
Abstract
The increasing effects of climate and global change oblige ecosystem-based management to adapt forestry practices to deal with uncertainties. Here we provide an overview to identify the challenges facing the boreal forest under projected future change, including altered natural disturbance regimes, biodiversity loss, increased forest fragmentation, the rapid loss of old-growth forests, and the need to develop novel silvicultural approaches. We specifically address subjects previously lacking from the ecosystem-based management framework, e.g., Indigenous communities, social concerns, ecological restoration, and impacts on aquatic ecosystems. We conclude by providing recommendations for ensuring the successful long-term management of the boreal biome facing climate change.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others
1 How Did We Get Here? A Perspective on Boreal Forest Management
Ecosystem degradation has intensified because of increased human pressure on natural systems worldwide (Foley et al., 2005; Rands et al., 2010). During the twentieth century, the world’s population increased from 1.6 to 7.7 billion people (Lutz et al., 2004), resulting in a greater demand for natural resources to meet the needs of this expanding population. Technological advances have increased our efficiency in exploiting ecosystems; humans now alter the environment faster and at a greater scale than ever before (Boserup, 1981; Puettmann et al., 2009). Nonetheless, societies depend on finite natural resources and ecosystem services (Perrow & Davy, 2002). The main activities causing ecosystem impacts are agriculture, industry, forestry, and urbanization, and this economic development has therefore led to the alteration of the original ecosystems across a large portion of the planet. This loss of an ecological–economic equilibrium has led to a need to further develop and apply the concept of sustainable development as a means of balancing resource exploitation, biological conservation, and social conditions for future generations (Quarrie, 1992; Rockström et al., 2009b; Steffen et al., 2015).
Forests account for 31% of the world’s land area (FAO, 2016), and forest resources are vital to the development of human societies (FAO, 2014). At least 18% of the world’s population uses wood to build their homes, 2.4 billion people cook by burning woody materials, and 90 million people in Europe and the United States use wood as an energy source for domestic heating. The boreal forest is the second-largest terrestrial biome in the world (Teodoru et al., 2009), covering 14 million km2, distributed in a circumpolar forest belt (Burton et al., 2003), and representing about 25% of the world’s forest (Dunn et al., 2007).
Currently, two-thirds of this biome is managed mainly for timber production (Gauthier et al., 2015b). Boreal forests are critical for the global wood supply, producing 37% of the world’s wood (Gauthier et al., 2015b). During the last century, forest management practices had timber production as their main goal. Logging activities to meet the demand for timber significantly affect this biome (Halme et al., 2013; Kuuluvainen & Siitonen, 2013; Messier et al., 2013; Puettmann et al., 2009). From 1990 to 2000, Canada recorded the most intense period of logging operations in the world, with forests harvested at more than two hectares per minute (Perrow & Davy, 2002).
The current global demand for wood is 1.5 billion m3, whereas it is expected to increase to between 2.3 and 3.5 billion m3 by 2050 (Smeets & Faaij, 2007). Thus, logging and related activities will likely have an ever-greater impact on the boreal forest in the near future, continuing the twentieth-century trend of expanded exploitation (Park & Wilson, 2007). For example, in Québec (Canada), the total volume of wood harvested over the past century increased steadily until 2005. In 1924, it was 13.9 million m3, rising to 21.9 million m3 in 2011. It should be noted that between 1997 and 2005, the volume harvested was more than 40 million m3, reaching its peak in 2005 at 45.64 million m3 (Duchesne & Ouimet, 2007; National Research Council, 2016). In the intensively managed boreal forests of Finland and Sweden, annual growth and harvesting have been increasing in the past 100 years. In the recent past (averaged over 2013–2017), an average of 68.3 and 82.8 million m3 were harvested, corresponding to 75% and 78% of the annual growth in these two countries (Korhonen et al., 2021; SLU, 2020). In Sweden, 20 of 28 million ha of forest is accessible for intensive forestry; therefore, these forested areas are currently on a transformation trajectory away from natural and resilient ecosystem conditions having multiple value chains (Angelstam et al., 2020).
The reduction, modification, and loss of forests are not recent phenomena; these human-related alterations to forest ecosystems trace the evolution and migration of human populations. Human activity is one of the key processes in the history of forest land transformation (Williams, 2003). The net loss in the global forest area between 2000 and 2010 was 5.2 million ha/yr (roughly the size of Costa Rica). This loss was 8.3 million ha/yr between 1990 and 2000 (FAO, 2011). However, the State of Canada’s Forests report (Natural Resources Canada, 2020) maintains that changes in forest area caused by deforestation are not significant in Canada, as it would take 40 years for Canada to lose 1% of its forest area under the most pessimistic forest harvesting scenario (Guindon et al., 2018). In Fennoscandian forests, deforestation due to forestry is prevented by legislation, i.e., clear-cut harvesting must be followed by forest regeneration; the main sustainability issues are related more to loss of biodiversity, ecosystem services, multiple value chains, and Indigenous and local cultures and less to sustained yield or deforestation.
Regardless of the region, climate change intensifies threats to forest health (Trumbore et al., 2015). The intensity and frequency of forest fires have increased in both Canada and the United States, exacerbated by prolonged drought episodes (attributed to climate change) and fire-suppression policies that have increased the amount of available fuel loads (FAO, 2009). From the projected cumulative impacts of fire, drought, and insects on timber volumes across North American boreal forest, the current level of harvesting could thus be difficult to maintain without implementing of adaptative measures (Boucher et al., 2018). In the Fennoscandian forests, the past decade has seen several years of exceptional forest fires, storms, and insect outbreaks, particularly in Sweden (Hlásny et al., 2021; Krikken et al., 2021; Valinger & Fridman, 2011). These events raise questions in regard to the vulnerability of a ubiquitous simplified forest management system—and also from a sustainable timber yield viewpoint—and advocate for the application of more diverse, ecosystem- and disturbance-based management perspectives (Berglund & Kuuluvainen, 2021). Droughts, insect outbreaks, and windstorms are particularly problematic for Norway spruce, which is favored as a commercial tree species in both countries, partly due to the extremely high ungulate browsing pressure on deciduous trees and pines in some parts of the region. The uncertainty associated with disturbances and their potential trajectories in future climates requires a profound reflection on the challenges faced by the boreal biome to achieve sustainable forest management in terms of wood material supply, biodiversity conservation, maintenance, and enhancement of forest carbon sinks, and the cultural values of forests.
Forestry activities in the recent decades have contributed to a decline in habitat diversity and productivity of forest ecosystems around the world, a phenomenon accelerated by climate change (Fischer & Lindenmayer, 2007; Lindenmayer & Fischer, 2007; Schütz, 1997). With increased social concerns about protecting biodiversity (Franklin et al., 2002), boreal forestry has begun to address goods and services other than timber production (Dobson et al., 1997; Gauthier et al., 2009; Halme et al., 2013; Kuuluvainen, 2002; Puettmann et al., 2009). The preservation of biodiversity and the modification of forestry practices to reduce their impact on ecosystems have emerged as two key issues in forest management (FAO, 2009; Lindenmayer & Franklin, 2002; Myers et al., 2000). These concerns confront traditional forest management, which focuses on a deterministic planning of harvesting and exploitation without considering changes, natural disturbances, social issues, uncertainty, and nonlinearity. Forest ecosystem-based management (FEM), in contrast, aims to bridge the gap between natural and managed forests to maintain the ecological integrity and biodiversity of ecosystems. FEM was specifically defined as
a management approach that aims to maintain healthy and resilient ecosystems by reducing the gaps between natural and managed landscapes to ensure, in the long term, the maintenance of multiple ecosystem functions and, consequently, to maintain the social and economic benefits derived from them (Gauthier et al., 2009).
This approach stems from the reflections on sustainable forest development, which emerged from the 1992 Earth Summit in Rio de Janeiro, and FEM has become increasingly applied within the boreal biome, especially in North America (Burton et al., 2003; Mitchell & Beese, 2002). FEM applies an ecosystem model to reconcile timber harvesting with the long-term maintenance of the structure, functioning, and ecological processes responsible for maintaining ecosystem services. This approach manages the forest through a holistic view (Kimmins, 1997) to ensure its integrity, biodiversity, and sustainability (Gauthier et al., 2009). However, the question arises: Is FEM a useful framework to deal with climate change and the associated impacts?
Our new definition considers FEM as
an adaptative management approach that aims to promote healthy and resilient forests under climate change to ensure the long-term maintenance of ecosystem functions and thereby retain the social and economic benefits they provide to society.
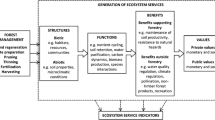
Thus, FEM is a promising solution for achieving sustainable forest management within a context of climate change, an approach able to include responses and solutions for all the challenges facing the boreal biome (Grenon et al., 2010). In this critical moment, scientific cooperation is essential to adapt forest management practices for the future. FEM within the boreal forest provides one of the last remaining global opportunities to proactively plan forest management for sustainable ecosystem and economic development. In this final chapter, we present the most important challenges facing the future boreal forest (Fig. 31.1). Our goal is to provide helpful recommendations and tools to reduce uncertainty and to justify how FEM can address future challenges within the second-largest terrestrial ecosystem in the world, the boreal biome.

The challenges facing boreal forests under global climate change
2 The Challenges of Sustainable Management in Boreal Forests Facing Climate Change
The need to adapt FEM to the future and develop measures to achieve sustainable forest management in the face of climate change led to a discussion among 147 researchers in forest sciences to build this book and produce a list outlining the challenges facing the boreal biome in terms of natural disturbances, silvicultural practices, biodiversity, landscape ecology, economy, and society (Fig. 31.1). In this section, we outline and assess the current state as a starting point to establish future research directions and applications.
2.1 Natural Disturbance Regime Change
Boreal forests are affected by various natural disturbances, including wildfires, weather-related disturbances (heat, drought, snow, and wind), insect outbreaks, and disease (Gauthier et al., 2015b; Price et al., 2013). These disturbances operate over a wide range of spatial and temporal scales and are among the core factors driving landscape dynamics and the structure, composition, and biodiversity in these forests (Berglund & Kuuluvainen, 2021; Price et al., 2013; Shorohova et al., 2011). Climate change can impact these forests by modifying the timing, extent, and severity of the natural disturbance regimes (Navarro et al., 2018a; Seidl et al., 2017). Thus, the most serious challenge will be to adapt FEM to this new reality. Here we describe and discuss the main observed and expected climate-driven changes in natural disturbance regimes.
2.1.1 Wildfires
Wildfires constitute a major natural disturbance in boreal forests, a natural process required to maintain the biodiversity and dynamics associated with these forests. Throughout the circumboreal biome, between 9 and 20 million ha of forest burn annually (Robinne et al., 2018) with considerable spatial and temporal variability (De Groot et al., 2013a; Gauthier et al., 2015a; Ryan, 2002). The North American boreal forest, for example, is characterized by relatively infrequent, high-intensity, stand-replacing crown fires that often completely burn extensive patches of forest. In contrast, the Eurasian boreal region experiences repeated low- to moderate-intensity surface fires characterized by low tree mortality (De Groot et al., 2013a; Robinne et al., 2018; Rogers et al., 2015). Human-caused fires occur mostly in areas where they are likely to be detected; this permits a rapid response from fire-suppression agencies, to often limit fire spread. Although humans cause a large proportion of fires in the boreal forest, lightning-caused fires account for most of the area burned in a given season (Robinne et al., 2018; Stocks et al., 2003).
Natural fire ignition and propagation depend on a combination of factors related to, on one hand, climate and weather (Eden et al., 2020; Seidl et al., 2017) and, on the other hand, fuel type and availability (De Groot et al., 2013a; Rogers et al., 2015). Consequently, ongoing climate change is expected to markedly alter future fire regimes. Fire weather could become more severe in the coming years (De Groot et al., 2013b; Flannigan et al., 2016), leading to drier fuels and favoring easier fire ignition and propagation (Flannigan et al., 2016). Despite uncertainties related to differences in climate change scenarios, fire weather is expected to be more severe in western Canada than in central Russia (De Groot et al., 2013b). Nevertheless, Russia is also expected to experience a marked increase in fire activity (De Groot et al., 2013b) and possibly a substantial increase in stand-replacing fires (Gauthier et al., 2015b). Although the fuel consumption rate will be higher in boreal stands in Canada, total carbon emissions could be higher from Russian boreal fires owing to a larger annually burned area (De Groot et al., 2013b). Longer and more active fire seasons will significantly affect boreal forests. Forest composition would shift toward an increased proportion of fire-tolerant and fire-resistant species (De Groot et al., 2013b). The amount of old-growth forest, associated with high biodiversity, would be greatly reduced to give way to landscapes dominated by young forests (Kuuluvainen & Gauthier, 2018). This phenomenon—resulting in fewer mature trees across the landscape because of the repeated occurrence of fires—increases the risks of regeneration failure, thereby leading to a gradual opening of forests (Jasinski & Payette, 2005; Kuuluvainen & Gauthier, 2018). For example, Splawinski et al. (2019) projected a progressive increase in the area affected by natural regeneration failure under climate change for northern Québec, culminating with a 65.8% loss under the worst-case scenario.
The global carbon cycle would also be affected as larger, more frequent, and more severe wildfires release higher levels of carbon into the atmosphere. Boreal forests therefore risk shifting from being carbon sinks to carbon sources (Walker et al., 2019), thereby amplifying this positive climate feedback.
2.1.2 Weather- and Climate-Related Disturbances: Heat, Drought, Wind, Floods, and Snow
Boreal forests are expected to experience large increases in temperature over the twenty-first century, accompanied by modest increases in precipitation in some regions (IPCC, 2014). These changes will lead to higher frequencies and intensities of extreme heat and drought events (Price et al., 2013). Heat- and drought-induced tree mortality has already increased over the last two decades (Allen et al., 2010). This phenomenon will likely be further exacerbated in the twenty-first century in the boreal biome (Gauthier et al., 2015b; Liu et al., 2013). Forest sensitivity to heat and drought events depends on such factors as the intensity and frequency of these events and the tolerance of tree species to heat/drought. Drought-intolerant aspen-dominated forests in western Canada, for example, have experienced very severe drought-induced diebacks at levels similar to postfire mortality (Michaelian et al., 2011). Overall, the driest regions of the boreal biome have been shown to be more sensitive to weather-induced diebacks (Gauthier et al., 2015b). Western Canadian boreal forests associated with a drier climate are already experiencing increased mortality rates (Boucher et al., 2018; Peng et al., 2011), whereas, at least for now, the moister forests of eastern Canada have been less affected (D’Orangeville et al., 2018).
Wind and snow, common disturbances in boreal forests, cause the uprooting and breakage of trees (Lavoie et al., 2019; Mitchell, 2013; Montoro Girona et al., 2019; Saad et al., 2017; Valinger & Fridman, 2011). These damages can rapidly alter forest structure, species composition, and the spatial and temporal availability of resources, in turn disrupting forest management and planning. Boreal forests have been recurrently affected by severe storms in the past, such as the Gudrun storm in 2005 in northern Europe and the Great Ice Storm of 1998 in eastern Canada. It remains unclear how storm regimes will be affected by climate change (Feser et al., 2015; Mölter et al., 2016); however, it is expected that increasing temperatures will favor an increased frequency and intensity of winds and greater snow loads (Gregow et al., 2011). Warmer winters will lead to shorter periods with frozen soil and greater loads of heavy humid snow; this combination heightens the likelihood of trees being uprooted and suffering stem breakage (Nykänen et al., 1997; Peltola et al., 1999). Furthermore, expected changes in tropical cyclone regimes could also increase windthrow impacts on boreal forests, as observed during the Sandy and Ophelia storm events reaching, respectively, Canadian forests in 2012 and Norwegian forests in 2017. Riparian forests may also be affected by an increase in flooding as unprecedented low and high spring discharge in recent decades—relative to the historical natural variability of the last 250 years—also suggests that the increase in flood frequency and magnitude originates from climate change (Nolin et al., 2021).
2.1.3 Insect Outbreaks and Diseases
Biotic agents, such as native or non-native insects and pathogens, constitute major disturbances in boreal forests. The most damaging insects for boreal tree species are defoliators, which eat leaves or needles, and bark beetles, which feed on phloem and cambium (MacLean, 2016). On the other hand, pathogens cause significant damage to all tree parts, i.e., foliage, stem, and roots, leading to reduced photosynthetic activity and water/nutrient uptake and producing structural problems (Malmström & Raffa, 2000; Natural Resources Canada, 2020). Climate and weather conditions affect the distributions and ecological dynamics of insects and pathogens and those of their hosts (Dukes et al., 2009; Malmström & Raffa, 2000). Although the life cycle of insects and pathogens responds mainly to temperature, pathogens are also sensitive to precipitation and humidity (Price et al., 2013). Pathogen-induced diseases reduce growth and productivity (Price et al., 2013) and cause widespread forest decline and mortality when co-occurring with other disturbance agents (Dukes et al., 2009). Insect outbreaks markedly impact forest productivity and dynamics by affecting tree growth, seed production, tree regeneration, and successional processes. Outbreaks are cyclic and often synchronous over large geographic areas; this leads to a region-wide mortality of host trees in a relatively short period. Spruce budworm (Choristoneura fumiferana) is the main defoliator of spruce and fir forests in North America, affecting extensive areas and causing important losses of timber supplies (Montoro Girona et al., 2018b; Régnière et al., 2012). In Eurasia, there are no comparable records of large-scale outbreaks of defoliators as that of the spruce budworm in North America. Although insect outbreaks generally have less severe punctual impacts on forest productivity and dynamics than fires, they often affect larger areas. For example, the Canadian Forest Service calculated that insects affected 15.6 million ha of Canadian forest in 2017, whereas 3.4 million ha of forests burned that same year (Natural Resources Canada, 2020).
Defoliating insect outbreaks have shown an increase in severity, extent, and duration over the recent decades, and their frequency and severity are expected to increase further (Navarro et al., 2018c; Zhang et al., 2014). Climate change will likely modify the geographic distribution of host trees within boreal forests and may alter both the range and outbreak potential of their associated insects and pathogens (Malmström & Raffa, 2000). Models project that future warmer winters and longer growing seasons will favor the northward expansion of the northern range limits of many insect pests (Dukes et al., 2009; Pureswaran et al., 2015; Régnière et al., 2012). Spruce budworm in eastern Canadian boreal forests experienced such a northward shift during the twentieth century (Navarro et al., 2018c). Similarly, an expansion in climatically suitable habitats at the beginning of the twenty-first century for the mountain pine beetle has facilitated the northward and higher-elevation expansion of outbreaks in western Canadian boreal forests (Kurz et al., 2008). Projections of climate change impacts on pathogen populations remain uncertain, as pathogen outbreaks are less predictable than insect outbreaks given the links of the former to precipitation levels (Dukes et al., 2009; Pautasso et al., 2015; Price et al., 2013). Researchers do agree, however, that pathogen activity in circumboreal forests will likely increase (Price et al., 2013).
2.1.4 Interactions Between Natural Disturbances
Interactions between natural disturbances are common across the boreal biome. The most frequently reported interaction is the increased flammability of forests induced by drought events, which enhances the frequency and severity of fires (Flannigan et al., 2016). Interactions between biotic, i.e., insect outbreaks and pathogens, and abiotic disturbances are also very frequent and are critical to the dynamics of biotic disturbance agents (Canelles et al., 2021; Nolin et al., 2021; Seidl et al., 2017). For example, the large number of dead trees resulting from drought, fire, or windfall can trigger a strong increase in insect populations and amplify the spread and intensity of outbreaks (Marini et al., 2013; Price et al., 2013). On the other hand, stands affected by insect outbreaks significantly increase the amount of flammable fuel and, therefore, a fire's potential spread and severity (James et al., 2017; Perrakis et al., 2014). Most interactions between disturbances tend to amplify the mutual effects of disturbance agents (Seidl et al., 2017). Outbreak severity may vary with forest composition at the landscape level (Lavoie et al., 2021). The long fire cycle may favor increasing outbreak severity stemming from the abundance of late-successional host trees (Bergeron & Leduc, 1998; Navarro et al., 2018c). On the contrary, a short fire cycle may increase the abundance of broadleaf stands and favor a better control by natural enemies (Cappuccino et al., 1998). Shifts between short and long fire cycles explain the dynamics of defoliators through the Holocene (see Chap. 2). The observations are of critical concern, as climate-induced increases in natural disturbances may be further intensified by such interactions, heightening the risk of exceeding ecological thresholds and tipping points.
2.2 Biodiversity Loss
Boreal forest landscapes have transformed rapidly over the last decades (Mori et al., 2021). In Fennoscandia, extensive changes have occurred since the 1950s when forestry methods became more mechanized and efficient, and clear-cutting was introduced, i.e., even-aged management (Esseen et al., 1997). Moreover, forestry and fire suppression have led to the disappearance of natural disturbances, replaced instead by anthropogenic disturbances, including thinning, clear-cutting, soil scarification, and the planting of conifers mostly in monospecific regimes (Esseen et al., 1997; Wallenius, 2011). Unlike forests structured by natural dynamics, managed forests often consist of even-aged monocultures that lack structural complexity, including an absence of coarse woody debris, snags, and old trees (Bengtsson et al., 2000; Löfman & Kouki, 2001). In North American boreal forest regions, forest composition and age structure are predicted to change over the twenty-first century because of climate change and increased anthropogenic pressure (Boulanger et al., 2016). These changes in forest composition and structures will have/have had serious consequences for the many species that rely on deciduous trees, deadwood, large-diameter trees, and complex horizontal and vertical structures of tree vegetation (Kuuluvainen, 2009; Ram et al., 2017; Regos et al., 2018; Virkkala, 2016).
In general, the structural complexity of habitats is strongly correlated with species richness for most taxonomic groups (Honnay et al., 2003; Lassau et al., 2005). In Fennoscandia, forest conditions remain determined by the transformation to even-aged, single-species conifer forests. This simplistic forest management has had severe consequences for forest biodiversity. In Finland, for example, the forest is the primary habitat for 31% of threatened species. For almost five-sixths of these threatened species, the primary driver of the population decrease is a change in forest habitat (Hyvärinen et al., 2019). This biodiversity loss continues to increase (Fig. 31.2). In Sweden, more than 50% of red-listed species are connected to forest habitats, 43% are dependent on forests, and 1,400 species are directly threatened by forest clear-cutting (Artdatabanken, 2020). In North American boreal forests, for a variety of species, it is predicted that populations will decrease, and their ranges, in terms of size and distribution, will shift through the loss of climate suitability and greater anthropogenic influence, i.e., habitat degradation and fragmentation (Cadieux et al., 2020; Woo-Durand et al., 2020).

