
Abstract
Monitoring forest responses to climate-smart forestry (CSF) is necessary to determine whether forest management is on track to contribute to the reduction and/or removal of greenhouse gas emissions and the development of resilient mountain forests. A set of indicators to assess “the smartness” of forests has been previously identified by combining indicators for sustainable forest management with the ecosystem services. Here, we discuss the remote sensing technologies suitable to assess those indicators grouped in forest resources, health and vitality, productivity, biological diversity, and protective functions criteria. Forest cover, growing stock, abiotic, biotic, and human-induced forest damage, and tree composition indicators can be readily assessed by using established remote sensing techniques. The emerging areas of phenotyping will help track genetic resource indicators. No single existing sensor or platform is sufficient on its own to assess all the individual CSF indicators, due to the need to balance fine-scale monitoring and satisfactory coverage at broad scales. The challenge of being successful in assessing the largest number and type of indicators (e.g., soil conditions) is likely to be best tackled through multimode and multifunctional sensors, increasingly coupled with new computational and analytical approaches, such as cloud computing, machine learning, and deep learning.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

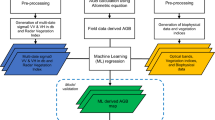
Keywords
- Forest cover
- Growing stock
- Multifunctional sensors
- Forest health
- Satellite, LiDAR, RADAR
- Climate-smart indicators Forest biodiversity
11.1 Introduction
Climate-smart forestry (CSF), as defined by Bowditch et al. (2020), consists of forest management practices that should enable both forests and society to transform, adapt to, and mitigate climate-induced changes. This definition is not far from the European Forest Institute (EFI) interpretation. Indeed, in EFI’s vision, CSF is an approach built on practices and active forest management targeted at reducing and/or removing greenhouse gas emissions to mitigate climate change, building resilient forests, and sustainably increasing forest productivity and incomes (Nabuurs et al. 2017; Kauppi et al. 2018). The economic dimension in the EFI’s point of view substitutes the social dimension of CSF on which Bowditch et al. (2020) focused. These two dimensions do not exclude each other: practices to stimulate forest productivity should not conflict with forestry practices aimed at growing forests able to contribute to the well-being of the people.
To determine whether forest management is on track to meet the goals of forest adaptation and mitigation to climate change, monitoring the forest response to practices applied during years of climate-smart forest management is necessary. Bowditch et al. (2020) selected a set of indicators to assess “the smartness” of forests, induced by forest management activities carried out in response to climate changes, by combining the pan-European indicators for sustainable forest management (SFM ) (FOREST EUROPE 2015) with the ecosystem services defined by the European Environment Agency in the Common International Classification of Ecosystem Services (CICES V5.1 2018, Haines-Young and Potschin 2018). The full list of indicators is reported in Chap. 2 of the book (Weatherall et al. 2021) together with their classification in core and peripheral groups according to their importance to assess the provision of forest ecosystem services .
