
Abstract
Numerous cell types are involved in maintenance of the intestinal tissue. However, the main players are cells of the epithelial lining and from the immune system. All these cells are communicating with each other and are strongly influenced by interactions with neighboring cells. Mono-cultivation of cells may provide valuable information as to understanding basic biology, but co-cultivation of more than one type of cells gives an opportunity to investigate effects of intercellular communication. Examples of systems for co-cultivation of epithelial cells and immune cells for the investigation of both direct cell–cell contact and communication with soluble factors will be discussed in this chapter. In addition, co-cultivation systems for differentiation to more specialized epithelial cells as microfold cells (M-cells) will be described.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Origin, Features and Mechanisms
The models described in this section include the cell lines Caco-2 and THP-1 in addition to peripheral mononuclear cells (PBMCs) and dendritic cells (DCs). Description of the origin, features and mechanisms of these cells are included in previous sections, and further information can be found in Chap. 10 for Caco-2, Chap. 14 for THP-1, Chap. 15 for PBMCs and Chap. 17 for DCs.
2 Relevance to Human In Vivo Situation
2.1 Co-culture Caco-2 and Dendritic Cells
Precise regulation of the intestinal barrier function is important for the maintenance of mucosal homeostasis and prevents the onset of uncontrolled inflammation (Pastorelli et al. 2013). A balanced and fine-tuned cross-talk between intestinal epithelial cells (IECs), immune cells and the intestinal microbiota is necessary to maintain the intestinal homeostasis (Artis 2008). Different immune cells communicate with each other as well as with epithelial cells by an impressive network of regulatory signals. The complexity of the mucosal immune system is difficult to mimic in vitro, but a co-culture system makes it possible to elaborate mechanisms involved in communication between epithelial cells and cells of the immune system. It is possible to study effects of soluble molecules as well as contact dependent mechanisms. DCs are professional antigen-presenting cells central to the regulation of both innate and adaptive immune responses at mucosal surfaces (Coombes and Powrie 2008). The intestinal epithelium is not only a barrier to microorganisms entering via the oral route, but epithelial cells are directly influencing the activating properties of mucosal DCs (Rimoldi et al. 2004). Communication between immune cells and epithelial cells can be studied by co-cultivation of Caco-2 cells with CD14+ monocyte-derived dendritic cells (mDC), PBMCs or THP-1 cells, a human monocytic cell line (described in Chap. 14). CD14+ monocytes isolated from blood are differentiated into DCs by stimulation with GM-CSF and IL-4. The phenotype of these in vitro differentiated DC resembles that of inflammatory DCs (Segura et al. 2013). Inflammatory DCs differentiate from monocytes recruited to inflamed tissue from blood during an inflammation. Therefore the CD14+ monocytes are phenotypically distinct from the bone marrow-derived DC precursors which give rise to tissue resident DCs (Steinman and Idoyaga 2010; Miller et al. 2012) (see Chap. 17 for further information on DCs).
2.2 Co-culture Caco-2 and B-cells (Raji)
The follicle-associated epithelium (FAE) overlying the organized lymphoid follicles of the gut-associated lymphoid tissues (GALT) including Peyer’s patches, are specialized for sampling luminal content. Approximately 10 % of the cells within the FAE are microfold (M) cells. M cells have unique morphological features that include a reduction in both glycocalyx and microvilli compared to other parts of the epithelial lining. They are highly specialized for phagocytosis and transcytosis of gut luminal content across the epithelium. Beneath the M-cell’s basolateral membrane there is an intraepithelial pocket where the luminal content samples are released after transcytosis. This dome structure contains various populations of lymphocytes, macrophages and dendritic cells. M-cells therefore provide an efficient delivery of gut luminal microorganisms and antigens to the underlying mucosal tissue and are therefore essential for mucosal immune responses (Gullberg et al. 2000; Kerneis et al. 1997). Cells with a M cell-similar phenotype can be obtained by co-cultivation of Caco-2 cells with Raji B cells (Gullberg et al. 2000).
3 Stability, Consistency and Reproducibility
Co-culture systems of epithelial cells and immune cells have not been much in use for investigation of food bioactive components, despite several methodological reports on such systems. This might indicate that the system gives inconsistent data or has a low level of reproducibility. Some of the co-culture systems described here combine the use of cell lines and primary cells. The cells lines will represent a stable and reproducible system with low variability from experiment to experiment. However, the response of PBMCs and monocyte derived DCs will depend on the donor and can lead to increased variability between experiments.
