
Abstract
Evaluation of wheat end-use quality in terms of loaf volume (LV) requires enormous time and labor inputs. Hence, many studies have attempted to use grain, flour and dough properties to predict LV. Many quantitative trait loci (QTL) underlying these traits have also been identified to facilitate breeding. However, correlations between such predictive tests and LV as well as their QTLs could be influenced by the environment. In this chapter, we review recent literature on the correlations and G × E interaction (GEI) of the bread making quality traits grain protein content (GPC), sodium dodecyl sulphate sedimentation volume (SV), dough rheological traits (DRT) and LV. We briefly discuss our results from the evaluation of a hexaploid wheat recombinant inbred line population for GPC, SV, LV and nine DRT by mixograph analysis in six year-location environments in India, which revealed that correlations between DRT and LV were not stable across environments. In addition, GEI measured in terms of principal components using Additive main effects and multiplicative interaction model showed up to 47 % contribution to the total variation of the traits, which was reflected in the location-specificity of QTLs expressed in single as well as multiple environments. Even though 16 QTL clusters for four to seven traits were identified, only one of them involved LV. The strong influence of the environment on complex interrelationships between DRT and the other end-use quality traits suggested that during breeding for wheat end-use quality, marker-based selection of these traits would be more efficient if specific agro-climatic zones are targeted separately.
You have full access to this open access chapter, Download conference paper PDF
Similar content being viewed by others


Keywords
Selection for End-Use Quality Traits in Hexaploid Wheat: The Challenges
Improving wheat grain quality to cater to its diverse end uses is necessary to make its cultivation profitable. The quality of the industrial products made from hexaploid wheat such as leavened loaf bread depends critically on the properties of the grain, flour, dough and its responses during baking. Selection in the early generations for the most perceptible indicator of bread making quality (BMQ) namely, loaf volume (LV) is difficult due to the requirement of substantial quantities of grain and lack of high-throughput in evaluating it. BMQ is determined by many complex traits like grain protein content (GPC), gluten strength, dough viscosity, extensibility and elasticity. Hence, to predict LV, dough rheological traits (DRT); gluten strength often determined by sodium dodecyl sulphate sedimentation volume (SV) and GPC have been evaluated. However, determining simple and reliable predictive tests for LV and identification of consistent quantitative trait loci (QTL) with substantial contribution for such traits to aid marker-assisted selection (MAS) have proved difficult. This is due to the complex inter-relationships among the traits, large number of QTLs governing them, their co-location and possible pleiotropic effects as well as genotype × environment interactions (GEI) (Mann et al. 2009; Li et al. 2013). In this Chapter, we discuss the recent literature on these aspects and present a brief summary of our work on QTL and GEI analyses of the BMQ contributing traits.
Predictive tests for LV need to have high correlations with it that are stable in diverse environmental conditions and are simple, quick, require small sample size and should be able to clearly distinguish between genotypes. The breeding material thus selected can eventually be evaluated for LV in advanced stages. Various instruments such as farinograph, alveograph and mixograph have been employed to assess dough rheological changes during processing for the quality control of the end products. Among these, the computerized mixograph instruments can measure DRT when flour-water blend is subjected to a fixed mixing time. Though mixograph characters like dough development time, peak height and width are often employed to evaluate BMQ, single or few mixograph parameters as predictors of loaf volume have not been conclusively established (Mann et al. 2009; Caffe-Treml et al. 2010). More recently, Li et al. (2013) suggested small-scale tests like swelling index of gluten and lactic acid retention capacity to predict LV; however, further studies on the stability of their correlations with LV are needed.
For both LV and its potentially predictive parameters, many QTLs have been identified; however, their consistent expression across environments is necessary for their effective use (Li et al. 2012; El-Feki et al. 2013 and references therein). We performed detailed GEI, correlation and QTL analyses of end-use quality traits in a recombinant inbred line (RIL) population of 105 individuals derived from a bread wheat cross HI977 × HD2329. HI977 (good BMQ) possesses Glu-A1 (2*), Glu-B1 (17 + 18) and Glu-D1 (5 + 10) and HD2329 has Glu-A1 (2*), Glu-B1 (7 + 9) and Glu-D1 (2 + 12) and is poor in BMQ. Phenotypic evaluations were performed in three agro-climatically diverse locations in India namely, Karnal, Kota and Pune for two consecutive years (2003–04; 2004–05) and 12 BMQ related traits, viz. GPC, SV, LV and nine mixograph traits that could quantitatively reflect the energy used during the mixing process, changes in dough consistency and elasticity, optimum dough development time and the rate of the gluten network breakdown during over-mixing were recorded (Table 33.1) (Elangovan et al. 2008, 2011; Prashant et al. 2015).