Remote sensing, “as the practice of deriving information about the Earth’s land and water surfaces using images acquired from an overhead perspective, using electromagnetic radiation in one or more regions of the electromagnetic spectrum, reflected or emitted from the Earth’s surfaces” (Campbell and Wynne 2011), can contribute to quantifying CSF indicators. As a general consideration, the benefits of remote sensing to monitor the forests as a result of the application of CSF practices are related to full coverage of forested areas in a relatively short time, repeatability of measurements, and availability of data for remote or inaccessible terrestrial areas (Koch 2015). Remote sensing plays an important role in mountain forest monitoring , i.e., forests at an elevation of 2500 m a.s.l. or higher, irrespective of the slope, or on land with an elevation of 300–2500 m and a slope with sharp changes in elevation within a short distance (Kapos et al. 2000). Because of their steep slopes and often-extreme climates and weather events, mountain forests are fragile ecosystems. Under a global change scenario, remote sensing technologies allow more complete spatial and temporal monitoring of climate-smart forests and forestry (e.g., to prevent and contrast illegal logging), including those in inaccessible mountain environments. Mountains are often data-scarce regions due to their remoteness and the harsh environment: in these contexts, remote sensing may provide one of the few methods for assessing the state of dynamic changes occurring in mountain forests (Weiss and Walsh 2009). Indeed, remote sensing overcomes the challenges of collecting field data in rugged terrain and the constraints imposed by the seasonality of access to many mountain environments. Generally, remote sensing in mountain areas is very similar to remote sensing elsewhere, but the complex topography common to mountainous regions, i.e., slope with sharp changes in elevation within a short distance, introduces several challenges unique to these environments (Weiss and Walsh 2009). Remote sensing products over mountain regions come with a larger measurement error than remote sensing products over flat terrain due to topographic effects (Li et al. 2014). In the case of satellite microwave radiometric data, for example, the error is particularly correlated to the mean values of the height and slope within the radiometric pixel, as well as to the standard deviation of the aspect and local incidence angle (Li et al. 2014). In optical images, corrections in preprocessing are in general required to reduce the spectral biases due to the topographic features that led to aspect-dependent illumination and reflectance differences, shadowing, and geometric distortion (Weiss and Walsh 2009). In other remote sensing data, such as the radio detection and ranging (RADAR), topography can result in distortions, such as foreshortening and layover on slopes and in areas of shadow that are not measured.
When assessing the CSF indicators in mountain forests by remote sensing, we have to consider that the temporal scale of monitoring needs to be adjusted for different indicators to ensure early detection of change is possible. Specific focus should be put on those indicators sensitive to climate change. Forest-based climate change indicators should complement SFM indicators by capturing the effects of climate change on the forest environment and the forest sector (Lorente et al. 2018).
In this chapter, we briefly describe the key aspects of remote sensing techniques for monitoring the climate smartness of forests. Next, we consider the techniques suitable to quantify indicators of forest resources, health and vitality, productivity, biological diversity, and protection considering specific challenges in mountain regions. Finally, considerations on future developments to assess climate smartness criteria in mountain forests are provided.
11.2 Remote Sensing of CSF Criteria in Mountain Forests: An Overview
Pan-European Criteria and Indicators (PECI) have proved to be a very helpful tool in providing solid information as the basis for the sustainable management of the forests in the pan-European region between policymakers, the private sector, and civil society over the years (FOREST EUROPE 2015). The role of the CSF indicators selected by Bowditch et al. (2020) is in line with the role of PECI.
The relevance of remote sensing in quantifying the CSF indicators is linked to the possibility to extract relevant variables from remotely sensed data. In some cases, it may be possible to make relevant direct measurements, but often remote sensing proxies can be used to represent indicator values (Ghaffarian et al. 2018). For example, from tree crown delineation processes applied to light detection and ranging (LiDAR) data, tree crowns can be segmented as well as tree height quantified, and, through allometric equations, the volume can be successively estimated. For these reasons, when using remote sensing data, it is important to identify the information to be derived from the data and the kind of product and information to be delivered as an expression of CSF indicator. Besides, the coverage of remote sensing data has to be investigated. While for satellite images, the coverage should be not a problem, in the case of LiDAR data, availability could be sparse in the area of interest, and the timing and frequency of data acquisition could differ among different areas. Despite their importance, terrestrial remote sensing techniques, such as terrestrial photogrammetry and terrestrial laser scanning (TLS), are not included in this chapter. The description of the development of TLS as a plot-scale measurement tool can be found in Newnham et al. (2015), and the current state of the art in the utilization of close-range sensing in forest monitoring is summarized in Vastaranta et al. (2020). For the sake of clarity, in close-range sensing are included technologies, such as terrestrial and mobile laser scanning as well as unmanned aerial vehicles (UAV), which are mainly used for collecting detailed information from single trees, forest patches or small forested landscapes (Vastaranta et al. 2020). It is worth underlining here that, based on the current published scientific literature, the capacity to characterize changes in forest ecosystems using close-range sensing has been recognized (Vastaranta et al. 2020) and, among close-range sensing techniques, terrestrial laser scanning should be viewed as a disruptive technology that requires a rethink of vegetation surveys and their application across a wide range of disciplines (Newnham et al. 2015). These technologies are potentially game-changing but outside the scope of this chapter. Here, we focus on the systems carried on spaceborne and airborne (both manned and unmanned) platforms.