The Caco-2/Raji B cell model is a simplified model that induces cells with M cell-like morphology. A gene expression study comparing the Caco-2/Raji model with human FAE suggest that the Caco-2 differentiation model is associated with some functional features of M cells. However the genes induced reflect the acquisition of a more general FAE phenotype (Lo et al. 2004). However, this is an easy accessible in vitro model system to study M cell-mediated translocation.
4 General Protocol
4.1 Co-culture of Caco-2/Human Monocyte Derived DCs (Include Contact Dependent Events)
In this co-cultivation system, the cells will be able to make direct cell–cell contacts between the immune cells and the basolateral side of the epithelial cells. Co-cultivation of the cells in this way makes it possible to investigate effects of both direct interactions between DCs and epithelial cells and communication by soluble factors.
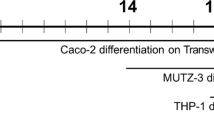
Caco-2 cells are seeded on the backside of filter inserts with 3 μm pores. 3 × 105 Caco-2 cells in 100 μL RPMI 1640 with 10 % FCS, are placed on the bottom side of the inserts placed in an inverted orientation. After overnight incubation, to allow attachment of the Caco-2 cells, the inserts are transferred to their normal orientation in a 24 well plate. The Caco-2 cells now face the lower chamber and are cultured as this for the next 14 days. Every other day the medium has to be exchanged with fresh RPMI 1640 with 10 % FCS. At day 14 add 1 × 106 CD14+ human monocytes in 500 μL RPMI 1640 with 10 % FCS, 25 ng/mL IL-4 and 50 ng/mL GM-CSF. At day 4 of co-cultivation remove the medium in the upper chamber and replace it with 500 μL RPMI 1640 with 25 ng/mL IL-4 and 50 ng/mL GM-CSF. The RPMI 1640 in the lower chamber should be exchanged every other day. Maintain the co-culture for 6 days before the experiments are conducted. Depending on the test compound, it could be added to either compartment. The lower chamber will reflect the apical side and the upper chamber the basolateral side of the intestinal epithelium. Incubation time for the test compound will depend on the type of compound and the read out system to be used and will have to be decided for each compound.
4.2 Caco-2/Human Monocyte Derived DCs (Soluble Factors)
In comparison with the above mentioned system co-cultivation of cells in this way will only make it possible to study effect of communication caused by soluble factors. The epithelial cells and the DCs will not be able to be in direct contact with each other and communication can therefore only happen by soluble factors diffusing through the filter.
300 μL of a 1 × 106/mL Caco-2 cells suspended in RPMI 1640 with 10 % FCS, are seeded on filter inserts with 0.4 μm pores. The cells are maintained at 37 °C in 5 % CO2 atmosphere for 14 days. The medium in both compartments should be exchanged with fresh RPMI 1640 with 10 % FCS, every other day. At day 14, 1 × 106 CD14+ human monocytes in RPMI-1640 with 10 % FCS, 25 ng/mL IL-4 and 50 ng/mL GM-CSF are added to the lower chamber, facing the basolateral side of the epithelial monolayer. The medium in the lower chamber is exchanged with fresh RPMI 1640 with 10 % FCS, 25 ng/mL IL-4 and 50 ng/mL GM-CSF at day 4 of the co-cultivation. Maintain the co-culture for 6 days before experiments are conducted. Continue to exchange the medium in the upper chamber with fresh RPMI 1640 with 10 % FCS every second day.
The upper chamber will reflect the apical side and the lower chamber the basolateral side of the intestinal epithelium. Incubation time for the test compound will depend on type of compound and the read out system to be used and will have to be decided for each compound.