The number of threatened species (bars) in Finland by phylum over time in forests (red) and other habitats (blue). Data obtained from the Natural Resources Institute Finland (2021)
Biodiversity plays a vital role in the functioning of forest ecosystems and is closely related to the health status of an ecosystem. Highly diverse systems are expected to be less prone to perturbations such as pest outbreaks. A healthier ecosystem provides higher-quality ecosystem services, and robust forest health correlates with forest productivity, thereby maximizing resource exploration in well-maintained forests (Bohn & Huth, 2017; Zhang et al., 2012). Ecosystems that deteriorate to an unsustainable level result in problems that are often very expensive, economically speaking, to reverse. Hence, the integration of biodiversity management as a target into current forestry practices becomes a priority. Over the last decades, forestry has drastically decreased legacies crucial for biodiversity. The challenge lies in bringing these critical components back into the boreal forest ecosystem without jeopardizing commercial forestry practices. Forests harbor multiple species, each with its own environmental and ecological requirements. This is particularly true for habitat specialists, which commonly require a precise range of environmental conditions or a specific diet. Forest structures and environmental conditions, e.g., moisture and light conditions, are important factors severely affected by forestry but are, nonetheless, critical for the occurrence of species from different taxonomic groups. The main challenge is to improve the overall habitat and landscape conditions that favor greater biodiversity. For example, deadwood quantity and quality, e.g., of varying decay stages, including standing and fallen trees, recently dead or decomposing stems, influence the occurrence of a variety of beetle species, polypores, and bird species. In Fennoscandia, managed forests harbor on average only 4 to 5 m3 of deadwood per hectare, whereas in natural forests, deadwood can exceed 100 m3 per hectare. Additionally, vertical structures, e.g., understory and uneven-aged forests, are important for food availability, nesting opportunities, and hiding spots for many boreal forest birds (Brokaw & Lent, 1999; Culbert et al., 2013; Eggers & Low, 2014).
The landscape structure plays an important role, as lichen and polypores are sensitive to habitat fragmentation and the lack of old-growth forests; therefore, large conservation areas benefit these species (Junninen & Komonen, 2011). For managed boreal forest landscapes to play a vital role in conserving biodiversity, these must include a mixture of habitats of varied successional stages, e.g., containing both early- and late-successional forest stages. Clear-cutting has long been held as an appropriate method for emulating natural disturbances (Mielikäinen & Hynynen, 2003). In naturally dynamic boreal forest systems, disturbances such as fire, insect outbreaks, and windfall contribute to high structural complexity; however, the ecological effects of clear-cutting differ from those of wildfire (Heikkala et al., 2016). Furthermore, forest dynamics in the boreal forest are also driven by various other natural, small-scale disturbance agents, like wind, pathogens, and insects, that have maintained a semicontinuous forest cover containing small gaps. The main challenge for forestry lies in reintroducing natural dynamics and restoring the natural systems while also minimizing any damage to forest production.
2.3 Loss of Old-Growth Forests
Old-growth forests are generally defined as stands at the end of forest succession, where post-disturbance cohorts are beginning to be replaced by new trees, human impacts are negligible, and low-severity disturbances are the primary drivers (Kneeshaw & Gauthier, 2003; Wirth et al., 2009). Specific structural attributes often distinguish these forests from younger stands, including a higher deadwood volume or a more complex structure (Kulha et al., 2019; Martin et al., 2018; Paillet et al., 2015; Wirth et al., 2009). Even within a given landscape, the concept of old-growth forest actually refers to a wide diversity of structures and composition that vary over time, depending on environmental conditions and the local disturbance history (Kulha et al., 2020; Martin et al., 2020a, d; Meigs et al., 2017; Portier et al., 2018). Old-growth forests hence typically consist of complex mosaics of uneven-aged stands. The long continuity of the forested state in these ecosystems is also vital for many disturbance-sensitive and low-dispersal species. For these reasons, old-growth forests provide a wide range of habitats, increase biodiversity, and provide numerous ecosystem services, such as carbon sequestration, water filtration, and cultural and aesthetic values (see Chap. 7; Keeton, 2018; Warren et al., 2018; Watson et al., 2018). Hence, old-growth forests are key elements of natural landscapes (Fig. 31.3). However, the climatic and fertility constraints in boreal landscapes may inhibit the development of old-growth attributes common to other biomes, such as very large trees or a complex vertical structure (Bergeron & Harper, 2009; Martin et al., 2020b, 2021b).

Old-growth boreal forests in eastern Canada are dominated by black spruce (Picea mariana (Mill.) BSP) and, to a lesser extent, balsam fir (Abies balsamea (L.) Mill.). These forests are defined by a strong heterogeneity of structures and microhabitats, shaped by specific natural disturbance histories and abiotic characteristics. The low diversity of tree species and their limited size may nevertheless make these stands appear—erroneously—as homogeneous. The difficulty in correctly identifying their heterogeneity may eventually lead to the disappearance of old-growth forests defined by distinctive functions and habitats not found elsewhere. Photo credits 1–9 Maxence Martin, 10 Frédéric L. Tremblay
The decreased area, diversity, connectivity, and functionality of boreal old-growth forests in managed landscapes represent major issues facing the boreal biome. Because of their remoteness and low productivity, boreal forests have long remained undisturbed by logging activities, particularly in the northern and eastern parts of Eurasia and in northern North America (Potapov et al., 2017; Venier et al., 2018; Wells et al., 2020). The development of industrial-scale forest management has nevertheless led to increased exploitation of these territories, especially since the mid-twentieth century (Boucher et al., 2017; Dupuis et al., 2020; Ostlund et al., 1997). Logging activities have therefore led to a loss of old-growth forest coverage (Cyr et al., 2009; Grondin et al., 2018; Ostlund et al., 1997), changes in tree-species composition (Boucher & Grondin, 2012; Boucher et al., 2015; Kuuluvainen et al., 2017), landscape homogenization and fragmentation (Haeussler & Kneeshaw, 2003; Löfman & Kouki, 2001; Schmiegelow & Mönkkönen, 2002), and decreased deadwood availability (Jonsson & Siitonen, 2012; Moussaoui et al., 2016). In certain regions, such as Fennoscandia, old-growth forests have almost completely disappeared and now represent a minimal part of the total forest cover (Forest Europe, 2015; Kuuluvainen et al., 2017; O’Brien et al., 2021; Potapov et al., 2017). Consequently, many species that depend on old-growth forests or associated elements, such as deadwood, are now threatened in the European boreal forests (Esseen et al., 1992; Jonsson & Siitonen, 2012; Tikkanen et al., 2006). In Canada and Russia, old-growth forests remain relatively abundant, but their areas are rapidly decreasing, and this is already causing major biodiversity issues (Aksenov et al., 1999; Bergeron et al., 2017; Cyr et al., 2009). For example, the level of fragmentation and degradation of old-growth forests in Canada has caused a collapse of woodland caribou (Rangifer tarandus caribou) populations (Venier et al., 2014). In Russia, saproxylic species are now facing a similar threat as they have experienced in Europe because of the development of forestry (Wallenius et al., 2010). Moreover, concerns have recently been raised about the characteristics of remnant old-growth forests in managed landscapes; these remnants are often defined by lower productivity or different structures than those observed in natural landscapes (Martin et al., 2020c, 2021a; Price et al., 2020). This implies that some habitats or ecosystem services specific to old-growth forests with higher economic value may be particularly at risk. A conservation approach focusing only on the area of old-growth forest to be conserved, without considering its quality, becomes insufficient. The challenges related to the protection of boreal old-growth forests therefore concern not only their size but also their diversity, connectivity, and functionality.
The expected effects of climate change on old-growth forests remain hard to project, as they can often be contradictory (Fig. 31.4). Late-successional boreal species may benefit or suffer under these future conditions (D’Orangeville et al., 2016, 2018; Thom et al., 2019). The warmer temperatures and longer growing season may enable a northward range expansion for southern boreal or hemiboreal species, such as European beech (Fagus sylvatica L.) or sugar maple (Acer saccharum Marsh) (Bouchard et al., 2019; D’Orangeville et al., 2018; Kramer et al., 2010). The replacement of shade-tolerant boreal species with new late-successional species, however, depends on the migration capacity of the latter, which remains uncertain (Bouchard et al., 2019). If the late-successional species cannot be replaced, developing old-growth forests dominated by pioneer species could be possible. Accordingly, Cumming et al. (2000) have observed forests dominated by trembling aspen (Populus tremuloides Michx.), driven by low- and moderate-severity secondary disturbances. Currently, this type of forest remains rare (Bergeron & Harper, 2009).

Conceptual and simplified flowchart of possible changes in the characteristics of boreal old-growth (OG) forests in the context of future climate change, including processes (italics), old-growth forest characteristics (bold), and possible management solutions (rounded boxes)
Climate change is also expected to increase the frequency and severity of natural disturbances in the coming decades (Bergeron et al., 2017; Kuuluvainen & Gauthier, 2018). This change in disturbance regime may potentially reduce remnant old-growth areas or increase the abundance of forests degraded by recurrent secondary disturbances, thereby eventually overwhelming stand resistance (Bergeron et al., 2017; Martin et al., 2019). Nevertheless, Bergeron et al. (2017) highlighted that in boreal landscapes, industrial-scale forest management based on short-rotation clear-cuts, i.e., rotation periods well below those of regional fire cycles, will remain the principal agent of the loss of old-growth forest, more than the projected increase in fire frequency and other changes in disturbance regimes (Fig. 31.4). Therefore, forest management will certainly have a much greater, immediate, and predictable impact than climate change on boreal old-growth forests. Although we require a better understanding of how climate change affects old-growth forests, the more immediate and pressing need is the proposal of management strategies that efficaciously protect these ecosystems. Combining forest monitoring networks (e.g., SmartForests Canada; Pappas et al., 2022) and mechanistic modeling (e.g., Fatichi et al., 2019) could enhance our process understanding and facilitate the development of sustainable forest management under environmental change.
2.4 Biotic Stress Factors as Underlying Drivers of Ecological Change
The ecosystem-based management of boreal forests emphasizes emulating natural disturbances to maintain the ecological composition, structure, functioning, and services provided by the boreal forest relative to historical baselines (Berglund & Kuuluvainen, 2021; Jackson et al., 2001; Kuuluvainen & Grenfell, 2012). Taking climate change into consideration when developing and implementing ecosystem-based management strategies involves integrating the effects of biotic stress factors that may slowly build up or involve sudden, and likely cumulative, extreme events. Similar to abiotic stress factors, biotic ones can act as underlying drivers of ecological change. In some situations, biotic stress factors can even override the effects of natural disturbances (Nuttle et al., 2013) or counteract the effects of climate change on forest ecosystems (Seidl et al., 2017; Speed et al., 2010; Vuorinen et al., 2020a). We can expect a spatiotemporal lag in trophic interactions under climate warming, as animals and pathogens from temperate forests move into the boreal forest at faster rates than most plants. The vulnerability of trees to combinations of the bottom-up effects of abiotic stresses and top-down effects of biotic effects is, however, difficult to predict, as such effects on plants can be positive, negative, or interactive (Canelles et al., 2021; Teshome et al., 2020; Vuorinen et al., 2020a, b).
Ecological change in the boreal biome occurs because of an increased intensity of biotic stressors, shifts in the climatic niches of temperate species toward the boreal zone, and invasive species. For example, increased moose (Alces alces) abundance can alter the composition, structure, and functioning of balsam fir forests in North America (McLaren & Peterson, 1994; Pastor et al., 1998) and mixed Scots pine and Norway spruce forests in Fennoscandia (Lorentzen et al., 2018), pushing succession along alternative pathways (De Vriendt et al., 2021) and ecological regimes (Gosse et al., 2011; Smith et al., 2010). In Poland, moose altered their behavior in response to higher temperatures by more frequent use of dense forests, which provide greater thermal shelter than open stands (Borowik et al., 2020). Moose populations are also appearing to shift to higher latitudes and altitudes in North America and China in response to warmer late-spring temperatures (Dou et al., 2013; Tape et al., 2016). In Canada, the distribution range of white-tailed deer (Odocoileus virginianus) has expanded northward since the mid-twentieth century, and climate data better explain this range shift than land-use metrics (Dawe & Boutin, 2016). Selective browsing by an introduced population of white-tailed deer north of this species’ historical range and in an area lacking predators has led to the conversion of balsam fir forest to spruce-dominated forest and parkland (Barrette et al., 2014, 2017). Climate warming will likely reduce forage limitations for ungulates in the boreal zone. Although a corresponding increase in predation pressure may counteract the population growth of these herbivores, full compensation is unlikely, as it would require apex predator populations to attain levels above historical numbers, an improbable scenario (Pasanen-Mortensen et al., 2017). Climate change, combined with globalization in the transportation of people and goods, has increased global incidents of invasions by alien species. These introductions now constitute one of the major threats to global biodiversity and planetary ecosystems (Bellard et al., 2013; Vitousek et al., 1996). Seidl et al. (2017) predicted that boreal forests will experience the most pronounced future changes in disturbances of all forest types. In boreal forests, exotic species of defoliator insects, earthworms, slugs, and pathogens, known drivers of major ecological change in forest dynamics, are increasingly observed (Sanderson et al., 2012).
2.5 Future Boreal Tree Growth at Risk
Wood formation is highly sensitive to temperature, and dendroecologists routinely use the growth rings of trees to reconstruct climate prior to the instrumental period (Fritts, 2001). Nonetheless, predicting the future productivity of boreal forest stands under projected increases in temperature and aridity remains extremely challenging. Changes in forest productivity will likely vary because of differences in stand composition and structure, site characteristics, variations in disturbance regimes, and regional and local climate anomalies.
Currently, forest productivity appears stimulated with warming in many cold-limited boreal regions of eastern Canada, Finland, Russia, and Asia (D’Orangeville et al., 2016, 2018; Kauppi et al., 2014; Lapenis et al., 2005; Loehle & Solarik, 2019; Myneni et al., 1997; Zhang et al., 2019a, b) despite important variations in climate sensitivity according to species, competition or stand development (Marchand et al., 2019); however, indicators of reduced growth have been observed in warmer or drier boreal regions (Barber et al., 2000; Beck et al., 2011). Potential growth increases may not necessarily translate into carbon storage gains. Indeed, a higher forest carbon pool hinges on higher mean tree longevity (Körner, 2017); however, higher growth rates have been observed in all forest ecosystems, including boreal forests, to reduce the longevity of trees as trees complete their natural life-span faster, e.g., higher susceptibility to windthrow with increasing size, or get harvested earlier (Brienen et al., 2020; Körner, 2017).
In addition to site factors, species-specific characteristics in terms of climate tolerance, adaptation, and migration capacity can also confound tree growth projections. In eastern Canada, drought-adapted species, such as jack pine (Pinus banksiana), are projected to cope well with projected future climate change (Aubin et al., 2018; Marchand et al., 2021). However, such predictions are based on historical data, whereas all boreal regions are on a warming trajectory that goes beyond that of the observed or reconstructed climate space. In addition, species’ vulnerabilities could depend on the future trajectory of disturbance regimes. The increased frequency and severity of climate anomalies, such as the 2004 drought in central boreal Canada that killed up to 80% of trees in some areas (Michaelian et al., 2011), wildfires (De Groot et al., 2013b) and the expansion of native or exotic forest pests, as seen with the recent mountain pine beetle outbreak in western North America (Cullingham et al., 2011; Robertson et al., 2009), could cancel out any gains in growth within the boreal forest.
2.6 Carbon Sequestration
Carbon sequestration, i.e., the storage of atmospheric CO2 in forested ecosystems (e.g., as tree biomass) at climate-relevant time scales (often referred to as a carbon sink), is one of the many services that forests offer to humanity. The terrestrial carbon sink presents a prominent natural climate solution, contributing to climate change mitigation by absorbing part of anthropogenic CO2 emissions (e.g., Cook-Patton et al., 2020). The circumboreal region includes more than 30% of the Earth’s forested area and represents one of the largest carbon storage pools (Pan et al., 2013). However, the strength of the boreal forest as a C sink varies markedly among regions, and the C sink response to global change remains uncertain. Extrapolating ecophysiological understanding of tree growth (e.g., Hilty et al., 2021) to the landscape and regional scales remains challenging; hence, predicting the fate of the terrestrial carbon sink under global change remains uncertain (Hof et al., 2021). Tree growth provides a carbon sink, yet the resulting ecosystem-level carbon sink stems from numerous interacting processes, including growth (and forest productivity, e.g., Hilty et al., 2021) and respiration (autotrophic and heterotrophic). Natural and anthropogenic disturbances, e.g., fires, insect outbreaks, could abruptly alter the ecosystem-level carbon balance, releasing part of the sequestered carbon back to the atmosphere (Ameray et al., 2021). Quantifying the carbon sequestration potential in the boreal region thus requires an accurate description of the residence times of the carbon stored in different pools (e.g., above vs. belowground; Friend et al., 2014; Pappas et al., 2020). Disentangling the C balance pools could facilitate a robust and quantitative description of the C sink strength and its fate under global change (see Chap. 10).
2.7 New Silvicultural Practices
Over the last century, anthropogenic disturbances have had a stronger impact than climate change on boreal forest/stand compositional changes because of the extensive use of even-aged approaches and the application of clear-cutting as the main silvicultural treatment e.g., in northeastern Canadian forests (Danneyrolles et al., 2019). The urgent need to diversify harvest treatments to reduce the homogenization of forest landscapes has become a priority. The difficulty stems from the past approaches to forest management, which failed to consider cumulative landscape-scale effects in forest planning and heavily favored the use of even-aged approaches and short rotation in boreal stands during the last half-century (see Chaps. 15, 16). The consequences of these decisions reduced the resilience of forest ecosystems facing climate change due to the homogenization and simplification of forest structure and composition across the forest landscapes (Franklin et al., 1997; Puettmann, 2011). This homogenization—promoting the dominance of even-aged stands—and simplification of forest structures affected many species that depend on deadwood, large-diameter trees, and complex horizontal and vertical structures of tree vegetation (Kuuluvainen, 2009). Climate change will alter tree growth and gradually replace existing tree species with more climatically suitable vegetation. Accordingly, these modifications will significantly impact post-disturbance recovery potential (Splawinski et al., 2019) and could affect post-harvest forest resilience and dynamics. Under climate change, the sustainability of forest management in North America is at risk, especially in regions currently characterized by a short fire cycle and low productivity (Gauthier et al., 2015a; Johnstone et al., 2016). Hence, new silvicultural tools and approaches will be required to maintain forest resilience and increase the adaptability of forest ecosystems to novel future conditions (Montoro Girona, 2017; Puettmann, 2011; Spies et al., 2012; Stephens et al., 2013). For Fennoscandian boreal forests, continuous-cover forestry is increasingly promoted to mitigate the loss of biodiversity, ecosystem services, and multiple value chains and ensure a long-term sustained timber yield (Peura et al., 2018; Pukkala, 2016). This shift calls for alternatives to the dominating, intensive rotation forestry. Berglund and Kuuluvainen (2021) proposed a shared (each one-third) distribution between clear-cutting, partial cutting and gap cutting combined with selective thinning on the basis of natural disturbance dynamics.
Partial harvests and variable retention forestry, in which only part of the stand is harvested, represent promising silvicultural approaches that can ensure a more diverse structure in managed forests by maintaining specific ecosystem attributes in the boreal biome, such as large living, dying, and dead trees, to favor greater biodiversity (Gustafsson et al., 2020; Kuuluvainen & Grenfell, 2012; Shorohova et al., 2019). Partial harvests and variable retention forestry include a broad range of treatments, which include commercial thinning (Gagné et al., 2012), selection cutting (Majcen, 1994), and shelterwood cuttings (Montoro Girona et al., 2017, 2018a, 2019; Prévost & DeBlois, 2014; Raymond & Bédard, 2017). Over the last decades, these treatments have been applied in North American and European boreal forests as an alternative to conventional clear-cutting to maintain structural attributes and ensure biodiversity and a continued timber supply (Hernández-Rodríguez et al., 2021; Kim et al., 2021; Moussaoui et al., 2020). However, these silvicultural treatments were initially developed in Europe, and there remains many questions in regard to their potential adaptability to North American boreal forests because of the few experiments and limited long-term monitoring of partial harvests in Canada and the United States (Bose et al., 2014; Montoro Girona et al., 2017).
Over the last 20 years, studies in both Canadian and European boreal forests have attempted to understand the effects of partial cutting on biodiversity and stand yields and have reported several sustainable benefits (Bescond et al., 2011; Brais et al., 2004; Webb et al., 2008). Partial harvesting preserves more favorable habitat attributes for various organisms by maintaining some residual stand structures within cutblocks (Fenton et al., 2013; Kim et al., 2021; Kuuluvainen & Grenfell, 2012; Moussaoui et al., 2016; Ruel et al., 2013). Partial harvesting also promotes increased residual tree growth in boreal forests by light thinning (Montoro Girona et al., 2016, 2017; Pothier et al., 2003; Thorpe et al., 2007). Despite these benefits, the main challenge in implementing partial harvesting in a context of climate change will be adapting this silvicultural treatment to the future conditions of North American boreal forests (species, stands, growth ratio) to develop its potential as a tool for ensuring carbon sequestration, biodiversity, and timber production while being implemented in an economic and financially cost-effective manner. Although additional research is required, these forest practices appear as means of providing an increased forest resistance and resilience to change and facilitating the boreal ecosystem’s ability to adapt to future conditions, e.g., drought and insect outbreaks.