11.3 Remote Sensing of Climate Smartness According to the Forest Resources
11.3.1 Defining Forest Resources in the Context of Climate Smartness
The area covered by forests is likely to change as the climate changes. There are also likely to be shifts in forest types due to changing temperatures and precipitation regimes. Forest area is expected to contract in the mountain and boreal regions and to expand in the temperate zone (Lucier et al. 2009; Wang et al. 2019). Natural changes in climate that occurred in past geological eras have determined analogous changes in forest cover, but for the present era, it will be difficult to isolate climate change from the other factors that are affecting the range of forest area (Lucier et al. 2009). Boreal forests are expected to move north due to climate change. Temperate forests are also expected to increase their area to the north but to a greater extent than boreal forests, which will reduce the total area of boreal forests (Burton et al. 2010).
Interactions among the impacts of climate change, land-use conversion, and unsustainable land-use practices are expected. Changes in water availability will be a key factor in the survival and growth of many forest species, although the response to prolonged droughts will vary among species and also among varieties of the same species (Lucier et al. 2009). Climate change will increase the risk of frequent and more intense fires, especially in areas where it leads to lower precipitation or longer dry periods, as in boreal forests (Burton et al. 2010), and forests in the Mediterranean and subtropical regions (Fischlin et al. 2009).
CSF is needed to increase the total forest area and avoid deforestation and to facilitate the use of wood products that store carbon and substitute emission-intensive fossil and nonrenewable products and materials (Verkerk et al. 2020). Deforestation and forest degradation account for about 12% of global anthropogenic carbon emissions, which is second only to fossil fuel combustion (Calders et al. 2020). Those emissions are partially compensated by forest growth, forestation, and the rebuilding of soil carbon pools following afforestation.
As forest resources are important for climate change mitigation, timely and accurate information about their status is needed. Indeed, assessing forest resources means assessing their extent in terms of area and their distribution, the volume of standing trees, and the carbon stock in woody biomass and soil. As a consequence, the maintenance and the appropriate enhancement of forest resources and their contribution to global carbon cycles are assessed by indicators that quantify the forest area, growing stock, carbon stock, and age structure and/or diameter distribution (FOREST EUROPE 2015).
11.3.2 Appropriate Remote Sensing Methods for the Monitoring of Forest Resource Indicators
Advances in remote sensing technologies drive innovations in forest resource assessments and monitoring at varying scales. Data acquired with spaceborne and airborne platforms provide us with higher spatial resolution, more frequent coverage, and increased spectral information than was available previously (Calders et al. 2020), allowing for frequent updates of forest information layers. Optical spaceborne sensors represent a consolidated opportunity to augment traditional data sources for large-area and sample-based forest inventories, especially for inventory updates (Falkowski et al. 2009). For example, Kempeneers et al. (2012) derived two pan-European forest maps and forest-type maps for the years 2000 and 2006 from MODIS medium-resolution, optical satellite imagery using an automatic processing technique. Knorn et al. (2009) produced a map of forest/non-forest cover of large areas in the Carpathian Mountains using chain classification of neighboring Landsat satellite images. High-resolution layers of tree cover density, dominant leaf type, and forest type are derived from semiautomatic classification algorithms applied on Sentinel-1 and Sentinel-2 images every 3 years (the first products were delivered in 2006, the last in 2018). These products, representing the status and evolution of the forest surface, are used for the assessment of pan-European forest resources (Copernicus Emergency Management Service 2020). Tree cover mapping based on Sentinel-2 images demonstrated high thematic overall accuracy in Europe, i.e., up to 90% (Ottosen et al. 2020). Among those based on aerial platforms, LiDAR or airborne laser scanning (ALS), typically multiphoton LiDAR, has become an operational technology in mapping, and it is used for inventorying forests. The feasibility of using single-photon LiDAR (SPL) for land cover classification has been recently studied in North Europe (Matikainen et al. 2020). The application of algorithms to LiDAR data, most of them based on geometric characteristics of point clouds, including mathematical morphology, and adaptive and robust filtering, allows separate vegetation points from ground points in a mountainous environment. The filtering process is an essential step for the generation of the digital terrain model (DTM), and makes possible the estimation of canopy height and the production of the canopy height model (CHM). LiDAR data can also enhance the capability to discriminate forest areas in satellite images, for example, QuickBird imagery (Hilker et al. 2008) and Sentinel-2 images (Fragoso-Campón et al. 2020), by fusion in the satellite data of metrics concerning the height.