4.3 Caco-2/THP-1 (Soluble Factors)
300 μL of a 1 × 106/mL Caco-2 cells suspended in RPMI 1640 with 10 % FCS, are seeded on filter inserts with 0.4 μm pores. The cells are maintained at 37 °C in 5 % CO2 atmosphere for 21 days, exchanging the medium with fresh RPMI 1640 containing 10 % FSC every other day. At day 18 the THP-1 cells are plated at a density of 2 × 105 cells/mL in RPMI 1640 with 10 % FCS, 200 ng/mL IL-4, 100 ng/mL GM-CSF, 20 ng/mL TNF-α and 200 ng/mL ionomycin in a 24 well plate for 3 days. At day 21 the medium of the THP-1 cells is exchanged with RPMI 1640 with 10 % FCS and the filter inserts with polarized Caco-2 monolayers are placed into the wells together with the differentiated THP-1 cells.
The upper chamber will reflect the apical side and the lower chamber the basolateral side of the intestinal epithelium. Incubation time for the test compound will depend on type of compound and the read out system to be used and will have to be decided for each compound.
4.4 Caco-2/PBMCs (Soluble Factors)
300 μL of a 1 × 106/mL Caco-2 cells suspended in RPMI-1640 with 10 % FCS are seeded on filter inserts with 0.4 μm pores. The cells are maintained at 37 °C in 5 % CO2 atmosphere for 21 days with exchanges of the medium with fresh RPMI 1640 with 10 % FSC every other day. At day 20, PBMCs are plated in the basolateral compartment at a density of 1–2 × 106 cells/mL RPMI-1640 with 10 % FCS. PBMCs can be activated by anti-CD3/CD28, mitogenic lectins or LPS (see Chap. 15 PBMCs).
The upper chamber will reflect the apical side and the lower chamber the basolateral side of the intestinal epithelium. Incubation time for the test compound will depend on type of compound and the read out system to be used and will have to be decided for each compound.
4.5 Caco-2/B Cells
300 μL of a 1 × 106/mL Caco-2 cells suspended in RPMI-1640 with 10 % FCS are seeded on filter inserts with 3.0 μm pores. The inserts are placed in a 24 well plate with 1 mL of RPMI 1640 with 10 % FCS. The cells are maintained at 37 °C in 5 % CO2 atmosphere for 14 days with exchanges of the medium with fresh RPMI 1640 with 10 % FSC, every other day. At day 14, 1 × 106 Raji cells in RPMI 1640 with 10 % FCS are added to the lower chamber, facing the basolateral side of the epithelial monolayer. Maintain the co-culture for 6 days. Continue to exchange the medium in the upper chamber with fresh RPMI 1640 with 10 % FCS every second day.
The upper chamber will reflect the apical side and the lower chamber the basolateral side of the intestinal epithelium. To investigate uptake through M-cells the test compound should be added to the apical side. The incubation time for the test compound must be investigated different compounds, but are usually in the range of minutes to a few hours.
In all of the above-mentioned systems, the effects of test compounds on co-cultures should be compared with effects on matched filters with Caco-2 monocultures. This will make it possible to identify whether the effects of the test compounds are related to cell–cell communication in the co-culture systems.
5 Assess Viability
The integrity of the Caco-2 monolayers should always be checked by measurement of TEER values, and filters with a TEER value below 300 Ω cm2 should not be used for further experiments.
The paracellular transport marker Lucifer Yellow (LY) is used to measure changes in the integrity of the Caco-2 cell layer. LY is added to the apical compartment at a concentration of 100 μM, in control wells and wells treated with test compound. The fluorescence of the LY transported to the basolateral side is then measured with a fluorescence microplate reader (Calatayud et al. 2012, and Chap. 10).
In comparison with polarized Caco-2 cells M-cells will lack microvilli on their apical surface which can be inspected by transmission electron microscope (TEM) or scanning electron microscope (SEM). Staining of the Caco-2 monolayer with fluorochrome-labeled Wheat Germ Agglutinin (WGA) or phalloidin (actin label) will give a continuous staining in a monoculture of Caco-2, however after co-cultivation of Caco-2 cells with Raji, M-cells are identified as a discontinuations in the staining pattern. This is due to the absence of microvilli. Corresponding monocultures of Caco-2 cells on matched filter inserts should be used as controls.
Lactate dehydrogenase (LDH) is also a marker of cytotoxicity and its release can be determined in the supernatant of the cell cultures (Decker and Lohmann-Matthes 1988). There are several commercial kits for determination of LDH concentration in culture supernatants.