Contributions of G, E and GEI Effects for DRT and BMQ Traits
The contributions of genotype (G) and environment (E) main-effects can be preliminarily judged by ANOVA (Tsilo et al. 2011; Li et al. 2013); however, its third component could include the GEI effects and the error. The Additive main effects and multiplicative interaction (AMMI) analysis (Nachit et al. 1992) combines additive and multivariate approaches and improves the analysis of non-additive GEI effects. In our studies on the HI977 × HD2329 population, LV indicated the highest contribution from G (38 %). However, for all the traits, either E (47–74 %) or GEI (42–57 %) were the main contributors. AMMI analysis showed 6–47 % contribution from GEI to total variation in terms of 1 to 4 principal component axes (Prashant et al. 2015). Similarly, Hristov et al. (2010) showed significant GEI contribution to LV, SV and GPC by AMMI analysis. It is possible that GEI could influence correlations among LV and its predictive tests thereby making it necessary to examine them individually in year-location combinations.
Trait Correlations and the Influence of the Environment
Using the average of the data from different years and locations (environments) to calculate trait correlations masks the effect of GEI and suppresses the patterns of trait inter-relationships across environments (Tsilo et al. 2011; Li et al. 2012). Mann et al. (2009) evaluated genetic correlations among DRT and BMQ traits in three individual environments that indicated consistent negative correlation between Maximum height of the Extensograph curve and LV. However, the stability of the correlations across multiple years in different locations could not be conclusively assessed since data were recorded at two locations, but in single year in one of them. We could efficiently document the repeatability of the correlations among LV, GPC, SV and DRT in HI977 × HD2329 RIL population as we assessed them in six individual year-location environments (Fig. 33.1). We found that correlations between 15 mixograph trait pairs were highly consistent since they were significant in five or in all the six environments. Among the rest, significant correlations were observed in one to four environments. MRI and MPT with LV and EPI, MRI and MPT with SV were significant in three environments and were the only DRT-BMQ trait pairs that showed fairly consistent correlations. However, majority of the correlations that were significant in only one or two environments involved LV, SV or GPC. Eleven trait pairs showed correlations that were inconsistent since they were positive or negative in different year-locations (Prashant et al. 2015). The environmental influence on correlations between predictive tests and LV, as well as significant GEI contributions suggested that even if a QTL with high contribution to predictive tests is detected, its stability across environments is important if it has to be considered for use in MAS for LV.

Pattern of correlations among dough rheological traits, loaf volume, SDS-sedimentation volume and grain protein content in HI977 × HD2329 population. ns- non-significant
Clustering and Location-Specificity of BMQ and DRT QTLs
Composite Interval Mapping (CIM) determined QTLs on all the chromosomes of bread wheat for LV, GPC, SV and DRT (Li et al. 2012; El-Feki et al. 2013; references therein). In addition, QTL clusters mainly for GPC, SV and DRT (Li et al. 2012) and for DRT and LV (Mann et al. 2009; Tsilo et al. 2011; Simons et al. 2012) suggested that as the traits are correlated, they could be controlled by common QTLs. But it needs to be noted that the HMW glutenin loci Glu-B1 or Glu-D1 often showed overlap with the largest of such clusters for DRT and LV and suggested that extensive studies to identify novel loci on other chromosomes are necessary for further improvement of wheat quality. In our analyses on HI977 ×HD2329 population, 158 QTLs for LV, SV, GPC and DRT were identified with 9–46 % contribution to phenotypic variance. There were 16 QTL clusters on nine chromosomes with co-locating QTLs for four to seven traits. Only a single QTL cluster on chromosome 5B showed co-location for LV and DRT.
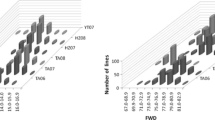
In many of the previous studies, large number of DRT and BMQ QTLs were detected in single environments (Mann et al. 2009; Tsilo et al. 2011; Li et al. 2012; Simons et al. 2012) and stable QTLs were very few, which often co-located with HMW glutenin loci (Mann et al. 2009; Tsilo et al. 2011). Examining their specificity to individual locations and pattern of detection across years within a location can help reveal their responses to GEI. In our study, 69 % of the QTLs were identified in single environments and only 15 % were detected in three or more environments. The significant influence of GEI was reflected in the pattern in which QTLs were detected across environments. We identified 109 QTLs for the traits analyzed, which were specifically expressed in single years in Karnal, Kota and Pune (Fig. 33.2). Interestingly, majority of them (47 %) were specific to Karnal and the rest were equally distributed in other two locations. In addition, the individual traits showed differential location-specificity for such QTLs. Furthermore, when the QTLs expressed in two or more environments were examined, 62 % showed expression in consecutive years in Karnal, followed by 34 % in Kota and only 4 % at Pune (Prashant et al. 2015).