2.8 Including Freshwater Systems Within Forest Management
Water-covered lands represent about 30% of the world boreal forest area (Benoy et al., 2007). In North America, for example, of the 6.3 million km2 of boreal forest, 850,000 km2 is covered by fresh surface waters, and about 1.27 million km2 is covered by peatlands, which, when combined, is equal in size to the country of Indonesia (Gingras et al., 2018). Aquatic and forest environments of the boreal landscape are highly connected because of a high number of contact zones that form a long and complex ecotone in which most aquatic–terrestrial interactions take place. Most organic matter and energy fluxes take their sources in forests and are transported toward aquatic environments by precipitation, freshets, and wind. Once the forest-sourced organic carbon reaches the aquatic habitat, one fraction is processed by the aquatic food web (Grosbois et al., 2020)—either assimilated into biomass or respired—one fraction is stored in the sediments of lakes, rivers, or wetlands, and another fraction is transferred to the atmosphere (Cole et al., 2007). Feedback fluxes from aquatic to forest environments are lower in magnitude because they rely on a faunal transfer of biomass, e.g., aquatic predation by terrestrial consumers and insect emergence from lakes, rivers, and ponds. These fluxes are of higher nutritional quality, however, than the organic matter transported from the forest to aquatic habitats; the latter is in an advanced state of decomposition. Because of these aquatic–terrestrial links and their major implications for forest ecosystem functions, the ecosystem-based management of boreal forests must include freshwater environments. Despite the aim of original ecosystem-based frameworks to manage the forest as a whole, these forest–aquatic system interactions have been largely neglected (see Chap. 29).
Given this strong connectivity between the aquatic and forest environments in the boreal biome, all disturbances affecting the boreal forest also impact aquatic environments within the same watershed. For example, wildfires increase the export of nitrogen and phosphorus to lakes (Lamontagne et al., 2000). Although this quantity represents a negligible amount of nutrients for forest ecosystems, the export of these new wildfire-released nutrients is an important complementary input to freshwaters. Wildfires also influence lake metabolism (Marchand et al., 2009), phytoplankton (Planas et al., 2000), zooplankton (Patoine et al., 2000), and, most likely, the entire aquatic food web. Nonetheless, very little information is available in the literature. This lack of data stems from the logistical challenge of studying wildfires and the traditional separation in the study of land-based disturbances and aquatic habitats. Forestry activities represent an anthropogenic disturbance that, in addition to causing an increased export of nutrients to lakes, produces an additional input of dissolved organic carbon (DOC), which affects the physiochemistry of a water body. The removal of wood drastically diminishes the forest’s capacity to retain precipitation and organic matter. The result is an enhanced release and transport into aquatic environments of terrestrial dissolved molecules and particles of inorganic and organic carbon. The new inputs of forest-derived dissolved organic molecules alter the water color (browning), and the added particles also increase turbidity in the water column. Both changes affect light penetration in the water column and therefore influence primary productivity, e.g., algal abundance (Steedman, 2000). Higher DOC concentrations can impact the entire food web and diminish fish growth (Benoît et al., 2016). Also, forest harvesting increases the mobility of methylmercury and increases its assimilation into the aquatic biomass of plankton and fish (Garcia & Carignan, 2000; Wu et al., 2018). The impact of other disturbances, such as insect outbreak or windthrow, on aquatic environments, has yet to be studied. Their influence on freshwater physicochemical properties, freshwater metabolism, and food webs therefore remains unknown. It is thus essential for future research to investigate how freshwater systems and their associated food webs react to land disturbances and adjust forestry operations to ensure the sustainable management of the boreal forest, especially in the context of future climate change.
2.9 Connectivity and Fragmentation
Connectivity and fragmentation are natural components in the continuum of habitat types and their transition zones, which comprise ecosystem configurations at multiple spatial and temporal scales. Both connectivity and fragmentation vary and change in response to natural disturbances and dynamics as well as anthropogenic influences. Intact forest landscapes and primary forests provide an ecological legacy that harbors intrinsic ecosystem resilience and adaptive capacity to withstand degradation (Lindner et al., 2010; Potapov et al., 2017; Sabatini et al., 2020; Venier et al., 2018) and avoid a sledgehammer effect where ecosystems risk entering new and potentially irreversible ecological states (Barnosky et al., 2012); this is particularly important in forests facing future climate change. With greater than 70% of the Earth’s land surface (Barnosky et al., 2012) and more than 80% of the remaining forests (Watson et al., 2018) modified by land use, however, the Anthropocene human footprint critically influences key ecological functions (Tucker et al., 2018). In addition to relocating natural forest frontiers (Potapov et al., 2017) and decreasing remaining intact forest landscape area (Svensson et al., 2020), extensive land use has also generically affected landscape matrix functionality and the remaining protected areas (Heino et al., 2015; Jones et al., 2018).
The consequences of anthropogenic-related forest fragmentation are increasingly debated (Ward et al., 2020). In many regions of the boreal biome, the natural configuration of forest landscapes has become seriously marked by systematic clear-cutting and monoculture–rotation forestry systems (Boucher et al., 2009; Peura et al., 2018) at rates beyond those of sustainability and biodiversity policies and environmental targets (Chazdon, 2018; Jonsson et al., 2019; Selva et al., 2020). The consequent forest landscape fragmentation has had consequences beyond the actual loss of primary forest area and the separation of a few larger areas into many smaller retained and set-aside patches. Edge effects penetrating the remaining patches also generate a proportionally larger loss of functional core areas (Harper et al., 2015; Pfeifer et al., 2017). A study of boreal Sweden, for example, revealed that systematic forest clear-cutting since the middle of the twentieth century has left behind only 6% primary forest core area for this biome (Svensson et al., 2019). In addition, artificial forest edges are created; these edges do not harbor the natural ecological attributes associated with natural edges (Haddad et al., 2015), further affecting biodiversity and ecosystem resilience. Hence, the re-creation of both structural and functional connectivity in landscapes typified by extensive clear-cutting forestry is challenging (Chazdon, 2018; Ward et al., 2020).
2.10 Collaborative Research and Indigenous Peoples
The relationship with the land is a foundation of Indigenous peoples’ identity, culture, and livelihood in boreal regions. Boreal landscapes are places of hunting, trapping, fishing, cultural and language learning, and healing. These practices provide access to a multitude of ecosystem services, both tangible and intangible, and contribute to the well-being of Indigenous people (Davidson-Hunt & Berkes, 2003; Saint-Arnaud et al., 2009). However, climate change and forest management drive major transformations of boreal forests and affect Indigenous people's relationship with the land (Fuentes et al., 2020; Turner & Clifton, 2009). Although Indigenous institutions play an increasing role in forest governance (Wyatt et al., 2019), the consideration of Indigenous values and perspectives remains the exception rather than the rule.
Collaborative research contributes to bridging the different perspectives on boreal landscapes and facilitates forest co-management through various means (Blackstock et al., 2007):
-
Collaborative research calls for the complementarity between Indigenous knowledge and science-based knowledge (e.g., Asselin, 2015; Suffice et al., 2017). Their combination extends the spatial and temporal scales of observations and provides a wider and comprehensive understanding of boreal environments (Bartlett et al., 2012; Lyver et al., 2018).
-
The bridging of Indigenous and scientific knowledge is based on the premise that every knowledge is situated within a knowledge system and is partial, and that there is no hierarchy between knowledge systems (Ericksen & Woodley, 2005). Following this principle, collaborative research legitimizes the knowledge creation process and the associated land management decisions.
-
Indigenous people are underrepresented in scientific research institutions, as are Indigenous concepts, methodologies, and ethics (Littlechild et al., 2021; McGregor, 2018). Collaborative research contributes to increasing the research capacity within Indigenous institutions and the Indigenous representation within scientific institutions.
Collaborative research, however, faces challenges. First, Indigenous and science-based knowledge belong to different knowledge systems, with their ontologies, epistemologies, and methodologies (Bartlett et al., 2012). Knowledge co-production requires sustained work at the boundary between knowledge systems to formulate research questions, make explicit the different perspectives of a phenomenon, and develop appropriate methodologies (Dam Lam et al., 2019; Robinson & Wallington, 2012). Second, research partnerships need to overcome existing mistrusts. On the one hand, Indigenous people have experienced negative relationships with scientists in the past, and confidence often needs to be rebuilt and be based on stronger research ethics (Canadian Institutes of Health Research, Natural Sciences and Engineering Research Council of Canada and Social Sciences and Humanities Research Council, 2018). On the other hand, scientists often seek to validate Indigenous knowledge with the same methods as for validating experimental ecological data (Gilchrist et al., 2005). Such an exercise is problematic because a piece of knowledge is taken out of the knowledge system that defines its meaning, sense, and scope and is imported into a different knowledge system without the contextual information required to appreciate its value and validity (Castleden et al., 2017). Extended collaborative work is thus needed to co-produce better-informed, legitimate, and valid knowledge, following both knowledge systems.
2.11 Resilient Forest Landscapes
The long-standing forest management approaches targeting only highly productive monocultures have homogenized and simplified forest ecosystems worldwide (Puettmann et al., 2009). Given that climate and global change entail multiple environmental and socioeconomic uncertainties, we urgently need novel forest management paradigms that (1) acknowledge this future uncertainty; (2) promote forest ecosystem resilience to altered disturbance regimes and climate change mainly through the functional diversification of tree communities; and (3) scale up the objectives and impacts of these novel silvicultural interventions from the stand to the landscape scale (Messier et al., 2019). The challenge lies in adopting a trait-based approach to rethink and redesign sustainable forest management plans (Cadotte et al., 2011). New management paradigms should foster functional diversity and redundancy within tree communities to actively turn boreal forests into ecosystems that are more resistant to known and unknown disturbances and have a higher capacity for adapting to novel environmental conditions.

Functional group classification of tree species commonly found in Canada's Boreal Shield and Mixedwood Plains ecozones. The eight functional traits used to classify the species are drought tolerance, shade tolerance, waterlogging tolerance, main seed dispersal vector, seed mass, wood density, leaf mass area, and taxonomic division. Of these eight traits, drought, shade, and waterlogging tolerance reflect a species susceptibility to environmental conditions (Niinemets & Valladares, 2006), whereas the other five relate to the capacity and mechanisms of a species to respond to natural disturbances. The bottom diagram presents the number of species, the number of functional groups, the functional diversity, and the functional redundancy of six typical tree communities. Vertical lines between the tree illustrations separate the functional groups. a This community consists of four species of the same functional group; it therefore has a very low functional diversity, although the functional redundancy is very strong because if a species disappears, the main functional traits remain in the stand. b This three-species community is relatively functionally poor, and the functional redundancy is very low because the loss of a species may compromise community functioning. c Functional diversity is moderate as up to three functional groups are present; however, as the loss of a species will certainly entail the loss of important traits, functional redundancy is very low. d This community is taxonomically and functionally richer than that of c and because half of the functional groups are represented by more than one species, redundancy is moderate. e In the most species-rich community, functional redundancy is very high because the three present functional groups are represented by more than one species. d This tree community is the most functionally diverse; however, six species are not sufficient to maintain all the diversity when a species is lost. Consequently, functional redundancy is very low
Plant functional traits determine, on one level, the fitness of each individual via their effects on growth, reproduction, and survival and thus their influence on ecosystem functioning, including productivity, competition for resources, and nutrient balancing (Lavorel & Garnier, 2002). On another level, traits also determine the strategies and responses of the species to changing environmental conditions and determine how tree populations respond to different environmental factors and recover from disturbances (Lavorel & Garnier, 2002). When explicitly considering effect traits, the higher the functional diversity of an ecosystem, the higher its overall productivity (Tilman et al., 1997); but when accounting for response traits, functional diversity indicates the variety of forms possessed by a community to resist environmental changes, recover from disturbances, and adapt to novel environmental conditions (Elmqvist et al., 2003). Functional redundancy acts as insurance for the ecosystem because the loss of a redundant species, i.e., a species that performs similar roles within the ecosystem and responds similarly to environmental stressors and disturbances, will not compromise ecosystem functioning and resistance. Therefore, high-functional diversity and redundancy translate into a more resilient ecosystem (Mori et al., 2013).
Specifying the management guidelines for a particular forest region requires not only quantifying the functional diversity and redundancy of current tree communities and the entire landscape (Aquilué et al., 2020) and determining other suitable species that could establish and grow in the region either naturally or through assisted migration. Some community-level functional dissimilarity measures can be partitioned in a way to quantify species-level contributions to overall functional diversity (Pavoine & Ricotta, 2019). Another way to approach this task is to group species not using a taxonomic-based method but rather via a trait-based method by clustering species into functionally homogeneous groups (Fig. 31.5). Species of the same functional group share similar functioning at the individual level, similar responses to environmental variations, and similar effects on the ecosystem (Cornelissen et al., 2003). Once tree species are clustered into functional groups, it is possible to identify the surplus functions and functional groups, as well as those less represented, and later target species having a greater potential to maximize functional diversity at both the community and landscape scales (Aquilué et al., 2021).
3 How to Face the Challenges Confronting the Boreal Biome? Looking Toward the Future: Implications for Ecosystem-Based Management
The future holds many challenges and much uncertainty. In this section, we outline the main directions and perspectives required to confront these challenges and the implications for ecosystem-based forest management in the future boreal biome.
3.1 Alteration of Natural Disturbance Regimes
As climate change is expected to trigger important changes in future disturbance regimes, forest management will face notable economic and ecological challenges. Setting management objectives is becoming increasingly complex, as forests are expected to provide various ecosystem services other than timber, including carbon emissions mitigation, biodiversity, protection, and recreative roles (Ameray et al., 2021; Thom & Seidl, 2016). With the increased risk of boreal forests losing large amounts of trees to natural disturbances, the main concern for the forestry sector is the possibility of significant timber shortfalls in the near future (Boulanger et al., 2019; Daniel et al., 2017; De Grandpré et al., 2018). Although large uncertainties remain, evaluating future risks at meaningful spatial and temporal scales is a crucial first step (Daniel et al., 2017; De Grandpré et al., 2018).
Ecosystem-based forest management aims to preserve natural forest attributes and processes by setting forestry strategies and targets on the basis of the variability of past disturbance regimes (Landres et al., 1999); for example, given the dominant role of wildfires in driving the dynamics of Canadian boreal forests, forest managers in Canada rely heavily on presettlement wildfire regimes to develop ecosystem-based management strategies (Thom & Seidl, 2016). In practice, this implies, for example, that the total annually burned or harvested area should not exceed that having burned under past fire regimes. Simultaneously integrating the risk of all-natural disturbances into ecosystem-based management strategies will be essential to ensure the applicability of this management approach under future conditions (Thom & Seidl, 2016).
Several management actions can be undertaken to handle the risks associated with future natural disturbance regimes in boreal forests. In areas facing high fire risks, governments should invest in fuel management to reduce the potential of fire occurrence and spread (De Groot et al., 2013b). Raising public awareness and producing prevention campaigns that explain the effects of forest fires can limit the occurrence of human-induced fires, likely to increase in frequency in drier conditions in the future. Other mitigation measures, such as favoring tree species that are less sensitive to insect outbreak, disease, drought, or fire, can be applied in these high-risk areas. Protecting foliage, reducing forest homogeneity, strategically removing fallen and weakened trees, thinning, debarking, and applying biological controls could all help reduce the risk of insect outbreak and disease (Ivantsova et al., 2019; Sturtevant et al., 2015). The rescheduling of harvests must also be considered to maximize timber production during a budworm outbreak (Sturtevant et al., 2015). Changes in the timing and intensity of management actions can also heighten the resistance of boreal forests to snow- and wind-related damage. Specifically, this resistance could be achieved by changing forest landscape-level structures, such as the distance to stand edges, decreasing stand height differences, or shortening harvest rotations (Díaz-Yáñez et al., 2019; Zeng et al., 2007; Zubizarreta-Gerendiain et al., 2017).
3.2 Biodiversity Loss and Forest Attributes
Ecosystem-based management could mediate between preserving ecological processes, economic goals, and social values. We may need to increase uneven-aged and continuous-cover forest management approaches to maintain desired levels of late-successional trees in forest landscapes. Moreover, uneven-aged managed stands tend to be less prone to windthrow than even-aged stands, and landscapes with larger old-forest patches are less vulnerable to fire (Leduc et al., 2015; Nevalainen, 2017; Pukkala, 2016). Measures to rehabilitate important forest structures and improve habitat quality for biodiversity conservation involve ecological restoration through simulating natural disturbances, e.g., prescribed burning and gap cutting (Hägglund et al., 2015; Hekkala et al., 2014; Hjältén et al., 2017; Versluijs et al., 2017), and the use of longer rotation cycles to ensure sufficient and adequate habitats for forest specialists (Roberge et al., 2018).
From a biodiversity perspective, the top priority is to ensure a sufficient proportion of old-growth and uneven-aged forests in the landscape (Kuuluvainen & Gauthier, 2018). To this end, an obvious measure would be to reduce harvesting levels (Daniel et al., 2017; Kuuluvainen & Gauthier, 2018). Management practices aiming to conserve post-disturbance legacy structures and old-forest attributes should also be promoted (Boulanger et al., 2019; De Grandpré et al., 2018; Kuuluvainen & Gauthier, 2018). At the stand scale, this can be achieved in part by promoting partial cutting over clear-cuts (Bose et al., 2014). In addition, salvage logging, i.e., the harvesting of disturbed forests, is a relatively common and increasingly used post-disturbance management strategy (Leverkus et al., 2018; Sturtevant et al., 2015). The use of salvage logging should, however, be carefully prescribed depending upon management objectives; for instance, as windthrow creates large amounts of deadwood, salvage logging should be avoided when priority is given to habitats that favor biodiversity (Nappi et al., 2011; Thorn et al., 2020). Nonetheless, salvage logging could offer a preferable treatment in recreational areas, for example, where large numbers of trees have been weakened by insects or disease and threaten to produce tree fall–related accidents (Ivantsova et al., 2019). All these measures could potentially generate economic losses; these losses could be compensated, to some degree, by valuing timber quality rather than quantity. Finally, a better understanding of fire–carbon feedbacks and deadwood dynamics resulting from insect outbreaks, disease, or weather-related disturbances would be necessary to better monitor carbon storage and release in boreal forests. This data would help develop improved, adapted management strategies that can limit carbon emissions.
3.3 Ecosystem-Based Management of Boreal Old-Growth Forests
Even-aged management based on short-rotation periods and clear-cutting has been, and will continue to be, the main cause of the loss of old-growth forest areas (Bergeron et al., 2017; Kuuluvainen & Gauthier, 2018; Martin et al., 2020c). The implementation of ecosystem-based management in boreal landscapes therefore requires a profound change in forestry practices. A combination of continuous-cover forestry, salvage logging, and clear-cutting with longer rotation periods, associated with the proactive mitigation of severe natural disturbances, has the potential to attain a balance between sustainable wood provision and environmental objectives in the context of climate change (Bergeron et al., 2006; Kuuluvainen, 2009; Leduc et al., 2015). Continuous-cover forestry can also be used to restore old-growth attributes in areas where these forests are currently absent, while limiting the anthropogenic impact on remnant old-growth forests (Eyvindson et al., 2021; Fenton et al., 2013; Montoro Girona et al., 2017). The remaining intact forests have an invaluable role as a natural reference from which to learn (Watson et al., 2018), and the conservation of the last large tracts of boreal old-growth forest must be a priority. In addition to the retention and continuous-cover forestry, natural disturbance–based management may provide a framework to increase the biodiversity, resiliency, and adaptive capacity of boreal forests (Kuuluvainen et al., 2021). Development of these management approaches based on scientific knowledge will heighten the flexibility to forest management, allowing it to maintain the structural diversity observed in old-growth forests and to better adapt to the new constraints caused by climate change (Fig. 31.4; Kuuluvainen et al., 2019). For example, in the event of conifer decline, it may be possible to carry out the assisted migration of shade-intolerant species following partial cutting.
Successful ecosystem-based management and the application of appropriate conservation and restoration activities needed to sustain old-growth forests require a detailed ecological understanding of the occurrence and dynamics of these key ecosystems. For example, the accurate identification and mapping of boreal old-growth forests, including their structural diversity, are necessary steps to define relevant protected old-growth areas that are representative of the preindustrial landscapes and to establish effective restoration targets. However, the efficacy of current survey methods, e.g., aerial photographic surveys, has been questioned (Martin et al., 2020b). Therefore, it is necessary to develop new tools, e.g., using LiDAR or UAV, to better identify the diversity and dynamics of old-growth forests in boreal landscapes. Furthermore, old-growth definitions can greatly vary between jurisdictions, including within the same country (see Chap. 7). This barrier severely limits the effectiveness of any strategy for old-growth conservation, for instance by reducing the coherence of measures taken by different actors and their degree of complexity. In the future, it will be necessary to continue the scientific effort initiated during the last decades to broaden our knowledge of old-growth boreal forests, a key for their preservation and sustainable management.
3.4 Tree Growth and Productivity
Effective management strategies require large-scale studies to better estimate key drivers of forest demographics under global change; these drivers include warmer temperatures, potential frost damage, and more severe water deficits. In turn, these drivers should be integrated into the next generation of forest growth models to simulate the interactive effects of climate change, insects, and disease on tree growth and mortality (Anderegg et al., 2015; Fatichi et al., 2019; McDowell et al., 2020). Until these data are available, risk reduction through portfolio diversification should drive management strategies for maintaining growth under global change. This strategy can include diversifying the composition of the boreal forest—a biome in which a few tree species dominate large tracts of landscapes—toward drought-adapted species and genotypes, thereby increasing the structural complexity of the forest (Messier et al., 2019). Another strategy is to increase the abundance of warm-adapted boreal broadleaf species, such as aspen and birch, to reduce fire risks while also increasing surface albedo (Astrup et al., 2018).
Maintaining stand productivity can also be ensured through stand-level interventions. Across a range of forest types in the United States, thinning has been shown to significantly increase resistance to greater water deficits (Bottero et al., 2017; D’Amato et al., 2013; Grant et al., 2013). In the boreal region of Canada, thinned balsam fir stands experiencing drought have shown increased resistance relative to natural stands, although important interactions with tree position and size, where larger or suppressed trees displayed lower resistance to these climate anomalies (Fig. 31.6). Revisiting long-term silvicultural studies, improving forest models, and moving away from business-as-usual forest management could yield the critical solutions for increasing growth resilience in the boreal forest in the context of global change.