Growing stock, i.e., the stem volume of living trees, is a basic variable to assess forest resources, and it is used as a basis for estimating the amount of carbon accumulated in living trees, thereby allowing for the assessment of harvesting possibilities and risks of disturbance (FOREST EUROPE 2015). Using satellite images, Päivinen et al. (2009) produced broadleaf, coniferous, and total growing stock maps for the pan-European forest area by combining the NOAA-AVHRR imagery and statistics derived from national forest inventories of European countries. Gallaun et al. (2010) did the same using MODIS imagery. At a smaller scale, Mura et al. (2018) used Sentinel-2 imagery to estimate growing stock volume in two forest areas in Italy, and for comparison, they used Landsat 8 OLI and RapidEye images. Since the application of ALS in forestry, models trained with local inventory data have been widely applied for growing stock estimation (Næsset 1997; Maltamo et al. 2006; Dalponte et al. 2009; Corona et al. 2014). The inference of growing stock volume is carried out by regression models built to correlate values of LiDAR metrics to the values of the ground-truth volume. The metrics can be extracted from the raw (point or waveform) LiDAR data or from the CHM, at tree or plot level, following an individual tree crown (ITC) or an area-based approach (AB). Examples of metrics computable from the ALS point cloud data are aboveground elevation of highest return, height percentiles, coefficient of variation of return height, skewness and kurtosis of returns height, non-ground percentage of total returns, etc. In the CHM, metrics, such as height per pixel, coefficient of variation of the height per pixel, or the sum of the heights of all the pixels in the plot, are calculated and used in the model building. The Geoscience Laser Altimeter System (GLAS) was a laser-ranging instrument able to provide large footprint waveform LiDAR datasets for global observations of Earth, which was aboard ICESat from 2003 to 2009. GLAS data have been used to extract canopy height and map the growing stock at 1 km spatial resolution for Spanish forest areas in combination with ground forest inventory data (Sánchez-Ruiz et al. 2016).
Carbon stock, i.e., the quantity of carbon in forest biomass, dead organic matter and soil, and harvested wood products, is linked to society’s efforts to mitigate climate change by reducing the net emissions of greenhouse gases to the atmosphere (FOREST EUROPE 2015). In the past, the use of satellite imagery to assess forest biomass, and consequently to estimate the carbon stock, was mainly based on the normalized difference vegetation index (NDVI, the index that quantifies vegetation by measuring the difference between near-infrared, which vegetation strongly reflects, and red light, which vegetation absorbs) datasets. For example, at regional scale, NDVI and enhanced vegetation index (EVI) extracted from MODIS images in combination with field data were used to model carbon stock in aboveground biomass of European beech forest in central Italy (Taghavi-Bayat et al. 2012). Attempts to explore the spatiotemporal changes in carbon stock have been conducted overlaying the vegetation maps of a region and NDVI datasets (Shi and Liu 2017). It is worth underlining that NDVI is largely determined by canopy dynamics, which from an ecological point of view have very little to do with dead wood, litter, and soil carbon. In addition, Hasenauer et al. (2017) highlighted that local daily climate data should be used, and stand density effects should be addressed to obtain realistic forest productivity estimates when using satellite imagery. Hence, these kinds of products should no longer be considered data products, but they have become model products. These shortcomings contributed to the BIOMASS Earth Explorer satellite of the European Space Agency (ESA) being selected to perform a global survey of Earth’s forests and see how they change, thanks to the data that will be used in carbon cycle calculations, over the course of BIOMASS’s 5-year mission set to start on 2022. With this launch, a fully polarimetric P-band synthetic aperture radar (SAR) will be available for the first time in space. Mutual gains will be made by combining BIOMASS data with data from other missions that will measure forest biomass, structure, height, and