Jeffries TC et al (2015) Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc Natl Acad Sci USA 112:15684–15689
Maestre FT, Eldridge DJ, Soliveres S et al (2016) Structure and functioning of dryland ecosystems in a changing world. Annu Rev Ecol Evol Syst 47:215–237
Maestre FT, Benito BM, Berdugo M et al (2021) Biogeography of global drylands. New Phytol 231(2):540–558
Marcolla B, Cescatti A, Manca G et al (2011) Climatic controls and ecosystem responses drive the inter-annual variability of the net ecosystem exchange of an alpine meadow. Agric for Meteorol 151:1233–1243
Martínez-Fernández et al., 2021 Martínez-Fernández J, Gonz´alez-Zamora A, Almendra-Martín L (2021) Soil moisture memory and soil properties: an analysis with the stored precipitation fraction. J Hydrol 593:125622
Martínez-Vilalta J, Lloret F (2016) Drought-induced vegetation shifts in terrestrial ecosystems: the key role of regeneration dynamics. Glob Planet Change 144:94–108
Maurer GE, Hallmark AJ, Brown RF et al (2020) Sensitivity of primary production to precipitation across the United States. Ecol Lett 23(3):527–536
Mayor AG, Kefi S, Bautista S et al (2013) Feedbacks between vegetation pattern and resource loss dramatically decrease ecosystem resilience and restoration potential in a simple dryland model. Landscape Ecol 28:931–942
Mayor AG, Bautista S, Rodriguez F et al (2019) Connectivity-mediated ecohydrological feedbacks and regime shifts in drylands. Ecosystems 22:1497–1511
Mbow C, Smith P, Skole D et al (2014) Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr Opin Environ Sustain 6:8–14
McGuire KJ, McDonnell JJ (2010) Hydrological connectivity of hillslopes and streams: characteristic time scales and nonlinearities. Water Resour Res 46(10):W10543
Meloni F, Nakamura GM, Granzotti CRF et al (2019) Vegetation cover reveals the phase diagram of patch patterns in drylands. Physica A 534:122048
Mendoza-Ponce A, Corona-Nunez R, Kraxner F et al (2018) Identifying effects of land use cover changes and climate change on terrestrial ecosystems and carbon stocks in Mexico. Glob Environ Chang 53:12–23
Midgley GF (2012) Biodiversity and ecosystem function. Science 335:174–175
Migliavacca M, Musavi T, Mahecha MD et al (2021) The three major axes of terrestrial ecosystem function. Nature 598:468–472
Millennium Ecosystem Assessment, 2005 Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: desertification synthesis. World Resources Institute, Washington, DC
Miralles-Wilhelm F (2022) Water is the middle child in global climate policy. Nat Clim Chang 12(2):110–112
Moon H, Guillod BP, Gudmundsson L et al (2019) Soil moisture effects on afternoon precipitation occurrence in current climate models. Geophys Res Lett 46(3):1861–1869
Moreno-Jiménez E, Plaza C, Saiz H et al (2019) Aridity and reduced soil micronutrient availability in global drylands. Nat Sustain 2:371–377
Müller F (1997) State-of-the-art in ecosystem theory. Ecol Model 100:135–161
Naeem S, Bunker DE, Hector A et al (2009) Biodiversity, ecosystem functioning, and human wellbeing: an ecological and economic perspective. Oxford University Press, Oxford
Nemani RR, Keeling CD, Hashimoto H, et al (2003) Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300:1560–1563
Nijp JJ, Temme AJ, van Voorn GA et al (2019) Spatial early warning signals for impending regime shifts: a practical framework for application in real-world landscapes. Glob Change Biol 25:1905–1921
Nowak DJ, Crane DE, Stevens JC (2006) Air pollution removal by urban trees and shrubs in the United States. Urban For Urban Green 4:115–123
Ochoa-Hueso R, Piñeiro J, Power S (2019) Decoupling of nutrient cycles in a Eucalyptus woodland under elevated CO2. J Ecol 107:2532–2540
Oliver TH, Heard MS, Isaac NJ et al (2015) Biodiversity and resilience of ecosystem functions. Trends Ecol Evol 30:673–684
Omuto CT, Vargas RR, Alim MS et al (2010) Mixed-effects modelling of time series NDVI-rainfall relationship for detecting human-induced loss of vegetation cover in drylands. J Arid Environ 74(11):1552–1563
Osem Y, Yavlovich H, Zecharia N et al (2013) Fire-free natural regeneration in water limited Pinus halepensis forests: a silvicultural approach. Eur J Forest Res 132:679–690