Variation in the drought response of balsam fir tree growth (1999–2017) in terms of a tree canopy position (ratio of tree height to average stand height; positive values indicate dominant trees), b tree size (diameter at breast height), and c stand density. The model was fitted to the standardized annual growth of 247 balsam fir trees sampled across 22 stands within the Montmorency Forest (Québec, Canada) between 1999 and 2017. A value of 100% predicted growth corresponds to the standardized mean annual diameter growth
3.5 Biotic Stressors
New challenges arising from shifting environmental conditions, species invasions, and human impacts will likely modify trophic interactions (Filbee-Dexter et al., 2017). Accounting for biotic stressors (in combination with abiotic ones) is essential when implementing an ecosystem-based forest management strategy, despite the risk of complicating the achievement of management targets (De Vriendt et al., 2021). These various ecological drivers and processes operate at different and often specific scales. The linking of management actions relative to wildlife and forests at appropriate spatial and temporal scales, as illustrated by Beguin et al. (2016), is essential for achieving ecosystem-based management goals (Fig. 31.7). This will require transdisciplinary collaboration to develop a consensus regarding those factors driving ecosystem succession and the identified management challenges, objectives, targets, and indicators (Gunderson, 2015). At the same time, we must develop flexible and adaptable processes able to deal with ecological surprises (Filbee-Dexter et al., 2017), alternative successional pathways (Hidding et al., 2013), and regime shifts (Folke et al., 2004; Gauthier et al., 2015b).

Modified from Beguin et al. (2016), CC BY 3.0 license
Wildlife and forest management actions at multiple temporal and spatial scales.
3.6 Forest Plantations as Silvicultural Tools for Adapting to New Conditions
Forest plantations, i.e., cultivated forest ecosystems established by planting, seeding, or both in the process of afforestation and reforestation, are often put forward in many parts of the world as a sustainable silvicultural tool for reconciling wood production and preserving the original natural forest (Paquette & Messier, 2010). Whereas forest plantations comprise less than 5% of forested lands, these areas provide 15% of the world’s timber production (Carnus et al., 2006). Nonetheless, biodiversity–ecosystem functioning must be considered when implementing plantations to ensure diverse forests, rather than large-scale monocultures, to favor a greater resistance to insect pests and diseases (Carnus et al., 2006; Paquette & Messier, 2010). Including conservation and various objectives into tree plantation planning will be a major future challenge for diversifying plantations. In addition to reducing pressure on natural forests, the use of forest plantations can restore some lost ecological forest services (Sedjo & Botkin, 1997), such as carbon sequestration (Ameray et al., 2021). Preventive planning is needed to deal with both the beneficial and detrimental effects of climate change. Managing plantations in an ecologically sustainable manner and planning for predicted future climate conditions will promote the maintenance of biodiversity at all scales (stand, landscape) and for all components (genes, species, communities). For example, by controlling invasive species, forest plantations can reduce non-natural competition and thus protect natural habitats, refuge networks, and tree populations that are either isolated or at margins of their distributions (Paquette & Messier, 2010). Moreover, we can accommodate the new climatic conditions by reforesting with local or locally adapted seed sources suitable for future conditions (Puettmann, 2011). However, natural disturbances regimes must to be taken into consideration, because if the fire regimes are shorter in the future, plantations could have a low chance of reaching commercial maturity in areas where burn rate will be shorter than the harvest age. The main challenge of forest plantations is to maintain the desired levels of stands in an ecologically sustainable manner by anticipatory planning and scientific management.
3.7 Silviculture in a Changing Context
Climate change heightens the variability and uncertainty of future ecological, economic, and social contexts. Maintaining and restoring biodiversity and the structural complexity of forests can promote forest resilience to human-induced pressures and impacts from climate change (Thompson et al., 2009). The structural and functional attributes of boreal forests are being altered quickly and significantly in response to a changing climate. FEM, however, promotes the diversification of silvicultural practices and the maintenance of forest stands of various structures. Given that these structures are in constant evolution in a changing context, FEM offers a means of prioritizing ecological processes that ensure the functioning of forest ecosystems. In this context, we must also consider factors other than timber production, including recreation, wildlife, and biodiversity conservation. The range of EBM-derived silvicultural practices can be a valuable tool for helping forests respond to a changing climate because these treatments generally aim to manage and sustain the growth, structure, and composition of forest vegetation for multiple objectives, including wildlife habitat, timber production, water resources, and recreation (Millar et al., 2007; Rist & Moen, 2013). There is thus a need to develop new silvicultural practices to increase forest resilience and promote the conservation of the ecosystem goods (wood and nonwood products) and services provided by forests, such as climate regulation, carbon sequestration, soil stabilization, nutrient cycling, and recreational values (Ameray et al., 2021; Hassan et al., 2005). To ensure that silviculture becomes a useful tool in rapidly changing realities, this discipline must build a new framework with a focus in three axes (Achim et al., 2021): (1) observational (monitor forest, detect changes, update data, integrate knowledge); (2) anticipative (integrate climate projections, predict future scenarios); and (3) adaptive (flexible implementation, risk acceptance, consideration of social acceptance). This new framework could be helpful to adapt silviculture to respond to new trends, needs, and preoccupations at the ecological, social, and economic scales, including a holistic conception of forest ecosystems under climate change.
3.8 Accounting for Freshwaters in the Sustainable Management of Forests
Forestry practices result in ecological consequences not only for the forest from which the trees are harvested but also for freshwaters within the harvested watershed. Including freshwater systems into the sustainable management of boreal forests is therefore vital in future management decisions. Maintaining healthy boreal freshwater systems benefits terrestrial food webs, which depend on the high nutritional quality feedback fluxes from the aquatic to land environments. Aquatic environments are key contributors to the ecological functioning of terrestrial food webs and are critical to many key boreal species; for example, moose and beaver feed largely on aquatic macrophytes (Bergman et al., 2018; Feldman et al., 2020; Fraser et al., 1984; Labrecque-Foy et al., 2020). Many terrestrial bird species also depend on the emergence of aquatic insects (Murakami & Nakano, 2002). Ensuring a healthy aquatic environment positively affects the health of terrestrial environments and, thus, of the entire boreal biome.
Currently, riparian buffer strips represent the best means of protecting aquatic habitats from terrestrial human disturbances such as forest harvesting. These buffer strips are applied universally to all forest types and ecosystems; however, they do not respond to all specific protection needs. Riparian buffer strips must be adapted to the physical and biological landscape. For example, strip width and composition should be adapted to the slope, soil type, and forest stand to reduce windthrow and high tree mortality. It is now crucial to understand the impact of forest disturbances on aquatic environments, as changing land use and climate are expected to increase the terrestrial influence on freshwaters, e.g., browning. Determining the resiliency of aquatic habitats and food webs will be especially important to define the maximum supported human pressure on boreal forests. Accounting for freshwaters within the forestry plans will therefore be essential for attaining the sustainable management of the boreal forest and ensuring the health of the entire boreal biome.

Boreal landscape sustainability according to Raworth’s (2017) doughnut economics and land-use experts from the Abitibiwinni and Ouje-Bougoumou First Nations
3.9 Green Infrastructure
Green infrastructure is a strategically planned management of natural and anthropic lands that aims to secure biodiversity, habitat resilience, and ecosystem services at multiple spatial scales (EC, 2013; Liquete et al., 2015; Wang & Banzhaf, 2018). Thus, green infrastructure promotes landscape-scale and holistic planning approaches that are based on remaining biodiversity and ecosystem-service hot spots and their functional connectivity within a landscape matrix subjected to ongoing climate and land-use changes. In forest landscapes having a legacy of extractive forestry, applying green infrastructure implies that forest patches are restored, matrix quality is improved, and connectedness is strengthened (Dondina et al., 2017; Heller & Zavaleta, 2009). In addition to preserving existing intact forest landscapes and primary forest as connectivity nodes, networks can be re-created within degraded or extensively transformed forest landscapes to secure functional habitats for species to move and spread. Forest edges, for example, are important transitional biotopes but also hold specific conservation values (Harper et al., 2015) and provide a functional green infrastructure in natural as well as need-to-be-restored anthropic forest landscapes in which clear-cut edges dominate (Esseen et al., 2016). Approaches are required that range from local species occurrence and microsites to habitats, landscapes, and entire regions to attain functional connectivity, i.e., a connectivity that supports the representative traits of species composition, habitat structures, and ecological processes (Heller & Zavaleta, 2009) and that goes beyond the protection of remaining biodiversity key habitats to forest landscape restoration and prestoration (Mansourian, 2018). Nonetheless, pan-national policies and policy implementation instruments and routines are needed to ensure that fragmentation (as a consequence) and connectivity (as a necessity) are accounted for in boreal sustainable forest management and governance.
3.10 A Sustainability Framework Emerging from Collaborative Research
An example of collaborative research is the partnership (2015–2021) between university researchers and the Abitibiwinni (Anishnaabeg) and Ouje-Bougoumou (Cree) First Nations, all located in boreal Québec, Canada, that evaluated the effects of environmental change on Indigenous landscapes. The main results can be synthesized in a sustainability framework inspired by Kate Raworth’s doughnut-shaped sustainability economics diagram (Raworth, 2017) (Fig. 31.8). The inner boundary of the doughnut represents the limits of human well-being (Nussbaum, 2011; Sen, 1980) and is defined as the capacity of the practices that are important for an Indigenous community, e.g., moose hunting, trapping, and education. The well-being boundary can be delineated with indicators of the resources abundance of, access to the land, quality of resources, and lived experience while on the land (Bélisle et al., 2021). The outer boundary represents the ecological limits of boreal landscapes (O’Neill et al., 2018; Rockström et al., 2009a). It is delineated by a set of influential factors identified by Indigenous land-use experts, e.g., forest road density, forest composition, and water quality (Bélisle & Asselin, 2021). In between these two zones, the sustainability zone represents the zone to be targeted and monitored for ecosystem management purposes.
3.11 The Functional Network Approach
From an ecosystem-based management perspective, forest resilience to global threats and uncertainty can be achieved through the functional diversification of tree communities. This diversification requires, critically, the identification of both functionally redundant and functionally rare species to prevent their likely decrease in abundance and promote their regeneration either naturally or by planting (Aquilué et al., 2021). Network theory can then be applied to target locations to optimize silvicultural interventions and attain greater levels of landscape resilience.
In the functional network approach, a forest landscape is represented as a network of forest stands. Tree species dispersal capacity dictates whether forest stands are connected; thus, the relative dispersal capacity within and between adjacent stands influences the dispersion of the associated functional traits for each tree species. Three network/landscape-level properties contribute to forest ecosystem resilience: functional connectivity, modularity, and node-level centrality (Fig. 31.9; Aquilué et al., 2020; Gonzalès & Parrott, 2012). A higher functional connectivity between forest stands facilitates the exchange of functionally distinct species and genes better adapted to novel environmental conditions and ensures a rapid tree recolonization of disturbed stands by seeds coming from the surrounding intact stands; these conditions contribute to an efficient reorganization of the ecosystem. Modular systems are organized in clusters of highly interconnected nodes that are loosely connected to nodes of other clusters. Modularity acts as an effective defense against the spread of pathogen outbreaks because modules buffer the spread of a perturbation and minimize the risk of ecosystem collapse (Gilarranz et al., 2017). Therefore, hypothetical modular forest landscapes will be functionally structured in clusters and less vulnerable to rapid, synchronized insect outbreaks. Finally, node centrality accounts for the different roles an element plays in the flow of energy, nutrients, organisms, and genes across the landscape (Bodin & Saura, 2010). Generally, central nodes concentrate most connections and/or bridge two subsets of nodes that would otherwise be disconnected. In a forest landscape, central stands (regardless of size) act as both source and sink for a large proportion of species and functional traits within the landscape.

Characterization of a fragmented forest landscape relying on two functional indicators (diversity, redundancy) and three properties of the spatial network (connectivity, centrality, modularity). The reference is a forested landscape in an agroforest mosaic in which forest stands are separated by agricultural fields, roads, and small villages. The enrichment strategy promotes natural regeneration of functionally rare species within the existing forest stands, and the plantations strategy aims to convert bare or agricultural land into woodlands using multispecies plantations to enhance the functional diversification of the landscape
Practitioners can evaluate current management plans and silvicultural practices at the landscape scale through the analyses of these five indicators—functional diversity, functional redundancy, functional connectivity, modularity, and centrality (Fig. 31.9). They can also assess novel, untested alternative management regimes, in particular where network analysis is combined with a modeling framework to evaluate future projections and determine the compounding impacts of natural disturbances, climate change, and silvicultural interventions on forest ecosystems (Hof et al., 2021; Mina et al., 2021). If tree planting is envisaged, it is advisable to consider species vulnerability to these natural disturbances that regularly or will likely impact a region in the near future (Mina et al., 2021). This extra challenge makes it even more important to merge environmental model predictions with expert knowledge. In short, foresters, climatologists, entomologists, modelers, economists, and other relevant experts must collaborate within a transdisciplinary framework to design ecosystem-based management plans that foster forest resilience to global change.
3.12 Restoration Under Climate Change
Restoration theory is generally based on the emulation of natural disturbances (Kuuluvainen, 2002). However, climate change will most likely impact disturbance regimes, such as by modifying the frequency and severity of forest fires, windthrow, floods, and insect infestations. Thus, future restoration efforts also must confront the challenge of climate change; we require a solid understanding of how global change may affect forest dynamics if we wish to safeguard ecosystem services and the biodiversity hosted by forests (Aerts & Honnay, 2011; Harris et al., 2006). The question is whether we should adapt ecological restoration to fit this new natural disturbance regime. One obstacle in this respect is that climate change effects are likely to be abrupt and unpredictable, where sudden and significant changes are inevitable in the next 20 years (Harris et al., 2006). As climate changes, the patterns of natural disturbances are also shifting, increasing uncertainty in terms of how the dynamics of forest ecosystems will evolve in the future (Johnstone et al., 2016). Paleoecological studies have demonstrated that the boreal forest has a substantial resilience to natural disturbances (Aakala et al., 2018; Carcaillet et al., 2001; Navarro et al., 2018a). However, alterations in disturbance regimes can also increase the vulnerability of forest ecosystems to degradation (Seidl et al., 2016). For example, a hurricane damaged 75 million m3 of Swedish spruce forests in 2005, resulting in an economic loss of $2 billion US (Valinger & Fridman, 2011). Consequently, ecological restoration must also consider the effects of the altered natural disturbances on ecosystem services. Disturbances operate at several and nested spatiotemporal scales, causing the heterogeneity of stands (age structure) and landscape patches (Kuuluvainen & Gauthier, 2018). Thus, to effectively restore forest ecosystems subjected to climate change, we must understand the factors involved in determining when, how, and where fire, insect outbreaks, and windthrow regimes have altered and will alter forest dynamics to restore for future scenarios rather than previous reference states.
3.13 Governance Policies
Governance policies have been essential for the implementation of FEM and in attempts to minimize future impacts of climate change. Moving forward, international and national legislative frameworks must better integrate climate change scenarios to enhance the protection of habitat types, species, and carbon sequestration. Local communities are also vulnerable to climate change through, for example, impacts on livelihoods and forest-based activities. These must also be considered in land-use planning at different scales. To the best of our knowledge, some topics have yet to be addressed within any country, such as ecological restoration within climate change strategies or functional landscape planning strategies. Moreover, some direct or partial conflicts remain between climate change adaptation and mitigation strategies and forest biodiversity goals, e.g., keeping deadwood versus removing logging residues. This is reflected in differences in policy direction among boreal countries. For example, the European Union considers climate change adaptation, natural resource sustainability, and restoration as central priorities, thereby becoming foci of environmental policy. In Canada, general plans and actions around climate change adaptations are occurring at multiple scales, including specific actions in the area of forest management adaptation, however there is little evidence of coordination or an overarching framework. In the case of restoration policies in Canada, the main focus is on specific species, rather than on risks to ecosystems, i.e., static conservation goals (Harris et al., 2006; Parks Canada, 2008). In the Russian Federation, ecological restoration continues to be missing from governance and climate policies, and commitments vary from minimal efforts to no action and are not at all consistent with the Paris Agreement. Given this situation, we suggest creating an active strategy for boreal countries to coordinate their sustainable forest management actions and ecological restoration policies under the umbrella of climate change.
4 Conclusions
Ongoing intensive forestry practices have significantly altered boreal forest ecosystems and will continue to do so, illustrated in part by the increased landscape fragmentation and homogenization of forest stands and a reduced species diversity. FEM is a promising approach for maintaining all ecosystem services, habitats, and functions provided by forests. However, in addition to intensive forestry, climate change is altering forest ecosystems around the world. Boreal forests face multiple challenges for which unpredictability and uncertainty will be the new baseline conditions, further complicating sustainable forest management. Thus, existing paradigms and the implementation of FEM must be sufficiently flexible and adaptable given that we will experience novel, previously unknown or unobserved scenarios. Solutions for safeguarding ecosystem services and biodiversity require a solid understanding of how global change will affect forest dynamics and processes (structure and function) (Table 31.1).
Altered climatic conditions will affect the distribution of forest species and impact disturbances, modify tree growth, and favor the arrival of invasive species; these consequences with therefore have significant economic and ecological implications for human societies. It is thus critical to (1) establish long-term monitoring networks documenting forest functioning across different boreal forest landscapes and environmental conditions; (2) rapidly develop reliable indicators of climate change to identify shifts within forest ecosystems; (3) develop new decision-support tools that can predict future scenarios in forest ecosystems; (4) promote international collaboration and cooperation among boreal countries; and (5) build governance policies (at international, national, and regional levels) that provide a legislative framework to ensure the application of sustainable forest management in the context of the greatest challenge currently facing humanity: climate change.
References
Aakala, T., Pasanen, L., Helama, S., et al. (2018). Multiscale variation in drought controlled historical forest fire activity in the boreal forests of eastern Fennoscandia. Ecological Monographs, 88(1), 74–91. https://doi.org/10.1002/ecm.1276.
Achim, A., Moreau, G., Coops, N. C., et al. (2021). The changing culture of silviculture. Forestry, 95(2), 143–152. https://doi.org/10.1093/forestry/cpab047.
Aerts, R., & Honnay, O. (2011). Forest restoration, biodiversity and ecosystem functioning. BMC Ecology, 11(1), 29. https://doi.org/10.1186/1472-6785-11-29.
Aksenov, D., Karpachevskiy, M., Lloyd, S., et al. (1999). The last of the last: the old-growth forests of boreal Europe. Helsinki: Taiga Rescue Network.
Allen, C. D., Macalady, A. K., Chenchouni, H., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 259(4), 660–684. https://doi.org/10.1016/j.foreco.2009.09.001.
Ameray, A., Bergeron, Y., Valeria, O., et al. (2021). Forest carbon management: A review of silvicultural practices and management strategies across boreal, temperate and tropical forests. Current Forestry Reports, 7(4), 245–266. https://doi.org/10.1007/s40725-021-00151-w.
Anderegg, W. R. L., Hicke, J. A., Fisher, R. A., et al. (2015). Tree mortality from drought, insects, and their interactions in a changing climate. New Phytologist, 208(3), 674–683. https://doi.org/10.1111/nph.13477.
Angelstam, P., Manton, M., Green, M., et al. (2020). Sweden does not meet agreed national and international forest biodiversity targets: A call for adaptive landscape planning. Landscape and Urban Planning, 202, 103838. https://doi.org/10.1016/j.landurbplan.2020.103838.
Aquilué, N., Filotas, É., Craven, D., et al. (2020). Evaluating forest resilience to global threats using functional response traits and network properties. Ecological Applications, 30(5), e02095. https://doi.org/10.1002/eap.2095.
Aquilué, N., Messier, C., Martins, K. T., et al. (2021). A simple-to-use management approach to boost adaptive capacity of forests to global uncertainty. Forest Ecology and Management, 481, 118692. https://doi.org/10.1016/j.foreco.2020.118692.
Artdatabanken. (2020). The Swedish redlist. Rodlistade arter i Sverige. ArtDatabanken SLU, Uppsala.
Asselin, H. (2015). Indigenous forest knowledge. In K. H.-S., Peh, R. Corlett, & Y. Bergeron (Eds.), Routledge handbook of forest ecology (pp. 586–596). London: Earthscan, Routledge.
Astrup, R., Bernier, P. Y., Genet, H., et al. (2018). A sensible climate solution for the boreal forest. Nature Climate Change, 8, 11–12. https://doi.org/10.1038/s41558-017-0043-3.
Aubin, I., Boisvert-Marsh, L., Kebli, H., et al. (2018). Tree vulnerability to climate change: Improving exposure-based assessments using traits as indicators of sensitivity. Ecosphere, 9(2), e02108. https://doi.org/10.1002/ecs2.2108.
Barber, V. A., Juday, G. P., & Finney, B. P. (2000). Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature, 405(6787), 668–673. https://doi.org/10.1038/35015049.
Barnosky, A. D., Hadly, E. A., Bascompte, J., et al. (2012). Approaching a state shift in Earth’s biosphere. Nature, 486(7401), 52–58. https://doi.org/10.1038/nature11018.
Barrette, M., Bélanger, L., De Grandpré, L., et al. (2014). Cumulative effects of chronic deer browsing and clear-cutting on regeneration processes in second-growth white spruce stands. Forest Ecology and Management, 329, 69–78. https://doi.org/10.1016/j.foreco.2014.06.020.
Barrette, M., Bélanger, L., De Grandpré, L., et al. (2017). Demographic disequilibrium caused by canopy gap expansion and recruitment failure triggers forest cover loss. Forest Ecology and Management, 401, 117–124. https://doi.org/10.1016/j.foreco.2017.07.012.
Bartlett, C., Marshall, M., & Marshall, A. (2012). Two-Eyed Seeing and other lessons learned within a co-learning journey of bringing together indigenous and mainstream knowledges and ways of knowing. Journal of Environmental Studies and Sciences, 2(4), 331–340. https://doi.org/10.1007/s13412-012-0086-8.
Beck, P. S. A., Juday, G. P., Alix, C., et al. (2011). Changes in forest productivity across Alaska consistent with biome shift. Ecology Letters, 14(4), 373–379. https://doi.org/10.1111/j.1461-0248.2011.01598.x.
Beguin, J., Tremblay, J. P., Thiffault, N., et al. (2016). Management of forest regeneration in boreal and temperate deer-forest systems: Challenges, guidelines, and research gaps. Ecosphere, 7(10), e01488. https://doi.org/10.1002/ecs2.1488.