6 Experimental Readout
Several methodological reports are describing co-culture conditions between Caco-2 and CD14+ monocyte derived DCs, THP-1 or PBMCs (Araujo and Sarmento 2013; Leonard et al. 2010; Schimpel et al. 2014; Antunes et al. 2013; Kerneis et al. 2000; Parlesak et al. 2004). However, there are still few reports where these co-culture systems have been used to scrutinize communication between epithelial cells and immune cells.
Apical TLR ligation on intestinal epithelial cells and its effect on PBMCs have been studied by co-cultivation of HT-29 and PBMCs (de Kivit et al. 2011). The TLR ligands are added to the apical side of the epithelial monolayer, whilst the PBMCs are stimulated with anti-CD3/CD28 antibodies and LPS. The release of cytokines to the culture supernatants, both in the basolateral and apical compartments, were investigated by ELISA. There is also a report on the effect of the anti-inflammatory drugs ibuprofen and prednisolone in a co-culture system of Caco-2 monolayers in co-culture with whole blood (Schmohl et al. 2012).
There is one report using a co-culture model of Caco-2 monolayers and THP-1 cells to study the function of the immediate-early response gene 1 (IEX-1) (Ishimoto et al. 2011). Overexpression or knock down of the IEX-1 gene in Caco-2 cells were induced by Lentivirus infection. Caco-2 cells were differentiated for 14 days on semi-permeable filter inserts. The THP-1 cells were differentiated for 4 days and then the filter inserts with Caco-2 were placed in the well with differentiated THP-1 cells. After co-cultivation, mRNA was extracted from both Caco-2 cells and THP-1. Changes in expression of genes coding for cytokines were investigated by qPCR.
The Caco-2/B cell model is widely used to study translocation of bacteria over M-cells. Translocation of fluorescence-labelled carboxylated latex or dextran particles was used to confirm in vitro induction of the M-cell like phenotype. Translocation of bacteria was measured by serial dilutions of basolateral culture medium on agar plates 2 h after addition of bacteria to the apical compartment (Finn et al. 2014). How dietary components affect translocation of bacteria over M-cells have also been investigated (Roberts et al. 2013). The test compound was added to the apical compartment for 30 min before infection with bacteria. After 2–4 h, the basolateral medium was harvested and bacteria enumerated following serial dilution on agar plates.
7 Advantages, Disadvantages and Limitations
These co-culture systems give the opportunity to study cell–cell communication, and most cell laboratories will have the possibility to establish at least some of these co-cultivation systems. The use of monocyte-derived DCs or PBMCs have the advantage of responding well with the production of cytokines, whilst THP-1 on the other side produce a limited number of cytokines. These protocols are time-consuming and therefore not suitable for high throughput screening. As for other in vitro culturing systems the data should be interpreted with care as they do not represent the complexity found in the in vivo situation.
8 Conclusions
The co-cultivation systems described here needs to be carefully validated in order to be able to harvest reliable experimental data related to communication between immune cells and epithelial cells. They can be used for screening purposes, however, due to time consumption they are generally unsuitable for high throughput screening. Co-culture of Caco-2 and Raji cells generates cells with a M-cell like phenotype. Such cells can give useful information regarding transport of test compounds through M-cells, however such studies will eventually have to be confirmed in an in vivo system.