change, including the NASA Global Ecosystem Dynamics Investigation (GEDI) LiDAR after its launch in December 2018, and the NASA-ISRO NISAR L- and S-band SAR, due for launch in 2022 (Quegan et al. 2019). Limitations of these missions have to be taken into consideration, for example, in the case of GEDI, the fact that it samples about 4% of the Earth’s land surface between 51.6° N and S latitude (Dubayah et al. 2020). Airborne S-Band SAR data have been used to estimate forest aboveground biomass in temperate mixed forests of the UK (Ningthoujam et al. 2016), while integrated spaceborne SAR data from COSMO-SkyMed (X band) and ALOS PALSAR (L band) with field inventory have been used to estimate the forest aboveground carbon (AGC) stock by Sinha et al. (2019). However, mapping biomass in mountain regions can be challenging as many regions include steep topography, making the use of RADAR data complex; for this, Mitchard et al. (2012) proposed to use a combination of terrain-corrected L-band RADAR data (ALOS PALSAR), spaceborne LiDAR data (ICESat GLAS), and ground-based data as a solution to this problem. Referring to ALS data, the most used approach for estimating carbon stocks is similar to that described for the estimation of the growing stock: it involves computing statistics from ALS point clouds for a specific pixel of forested land and relating these to carbon estimates obtained from field plots in a regression framework (Jucker et al. 2017). Currently, efforts are moving from this AB approach toward a tree-centric approach for integrating tree-level ALS data into biomass monitoring programs. Two solutions are most commonly applied: the first is to use tree height and crown dimensions, computed for a single tree after its segmentation, to predict diameters, allowing the biomass to be estimated using existing allometric equations (e.g., Dalponte and Coomes 2016); the second is to develop equations that estimate biomass directly from tree height and crown size, thus bypassing diameter altogether (Jucker et al. 2017).
Regarding the carbon in soils, Rasel et al. (2017) explored the possibility of developing a model based on variables (i.e., elevation, forest type, and aboveground biomass) extracted from LiDAR data and WorldView-2 imagery to estimate soil carbon stocks. It is evident from this kind of approach that soil carbon content cannot be measured directly by LiDAR data; hence, the problem is to understand how much modelling is acceptable to still consider its result an observational product.
With reference to the age structure indicator, information concerning the age-class structure of forests, and for uneven-aged forests, their diameter distributions, is important for understanding the history of forests and their likely future development, for assessing the harvesting potential, and for providing insights into biodiversity and recreation, which are generally more favorable in uneven-aged and old even-aged forests than in young even-aged forests (FOREST EUROPE 2015). It is known that the diameter of a tree can generally be modelled as a function of tree height or tree crown or measures related to stand structure (Filipescu et al. 2012) and derived from LiDAR data (Thomas et al. 2008; Salas et al. 2010; Bergseng et al. 2015; Spriggs et al. 2017; Arias-Rodil et al. 2018). Recently, harvester-mounted and ALS data (Maltamo et al. 2019) as well as SPOT-5 satellite imagery and field sample data (Peuhkurinen et al. 2018) have been used to estimate stand-level stem diameter distribution. Forest types (i.e., pole-stage, young, adult, mature, and old-growth forests) have been predicted using classification trees from LiDAR data (Torresan et al. 2016). Global forest canopy height products have been derived from GLAS, revealing a global latitudinal gradient in canopy height, increasing toward the equator, as well as coarse forest disturbance patterns (Simard et al. 2011), and also from MODIS and GLAS data using image segmentation (Lefsky 2010). Global canopy cover distributions were analyzed using observations from GLAS (Tang et al. 2019), and it was discovered that the estimates were sensitive to canopy cover dynamics even over dense forests with cover exceeding 80% and were able to better characterize biome-level gradients and canopy cover distributions than the existing products derived from conventional optical remote sensing.