Bélisle, A. C., & Asselin, H. (2021). A collaborative typology of boreal Indigenous landscapes. Canadian Journal of Forest Research, 51(9), 1253–1262. https://doi.org/10.1139/cjfr-2020-0369.
Bélisle, A. C., Wapachee, A., & Asselin, H. (2021). From landscape practices to ecosystem services: Landscape valuation in Indigenous contexts. Ecological Economics, 179, 106858. https://doi.org/10.1016/j.ecolecon.2020.106858.
Bellard, C., Thuiller, W., Leroy, B., et al. (2013). Will climate change promote future invasions? Global Change Biology, 19(12), 3740–3748. https://doi.org/10.1111/gcb.12344.
Bengtsson, J., Nilsson, S. G., Franc, A., et al. (2000). Biodiversity, disturbances, ecosystem function and management of European forests. Forest Ecology and Management, 132(1), 39–50. https://doi.org/10.1016/S0378-1127(00)00378-9.
Benoît, P. O., Beisner, B. E., & Solomon, C. T. (2016). Growth rate and abundance of common fishes is negatively related to dissolved organic carbon concentration in lakes. Canadian Journal of Fisheries and Aquatic Sciences, 73(8), 1230–1236. https://doi.org/10.1139/cjfas-2015-0340.
Benoy, G., Cash, K., McCauley, E., et al. (2007). Carbon dynamics in lakes of the boreal forest under a changing climate. Environmental Reviews, 15, 175–189. https://doi.org/10.1139/a07-006.
Bergeron, Y., & Harper, K. A. (2009). Old-growth forests in the Canadian boreal: The exception rather than the rule? In C. Wirth, G. Gleixner, & M. Heimann (Eds.), Old-growth forests. Ecological studies (analysis and synthesis). Berlin, Heidelberg: Springer.
Bergeron, Y., & Leduc, A. (1998). Relationships between change in fire frequency and mortality due to spruce budworm outbreak in the southeastern Canadian boreal forest. Journal of Vegetation Science, 9(4), 492–500. https://doi.org/10.2307/3237264.
Bergeron, Y., Cyr, D., Drever, C. R., et al. (2006). Past, current, and future fire frequencies in Quebec’s commercial forests: Implications for the cumulative effects of harvesting and fire on age-class structure and natural disturbance-based management. Canadian Journal of Forest Research, 36(11), 2737–2744. https://doi.org/10.1139/x06-177.
Bergeron, Y., Vijayakumar, D. B. I. P., Ouzennou, H., et al. (2017). Projections of future forest age class structure under the influence of fire and harvesting: Implications for forest management in the boreal forest of eastern Canada. Forestry, 90(4), 485–495. https://doi.org/10.1093/forestry/cpx022.
Berglund, H., & Kuuluvainen, T. (2021). Representative boreal forest habitats in northern Europe, and a revised model for ecosystem management and biodiversity conservation. Ambio, 50, 1003–1017. https://doi.org/10.1007/s13280-020-01444-3.
Bergman, B. G., Bump, J. K., & Romanski, M. C. (2018). Revisiting the role of aquatic plants in beaver habitat selection. American Midland Naturalist, 179(2), 222–246. https://doi.org/10.1674/0003-0031-179.2.222.
Bescond, H., Fenton, N. J., & Bergeron, Y. (2011). Partial harvests in the boreal forest: Response of the understory vegetation five years after harvest. The Forestry Chronicle, 87(1), 86–98. https://doi.org/10.5558/tfc87086-1.
Blackstock, K. L., Kelly, G. J., & Horsey, B. L. (2007). Developing and applying a framework to evaluate participatory research for sustainability. Ecological Economics, 60(4), 726–742. https://doi.org/10.1016/j.ecolecon.2006.05.014.
Bodin, Ö., & Saura, S. (2010). Ranking individual habitat patches as connectivity providers: Integrating network analysis and patch removal experiments. Ecological Modelling, 221(19), 2393–2405. https://doi.org/10.1016/j.ecolmodel.2010.06.017.
Bohn, F. J., & Huth, A. (2017). The importance of forest structure to biodiversity-productivity relationships. Royal Society Open Science, 4(1), 160521. https://doi.org/10.1098/rsos.160521.
Borowik, T., Ratkiewicz, M., Maślanko, W., et al. (2020). Too hot to handle: Summer space use shift in a cold-adapted ungulate at the edge of its range. Landscape Ecology, 35(6), 1341–1351. https://doi.org/10.1007/s10980-020-01018-4.
Bose, A. K., Harvey, B. D., Brais, S., et al. (2014). Constraints to partial cutting in the boreal forest of Canada in the context of natural disturbance-based management: A review. Forestry, 87(1), 11–28. https://doi.org/10.1093/forestry/cpt047.
Boserup, E. (1981). Population and technological change: A study of long-term trends. University of Chicago Press.
Bottero, A., D’Amato, A. W., Palik, B. J., et al. (2017). Density-dependent vulnerability of forest ecosystems to drought. Journal of Applied Ecology, 54(6), 1605–1614. https://doi.org/10.1111/1365-2664.12847.
Bouchard, M., Aquilué, N., Périé, C., et al. (2019). Tree species persistence under warming conditions: A key driver of forest response to climate change. Forest Ecology and Management, 442, 96–104. https://doi.org/10.1016/j.foreco.2019.03.040.
Boucher, D., De Grandpré, L., Kneeshaw, D. D., et al. (2015). Effects of 80 years of forest management on landscape structure and pattern in the eastern Canadian boreal forest. Landscape Ecology, 30, 1913–1929. https://doi.org/10.1007/s10980-015-0220-6.
Boucher, D., Boulanger, Y., Aubin, I., et al. (2018). Current and projected cumulative impacts of fire, drought, and insects on timber volumes across Canada. Ecological Applications, 28(5), 1245–1259. https://doi.org/10.1002/eap.1724.
Boucher, Y., & Grondin, P. (2012). Impact of logging and natural stand-replacing disturbances on high-elevation boreal landscape dynamics (1950–2005) in eastern Canada. Forest Ecology and Management, 263, 229–239. https://doi.org/10.1016/j.foreco.2011.09.012.
Boucher, Y., Arseneault, D., Sirois, L., et al. (2009). Logging pattern and landscape changes over the last century at the boreal and deciduous forest transition in Eastern Canada. Landscape Ecology, 24(2), 171–184. https://doi.org/10.1007/s10980-008-9294-8.
Boucher, Y., Perrault-Hébert, M., Fournier, R., et al. (2017). Cumulative patterns of logging and fire (1940–2009): Consequences on the structure of the eastern Canadian boreal forest. Landscape Ecology, 32(2), 361–375. https://doi.org/10.1007/s10980-016-0448-9.
Boulanger, Y., Taylor, A. R., Price, D. T., et al. (2016). Climate change impacts on forest landscapes along the Canadian southern boreal forest transition zone. Landscape Ecology, 32(7), 1415–1431. https://doi.org/10.1007/s10980-016-0421-7.
Boulanger, Y., Arseneault, D., Boucher, Y., et al. (2019). Climate change will affect the ability of forest management to reduce gaps between current and presettlement forest composition in southeastern Canada. Landscape Ecology, 34(1), 159–174. https://doi.org/10.1007/s10980-018-0761-6.
Brais, S., Harvey, B. D., Bergeron, Y., et al. (2004). Testing forest ecosystem management in boreal mixedwoods of northwestern Quebec: Initial response of aspen stands to different levels of harvesting. Canadian Journal of Forest Research, 34(2), 431–446. https://doi.org/10.1139/x03-144.
Brienen, R. J. W., Caldwell, L., Duchesne, L., et al. (2020). Forest carbon sink neutralized by pervasive growth-lifespan trade-offs. Nature Communications, 11(1), 4241. https://doi.org/10.1038/s41467-020-17966-z.
Brokaw, N. V. L., & Lent, R. A. (1999). Vertical structure. In M. Hunter (Ed.), Maintaining biodiversity in forest ecosystems (pp. 373–399). Cambridge University Press.
Burton, P. J., Messier, C., Weetman, G., et al. (2003). The current state of boreal forestry and the drive for change. In P. J. Burton, C. Messier, D. W. Smith, & W. L. Adamowicz (Eds.), Towards sustainable management of the boreal forest (pp. 1–40). NRC Research Press.
Cadieux, P., Boulanger, Y., Cyr, D., et al. (2020). Projected effects of climate change on boreal bird community accentuated by anthropogenic disturbances in western boreal forest, Canada. Diversity and Distributions, 26(6), 668–682. https://doi.org/10.1111/ddi.13057.
Cadotte, M. W., Carscadden, K., & Mirotchnick, N. (2011). Beyond species: Functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 48(5), 1079–1087. https://doi.org/10.1111/j.1365-2664.2011.02048.x.
Canadian Institutes of Health Research, Natural Sciences and Engineering Council of Canada, Social Sciences and Humanities Research Council. (2018). Tri-council policy statement ethical conduct for research involving humans Secretariat on Responsible Conduct of Research, Ottawa, p. 231.
Canelles, Q., Aquilué, N., James, P. M. A., et al. (2021). Global review on interactions between insect pests and other forest disturbances. Landscape Ecology, 36(4), 945–972. https://doi.org/10.1007/s10980-021-01209-7.
Cappuccino, N., Lavertu, D., Bergeron, Y., et al. (1998). Spruce budworm impact, abundance and parasitism rate in a patchy landscape. Oecologia, 114(2), 236–242. https://doi.org/10.1007/s004420050441.
Carcaillet, C., Bergeron, Y., Richard, P. J. H., et al. (2001). Change of fire frequency in the eastern Canadian boreal forests during the Holocene: Does vegetation composition or climate trigger the fire regime? Journal of Ecology, 89(6), 930–946. https://doi.org/10.1046/j.0022-0477.2001.00614.x https://doi.org/10.1111/j.1365-2745.2001.00614.x.
Carignan, R., & Steedman, R. J. (2000). Impacts of major watershed perturbations on aquatic ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 57(S2), 1–4. https://doi.org/10.1139/f00-172.
Carnus, J. M., Parrotta, J., Brockerhoff, E., et al. (2006). Planted Forests and Biodiversity. Journal of Forestry, 104(2), 65–77. https://doi.org/10.1093/jof/104.2.65.
Castleden, H. E., Hart, C., Harper, S., et al. (2017). Implementing indigenous and western knowledge systems in water research and management (Part 1): A systematic realist review to inform water policy and governance in Canada. International Indigenous Policy Journal, 8(4):1–33 https://doi.org/10.18584/iipj.2017.8.4.6.
Chazdon, R. L. (2018). Protecting intact forests requires holistic approaches. Nature Ecology and Evolution, 2(6), 915–915. https://doi.org/10.1038/s41559-018-0546-y.
Cole, J. J., Prairie, Y. T., Caraco, N. F., et al. (2007). Plumbing the global carbon cycle: Integrating inland waters into the terrestrial carbon budget. Ecosystems, 10(1), 172–185. https://doi.org/10.1007/s10021-006-9013-8.
Cook-Patton, S. C., Leavitt, S. M., Gibbs, D., et al. (2020). Mapping carbon accumulation potential from global natural forest regrowth. Nature, 585(7826), 545–550. https://doi.org/10.1038/s41586-020-2686-x.
Cornelissen, J. H. C., Lavorel, S., Garnier, E., et al. (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 51(4), 335–380. https://doi.org/10.1071/BT02124.
Culbert, P. D., Radeloff, V. C., Flather, C. H., et al. (2013). The influence of vertical and horizontal habit at structure on nationwide patterns of avian biodiversity. The Auk, 130(4), 656–665. https://doi.org/10.1525/auk.2013.13007.
Cullingham, C. I., Cooke, J. E. K., Dang, S., et al. (2011). Mountain pine beetle host-range expansion threatens the boreal forest. Molecular Ecology, 20(10), 2157–2171. https://doi.org/10.1111/j.1365-294X.2011.05086.x.
Cumming, S. G., Schmiegelow, F. K. A., & Burton, P. J. (2000). Gap dynamics in boreal aspen stands: Is the forest older than we think? Ecological Applications, 10(3), 744–759. https://doi.org/10.1890/1051-0761(2000)010[0744:GDIBAS]2.0.CO;2.
Cyr, D., Gauthier, S., Bergeron, Y., et al. (2009). Forest management is driving the eastern North American boreal forest outside its natural range of variability. Frontiers in Ecology and the Environment, 7(10), 519–524. https://doi.org/10.1890/080088.
D'Amato, A. W., Bradford, J. B., Fraver, S., et al. (2011). Forest management for mitigation and adaptation to climate change: Insights from long-term silviculture experiments. Forest Ecology and Management, 262, 803–816. https://doi.org/10.1016/j.foreco.2011.05.014.
D’Amato, A. W., Bradford, J. B., Fraver, S., et al. (2013). Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems. Ecological Applications, 23(8), 1735–1742. https://doi.org/10.1890/13-0677.1.
D’Orangeville, L., Duchesne, L., Houle, D., et al. (2016). Northeastern North America as a potential refugium for boreal forests in a warming climate. Science, 352(6292), 1452–1455. https://doi.org/10.1126/science.aaf4951.
D’Orangeville, L., Houle, D., Duchesne, L., et al. (2018). Beneficial effects of climate warming on boreal tree growth may be transitory. Nature Communications, 9(1), 3213. https://doi.org/10.1038/s41467-018-05705-4.
Dam Lam, R., Gasparatos, A., Chakraborty, S., et al. (2019). Multiple values and knowledge integration in indigenous coastal and marine social-ecological systems research: A systematic review. Ecosystem Services, 37, 100910. https://doi.org/10.1016/j.ecoser.2019.100910.
Daniel, C. J., Ter-Mikaelian, M. T., Wotton, B. M., et al. (2017). Incorporating uncertainty into forest management planning: Timber harvest, wildfire and climate change in the boreal forest. Forest Ecology and Management, 400, 542–554. https://doi.org/10.1016/j.foreco.2017.06.039.
Danneyrolles, V., Dupuis, S., Fortin, G., et al. (2019). Stronger influence of anthropogenic disturbance than climate change on century-scale compositional changes in northern forests. Nature Communications, 10(1), 1265. https://doi.org/10.1038/s41467-019-09265-z.
Davidson-Hunt, I., & Berkes, F. (2003). Learning as you journey: Anishinaabe perception of social-ecological environments and adaptive learning. Conservation Ecology, 8(1):art5. https://doi.org/10.5751/es-00587-080105.
Dawe, K. L., & Boutin, S. (2016). Climate change is the primary driver of white-tailed deer (Odocoileus virginianus) range expansion at the northern extent of its range; land use is secondary. Ecology and Evolution, 6(18), 6435–6451. https://doi.org/10.1002/ece3.2316.
De Grandpré, L., Waldron, K., Bouchard, M., et al. (2018). Incorporating insect and wind disturbances in a natural disturbance-based management framework for the boreal forest. Forests, 9(8), 471. https://doi.org/10.3390/f9080471.
De Groot, W. J., Cantin, A. S., Flannigan, M. D., et al. (2013a). A comparison of Canadian and Russian boreal forest fire regimes. Forest Ecology and Management, 294, 23–34. https://doi.org/10.1016/j.foreco.2012.07.033.
De Groot, W. J., Flannigan, M. D., & Cantin, A. S. (2013b). Climate change impacts on future boreal fire regimes. Forest Ecology and Management, 294, 35–44. https://doi.org/10.1016/j.foreco.2012.09.027.
De Vriendt, L., Lavoie, S., Barrette, M., et al. (2021). From delayed succession to alternative successional trajectory: How different moose browsing pressures contribute to forest dynamics following clear-cutting. Journal of Vegetation Science, 32(1), e12945. https://doi.org/10.1111/jvs.12945.
Díaz-Yáñez, O., Arias-Rodil, M., Mola-Yudego, B., et al. (2019). Simulating the effects of wind and snow damage on the optimal management of Norwegian spruce forests. Forestry, 92(4), 406–416. https://doi.org/10.1093/forestry/cpz031.
Dobson, A. P., Bradshaw, A. D., & Baker, A. J. M. (1997). Hopes for the future: Restoration ecology and conservation biology. Science, 277(5325), 515–522. https://doi.org/10.1126/science.277.5325.515.
Dondina, O., Orioli, V., D’Occhio, P., et al. (2017). How does forest species specialization affect the application of the island biogeography theory in fragmented landscapes? Journal of Biogeography, 44(5), 1041–1052. https://doi.org/10.1111/jbi.12827.
Dou, H., Jiang, G., Stott, P., et al. (2013). Climate change impacts population dynamics and distribution shift of moose (Alces alces) in Heilongjiang Province of China. Ecological Research, 28(4), 625–632. https://doi.org/10.1007/s11284-013-1054-9.
Duchesne, L., & Ouimet, R. (2007). Une histoire de perturbations! Les changements de composition dans la forêt du Québec méridional au cours des 30 dernières années. Québec: Direction de la recherche forestière, ministère des Forêts, de la Faune et des Parcs.
Dukes, J. S., Pontius, J., Orwig, D., et al. (2009). Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Canadian Journal of Forest Research, 39, 231–248. https://doi.org/10.1139/X08-171.
Dunn, A. L., Barford, C. C., Wofsy, S. C., et al. (2007). A long-term record of carbon exchange in a boreal black spruce forest: Means, responses to interannual variability, and decadal trends. Global Change Biology, 13(3), 577–590. https://doi.org/10.1111/j.1365-2486.2006.01221.x.
Dupuis, S., Danneyrolles, V., Laflamme, J., et al. (2020). Forest transformation following European settlement in the Saguenay-Lac-St-Jean valley in eastern Québec, Canada. Frontiers in Ecology and Evolution, 8(August), 1–13. https://doi.org/10.3389/fevo.2020.00257.
Eden, J. M., Krikken, F., & Drobyshev, I. (2020). An empirical prediction approach for seasonal fire risk in the boreal forests. International Journal of Climatology, 40(5), 2732–2744. https://doi.org/10.1002/joc.6363.
Eggers, S., & Low, M. (2014). Differential demographic responses of sympatric Parids to vegetation management in boreal forest. Forest Ecology and Management, 319, 169–175. https://doi.org/10.1016/j.foreco.2014.02.019.
Elmqvist, T., Folke, C., Nystrom, M., et al. (2003). Response diversity, ecosystem change, and resilience. Frontiers in Ecology and the Environment, 1, 488–494. https://doi.org/10.1890/1540-9295(2003)001[0488:RDECAR]2.0.CO;2.
Ericksen, P., & Woodley, E. (2005). Using multiple knowledge systems: Benefits and challenges. In A. M. E. Assessment (Ed.), Ecosystems and human well-being (pp. 85–117). World Resources Institute.
Esseen, P. A., Ehnström, B., Ericson, L., et al. (1992). Boreal forests—the focal habitats of Fennoscandia. In L. Hansson (Ed.), Ecological principles of nature conservation (pp. 252–325). Elsevier Applied Science.
Esseen, P. A., Ehnström, B., Ericson, L., et al. (1997). Boreal forests. Ecological Bulletins, 46, 16–47.
Esseen, P. A., Hedström Ringvall, A., Harper, K. A., et al. (2016). Factors driving structure of natural and anthropogenic forest edges from temperate to boreal ecosystems. Journal of Vegetation Science, 27(3), 482–492. https://doi.org/10.1111/jvs.12387.
European Commission (EC). (2013). Communication from the Commission to the European Parliament, the Council, The European Economic and Social Committee and the Committee of the Regions: Green infrastructure (GI)—Enhancing Europe’s natural capital: COM/2013/0249. Brussels: European Commission.
Eyvindson, K., Duflot, R., Triviño, M., et al. (2021). High boreal forest multifunctionality requires continuous cover forestry as a dominant management. Land Use Policy, 100, 104918. https://doi.org/10.1016/j.landusepol.2020.104918.
Fatichi, S., Pappas, C., Zscheischler, J., et al. (2019). Modelling carbon sources and sinks in terrestrial vegetation. New Phytologist, 221(2), 652–668. https://doi.org/10.1111/nph.15451.
Feldman, M. J., Girona, M. M., Grosbois, G., et al. (2020). Why do beavers leave home? Lodge abandonment in an invasive population in Patagonia. Forests, 11(11), 1161. https://doi.org/10.3390/f11111161.
Fenton, N., Simard, M., & Bergeron, Y. (2009). Emulating natural disturbances: The role of silviculture in creating even-aged and complex structures in the black spruce boreal forest of eastern North America. Journal of Forest Research, 14, 258–267. https://doi.org/10.1007/s10310-009-0134-8.
Fenton, N. J., Imbeau, L., Work, T., et al. (2013). Lessons learned from 12 years of ecological research on partial cuts in black spruce forests of northwestern Québec. The Forestry Chronicle, 89(03), 350–359. https://doi.org/10.5558/tfc2013-065.
Feser, F., Barcikowska, M., Krueger, O., et al. (2015). Storminess over the North Atlantic and northwestern Europe—A review. Quarterly Journal Royal Meteorological Society, 141(687), 350–382. https://doi.org/10.1002/qj.2364.
Filbee-Dexter, K., Pittman, J., Haig, H. A., et al. (2017). Ecological surprise: Concept, synthesis, and social dimensions. Ecosphere, 8(12), e02005. https://doi.org/10.1002/ecs2.2005.
Fischer, J., & Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: A synthesis. Global Ecology and Biogeography, 16(3), 265–280. https://doi.org/10.1111/j.1466-8238.2007.00287.x.
Flannigan, M. D., Wotton, B. M., Marshall, G. A., et al. (2016). Fuel moisture sensitivity to temperature and precipitation: Climate change implications. Climatic Change, 134(1–2), 59–71. https://doi.org/10.1007/s10584-015-1521-0.
Foley, J. A., DeFries, R., Asner, G. P., et al. (2005). Global consequences of land use. Science, 309(5734), 570–574. https://doi.org/10.1126/science.1111772.
Folke, C., Carpenter, S., Walker, B., et al. (2004). Regime shifts, resilience, and biodiversity in ecosystem management. Annual Review of Ecology, Evolution, and Systematics, 35(1), 557–581. https://doi.org/10.1146/annurev.ecolsys.35.021103.105711.
Food and Agriculture Organization of the United Nations (FAO). (2009). State of the world’s forests 2009 (p. 168). Electronic Publishing Policy and Support Branch, Communication Division.