Ostrom E (2009) A general framework for analyzing sustainability of social-ecological systems. Science 325(5939):419–422
Özkan U, Gökbulak F (2017) Effect of vegetation change from forest to herbaceous vegetation cover on soil moisture and temperature regimes and soil water chemistry. CATENA 149:158–166
Peng S, Piao S, Ciais P et al (2013) Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 501:88–92
Peters D, Havstad K, Archer S et al (2015) Beyond desertification: new paradigms for dryland landscapes. Front Ecol Environ 13:4–12
Peterson G, Allen CR, Holling CS (1998) Ecological resilience, biodiversity, and scale. Ecosystems 1:6–18
Piao S, Tan J, Chen A et al (2015) Leaf onset in the northern hemisphere triggered by daytime temperature. Nat Commun 6:6911
Pillar VD, Chiarucci A, Pärtel M et al (2018) Linking biodiversity to ecosystems: a task for plant community ecologists. J Veg Sci 29:1–3
Poulter B, Frank D, Ciais P et al (2014) Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 509:600–603
Quan Q, Tian D, Luo Y, et al (2019) Water scaling of ecosystem carbon cycle feedback to climate warming. Sci Adv 5(8):eaav1131
Rapport DJ, Whitford WG (1999) How ecosystems respond to stress: common properties of arid and aquatic systems. Bioscience 49:193–203
Ravi S, Breshears DD, Huxman TE et al (2010) Land degradation in drylands: interactions among hydrologic–aeolian erosion and vegetation dynamics. Geomorphology 116:236–245
Reichstein M, Bahn M, Ciais P et al (2013) Climate extremes and the carbon cycle. Nature 500:287–295
Reynolds JF, Stafford Smith DM, Lambin EF et al (2007) Global desertification: building a science for dryland development. Science 316:847–851
Richardson T, Forster P, Smith C et al (2019) Efficacy of climate forcings in PDRMIP models. J Gerontol Ser A Biol Med Sci 124:12824–12844
Rietkerk M, Dekker SC, de Ruiter PC et al (2004) Self-organized patchiness and catastrophic shifts in ecosystems. Science 305:1926
Rodriguez-Caballero E, Canton Y, Chamizo S et al (2013) Soil loss and runoff in semiarid ecosystems: a complex interaction between biological soil crusts, micro-topography, and hydrological drivers. Ecosystems 16:529–546
Ryo M, Aguilar-Trigueros CA, Pinek L et al (2019) Basic principles of temporal dynamics. Trends Ecol Evol 34(8):723–733
Saco PM, Moreno-de las Heras M, Keesstra S, et al. (2018) Vegetation and soil degradation in drylands: non linear feedbacks and early warning signals. Curr Opin Environ Sci Health 5:67–72
Saco PM, Rodríguez JF, Moreno-de las Heras M, et al. (2020) Using hydrological connectivity to detect transitions and degradation thresholds: applications to dryland systems. CATENA 186:104354
Sarah P (2004) Nonlinearity of ecogeomorphic processes along Mediterranean-arid transect. Geomorphology 60:303–317
Scheffer M, Carpenter SR, Foley JA et al (2001) Catastrophic shifts in ecosystems. Nature 413:591–596
Scheffer M, Carpenter SR, Lenton TM et al (2012) Anticipating critical transitions. Science 338:344–348
Scheffer M, Barrett S, Carpenter SR et al (2015) Creating a safe operating space for iconic ecosystems. Science 347:1317–1319
Schimel D, Melillo J, Tian H, et al (2000) Contribution of increasing CO2 and climate to carbon storage by ecosystems in the United States. Science 287:2004–2006
Schlaepfer DR, Bradford JB, Lauenroth WK et al (2017) Climate change reduces extent of temperate drylands and intensifies drought in deep soils. Nat Commun 8:14196
Schlesinger WH, Reynolds JF, Cunningham GL et al (1990) Biological feedbacks in global desertification. Science 247:1043
Schlüter M, Müller B, Frank K (2019) The potential of models and modeling for social-ecological systems research. Ecol Soc 24(1):31
Schröder A, Persson L, De Roos AM (2005) Direct experimental evidence for alternative stable states: a review. Oikos 110:3–19
Schowalter TD (2011) Ecosystem structure and function. In: Schowalter TD (ed) Insect ecology (3rd Edition). Academic Press, San Diego, pp. 327–358
Schwinning S, Sala OE (2004) Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 141:211–220
Schwinning S, Sala OE, Loik ME et al (2004) Thresholds, memory, and seasonality: understanding pulse dynamics in arid/semi-arid ecosystems. Oecologia 141:191–193