ALS is a powerful source of data to compute the new indicators of climate smartness defined by Bowditch et al. (2020) related to vertical and horizontal forest structure. The first experiences in the application of ALS data for the assessment of vertical and horizontal forest structure are attributable to Friedlaender and Koch (2000). Successively, vertical distribution of tree crowns in terms of layers has been analyzed and characterized by Zimble et al. (2003) using ALS-derived tree heights, which allowed detecting differences in the continuous nature of vertical structure forest and specifically allowed two classes of vertical forest structure to be distinguished. Vertical distribution has been derived in multi-story stands, stratifying the ALS point cloud to canopy layers and segmenting individual tree crowns within each layer using a DSM-based tree segmentation method (Hamraz et al. 2017). Horizontal and vertical distribution of forest canopy has been derived using two point clouds from a UAV: one obtained by applying the Structure from Motion technique to digital photographs and the other one obtained from a LiDAR system (Wallace et al. 2016). Results indicate that both techniques are capable of providing information that can be used to describe canopy properties in areas of relatively low canopy closure. A comparison between waveform ALS data and discrete return ALS data, using TLS data as an independent validation, to describe the 3D structure of vegetation canopies (Anderson et al. 2015) highlighted that discrete return ALS data provide more biased and less consistent measurements of woodland canopy height than waveform ALS data. Besides, discrete return ALS data performed poorly in describing the canopy understory, compared to waveform data, but waveform ALS carried a higher data processing cost.
The slenderness coefficient, i.e., the ratio of tree total height to DBH, is a fundamental attribute for determining tree and stand stability (Vincent et al. 2012), but despite using very high-density LiDAR point cloud from aerial platforms, the stem is not sufficiently visible for accurate DBH extraction. This is only really feasible from ground-based LiDAR or photogrammetry data.
11.4 Remote Sensing of Climate Smartness According to the Forest Health and Vitality
11.4.1 Defining Health and Vitality in the Context of Climate Smartness
The state of health and vitality of a forest is determined by considering various factors, such as age, structure, composition, function, vigor, the presence of unusual levels of insects or disease, and resilience to disturbance. Climate change may have profound impacts on the health and vitality of the forests. In some cases, vitality may increase due to a combination of carbon dioxide fertilization and a more favorable climate. However, in many cases, benefits of carbon dioxide increase on tree growth may be outweighed by increasing drought- and heat-induced tree mortality (Allen et al. 2010), and increasing temperatures can favor the growth of insect populations that are particularly detrimental to the health of forests composed of few tree species (Lucier et al. 2009), as in the case of alpine forests affected by the spruce bark beetle (Ips typographus L.). Longer harvesting periods, increased storm damage, and longer spore-production season seem to be the causes of the increase in infestations of root and bud rot by the fungus Heterobasidion parviporum Niemelä & Korhonen in coniferous forests in North Europe (Burton et al. 2010).
The criterion of forest health and vitality addresses one of the main concerns of the European countries at the start of the pan-European process. This criterion includes indicators of soil conditions; forest damage by abiotic, biotic, and human-induced agents; defoliation; deposition; and concentration of air pollutants on forests (FOREST EUROPE 2015).
11.4.2 Appropriate Remote Sensing Methods for the Monitoring of Health and Vitality Indicators
Indicators of soil conditions, defined in terms of carbon, water, and nutrient concentrations, are generally not directly measurable by remote sensing techniques in forest environments, as exposed soil is rarely visible. However, SAR and microwave radiometer systems using long wavelengths (e.g., L-Band) have been used to measure surface (0–5 cm depth) soil moisture under forest canopies, but modelling of the scattering and absorption effects of the canopy is required, and the effect of the litter layer and surface roughness must be considered. Root-zone soil moisture can be estimated by assimilation of such data into land-surface or hydrological models, but such approaches may be less successful in mountainous and densely forested areas (Pablos et al. 2018). A review of modelling the passive microwave signature from land surfaces can be found in Wigneron et al. (2017), and some assessment of accuracy over forested areas is provided in Vittucci et al. (2016). The coarse resolution of passive microwave radiometer satellite missions, such as Soil Moisture Ocean Salinity (SMOS) and Soil Moisture Active Passive (SMAP), largely limits their application to regional scales and above. The potential for the use of active RADAR for soil moisture retrieval has also been explored, with good agreement with ground measurements, using L- and P-band polarimetric airborne SAR in jack pine forest stands (Moghaddam et al. 2000), and for soil moisture variations retrieval from ERS SAR satellite data in a recently burned black spruce forest in Alaska (Kasischke et al. 2007).