Food and Agriculture Organization of the United Nations (FAO). (2011). State of the world’s forest 2011. Electronic Publishing Policy and Support Branch.
Food and Agriculture Organization of the United Nations (FAO). (2014). State of the world’s forest: Enhancing the socioeconomic benefits from forests. Electronic Publishing Policy and Support Branch.
Food and Agriculture Organization of the United Nations (FAO). (2016). State of the world’s forest 2016. Electronic Publishing Policy and Support Branch.
Forest Europe. (2015). State of Europe's forests 2015. In Ministerial Conference on the Protection of Forests in Europe, Madrid: Forest Europe, Liaison Unit Madrid.
Franklin, J. F., Berg, D. F., Thornburg, D., et al. (1997). Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In K. A. Kohm & J. F. Franklin (Eds.), Creating a forestry for the 21st century: The science of ecosystem management (pp. 111–140). Island Press.
Franklin, J. F., Spies, T. A., Pelt, R. V., et al. (2002). Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. Forest Ecology and Management, 155(1–3), 399–423. https://doi.org/10.1016/S0378-1127(01)00575-8.
Fraser, D., Chavez, E. R., & Paloheimo, J. E. (1984). Aquatic feeding by moose: Selection of plant species and feeding areas in relation to plant chemical composition and characteristics of lakes. Canadian Journal of Zoology, 62(1), 80–87. https://doi.org/10.1139/z84-014.
Friend, A. D., Lucht, W., Rademacher, T. T., et al. (2014). Carbon residence time dominates uncertainty in terrestrial vegetation responses to future climate and atmospheric CO2. Proceedings of the National Academy of Sciences of the United States of America, 111(9), 3280. https://doi.org/10.1073/pnas.1222477110.
Fritts, H. C. (2001). Tree rings and climate (p. 584). Caldwell: Blackburn Press.
Fuentes, L., Asselin, H., Bélisle, A. C., et al. (2020). Impacts of environmental changes on well-being in Indigenous communities in Eastern Canada. International Journal of Environmental Research and Public Health, 17(2), 637. https://doi.org/10.3390/ijerph17020637.
Gagné, L., Lavoie, L., & Binot, J. M. (2012). Growth and mechanical properties of wood after commercial thinning in a 32-year-old white spruce (Picea glauca) plantation. Canadian Journal of Forest Research, 42(2), 291–302. https://doi.org/10.1139/X11-181.
Garcia, E., & Carignan, R. (2000). Mercury concentrations in northern pike (Esox lucius) from boreal lakes with logged, burned, or undisturbed catchments. Canadian Journal of Fisheries and Aquatic Sciences, 57, 129–135. https://doi.org/10.1139/f00-126.
Gauthier, S., Villancourt, M. A., Leduc, A., et al. (2009). Ecosystem management in the boreal forest. Presses de l’Université de Québec.
Gauthier, S., Bernier, P. Y., Boulanger, Y., et al. (2015a). Vulnerability of timber supply to projected changes in fire regime in Canada’s managed forests. Canadian Journal of Forest Research, 45, 1439–1447. https://doi.org/10.1139/cjfr-2015-0079.
Gauthier, S., Bernier, P., Kuuluvainen, T., et al. (2015b). Boreal forest health and global change. Science, 349, 819–822. https://doi.org/10.1126/science.aaa9092.
Gilarranz, L. J., Rayfield, B., Liñán-Cembrano, G., et al. (2017). Effects of network modularity on the spread of perturbation impact in experimental metapopulations. Science, 357(6347), 199–201. https://doi.org/10.1126/science.aal4122.
Gilchrist, G., Mallory, M., & Merkel, F. (2005). Can local ecological knowledge contribute to wildlife management? Case studies of migratory birds. Ecology and Society, 10(1), 12. https://doi.org/10.5751/ES-01275-100120.
Gingras, B., Slattery, S., Smith, K., et al. (2018). Boreal wetlands of Canada and the United States of America. In C. M. Finlayson, G. R. Milton, R. C. Prentice, & N. C. Davidson (Eds.), The wetland book: II: Distribution, description, and conservation (pp. 521–542). Springer.
Glaz, P., Sirois, P., Archambault, P., et al. (2014). Impact of forest harvesting on trophic structure of eastern Canadian Boreal Shield lakes: Insights from stable isotope analyses. PLoS ONE, 9(4), e96143. https://doi.org/10.1371/journal.pone.0096143.
Glaz, P., Gagné, J. P., Archambault, P., et al. (2015). Impact of forest harvesting on water quality and fluorescence characteristics of dissolved organic matter in eastern Canadian Boreal Shield lakes in summer. Biogeosciences, 12(23), 6999–7011. https://doi.org/10.5194/bg-12-6999-2015.
Gonzalès, R., & Parrott, L. (2012). Network theory in the assessment of the sustainability of social-ecological systems. Geography Compass, 6(2), 76–88. https://doi.org/10.1111/j.1749-8198.2011.00470.x.
Gosse, J., Hermanutz, L., McLaren, B., et al. (2011). Degradation of boreal forests by nonnative herbivores in Newfoundland’s national parks: Recommendations for ecosystem restoration. Natural Areas Journal, 31(4), 331–339. https://doi.org/10.3375/043.031.0403.
Grant, G. E., Tague, C. L., & Allen, C. D. (2013). Watering the forest for the trees: An emerging priority for managing water in forest landscapes. Frontiers in Ecology and the Environment, 11(6), 314–321. https://doi.org/10.1890/120209.
Gregow, H., Peltola, H., Laapas, M., et al. (2011). Combined occurrence of wind, snow loading and soil frost with implications for risks to forestry in Finland under the current and changing climatic conditions. Silva Fennica, 45(1), 30. https://doi.org/10.14214/sf.30.
Grenon, F., Jetté, J., & Leblanc, M. (2010). Manuel de référence pour l’aménagement écosystémique des forêts au Québec–Module 1-Fondements et démarche de la mise en oeuvre. Québec: CERFO, ministère des Ressources naturelles et de la Faune.
Grondin, P., Gauthier, S., Poirier, V., et al. (2018). Have some landscapes in the eastern Canadian boreal forest moved beyond their natural range of variability? Forest Ecosystems, 5(1), 30. https://doi.org/10.1186/s40663-018-0148-9.
Grosbois, G., Vachon, D., Del Giorgio, P. A., et al. (2020). Efficiency of crustacean zooplankton in transferring allochthonous carbon in a boreal lake. Ecology, 101(6), e03013. https://doi.org/10.1002/ecy.3013.
Guindon, L., Bernier, P., Gauthier, S., et al. (2018). Missing forest cover gains in boreal forests explained. Ecosphere, 9(1), e02094. https://doi.org/10.1002/ecs2.2094.
Gunderson, L. (2015). Lessons from adaptive management: Obstacles and outcomes. In C. R. Allen & A. S. Garmestani (Eds.), Adaptive management of social-ecological systems (pp. 27–38). Springer.
Gustafsson, L., Bauhus, J., Asbeck, T., et al. (2020). Retention as an integrated biodiversity conservation approach for continuous-cover forestry in Europe. Ambio, 49(1), 85–97. https://doi.org/10.1007/s13280-019-01190-1.
Haddad, N. M., Brudvig, L. A., Clobert, J., et al. (2015). Habitat fragmentation and its lasting impact on Earth’s ecosystems. Science Advances, 1(2), e1500052. https://doi.org/10.1126/sciadv.1500052.
Haeussler, S., & Kneeshaw, D. (2003). Comparing forest management to natural processes. In P. J. Burton, C. Messier, D. W. Smith, & W. L. Adamowicz (Eds.), Towards sustainable management of the boreal forest (pp. 307–368). NRC Research Press.
Hägglund, R., Hekkala, A. M., Hjältén, J., et al. (2015). Positive effects of ecological restoration on rare and threatened flat bugs (Heteroptera: Aradidae). Journal of Insect Conservation, 19(6), 1089–1099. https://doi.org/10.1007/s10841-015-9824-z.
Halme, P., Allen, K. A., Auniņš, A., et al. (2013). Challenges of ecological restoration: Lessons from forests in northern Europe. Biological Conservation, 167, 248–256. https://doi.org/10.1016/j.biocon.2013.08.029.
Harper, K. A., Macdonald, S. E., Mayerhofer, M. S., et al. (2015). Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. Journal of Ecology, 103, 550–562. https://doi.org/10.1111/1365-2745.12398.
Harris, J. A., Hobbs, R. J., Higgs, E., et al. (2006). Ecological restoration and global climate change. Restoration Ecology, 14(2), 170–176. https://doi.org/10.1111/j.1526-100X.2006.00136.x.
Hassan, R. M., Scholes, R. J., Ash, N., et al. (2005). Ecosystems and human well-being : Current state and trends : Findings of the Condition and Trends Working Group of the Millennium Ecosystem Assessment. Island Press.
Heikkala, O., Seibold, S., Koivula, M., et al. (2016). Retention forestry and prescribed burning result in functionally different saproxylic beetle assemblages than clear-cutting. Forest Ecology and Management, 359, 51–58. https://doi.org/10.1016/j.foreco.2015.09.043.
Heino, M., Kummu, M., Makkonen, M., et al. (2015). Forest loss in protected areas and intact forest landscapes: A global analysis. PLoS ONE, 10(10), e0138918. https://doi.org/10.1371/journal.pone.0138918.
Hekkala, A. M., Tarvainen, O., & Tolvanen, A. (2014). Dynamics of understory vegetation after restoration of natural characteristics in the boreal forests in Finland. Forest Ecology and Management, 330, 55–66. https://doi.org/10.1016/j.foreco.2014.07.001.
Heller, N. E., & Zavaleta, E. S. (2009). Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biological Conservation, 142(1), 14–32. https://doi.org/10.1016/j.biocon.2008.10.006.
Hernández-Rodríguez, E., Escalera-Vázquez, L. H., García-Ávila, D., et al. (2021). Reduced-impact logging maintain high moss diversity in temperate forests. Forests, 12(4), 383. https://doi.org/10.3390/f12040383.
Hidding, B., Tremblay, J. P., & Côté, S. D. (2013). A large herbivore triggers alternative successional trajectories in the boreal forest. Ecology, 94(12), 2852–2860. https://doi.org/10.1890/12-2015.1.
Hilty, J., Muller, B., Pantin, F., et al. (2021). Plant growth: The what, the how, and the why. New Phytologist, 232(1), 25–41. https://doi.org/10.1111/nph.17610.
Hjältén, J., Hägglund, R., Löfroth, T., et al. (2017). Forest restoration by burning and gap cutting of voluntary set-asides yield distinct immediate effects on saproxylic beetles. Biodiversity and Conservation, 26(7), 1623–1640. https://doi.org/10.1007/s10531-017-1321-0.
Hlásny, T., König, L., Krokene, P., et al. (2021). Bark beetle outbreaks in Europe: State of knowledge and ways forward for management. Current Forestry Reports, 7(3), 138–165. https://doi.org/10.1007/s40725-021-00142-x.
Hof, A. R., Montoro Girona, M., Fortin, M.-J., et al. (2021). Editorial: Using landscape simulation models to help balance conflicting goals in changing forests. Frontiers in Ecology and Evolution. https://doi.org/10.3389/fevo.2021.795736.
Honnay, O., Piessens, K., Van Landuyt, W., et al. (2003). Satellite based land use and landscape complexity indices as predictors for regional plant species diversity. Landscape and Urban Planning, 63(4), 241–250. https://doi.org/10.1016/S0169-2046(02)00194-9.
Hyvärinen, E., Juslén, A., Kemppainen, E., et al. (2019). Suomen lajien uhanalaisuus—Punainen kirja 2019/The 2019 Red List of Finnish Species. Ympäristöministeriö and Suomen ympäristökeskus/Ministry of the Environment and Finnish Environment Institute.
Intergovernmental Panel on Climate Change (IPCC). (2014). Climate Change 2014: Synthesis report. Contribution of Working Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change (p. 151). Geneva: IPCC.
Ivantsova, E. D., Pyzhev, A. I., & Zander, E. V. (2019). Economic consequences of insect pests outbreaks in boreal forests: A literature review. Journal of Siberian Federal University. Humanities & Social Sciences, 12(4), 627–642. https://doi.org/10.17516/1997-1370-0417.
Jackson, J. B. C., Kirby, M. X., Berger, W. H., et al. (2001). Historical overfishing and the recent collapse of coastal ecosystems. Science, 293(5530), 629–637. https://doi.org/10.1126/science.1059199.
James, P. M. A., Robert, L. E., Wotton, B. M., et al. (2017). Lagged cumulative spruce budworm defoliation affects the risk of fire ignition in Ontario, Canada. Ecological Applications, 27(2), 532–544. https://doi.org/10.1002/eap.1463.
Jasinski, J. P. P., & Payette, S. (2005). The creation of alternative stable states in the southern boreal forest, Québec, Canada. Ecological Monographs, 75(4), 561–583. https://doi.org/10.1890/04-1621.
Johnstone, J. F., Allen, C. D., Franklin, J. F., et al. (2016). Changing disturbance regimes, ecological memory, and forest resilience. Frontiers in Ecology and the Environment, 14(7), 369–378. https://doi.org/10.1002/fee.1311.
Jones, K. R., Venter, O., Fuller, R. A., et al. (2018). One-third of global protected land is under intense human pressure. Science, 360(6390), 788–791. https://doi.org/10.1126/science.aap9565.
Jonsson, B. G., & Siitonen, J. (2012). Dead wood and sustainable forest management. In B. G. Jonsson, J. N. Stokland, & J. Siitonen (Eds.), Biodiversity in dead wood (pp. 302–337). Cambridge: Cambridge University Press.
Jonsson, B. G., Svensson, J., Mikusiński, G., et al. (2019). European Union’s last intact forest landscapes are at a value chain crossroad between multiple use and intensified wood production. Forests, 10(7), 564. https://doi.org/10.3390/f10070564.
Junninen, K., & Komonen, A. (2011). Conservation ecology of boreal polypores: A review. Biological Conservation, 144(1), 11–20. https://doi.org/10.1016/j.biocon.2010.07.010.
Kauppi, P. E., Posch, M., & Pirinen, P. (2014). Large impacts of climatic warming on growth of boreal forests since 1960. PLoS ONE, 9(11), e111340. https://doi.org/10.1371/journal.pone.0111340.
Keeton, W. S. (2018). Source or sink? Carbon dynamics in eastern old-growth forests and their role in climate change mitigation. In A. M. Barton & W. S. Keeton (Eds.), Ecology and recovery of eastern old-growth forests (pp. 267–288). Island Press/Center for Resource Economics.
Kim, S., Axelsson, E. P., Girona, M. M., et al. (2021). Continuous-cover forestry maintains soil fungal communities in Norway spruce dominated boreal forests. Forest Ecology and Management, 480, 118659. https://doi.org/10.1016/j.foreco.2020.118659.
Kimmins, J. P. (1997). Forest ecology—a foundation for sustainable forest management and environmental ethics in forestry. Prentice Hall.
Klaus, M., Geibrink, E., Jonsson, A., et al. (2018). Greenhouse gas emissions from boreal inland waters unchanged after forest harvesting. Biogeosciences, 15(18), 5575–5594. https://doi.org/10.5194/bg-15-5575-2018.
Kneeshaw, D. D., & Gauthier, S. (2003). Old growth in the boreal forest: A dynamic perspective at the stand and landscape level. Environmental Reviews, 11, S99–S114. https://doi.org/10.1139/a03-010.
Korhonen, K., Ahola, A., Heikkinen, J., et al. (2021). Forests of Finland 2014–2018 and their development 1921–2018. Silva Fennica, 55(5), 10662. https://doi.org/10.14214/sf.10662.
Körner, C. (2017). A matter of tree longevity. Science, 355(6321), 130–131. https://doi.org/10.1126/science.aal2449.
Kramer, K., Degen, B., Buschbom, J., et al. (2010). Modelling exploration of the future of European beech (Fagus sylvatica L.) under climate change-Range, abundance, genetic diversity and adaptive response. Forest Ecology and Management, 259(11), 2213–2222. https://doi.org/10.1016/j.foreco.2009.12.023.
Krikken, F., Lehner, F., Haustein, K., et al. (2021). Attribution of the role of climate change in the forest fires in Sweden 2018. Natural Hazards and Earth System Sciences, 21(7), 2169–2179. https://doi.org/10.5194/nhess-21-2169-2021.
Kritzberg, E. S., Hasselquist, E. M., Škerlep, M., et al. (2020). Browning of freshwaters: Consequences to ecosystem services, underlying drivers, and potential mitigation measures. Ambio, 49(2), 375–390. https://doi.org/10.1007/s13280-019-01227-5.
Kulha, N., Pasanen, L., Holmström, L., et al. (2019). At what scales and why does forest structure vary in naturally dynamic boreal forests? An analysis of forest landscapes on two continents. Ecosystems, 22(4), 709–724. https://doi.org/10.1007/s10021-018-0297-2.
Kulha, N., Pasanen, L., Holmström, L., et al. (2020). The structure of boreal old-growth forests changes at multiple spatial scales over decades. Landscape Ecology, 35(4), 843–858. https://doi.org/10.1007/s10980-020-00979-w.
Kurz, W. A., Stinson, G., Rampley, G. J., et al. (2008). Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proceedings of National Academy of Sciences of the United States of America, 105(5), 1551–1555. https://doi.org/10.1073/pnas.0708133105.
Kuuluvainen, T. (2002). Natural variability of forests as a reference for restoring and managing biological diversity in boreal Fennoscandia. Silva Fennica, 36(1), 552. https://doi.org/10.14214/sf.552.
Kuuluvainen, T. (2009). Forest management and biodiversity conservation based on natural ecosystem dynamics in northern Europe: The complexity challenge. Ambio, 38(6), 309–315. https://doi.org/10.1579/08-A-490.1.
Kuuluvainen, T., & Grenfell, R. (2012). Natural disturbance emulation in boreal forest ecosystem management—theories, strategies, and a comparison with conventional even-aged management. Canadian Journal of Forest Research, 42(7), 1185–1203. https://doi.org/10.1139/x2012-064.
Kuuluvainen, T., & Siitonen, J. (2013). Fennoscandian boreal forests as complex adaptive systems. Properties, management challenges and opportunities. In C. Messier, K. J. Puettman, & K. D. Coates (Eds.), Managing forests as complex adaptive systems. Building resilience to the challenge of global change (pp. 244–268). London: The Earthscan Forest Library, Routledge.
Kuuluvainen, T., & Gauthier, S. (2018). Young and old forest in the boreal: Critical stages of ecosystem dynamics and management under global change. Forestry Ecosystems, 5(1), 26. https://doi.org/10.1186/s40663-018-0142-2.
Kuuluvainen, T., Hofgaard, A., Aakala, T., et al. (2017). North Fennoscandian mountain forests: History, composition, disturbance dynamics and the unpredictable future. Forest Ecology and Management, 385, 140–149. https://doi.org/10.1016/j.foreco.2016.11.031.
Kuuluvainen, T., Lindberg, H., Vanha-Majamaa, I., et al. (2019). Low-level retention forestry, certification, and biodiversity: Case Finland. Ecological Processes, 8(1), 47. https://doi.org/10.1186/s13717-019-0198-0.
Kuuluvainen, T., Angelstam, P., Frelich, L., et al. (2021). Natural disturbance-based forest management: Moving beyond retention and continuous-cover forestry. Frontiers in Forests and Global Change, 4(24), 629020. https://doi.org/10.3389/ffgc.2021.629020.
Labrecque-Foy, J.-P., Morin, H., & Girona, M. M. (2020). Dynamics of territorial occupation by North American beavers in Canadian boreal forests: A novel dendroecological approach. Forests, 11(2), 221. https://doi.org/10.3390/f11020221.
Lamontagne, S., Carignan, R., D’Arcy, P., et al. (2000). Element export in runoff from eastern Canadian Boreal Shield drainage basins following forest harvesting and wildfires. Canadian Journal of Fisheries and Aquatic Sciences, 57(S2), 118–128. https://doi.org/10.1139/f00-108.
Landres, P. B., Morgan, P., & Swanson, F. J. (1999). Overview of the use of natural variability concepts in managing ecological systems. Ecological Applications, 9(4), 1179–1188. https://doi.org/10.1890/1051-0761(1999)009[1179:OOTUON]2.0.CO;2.
Lapenis, A., Shvidenko, A., Shepaschenko, D., et al. (2005). Acclimation of Russian forests to recent changes in climate. Global Change Biology, 11(12), 2090–2102. https://doi.org/10.1111/j.1365-2486.2005.001069.x.
Lassau, S. A., Hochuli, D. F., Cassis, G., et al. (2005). Effects of habitat complexity on forest beetle diversity: Do functional groups respond consistently? Diversity and Distributions, 11(1), 73–82. https://doi.org/10.1111/j.1366-9516.2005.00124.x.
Lavoie, J., Girona, M. M., & Morin, H. (2019). Vulnerability of conifer regeneration to spruce budworm outbreaks in the Eastern Canadian boreal forest. Forests, 10(10), 1–14. https://doi.org/10.3390/f10100850.
Lavoie, J., Montoro Girona, M., Grosbois, G., et al. (2021). Does the type of silvicultural practice influence spruce budworm defoliation of seedlings? Ecosphere, 12(4), 17. https://doi.org/10.1002/ecs2.3506.
Lavorel, S., & Garnier, E. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Functional Ecology, 16(5), 545–556. https://doi.org/10.1046/j.1365-2435.2002.00664.x.
Leduc, A., Bernier, P. Y., Mansuy, N., et al. (2015). Using salvage logging and tolerance to risk to reduce the impact of forest fires on timber supply calculations. Canadian Journal of Forest Research, 45(4), 480–486. https://doi.org/10.1139/cjfr-2014-0434.
Leverkus, A. B., Lindenmayer, D. B., Thorn, S., et al. (2018). Salvage logging in the world’s forests: Interactions between natural disturbance and logging need recognition. Global Ecology and Biogeography, 27(10), 1140–1154. https://doi.org/10.1111/geb.12772.
Lindenmayer, D. B., & Franklin, J. F. (2002). Conserving forest biodiversity: A comprehensive multiscaled approach. Island Press.