Seddon N, Smith A, Smith P et al (2021) Getting the message right on nature-based solutions to climate change. Glob Change Biol 27(8):1518–1546
Seto KC, Reenberg A, Boone CG et al (2012) Urban land teleconnections and sustainability. Proc Natl Acad Sci USA 109:7687–7692
Seymour F, Harris NL (2019) Reducing tropical deforestation. Science 365:756–757
Sharrow SH, Ismail S (2004) Carbon and nitrogen storage in agroforests, tree plantations, and pastures in western Oregon, USA. Agrofor Syst 60:123–130
Sheffield J, Wood EF, Roderick ML (2012) Little change in global drought over the past 60 years. Nature 491(7424):435–438
Siefert A, Violle C, Chalmandrier L et al (2015) A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol Lett 18:1406–1419
Smiraglia D, Ceccarelli T, Bajocco S et al (2016) Linking trajectories of land change, land degradation processes and ecosystem services. Environ Res 147:590–600
Soliveres S, Maestre FT, Eldridge DJ et al (2014) Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands. Glob Ecol Biogeogr 23:1408–1416
Stavi I, Yizhaq H, Szitenberg A et al (2021) Patch-scale to hillslope-scale geodiversity alleviates susceptibility of dryland ecosystems to climate change: insights from the Israeli Negev. Curr Opin Environ Sustain 50:129–137
Strassburg BBN, Rodrigues ASL, Gusti M et al (2012) Impacts of incentives to reduce emissions from deforestation on global species extinctions. Nat Clim Chang 2:350–355
Teague WR, Apfelbaum S, Lal R et al (2016) The role of ruminants in reducing agriculture’s carbon footprint in North America. J Soil Water Conserv 71(2):156–164
Tietjen B, Jeltsch F, Zehe E et al (2010) Effects of climate change on the coupled dynamics of water and vegetation in drylands. Ecohydrology 3:226–237
Turnbull L, Wainwright J (2019) From structure to function: understanding shrub encroachment in drylands using hydrological and sediment connectivity. Ecol Ind 98:608–618
Turnbull L, Wilcox BP, Belnap J et al (2012) Understanding the role of ecohydrological feedbacks in ecosystem state change in drylands. Ecohydrology 5:174–183
Ukkola AM, Prentice IC, Keenan TF et al (2015) Reduced streamflow in water-stressed climates consistent with CO2 effects on vegetation. Nat Clim Chang 6:75–78
Urgeghe AM, Breshears DD, Martens SN et al (2010) Redistribution of runoff among vegetation patch types: on ecohydrological optimality of herbaceous capture of run-on Rangeland. Ecol Manag 63:497–504
Ursino N (2019) Dynamic models of socio-ecological systems predict catastrophic shifts following unsustainable development. Sci Total Environ 654:890–894
Valone TJ, Meyer M, Brown JH et al (2002) Timescale of perennial grass recovery in desertified arid grasslands following livestock removal. Conserv Biol 16(4):995–1002
Vandandorj S, Eldridge DJ, Travers SK et al (2017) Microsite and grazing intensity drive infiltration in a semiarid woodland. Ecohydrology 10:e1831
Voosen P (2022) Hot’ climate models exaggerate earth impacts. Science 376:685
Walker TW, Adams AFR (1958) Studies on soil organic matter: I Influence of phosphorus content of parent materials on accumulations of carbon, nitrogen, sulfur, and organic phosphorus in grassland soils. Soil Sci 85:307–318
Wang SQ, Yu GR (2008) Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecol Sin 28:3937–3947
Wang L, Manzoni S, Ravi S et al (2015) Dynamic interactions of ecohydrological and biogeochemical processes in water-limited systems. Ecosphere 6(8):1–27
Wang B, Waters C, Orgill S et al (2018) High resolution mapping of soil organic carbon stocks using remote sensing variables in the semi-arid rangelands of eastern Australia. Sci Total Environ 630:367–378
Wang J, Zhen J, Hu W et al (2023a) Remote sensing of soil degradation: progress and perspective. Int Soil Water Conserv Res 11(3):429–454
Wang H, Liu Y, Wang Y et al (2023b) Land cover change in global drylands: a review. Sci Total Environ 863:160943
Wang-Erlandsson L, Fetzer I, Keys PW et al (2018) Remote land use impacts on river flows through atmospheric teleconnections. Hydrol Earth Syst Sci 22:4311–4328