Remote sensing can significantly contribute to measuring and monitoring forest damage from abiotic (e.g., drought, winter injury, wind storms, avalanche, landslide, fires, air pollution) and biotic (insect pests, diseases) stresses, which influence key biophysical and biochemical parameters of the tree canopy and structure. Large-scale, stand-replacing, disturbance events leading to significant loss of tree cover can be readily monitored with satellite or aerial imagery using change detection approaches. Over large areas, extensive satellite image time series, such as the Landsat archive, have been used to map tree cover loss, using methods such as the vegetation change tracker algorithm (Masek et al. 2013), and to capture both slowly evolving and abrupt changes in forest cover using LandTrendr – temporal segmentation algorithm (Kennedy et al. 2010). IKONOS satellite imagery with Tasseled Cap transformation and edge enhancements have been tested for mapping of snow avalanche paths (Walsh et al. 2004) and RapidEye imagery for detecting windthrow damage based on pre- and post-storm object-based change detection (Einzmann et al. 2017). Remote sensing has also been widely applied in monitoring both active fires (using thermal sensors) and fire severity and in detecting fire scar (Szpakowski and Jensen 2019). Although both stand-replacing disturbances and finer-scale abiotic or biotic forest damage and decline can be measured, attribution of the specific cause of observed stress or disturbance can frequently be more challenging. But it can potentially be achieved through consideration of the spectral properties (e.g., dead and burnt materials resulting from fires; McDowell et al. 2015), temporal signatures and specific symptom progression (e.g., for pest or disease; Stone and Mohammed, 2017), and spatial patterns of disturbance. Regarding forest damage by human-induced agents, Kennedy et al. (2007) used distinctive temporal signatures in the progression of spectral properties before and after an event to identify the timing of disturbance events, such as clear-cuts and thinnings from a dense stack of Landsat TM images, and to attribute the type using a series of rules. Hilker et al. (2011) utilized spatial characteristics of disturbed patches (patch size, core area, and contiguity), along with the date of disturbance, to attribute disturbance types in Alberta, Canada, using a regression tree classification method, while Hermosilla et al. (2015) utilized spectral, temporal, and geometric metrics from Landsat time series to attribute disturbance as fire, harvesting, road, and non-stand-replacing changes with a 91.6% accuracy level. Baumann et al. (2014) developed a method to separate windfall disturbance from clearcut forest harvesting activity using Landsat data, after Tasseled Cap transformation, obtaining classification accuracy over 75% for windfall areas and better results for larger disturbance patches. The classification was based on spectral differences between the disturbance types, such as lower brightness (due to shadows from remaining biomass) and higher wetness for the windfall areas. Often ancillary data, such as meteorological observations, information on known disturbance agents or events (e.g., storm paths, species ranges for hosts and pests, indices of fire risk), and additional field-based monitoring observations, are needed to reliably attribute or confirm causes, often through integration with modelling (McDowell et al. 2015).