Lindenmayer, D. B., & Fischer, J. (2007). Tackling the habitat fragmentation panchreston. Trends in Ecology & Evolution, 22(3), 127–132. https://doi.org/10.1016/j.tree.2006.11.006.
Lindner, M., Maroschek, M., Netherer, S., et al. (2010). Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecology and Management, 259(4), 698–709. https://doi.org/10.1016/j.foreco.2009.09.023.
Liquete, C., Kleeschulte, S., Dige, G., et al. (2015). Mapping green infrastructure based on ecosystem services and ecological networks: A Pan-European case study. Environmental Science & Policy, 54, 268–280. https://doi.org/10.1016/j.envsci.2015.07.009.
Littlechild, D. B., Finegan, C., & McGregor, D. (2021). “Reconciliation” in undergraduate education in Canada: The application of Indigenous knowledge in conservation. Facets, 6(1), 665–685. https://doi.org/10.1139/facets-2020-0076.
Liu, H., Park Williams, A., Allen, C. D., et al. (2013). Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Global Change Biology, 19(8), 2500–2510. https://doi.org/10.1111/gcb.12217.
Loehle, C., & Solarik, K. A. (2019). Forest growth trends in Canada. The Forestry Chronicle, 95(03), 183–195. https://doi.org/10.5558/tfc2019-027.
Löfman, S., & Kouki, J. (2001). Fifty years of landscape transformation in managed forests of southern Finland. Scandinavian Journal of Forest Research, 16(1), 44–53. https://doi.org/10.1080/028275801300004406.
Lorentzen Kolstad, A., Austrheim, G., Solberg, E. J., et al. (2018). Pervasive moose browsing in boreal forests alters successional trajectories by severely suppressing keystone species. Ecosphere, 9(10), e02458. https://doi.org/10.1002/ecs2.2458.
Lutz, W., Sanderson, W. C., & Scherbov, S. (2004). The end of world population growth in the 21st century: New challenges for human capital formation and sustainable development. Routledge.
Lyver, P. O. B., Richardson, S. J., Gormley, A. M., et al. (2018). Complementarity of indigenous and western scientific approaches for monitoring forest state. Ecological Applications, 28(7), 1909–1923. https://doi.org/10.1002/eap.1787.
MacLean, D. A. (2016). Impacts of insect outbreaks on tree mortality, productivity, and stand development. The Canadian Entomologist, 148, S138–S159. https://doi.org/10.4039/tce.2015.24.
Majcen, Z. (1994). History of selection cutting in uneven-aged forests in Quebec (Historique des coupes de jardinage dans les forêts inéquiennes au Québec). Revue Forestière Française, 45(4), 375–384. https://doi.org/10.4267/2042/26556.
Malmström, C. M., & Raffa, K. F. (2000). Biotic disturbance agents in the boreal forest: Considerations for vegetation change models. Global Change Biology, 6, 35–48. https://doi.org/10.1046/j.1365-2486.2000.06012.x.
Mansourian, S. (2018). In the eye of the beholder: Reconciling interpretations of forest landscape restoration. Land Degradation and Development, 29(9), 2888–2898. https://doi.org/10.1002/ldr.3014.
Marchand, D., Prairie, Y. T., & del Giorgio, P. A. (2009). Linking forest fires to lake metabolism and carbon dioxide emissions in the boreal region of Northern Québec. Global Change Biology, 15(12), 2861–2873. https://doi.org/10.1111/j.1365-2486.2009.01979.x.
Marchand, W., Girardin, M. P., Hartmann, H., et al. (2019). Taxonomy, together with ontogeny and growing conditions, drives needleleaf species’ sensitivity to climate in boreal North America. Global Change Biology, 25(8), 2793–2809. https://doi.org/10.1111/gcb.14665.
Marchand, W., Girardin, M. P., Hartmann, H., et al. (2021). Contrasting life-history traits of black spruce and jack pine influence their physiological response to drought and growth recovery in northeastern boreal Canada. Science of the Total Environment, 794, 148514. https://doi.org/10.1016/j.scitotenv.2021.148514.
Marini, L., Lindelöw, Å., Jönsson, A. M., et al. (2013). Population dynamics of the spruce bark beetle: A long-term study. Oikos, 122(12), 1768–1776. https://doi.org/10.1111/j.1600-0706.2013.00431.x.
Martin, M., Fenton, N., & Morin, H. (2018). Structural diversity and dynamics of boreal old-growth forests case study in Eastern Canada. Forest Ecology and Management, 422, 125–136. https://doi.org/10.1016/j.foreco.2018.04.007.
Martin, M., Morin, H., & Fenton, N. J. (2019). Secondary disturbances of low and moderate severity drive the dynamics of eastern Canadian boreal old-growth forests. Annals of Forest Science, 76(4), 108. https://doi.org/10.1007/s13595-019-0891-2.
Martin, M., Boucher, Y., Fenton, N. J., et al. (2020a). Forest management has reduced the structural diversity of residual boreal old-growth forest landscapes in Eastern Canada. Forest Ecology and Management, 458, 117765. https://doi.org/10.1016/j.foreco.2019.117765.
Martin, M., Fenton, N. J., & Morin, H. (2020b). Boreal old-growth forest structural diversity challenges aerial photographic survey accuracy. Canadian Journal of Forest Research, 50(2), 155–169. https://doi.org/10.1139/cjfr-2019-0177.
Martin, M., Girona, M. M., & Morin, H. (2020c). Driving factors of conifer regeneration dynamics in eastern Canadian boreal old-growth forests. PLoS ONE, 15(7), e0230221. https://doi.org/10.1371/journal.pone.0230221.
Martin, M., Krause, C., Fenton, N. J., et al. (2020d). Unveiling the diversity of tree growth patterns in boreal old-growth forests reveals the richness of their dynamics. Forests, 11(3), 252. https://doi.org/10.3390/f11030252.
Martin, M., Fenton, N. J., & Morin, H. (2021a). Tree-related microhabitats and deadwood dynamics form a diverse and constantly changing mosaic of habitats in boreal old-growth forests. Ecological Indicators, 128, 107813. https://doi.org/10.1016/j.ecolind.2021.107813.
Martin, M., Grondin, P., Lambert, M.-C., et al. (2021b). Compared to wildfire, management practices reduced old-growth forest diversity and functionality in primary boreal landscapes of eastern Canada. Frontiers in Forests and Global Change, 4, 1–16. https://doi.org/10.3389/ffgc.2021.639397.
McDowell, N. G., Allen, C. D., Anderson-Teixeira, K., et al. (2020). Pervasive shifts in forest dynamics in a changing world. Science, 368(6494), eaaz9463. https://doi.org/10.1126/science.aaz9463.
McGregor, D. (2018). From “decolonized” to reconciliation research in Canada: Drawing from indigenous research paradigms. Acme, 17(3), 810–831.
McLaren, B. E., & Peterson, R. O. (1994). Wolves, moose, and tree rings on Isle Royale. Science, 266(5190), 1555–1558. https://doi.org/10.1126/science.266.5190.1555.
Meigs, G. W., Morrissey, R. C., Bače, R., et al. (2017). More ways than one: Mixed-severity disturbance regimes foster structural complexity via multiple developmental pathways. Forest Ecology and Management, 406, 410–426. https://doi.org/10.1016/j.foreco.2017.07.051.
Messier, C., Puettmann, K. J., & Coates, K. D. (Eds.). (2013). Managing forests as complex adaptive systems: Building resilience to the challenge of global change (p. 368). Routledge.
Messier, C., Bauhus, J., Doyon, F., et al. (2019). The functional complex network approach to foster forest resilience to global changes. Forest Ecosystems, 6(1), 21. https://doi.org/10.1186/s40663-019-0166-2.
Michaelian, M., Hogg, E. H., Hall, R. J., et al. (2011). Massive mortality of aspen following severe drought along the southern edge of the Canadian boreal forest. Global Change Biology, 17(6), 2084–2094. https://doi.org/10.1111/j.1365-2486.2010.02357.x.
Mielikäinen, K., & Hynynen, J. (2003). Silvicultural management in maintaining biodiversity and resistance of forests in Europe–boreal zone: Case Finland. Journal of Environmental Management, 67(1), 47–54. https://doi.org/10.1016/S0301-4797(02)00187-1.
Mikusiński, G., Orlikowska, E. H., Bubnicki, J. W., et al. (2021). Strengthening the network of high conservation value forests in boreal landscapes. Frontiers in Ecology and Evolution, 8, 595730. https://doi.org/10.3389/fevo.2020.595730.
Millar, C. I., Stephenson, N. L., & Stephens, S. L. (2007). Climate change and forests of the future: Managing in the face of uncertainty. Ecological Applications, 17(8), 2145–2151. https://doi.org/10.1890/06-1715.1.
Mina, M., Messier, C., Duveneck, M., et al. (2021). Network analysis can guide resilience-based management in forest landscapes under global change. Ecological Applications, 31(1), e2221. https://doi.org/10.1002/eap.2221.
Mitchell, S. J. (2013). Wind as a natural disturbance agent in forests: A synthesis. Forestry, 86(2), 147–157. https://doi.org/10.1093/forestry/cps058.
Mitchell, S. J., & Beese, W. J. (2002). The retention system: Reconciling variable retention with the principles of silvicultural systems. The Forestry Chronicle, 78(3), 397–403. https://doi.org/10.5558/tfc78397-3.
Mölter, T., Schindler, D., Albrecht, A. T., et al. (2016). Review on the projections of future storminess over the North Atlantic European region. Atmosphere, 7(4), 60. https://doi.org/10.3390/atmos7040060.
Montoro Girona, M. (2017). À la recherche de l’aménagement durable en forêt boréale: croissance, mortalité et régénération des pessières noires soumises à différents systèmes sylvicoles. Ph.D. thesis, Université du Québec à Chicoutimi.
Montoro Girona, M., Morin, H., Lussier, J. M., et al. (2016). Radial growth response of black spruce stands ten years after experimental shelterwoods and seed-tree cuttings in boreal forest. Forests, 7(10), 1–20. https://doi.org/10.3390/f7100240.
Montoro Girona, M., Rossi, S., Lussier, J. M., et al. (2017). Understanding tree growth responses after partial cuttings: A new approach. PLoS ONE, 12(2), e0172653. https://doi.org/10.1371/journal.pone.0172653.
Montoro Girona, M., Lussier, J. M., Morin, H., et al. (2018a). Conifer regeneration after experimental shelterwood and seed-tree treatments in boreal forests: Finding silvicultural alternatives. Frontiers in Plant Science, 9(August), 1145. https://doi.org/10.3389/fpls.2018.01145.
Montoro Girona, M., Navarro, L., & Morin, H. (2018b). A secret hidden in the sediments: Lepidoptera scales. Frontiers in Ecology and Evolution, 6, 2. https://doi.org/10.3389/fevo.2018.00002.
Montoro Girona, M., Morin, H., Lussier, J.-M., et al. (2019). Post-cutting mortality following experimental silvicultural treatments in unmanaged boreal forest stands. Frontiers in Forests and Global Change, 2, 4. https://doi.org/10.3389/ffgc.2019.00004.
Mori, A. S., Furukawa, T., & Sasaki, T. (2013). Response diversity determines the resilience of ecosystems to environmental change. Biological Reviews, 88(2), 349–364. https://doi.org/10.1111/brv.12004.
Mori, A. S., Dee, L. E., Gonzalez, A., et al. (2021). Biodiversity–productivity relationships are key to nature-based climate solutions. Nature Climate Change, 11(6), 543–550. https://doi.org/10.1038/s41558-021-01062-1.
Moussaoui, L., Fenton, N. J., Leduc, A., et al. (2016). Can retention harvest maintain natural structural complexity? A comparison of post-harvest and post-fire residual patches in boreal forest. Forests, 7(10), 243. https://doi.org/10.3390/f7100243.
Moussaoui, L., Leduc, A., Girona, M. M., et al. (2020). Success factors for experimental partial harvesting in unmanaged boreal forest: 10-year stand yield results. Forests, 11(11), 1199. https://doi.org/10.3390/f11111199.
Murakami, M., & Nakano, S. (2002). Indirect effect of aquatic insect emergence on a terrestrial insect population through by birds predation. Ecology Letters, 5(3), 333–337. https://doi.org/10.1046/j.1461-0248.2002.00321.x.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., et al. (2000). Biodiversity hotspots for conservation priorities. Nature, 403(6772), 853–858. https://doi.org/10.1038/35002501.
Myneni, R. B., Keeling, C. D., Tucker, C. J., et al. (1997). Increased plant growth in the northern high latitudes from 1981 to 1991. Nature, 386(6626), 698–702. https://doi.org/10.1038/386698a0.
Nagel, L. M., Palik, B. J., Battaglia, M. A., et al. (2017). Adaptive silviculture for climate change: A national experiment in manager-scientist partnerships to apply an adaptation framework. Journal of Forestry, 115(3), 167–178. https://doi.org/10.5849/jof.16-039.
Nappi, A., Dery, S., Bujold, F., et al. (2011). Harvesting in burned forests—Issues and orientations for ecosystem-based management. Québec: Ministère des Ressources naturelles et de la Faune, Direction de l'Environnement et de la Protection des Forêts.
National Research Council (NRC). (2016). Données statistiques des ressources forestières. https://scf.rncan.gc.ca/profilstats.
Natural Resources Canada. (2020). The state of Canada’s forests. Annual report 2019 (p. 80). Ottawa: Natural Resources Canada, Canadian Forest Service.
Natural Resources Institute Finland. (2021). Statistics database: Number of threatened species. Helsinki: Natural Resources Institute Finland.
Navarro, L., Harvey, A. É., & Morin, H. (2018a). Lepidoptera wing scales: A new paleoecological indicator for reconstructing spruce budworm abundance. Canadian Journal of Forest Research, 48(3), 302–308. https://doi.org/10.1139/cjfr-2017-0009.
Navarro, L., Harvey, A. É., Ali, A., et al. (2018b). A Holocene landscape dynamic multiproxy reconstruction: How do interactions between fire and insect outbreaks shape an ecosystem over long time scales? PLoS ONE, 13(10), e0204316. https://doi.org/10.1371/journal.pone.0204316.
Navarro, L., Morin, H., Bergeron, Y., et al. (2018c). Changes in spatiotemporal patterns of 20th century spruce budworm outbreaks in eastern Canadian boreal forests. Frontiers in Plant Science 9, 1905. https://doi.org/10.3389/fpls.2018c.01905.
Niinemets, Ü., & Valladares, F. (2006). Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecological Monographs, 76(4), 521–547. https://doi.org/10.1890/0012-9615(2006)076[0521:TTSDAW]2.0.CO;2.
Nolin, A. F., Tardif, J. C., Conciatori, F., et al. (2021). Spatial coherency of the spring flood signal among major river basins of eastern boreal Canada inferred from flood rings. Journal of Hydrology, 596, 126084. https://doi.org/10.1016/j.jhydrol.2021.126084.
Nussbaum, M. C. (2011). Creating capabilities: The human development approach. Harvard University Press.
Nuttle, T., Royo, A. A., Adams, M. B., et al. (2013). Historic disturbance regimes promote tree diversity only under low browsing regimes in eastern deciduous forest. Ecological Monographs, 83(1), 3–17. https://doi.org/10.1890/11-2263.1.
Nykänen M.-L., Broadgate, M., Kellomäki, S., et al. (1997). Factors affecting snow damage of trees with particular reference to European conditions. Silva Fennica, 31(2), 5618. https://doi.org/10.14214/sf.a8519.
O'Hara, K. L. (2016). What is close-to-nature silviculture in a changing world? Forestry, 89(1), 1–6. https://doi.org/10.1093/forestry/cpv043.
O’Brien, L., Schuck, A., Fraccaroli, C., et al. (2021). Protecting old-growth forests in Europe—a review of scientific evidence to inform policy implementation. Final report. Joensuu: European Forest Institute.
O’Neill, D. W., Fanning, A. L., Lamb, W. F., et al. (2018). A good life for all within planetary boundaries. Nature Sustainability, 1(2), 88–95. https://doi.org/10.1038/s41893-018-0021-4.
Ostlund, L., Zackrisson, O., & Axelsson, A. L. (1997). The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Canadian Journal of Forest Research, 27(8), 1198–1206. https://doi.org/10.1139/x97-070.
Paillet, Y., Pernot, C., Boulanger, V., et al. (2015). Quantifying the recovery of old-growth attributes in forest reserves: A first reference for France. Forest Ecology and Management, 346, 51–64. https://doi.org/10.1016/j.foreco.2015.02.037.
Pan, Y., Birdsey, R. A., Phillips, O. L., et al. (2013). The structure, distribution, and biomass of the world’s forests. Annual Review of Ecology, Evolution, and Systematics, 44(1), 593–622. https://doi.org/10.1146/annurev-ecolsys-110512-135914.
Pappas, C., Maillet, J., Rakowski, S., et al. (2020). Aboveground tree growth is a minor and decoupled fraction of boreal forest carbon input. Agricultural and Forest Meteorology, 290, 108030. https://doi.org/10.1016/j.agrformet.2020.108030.
Pappas, C., Bélanger, N., Bergeron, Y., et al. (2022). Smartforests Canada: A network of monitoring plots for forest management under environmental change. In R. Tognetti, M. Smith, & P. Panzacchi (Eds.), Climate-smart forestry in mountain regions (pp. 521–543). Springer International Publishing.
Paquette, A., & Messier, C. (2010). The role of plantations in managing the world’s forests in the Anthropocene. Frontiers in Ecology and the Environment, 8(1), 27–34. https://doi.org/10.1890/080116.
Paradis, L., Thiffault, E., & Achim, A. (2019). Comparison of carbon balance and climate change mitigation potential of forest management strategies in the boreal forest of Quebec (Canada). Forestry, 92(3), 264–277. https://doi.org/10.1093/forestry/cpz004.
Park, A., & Wilson, E. R. (2007). Beautiful plantations: Can intensive silviculture help Canada to fulfill ecological and timber production objectives? The Forestry Chronicle, 83(6), 825–839. https://doi.org/10.5558/tfc83825-6.
Parks Canada. (2008). Principles and guidelines for ecological restoration in Canada’s protected natural areas. Gatineau: National Parks Directorate, Parks Canada.
Pasanen-Mortensen, M., Elmhagen, B., Lindén, H., et al. (2017). The changing contribution of top-down and bottom-up limitation of mesopredators during 220 years of land use and climate change. Journal of Animal Ecology, 86(3), 566–576. https://doi.org/10.1111/1365-2656.12633.
Pastor, J., Dewey, B., Moen, R., et al. (1998). Spatial patterns in the moose-forest-soil ecosystem on Isle Royale, Michigan, USA. Ecological Applications, 8(2), 411–424. https://doi.org/10.1890/1051-0761(1998)008[0411:SPITMF]2.0.CO;2.
Patoine, A., Pinel-Alloul, B., Prepas, E. E., et al. (2000). Do logging and forest fires influence zooplankton biomass in Canadian Boreal Shield lakes? Canadian Journal of Fisheries and Aquatic Sciences, 57(S2), 155–164. https://doi.org/10.1139/f00-105.
Pautasso, M., Schlegel, M., & Holdenrieder, O. (2015). Forest health in a changing world. Microbial Ecology, 69(4), 826–842. https://doi.org/10.1007/s00248-014-0545-8.
Pavoine, S., & Ricotta, C. (2019). Measuring functional dissimilarity among plots: Adapting old methods to new questions. Ecological Indicators, 97, 67–72. https://doi.org/10.1016/j.ecolind.2018.09.048.
Peltola, H., Kellomäki, S., Väisänen, H., et al. (1999). A mechanistic model for assessing the risk of wind and snow damage to single trees and stands of Scots pine, Norway spruce, and birch. Canadian Journal of Forest Research, 29(6), 647–661. https://doi.org/10.1139/x99-029.
Peng, C., Ma, Z., Lei, X., et al. (2011). A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nature Climate Change, 1(9), 467–471. https://doi.org/10.1038/nclimate1293.
Perrakis, D. D. B., Lanoville, R. A., Taylor, S. W., et al. (2014). Modeling wildfire spread in mountain pine beetle-affected forest stands, British Columbia, Canada. Fire Ecology, 10(2), 10–35. https://doi.org/10.4996/fireecology.1002010.
Perrow, M. R., & Davy, A. J. (Eds.), (2002). Handbook of ecological restoration (p. 444). Cambridge University Press.
Peura, M., Burgas, D., Eyvindson, K., et al. (2018). Continuous cover forestry is a cost-efficient tool to increase multifunctionality of boreal production forests in Fennoscandia. Biological Conservation, 217, 104–112. https://doi.org/10.1016/j.biocon.2017.10.018.
Pfeifer, M., Lefebvre, V., Peres, C. A., et al. (2017). Creation of forest edges has a global impact on forest vertebrates. Nature, 551(7679), 187–191. https://doi.org/10.1038/nature24457.
Pinel-Alloul, B., Prepas, E., Planas, D., et al. (2002). Watershed impacts of logging and wildfire: Case studies in Canada. Lake and Reservoir Management, 18(4), 307–318. https://doi.org/10.1080/07438140209353937.
Planas, D., Desrosiers, M., Groulx, S. R., et al. (2000). Pelagic and benthic algal responses in eastern canadian Boreal Shield Lakes following harvesting and wildfires. Canadian Journal of Fisheries and Aquatic Sciences, 57, 136–145. https://doi.org/10.1139/cjfas-57-s2-136 https://doi.org/10.1139/f00-130.
Portier, J., Gauthier, S., Cyr, G., et al. (2018). Does time since fire drive live aboveground biomass and stand structure in low fire activity boreal forests? Impacts on their management. Journal of Environmental Management, 225, 346–355. https://doi.org/10.1016/j.jenvman.2018.07.100.
Potapov, P., Hansen, M. C., Laestadius, L., et al. (2017). The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Science Advances, 3(1), e1600821. https://doi.org/10.1126/sciadv.1600821.
Pothier, D., Prévost, M., & Auger, I. (2003). Using the shelterwood method to mitigate water table rise after forest harvesting. Forest Ecology and Management, 179(1–3), 573–583. https://doi.org/10.1016/S0378-1127(02)00530-3.
Prévost, M., & DeBlois, J. (2014). Shelterwood cutting to release coniferous advance growth and limit aspen sucker development in a boreal mixedwood stand. Forest Ecology and Management, 323, 148–157. https://doi.org/10.1016/j.foreco.2014.03.015.