References
Antunes F, Andrade F, Araujo F et al (2013) Establishment of a triple co-culture in vitro cell models to study intestinal absorption of peptide drugs. Eur J Pharm Biopharm 83:427–435
Araujo F, Sarmento B (2013) Towards the characterization of an in vitro triple co-culture intestine cell model for permeability studies. Int J Pharm 458:128–134
Artis D (2008) Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol 8:411–420
Calatayud M, Vazquez M, Devesa V et al (2012) In vitro study of intestinal transport of inorganic and methylated arsenic species by Caco-2/HT29-MTX cocultures. Chem Res Toxicol 25:2654–2662
Coombes JL, Powrie F (2008) Dendritic cells in intestinal immune regulation. Nat Rev Immunol 8:435–446
de Kivit S, van Hoffen E, Korthagen N et al (2011) Apical TLR ligation of intestinal epithelial cells drives a Th1-polarized regulatory or inflammatory type effector response in vitro. Immunobiology 216:518–527
Decker T, Lohmann-Matthes ML (1988) A quick and simple method for the quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis factor (TNF) activity. J Immunol Methods 115:61–69
Finn R, Ahmad T, Coffey ET et al (2014) Translocation of Vibrio parahaemolyticus across an in vitro M cell model. FEMS Microbiol Lett 350:65–71
Gullberg E, Leonard M, Karlsson J et al (2000) Expression of specific markers and particle transport in a new human intestinal M-cell model. Biochem Biophys Res Commun 279:808–813
Ishimoto Y, Satsu H, Totsuka M et al (2011) IEX-1 suppresses apoptotic damage in human intestinal epithelial Caco-2 cells induced by co-culturing with macrophage-like THP-1 cells. Biosci Rep 31:345–351
Kerneis S, Bogdanova A, Kraehenbuhl JP et al (1997) Conversion by Peyer’s patch lymphocytes of human enterocytes into M cells that transport bacteria. Science 277:949–952
Kerneis S, Caliot E, Stubbe H et al (2000) Molecular studies of the intestinal mucosal barrier physiopathology using cocultures of epithelial and immune cells: a technical update. Microbes Infect 2:1119–1124
Leonard F, Collnot EM, Lehr CM (2010) A three-dimensional coculture of enterocytes, monocytes and dendritic cells to model inflamed intestinal mucosa in vitro. Mol Pharm 7:2103–2119
Lo D, Tynan W, Dickerson J et al (2004) Cell culture modeling of specialized tissue: identification of genes expressed specifically by follicle-associated epithelium of Peyer’s patch by expression profiling of Caco-2/Raji co-cultures. Int Immunol 16:91–99
Miller JC, Brown BD, Shay T et al (2012) Deciphering the transcriptional network of the dendritic cell lineage. Nat Immunol 13:888–899
Mora JR, Bono MR, Manjunath N et al (2003) Selective imprinting of gut-homing T cells by Peyer’s patch dendritic cells. Nature 424:88–93
Mowat AM (2003) Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev Immunol 3:331–341
Parlesak A, Haller D, Brinz S et al (2004) Modulation of cytokine release by differentiated CACO-2 cells in a compartmentalized coculture model with mononuclear leucocytes and nonpathogenic bacteria. Scand J Immunol 60:477–485
Pastorelli L, De Salvo C, Mercado JR et al (2013) Central role of the gut epithelial barrier in the pathogenesis of chronic intestinal inflammation: lessons learned from animal models and human genetics. Front Immunol 4:280
Rimoldi M, Chieppa M, Vulcano M et al (2004) Intestinal epithelial cells control dendritic cell function. Ann N Y Acad Sci 1029:66–74
Roberts CL, Keita AV, Parsons BN et al (2013) Soluble plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J Nutr Biochem 24:97–103
Schimpel C, Teubl B, Absenger M et al (2014) Development of an advanced intestinal in vitro triple culture permeability model to study transport of nanoparticles. Mol Pharm 11(3):808–818
Schmohl M, Schneiderhan-Marra N, Baur N et al (2012) Characterization of immunologically active drugs in a novel organotypic co-culture model of the human gut and whole blood. Int Immunopharmacol 14:722–728
Segura E, Touzot M, Bohineust A et al (2013) Human inflammatory dendritic cells induce Th17 cell differentiation. Immunity 38:336–348
Stagg AJ, Kamm MA, Knight SC (2002) Intestinal dendritic cells increase T cell expression of alpha4beta7 integrin. Eur J Immunol 32:1445–1454
Steinman RM (2007) Dendritic cells: understanding immunogenicity. Eur J Immunol 37(Suppl 1):S53–S60
Steinman RM (2012) Decisions about dendritic cells: past, present, and future. Annu Rev Immunol 30:1–22
Steinman RM, Idoyaga J (2010) Features of the dendritic cell lineage. Immunol Rev 234:5–17
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this chapter
Cite this chapter
Kleiveland, C.R. (2015). Co-culture Caco-2/Immune Cells. In: Verhoeckx, K., et al. The Impact of Food Bioactives on Health. Springer, Cham. https://doi.org/10.1007/978-3-319-16104-4_18
Download citation
DOI: https://doi.org/10.1007/978-3-319-16104-4_18
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-15791-7
Online ISBN: 978-3-319-16104-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)