Windisch MG, Davin EL, Seneviratne SI (2021) Prioritizing forestation based on biogeochemical and local biogeophysical impacts. Nat Clim Chang 11(10):867–871
Xu L, Myneni RB, Chapin Iii FS et al (2013) Temperature and vegetation seasonality diminishment over northern lands. Nat Clim Chang 3(6):581–586
Xu H, Wang X, Yang T (2017) Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Sci Total Environ 579:1658–1674
Xu H, Wang X, Zhao C et al (2018) Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric for Meteorol 262:1–13
Xu C, McDowell NG, Fisher RA et al (2019) Increasing impacts of extreme droughts on vegetation productivity under climate change. Nat Clim Chang 9:948–953
Yahdjian L, Gherardi L, Sala OE (2011) Nitrogen limitation in arid-subhumid ecosystems: a meta-analysis of fertilization studies. J Arid Environ 75:675–680
Yang Z, Baoyin T, Minggagud H et al (2017) Recovery succession drives the convergence, and grazing versus fencing drives the divergence of plant and soil N/P stoichiometry in a semiarid steppe of Inner Mongolia. Plant Soil 420:303–314
Yuan W, Zheng Y, Piao S, et al. (2019) Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci Adv 5:eaax1396
Yu Z, Yang G, Zuo S, Jørgensen G, et al (2020) Critical review on the cooling effect of urban blue-green space: a threshold-size perspective. Urban For. Urban Green 49:126630
Zaitchik BF, Rodell M, Biasutti M et al (2023) Wetting and drying trends under climate change. Nat Water 1(6):502–513
Zehe E, Elsenbeer H, Lindenmaier F et al (2007) Patterns of predictability in hydrological threshold systems. Water Resour Res 43:W07434
Zhang Y, Peng C, Li W et al (2013) Monitoring and estimating drought-induced impacts on forest structure, growth, function, and ecosystem services using remote-sensing data: recent progress and future challenges. Environ Rev 21(2):103–115
Zhang W, Kato E, Bhandary P et al (2016) Awareness and perceptions of ecosystem services in relation to land use types: evidence from rural communities in Nigeria. Ecosyst Serv 22:150–160
Zhang C, Wang Y, Jia X et al (2020) Impacts of shrub introduction on soil properties and implications for dryland revegetation. Sci Total Environ 742:140498
Zhao W, Hu Z, Guo Q et al (2020) Contributions of climatic factors to interannual variability of the vegetation index in Northern China Grasslands. J Clim 33:175–183
Zhao W, Yu X, Liu Y et al (2021) Distinct vegetation response to drying and wetting trends across an aridity threshold. Environ Res Commun 3:025002
Zhao W, Yu X, Xu C et al (2022) Dynamic traceability effects of soil moisture on the precipitation–vegetation association in drylands. J Hydrol 615:128645
Zhao Y, Wang X, Chen F et al (2023) Soil organic matter enhances aboveground biomass in alpine grassland under drought. Geoderma 433:116430
Zhou J, Fu B, Gao G et al (2016) Effects of precipitation and restoration vegetation on soil erosion in a semi-arid environment in the Loess Plateau, China. CATENA 137:1–11
Zhou S, Williams AP, Lintner BR et al (2021) Soil moisture–atmosphere feedbacks mitigate declining water availability in drylands. Nat Clim Chang 11:38–44
Zhu Z, Wang S, Woodcock CE (2015) Improvement and expansion of the Fmask algorithm: cloud, cloud shadow, and snow detection for Landsats 4–7, 8, and Sentinel 2 images. Remote Sens Environ 159:269–277
Zika M, Erb K-H (2009) The global loss of net primary production resulting from human-induced soil degradation in drylands. Ecol Econ 69:310–318
Zurlini G, Jones KB, Riitters KH et al (2014) Early warning signals of regime shifts from cross-scale connectivity of land-cover patterns. Ecol Ind 45:549–560
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2024 The Author(s)
About this chapter

Cite this chapter
Yu, X., Liu, Y., Niu, S., Zhao, W., Fu, C., Chen, Z. (2024). Structure, Functions, and Interactions of Dryland Ecosystems. In: Fu, B., Stafford-Smith, M. (eds) Dryland Social-Ecological Systems in Changing Environments. Springer, Singapore. https://doi.org/10.1007/978-981-99-9375-8_3
Download citation
DOI: https://doi.org/10.1007/978-981-99-9375-8_3
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-9374-1
Online ISBN: 978-981-99-9375-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)