Price, D. T., Alfaro, R. I., Brown, K. J., et al. (2013). Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environmental Reviews, 21(4), 322–365. https://doi.org/10.1139/er-2013-0042.
Price, K., Holt, R., & Daust, D. (2020). BC’s old growth forest : A last stand for biodiversity. Victoria: Sierra Club BC.
Puettmann, K. J. (2011). Silvicultural challenges and options in the context of global change: “simple” fixes and opportunities for new management approaches. Journal of Forestry, 109(6), 321–331. https://doi.org/10.1093/jof/109.6.321.
Puettmann, K. J., Coates, K. D. D., & Messier, C. C. (2009). A critique of silviculture: Managing for complexity. Island Press.
Pukkala, T. (2016). Which type of forest management provides most ecosystem services? Forestry Ecosystems, 3(1), 9. https://doi.org/10.1186/s40663-016-0068-5.
Pureswaran, D. S., De Grandpré, L., Paré, D., et al. (2015). Climate-induced changes in host tree–insect phenology may drive ecological state-shift in boreal forests. Ecology, 96(6), 1480–1491. https://doi.org/10.1890/13-2366.1.
Quarrie, J. (Ed.). (1992). Earth Summit ‘92. The United Nations Conference on Environment and Development, Rio de Janeiro 1992. Rome: Food and Agriculture Organization of the United Nations.
Ram, D., Axelsson, A. L., Green, M., et al. (2017). What drives current population trends in forest birds—forest quantity, quality or climate? A large-scale analysis from northern Europe. Forest Ecology and Management, 385, 177–188. https://doi.org/10.1016/j.foreco.2016.11.013.
Rands, M. R. W., Adams, W. M., Bennun, L., et al. (2010). Biodiversity conservation: Challenges beyond 2010. Science, 329(5997), 1298–1303. https://doi.org/10.1126/science.1189138.
Raworth, K. (2017). Doughnut economics: Seven ways to think like a 21st century economist. Chelsea Green Publishing.
Raymond, P., & Bédard, S. (2017). The irregular shelterwood system as an alternative to clearcutting to achieve compositional and structural objectives in temperate mixedwood stands. Forest Ecology and Management, 398, 91–100. https://doi.org/10.1016/j.foreco.2017.04.042.
Régnière, J., St-Amant, R., & Duval, P. (2012). Predicting insect distributions under climate change from physiological responses: Spruce budworm as an example. Biological Invasions, 14(8), 1571–1586. https://doi.org/10.1007/s10530-010-9918-1.
Regos, A., Imbeau, L., Desrochers, M., et al. (2018). Hindcasting the impacts of land-use changes on bird communities with species distribution models of Bird Atlas data. Ecological Applications, 28(7), 1867–1883. https://doi.org/10.1002/eap.1784.
Rist, L., & Moen, J. (2013). Sustainability in forest management and a new role for resilience thinking. Forest Ecology and Management, 310, 416–427. https://doi.org/10.1016/j.foreco.2013.08.033.
Roberge, J. M., Öhman, K., Lämås, T., et al. (2018). Modified forest rotation lengths: Long-term effects on landscape-scale habitat availability for specialized species. Journal of Environmental Management, 210, 1–9. https://doi.org/10.1016/j.jenvman.2017.12.022.
Robertson, C., Nelson, T. A., Jelinski, D. E., et al. (2009). Spatial-temporal analysis of species range expansion: The case of the mountain pine beetle, Dendroctonus ponderosae. Journal of Biogeography, 36(8), 1446–1458.https://doi.org/10.1111/j.1365-2699.2009.02100.x.
Robinne, F. N., Burns, J., Kant, P., et al. (2018). Global fire challenges in a warming world. IUFRO.
Robinson, C. J., & Wallington, T. J. (2012). Boundary work: Engaging knowledge systems in co-management of feral animals on Indigenous lands. Ecology and Society, 17(2), 16. https://doi.org/10.5751/ES-04836-170216.
Rockström, J., Steffen, W., Noone, K., et al. (2009a). Planetary boundaries: Exploring the safe operating space for humanity. Ecology and Society, 14(2), 32. https://doi.org/10.5751/ES-03180-140232.
Rockström, J., Steffen, W., Noone, K., et al. (2009b). A safe operating space for humanity. Nature, 461(7263), 472–475. https://doi.org/10.1038/461472a.
Rogers, B. M., Soja, A. J., Goulden, M. L., et al. (2015). Influence of tree species on continental differences in boreal fires and climate feedbacks. Nature Geoscience, 8(3), 228–234. https://doi.org/10.1038/ngeo2352.
Ruel, J. C., Pin, D., & Cooper, K. (2001). Windthrow in riparian buffer strips: Effect of wind exposure, thinning and strip width. Forest Ecology and Management, 143, 105–113. https://doi.org/10.1016/S0378-1127(00)00510-7.
Ruel, J. C., Fortin, D., & Pothier, D. (2013). Partial cutting in old-growth boreal stands: An integrated experiment. The Forestry Chronicle, 89(3), 360–369. https://doi.org/10.5558/tfc2013-066.
Ryan, K. C. (2002). Dynamic interactions between forest structure and fire behavior in boreal ecosystems. Silva Fennica, 36(1), 548. https://doi.org/10.14214/sf.548.
Saad, C., Boulanger, Y., Beaudet, M., et al. (2017). Potential impact of climate change on the risk of windthrow in eastern Canada’s forests. Climatic Change, 143, 487–501. https://doi.org/10.1007/s10584-017-1995-z.
Sabatini, F. M., Keeton, W. S., Lindner, M., et al. (2020). Protection gaps and restoration opportunities for primary forests in Europe. Diversity and Distributions, 26(12), 1646–1662. https://doi.org/10.1111/ddi.13158.
Saint-Arnaud, M., Asselin, H., Dubé, C., et al. (2009). Developing criteria and indicators for aboriginal forestry: Mutual learning through collaborative research. In M. Stevenson & D. C. Natcher (Eds.), Changing the culture of forestry in Canada: Building effective institutions for Aboriginal engagement in sustainable forest management (pp. 85–105). Canadian Circumpolar Institute Press.
Sanderson, L. A., Mclaughlin, J. A., & Antunes, P. M. (2012). The last great forest: A review of the status of invasive species in the North American boreal forest. Forestry, 85(3), 329–340. https://doi.org/10.1093/forestry/cps033.
Schmiegelow, F. K. A., & Mönkkönen, M. (2002). Habitat loss and fragmentation in dynamic landscapes: Avian perspectives from the boreal forest. Ecological Applications, 12(2), 375–389. https://doi.org/10.1890/1051-0761(2002)012[0375:HLAFID]2.0.CO;2.
Schütz, J. P. (1997). Sylviculture 2: La gestion des forets irregulieres et melangees. Presses Polytechniques et Universitaires Romandes.
Sedjo, R. A., & Botkin, D. (1997). Using forest plantations to spare natural forests. Environment, 39(10), 14–30. https://doi.org/10.1080/00139159709604776.
Seidl, R., Spies, T. A., Peterson, D. L., et al. (2016). Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. Journal of Applied Ecology, 53(1), 120–129. https://doi.org/10.1111/1365-2664.12511.
Seidl, R., Thom, D., Kautz, M., et al. (2017). Forest disturbances under climate change. Nature Climate Change, 7(6), 395–402. https://doi.org/10.1038/nclimate3303.
Selva, N., Chylarecki, P., Jonsson, B.-G., et al. (2020). Misguided forest action in EU Biodiversity Strategy. Science, 368(6498), 1438–1439. https://doi.org/10.1126/science.abc9892.
Sen, A. K. (1980). Equality of what? In S. McMurrin (Ed.), Tanner Lectures on Human Values (pp. 197–220). Cambridge University Press.
Senez-Gagnon, F., Thiffault, E., Paré, D., et al. (2018). Dynamics of detrital carbon pools following harvesting of a humid eastern Canadian balsam fir boreal forest. Forest Ecology and Management, 430, 33–42. https://doi.org/10.1016/j.foreco.2018.07.044.
Shorohova, E., Kneeshaw, D., Kuuluvainen, T., et al. (2011). Variability and dynamics of old-growth forests in the circumboreal zone: Implications for conservation, restoration and management. Silva Fennica, 45(5), 72. https://doi.org/10.14214/sf.72.
Shorohova, E., Sinkevich, S., Kryshen, A., et al. (2019). Variable retention forestry in European boreal forests in Russia. Ecological Processes, 8(1), 34. https://doi.org/10.1186/s13717-019-0183-7.
Smeets, E. M. W., & Faaij, A. P. C. (2007). Bioenergy potentials from forestry in 2050. Climatic Change, 81(3–4), 353–390. https://doi.org/10.1007/s10584-006-9163-x.
Smith, C., Beazley, K. F., & Duinker, P., et al (2010). The impact of moose (Alces alces andersoni) on forest regeneration following a severe spruce budworm outbreak in the Cape Breton highlands, Nova Scotia, Canada. Alces, 46, 135–150.
Speed, J. D. M., Austrheim, G., Hester, A. J., et al. (2010). Experimental evidence for herbivore limitation of the treeline. Ecology, 91(11), 3414–3420. https://doi.org/10.1890/09-2300.1.
Spies, T. A., Lindenmayer, D. B., Gill, A. M., et al. (2012). Challenges and a checklist for biodiversity conservation in fire-prone forests: Perspectives from the Pacific Northwest of USA and southeastern Australia. Biological Conservation, 145(1), 5–14. https://doi.org/10.1016/j.biocon.2011.09.008.
Splawinski, T. B., Cyr, D., Gauthier, S., et al. (2019). Analyzing risk of regeneration failure in the managed boreal forest of northwestern Quebec. Canadian Journal of Forest Research, 49(6), 680–691. https://doi.org/10.1139/cjfr-2018-0278.
Steedman, R. J. (2000). Effects of experimental clearcut logging on water quality in three small boreal forest lake trout (Salvelinus namaycush) lakes. Canadian Journal of Fisheries and Aquatic Sciences, 57(S2), 92–96. https://doi.org/10.1139/f00-119.
Steffen, W., Richardson, K., Rockström, J., et al. (2015). Sustainability. Planetary boundaries: Guiding human development on a changing planet. Science, 347(6223), 1259855. https://doi.org/10.1126/science.1259855.
Stephens, S. L., Agee, J. K., Fulé, P. Z., et al. (2013). Managing forests and fire in changing climates. Science, 342(6154), 41–42. https://doi.org/10.1126/science.1240294.
Stocks, B. J., Mason, J. A., Todd, J. B., et al. (2003). Large forest fires in Canada, 1959–1997. Journal of Geophysical Research, 108(1), 8149. https://doi.org/10.1029/2001jd000484.
Sturtevant, B. R., Cooke, B. J., Kneeshaw, D. D., et al. (2015). Modeling insect disturbance across forested landscapes: Insights from the spruce budworm. In A. H. Perera, B. R. Sturtevant & L. J. Buse (Eds.), Simulation modeling of forest landscape disturbances (pp. 93–134). Springer International Publishing.
Suffice, P., Asselin, H., Imbeau, L., et al. (2017). More fishers and fewer martens due to cumulative effects of forest management and climate change as evidenced from local knowledge. Journal of Ethnobiology and Ethnomedicine, 13(1), 51. https://doi.org/10.1186/s13002-017-0180-9.
Svensson, J., Andersson, J., Sandström, P., et al. (2019). Landscape trajectory of natural boreal forest loss as an impediment to green infrastructure. Conservation Biology, 33(1), 152–163. https://doi.org/10.1111/cobi.13148.
Svensson, J., Bubnicki, J. W., Jonsson, B. G., et al. (2020). Conservation significance of intact forest landscapes in the Scandinavian Mountains Green Belt. Landscape Ecology, 35(9), 2113–2131. https://doi.org/10.1007/s10980-020-01088-4.
Swedish University of Agricultural Sciences (SLU). (2020). Forest statistics 2020 Official statistics of Sweden. Umeå: Swedish University of Agricultural Sciences.
Tape, K. D., Gustine, D. D., Ruess, R. W., et al. (2016). Range expansion of moose in arctic Alaska linked to warming and increased shrub habitat. PLoS ONE, 11(4), e0152636. https://doi.org/10.1371/journal.pone.0152636.
Teodoru, C. R., del Giorgio, P. A., Prairie, Y. T., et al. (2009). Patterns in pCO2 in boreal streams and rivers of northern Quebec, Canada. Global Biogeochemical Cycles, 23(2):n/a. https://doi.org/10.1029/2008GB003404.
Teshome, D. T., Zharare, G. E., & Naidoo, S. (2020). The threat of the combined effect of biotic and abiotic stress factors in forestry under a changing climate. Frontiers in Plant Science, 11, 601009. https://doi.org/10.3389/fpls.2020.601009.
Thiffault, N., & Pinno, B. D. (2021). Enhancing forest productivity, value, and health through silviculture in a changing world. Forests, 12(11), 1550. https://doi.org/10.3390/f12111550.
Thom, D., & Seidl, R. (2016). Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biological Reviews, 91(3), 760–781. https://doi.org/10.1111/brv.12193.
Thom, D., Golivets, M., Edling, L., et al. (2019). The climate sensitivity of carbon, timber, and species richness covaries with forest age in boreal-temperate North America. Global Change Biology, 25, 2446–2458. https://doi.org/10.1111/gcb.14656.
Thompson, I., Mackey, B., McNulty, S., et al. (2009). Forest resilience, biodiversity, and climate change: a synthesis of the biodiversity/resilience/stability relationship in forest ecosystems (pp. 1–67). Montreal: Secretariat of the Convention on Biological Diversity.
Thorn, S., Chao, A., Georgiev, K. B., et al. (2020). Estimating retention benchmarks for salvage logging to protect biodiversity. Nature Communications, 11(1), 4762. https://doi.org/10.1038/s41467-020-18612-4.
Thorpe, H. C., Thomas, S. C., & Caspersen, J. P. (2007). Residual-tree growth responses to partial stand harvest in the black spruce (Picea mariana) boreal forest. Canadian Journal of Forest Research, 37(9), 1563–1571. https://doi.org/10.1139/X07-148.
Tikkanen, O. P., Martikainen, P., Hyvärinen, E., et al. (2006). Red-listed boreal forest species of Finland: Associations with forest structure, tree species, and decaying wood. Annales Zoologici Fennici, 43(4), 373–383.
Tilman, D., Knops, J., Wedin, D., et al. (1997). The influence of functional diversity and composition on ecosystem processes. Science, 277, 1300–1302. https://doi.org/10.1126/science.277.5330.1300.
Trumbore, S., Brando, P., & Hartmann, H. (2015). Forest health and global change. Science, 349(6250), 814–818. https://doi.org/10.1126/science.aac6759.
Tucker, M. A., Böhning-Gaese, K., Fagan, W. F., et al. (2018). Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science, 359(6374), 466–469. https://doi.org/10.1126/science.aam9712.
Turner, N. J., & Clifton, H. (2009). “It’s so different today”: Climate change and indigenous lifeways in British Columbia, Canada. Global Environmental Change, 19(2), 180–190. https://doi.org/10.1016/j.gloenvcha.2009.01.005.
Valinger, E., & Fridman, J. (2011). Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. Forest Ecology and Management, 262(3), 398–403. https://doi.org/10.1016/j.foreco.2011.04.004.
Venier, L. A., Thompson, I. D., Fleming, R., et al. (2014). Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forests. Environmental Reviews, 22(4), 457–490. https://doi.org/10.1139/er-2013-0075.
Venier, L. A., Walton, R., Thompson, I. D., et al. (2018). A review of the intact forest landscape concept in the Canadian boreal forest: Its history, value, and measurement. Environmental Reviews, 26(4), 369–377. https://doi.org/10.1139/er-2018-0041.
Versluijs, M., Eggers, S., Hjältén, J., et al. (2017). Ecological restoration in boreal forest modifies the structure of bird assemblages. Forest Ecology and Management, 401, 75–88. https://doi.org/10.1016/j.foreco.2017.06.055.
Virkkala, R. (2016). Long-term decline of southern boreal forest birds: Consequence of habitat alteration or climate change? Biodiversity and Conservation, 25(1), 151–167. https://doi.org/10.1007/s10531-015-1043-0.
Vitousek, P. M., D’Antonio, C. M., Loope, L. L., et al. (1996). Biological invasions as global environmental change. American Scientist, 84(5), 468–478.
Vuorinen, K. E. M., Kolstad, A. L., De Vriendt, L., et al. (2020a). Cool as a moose: How can browsing counteract climate warming effects across boreal forest ecosystems? Ecology, 101(11), e03159. https://doi.org/10.1002/ecy.3159.
Vuorinen, K. E. M., Rao, S. J., Hester, A. J., et al. (2020b). Herbivory and climate as drivers of woody plant growth: Do deer decrease the impacts of warming? Ecological Applications, 30(6), e02119. https://doi.org/10.1002/eap.2119.
Walker, X. J., Baltzer, J. L., Cumming, S. G., et al. (2019). Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature, 572(7770), 520–523. https://doi.org/10.1038/s41586-019-1474-y.
Wallenius, T. (2011). Major decline in fires in coniferous forests—reconstructing the phenomenon and seeking for the cause. Silva Fennica, 45(1), 36. https://doi.org/10.14214/sf.36.
Wallenius, T., Niskanen, L., Virtanen, T., et al. (2010). Loss of habitats, naturalness and species diversity in Eurasian forest landscapes. Ecological Indicators, 10(6), 1093–1101. https://doi.org/10.1016/j.ecolind.2010.03.006.
Wang, J., & Banzhaf, E. (2018). Towards a better understanding of Green Infrastructure: A critical review. Ecological Indicators, 85, 758–772. https://doi.org/10.1016/j.ecolind.2017.09.018.
Ward, M., Saura, S., Williams, B., et al. (2020). Just ten percent of the global terrestrial protected area network is structurally connected via intact land. Nature Communications, 11(1), 4563. https://doi.org/10.1038/s41467-020-18457-x.
Warren, D. R., Keeton, W. S., Bechtold, H. A., et al. (2018). Forest-stream interactions in eastern old-growth forests. In A. M. Barton & W. S. Keeton (Eds.), Ecology and recovery of eastern old-growth forests (pp. 159–178). Island Press/Center for Resource Economics.
Watson, J. E. M., Evans, T., Venter, O., et al. (2018). The exceptional value of intact forest ecosystems. Nature Ecology and Evolution, 2(4), 599–610. https://doi.org/10.1038/s41559-018-0490-x.
Webb, A., Buddle, C. M., Drapeau, P., et al. (2008). Use of remnant boreal forest habitats by saproxylic beetle assemblages in even-aged managed landscapes. Biological Conservation, 141(3), 815–826. https://doi.org/10.1016/j.biocon.2008.01.004.
Wells, J. V., Dawson, N., Culver, N., et al. (2020). The state of conservation in North America’s boreal forest: Issues and opportunities. Frontiers in Forests and Global Change, 3, 90. https://doi.org/10.3389/ffgc.2020.00090.
Williams, M. (2003). Deforesting the earth: From prehistory to global crisis. University of Chicago Press.
Wirth, C., Messier, C., Bergeron, Y., et al. (2009). Old-growth forest definitions: A pragmatic view. In C. Wirth, G. Gleixner, & M. Heimann (Eds.), Old-growth forests: Function, fate and value (pp. 11–33). Springer.
Woo-Durand, C., Matte, J.-M., Cuddihy, G., et al. (2020). Increasing importance of climate change and other threats to at-risk species in Canada. Environmental Reviews, 28(4), 449–456. https://doi.org/10.1139/er-2020-0032.
Wu, P., Bishop, K., von Brömssen, C., et al. (2018). Does forest harvest increase the mercury concentrations in fish? Evidence from Swedish lakes. Science of the Total Environment, 622–623, 1353–1362. https://doi.org/10.1016/j.scitotenv.2017.12.075.
Wyatt, S., Hebert, M., Fortier, J. F., et al. (2019). Strategic approaches to Indigenous engagement in natural resource management: Use of collaboration and conflict to expand negotiating space by three Indigenous nations in Quebec, Canada. Canadian Journal of Forest Research, 49(4), 375–387. https://doi.org/10.1139/cjfr-2018-0253.
Zeng, H., Pukkala, T., & Peltola, H. (2007). The use of heuristic optimization in risk management of wind damage in forest planning. Forest Ecology and Management, 241(1–3), 189–199. https://doi.org/10.1016/j.foreco.2007.01.016.
Zhang, X., Lei, Y., Ma, Z., et al. (2014). Insect-induced tree mortality of boreal forests in eastern Canada under a changing climate. Ecology and Evolution, 4(12), 2384–2394. https://doi.org/10.1002/ece3.988.
Zhang, X., Bai, X., Hou, M., et al. (2019a). Warmer winter ground temperatures trigger rapid growth of dahurian larch in the permafrost forests of northeast China. Journal of Geophysical Research. Biogeosciences, 124(5), 1088–1097. https://doi.org/10.1029/2018JG004882.
Zhang, X., Manzanedo, R. D., D’Orangeville, L., et al. (2019b). Snowmelt and early to mid-growing season water availability augment tree growth during rapid warming in southern Asian boreal forests. Global Change Biology, 25(10), 3462–3471. https://doi.org/10.1111/gcb.14749.
Zhang, Y., Chen, H. Y. H., & Reich, P. B. (2012). Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. Journal of Ecology, 100(3), 742–749. https://doi.org/10.1111/j.1365-2745.2011.01944.x.
Zubizarreta-Gerendiain, A., Pukkala, T., & Peltola, H. (2017). Effects of wind damage on the optimal management of boreal forests under current and changing climatic conditions. Canadian Journal of Forest Research, 47(2), 246–256. https://doi.org/10.1139/cjfr-2016-0226.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Girona, M.M. et al. (2023). Challenges for the Sustainable Management of the Boreal Forest Under Climate Change. In: Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y. (eds) Boreal Forests in the Face of Climate Change. Advances in Global Change Research, vol 74. Springer, Cham. https://doi.org/10.1007/978-3-031-15988-6_31
Download citation
DOI: https://doi.org/10.1007/978-3-031-15988-6_31
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-15987-9
Online ISBN: 978-3-031-15988-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)