Location-specificity of QTLs for dough rheological traits, loaf volume, SDS-sedimentation volume and grain protein content detected in single years in HI977 × HD2329 population
Conclusions
The GEI contributions, trait correlations and QTL clusters reinforce the view that wheat quality is achieved by complex gene repertoire and action underlying the levels and composition of grain constituents. The QTL clusters provide leads for characterization of genomic regions controlling end-use quality traits in wheat. In the coming years, genetic maps with better resolution and the wheat genome sequence would help resolve pleiotropic gene effects and multiple genes with close linkage. The location-specificity of QTLs detected in both single-environment and across years in HI977 × HD2329 population suggested that for traits exhibiting high GEI, it might be beneficial to target specific agro-climatic zones while breeding varieties for quality traits. In addition, QTL validation in those zones in multiple years using many wheat genotypes is necessary for their use in MAS.
References
Caffe-Treml M, Glover KD, Krishnan PG, Hareland GA (2010) Variability and relationships among Mixolab, Mixograph, and baking parameters based on multi-environment spring wheat trials. Cereal Chem 87:574–580
Elangovan M, Rai R, Dholakia BB et al (2008) Molecular genetic mapping of quantitative trait loci associated with loaf volume in hexaploid wheat (Triticum aestivum). J Cereal Sci 47:587–598
Elangovan M, Dholakia BB, Rai R et al (2011) Mapping QTL associated with agronomic traits in bread wheat (Triticum aestivum L.). J Wheat Res 3:14–23
El-Feki WM, Byrne PF, Reid SD et al (2013) Quantitative trait locus mapping for end-use quality traits in hard winter wheat under contrasting soil moisture levels. Crop Sci 53:1953–1967
Hristov N, Mladenov N, Djuric V et al (2010) Genotype by environment interactions in wheat quality breeding programs in southeast Europe. Euphytica 174:315–324
Li Y, Zhou R, Wang J et al (2012) Novel and favorable QTL allele clusters for end-use quality revealed by introgression lines derived from synthetic wheat. Mol Breed 29:627–643
Li Y, Wua Y, Hernandez-Espinosa N, Peña RJ (2013) The influence of drought and heat stress on the expression of end-use quality parameters of common wheat. J Cereal Sci 57:73–78
Mann G, Diffey S, Cullis B et al (2009) Genetic control of wheat quality: interactions between chromosomal regions determining protein content and composition, dough rheology, and sponge and dough baking properties. Theor Appl Genet 118:1519–1537
Nachit MM, Nachit G, Ketata H et al (1992) Use of AMMI and linear regression models to analyze genotype-environment interaction in durum wheat. Theor Appl Genet 83:597–601
Prashant R, Elangovan M, Rai R et al (2015) Genotype x environment interactions and QTL clusters underlying dough rheology traits in Triticum aestivum L. J Cereal Sci 64:82–91
Simons K, Anderson JA, Mergoum M et al (2012) Genetic mapping analysis of bread-making quality traits in spring wheat. Crop Sci 52:2182–2197
Tsilo TJ, Simsek S, Ohm J-B et al (2011) Quantitative trait loci influencing endosperm texture, dough-mixing strength, and bread-making properties of the hard red spring wheat breeding lines. Genome 54:460–470
Acknowledgments
We gratefully acknowledge the Department of Biotechnology (DBT), Government of India for financial support for this work and Bio-CARe Fellowship for RP and the Council of Scientific and Industrial Research, Government of India research fellowships for RP and EM. We thank Drs. Meena Lagu and Bhushan Dholakia, CSIR-NCL, Pune, for help in planning the experiment and Dr. Marion Röder, IPK, Gatersleben, Germany, for facilitating SSR genotyping of the HI977 × HD2329 population.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this paper
Cite this paper
Prashant, R. et al. (2015). Complex G × E Interactions and QTL Clusters Govern End-Use Quality Traits in Hexaploid Wheat. In: Ogihara, Y., Takumi, S., Handa, H. (eds) Advances in Wheat Genetics: From Genome to Field. Springer, Tokyo. https://doi.org/10.1007/978-4-431-55675-6_33
Download citation
DOI: https://doi.org/10.1007/978-4-431-55675-6_33
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-55674-9
Online ISBN: 978-4-431-55675-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)