To monitor changes in forest vitality and stress in individual trees, a range of biophysical and biochemical parameters can be estimated with remote sensing. These include defoliation, alterations in pigment concentrations, reduced photosynthesis and light-use efficiency, changes in water relations and hydraulic transport, including leaf water content and evapotranspiration rates, and changes in leaf cell structure due to senescence or wilting. Detailed reviews of the use of remote sensing systems to detect such changes resulting from pests and disease (Chen and Meentemeyer 2016) and die-off from abiotic stress , such as drought (Huang et al. 2019), provide an in-depth insight into the capability of different sensor systems. Optical sensors are capable of detecting noticeable changes in foliar color due to changes in pigment concentrations or photosynthetic activity. A wide range of spectral indices have been developed to estimate pigment concentrations (e.g., the carotenoid reflectance index; Gitelson et al. 2002), light-use efficiency (LUE), or photosynthetic activity from multispectral or hyperspectral data, and a detailed summary of such methods in monitoring forest decline and disturbance can be found in Pontius et al. (2020). Widely used LUE indices, such as the photosynthetic reflectance index (PRI; Gamon et al. 1997) and chlorophyll/carotenoid index (CCI; Gamon et al. 2016), have allowed detection of water stress (Hernández-Clemente et al. 2011; Dotzler et al. 2015) and forest pests, e.g., Peña and Altmann (2009) use the PRI to detect aphid-induced stress in the Chilean Andes. Such methods require hyperspectral data, which can be costly and hard to acquire and can be sensitive to illumination conditions and canopy structure. Some early-stage pest and disease symptoms cannot be easily observed in the visible range but can be identified by sensors with the capacity to detect the near- and shortwave- infrared spectrum, where reflectance is strongly influenced by the structure of leaf mesophyll tissue and water content. A common example is the use of the NDVI, which can be sensitive to defoliation and to changing chlorophyll levels but also prone to saturation at high values of leaf area index (LAI). Optical remote sensing has been extensively tested for the survey of damage by mountain pine beetle in the Canadian and American Rocky Mountains, ranging from visual interpretation of aerial photography (e.g., Klein 1982) to the use of Landsat time series and spectral indices related to needle water content for automated mapping of red attack stages (e.g., Skakun et al. 2003). More recently, efforts have been made to detect early “green” attack stages of mountain pine beetle infestation, using hyperspectral data (e.g., Fassnacht et al. 2014) and with Mullen et al. (2018) using high-resolution WorldView-2 satellite imagery to detect differences in spectral properties of individual tree crowns, especially in the near-infrared region. Other remote sensing methods attempt to measure changes in photosynthesis or plant functioning more directly. Thermal measurements are sensitive to changes in leaf temperature caused by reduced evapotranspiration as a result of stomatal closure following water stress . Such measurements have been used in the detection of red-band needle blight in pine from a UAV platform (Smigaj et al. 2019) but are highly sensitive to changes in meteorological conditions and time of acquisition. The ECOSTRESS instrument on the International Space Station aims to provide thermal-derived estimates of water stress over much larger extents. A wide range of optical spectral indices (e.g., shortwave infrared-based normalized difference water index, Gao 1996, or moisture stress index, Hunt and Rock 1989) have been proposed to detect vegetation drought stress , but methods have also been proposed based on passive microwave sensors (e.g., estimation of relative water content based on vegetation optical depth, Rao et al. 2019) and solar-induced chlorophyll fluorescence (SIF) of leaves. SIF results from the emittance of light during photosynthetic activity and, therefore, allows for tracking of photosynthetic activity, phenology, and estimation of GPP, but it is influenced by plant stress when excess light is present and LUE is low. SIF has been used to detect stress due to environmental conditions in forest environments (e.g., the Lägeren forest site in Switzerland, where an abrupt decrease in SIF was shown to relate to a heat wave and an aphid outbreak causing early leaf senescence, Paul-Limoges et al. 2018) and has significant future potential with the expected 2022 launch of the ESA FLEX Earth Explorer mission; however, such measurements are again influenced by the diurnal timing of acquisition and the species being observed.
Defoliation is a key indicator of forest decline and has been extensively monitored using optical, LiDAR, and RADAR data at a range of scales from individual trees to landscape. For example, Olsson et al. (2016) use MODIS NDVI products to monitor insect outbreak-linked defoliation in a subalpine birch forest in Sweden, in near real time. Through the use of an NDVI time series, 74% of defoliation was detected where pixels comprised at least 50% birch forest cover, but with some significant misclassification of undisturbed forest, depending on threshold selection. In contrast, Meng et al. (2018) map defoliation at the individual tree level, based on airborne hyperspectral imaging and LiDAR metrics. They show red-edge and near-infrared wavelength regions to be sensitive to defoliation due to gypsy moth at the crown scale and demonstrate the superior capability of LiDAR structural and intensity metrics to predict leaf area. Meiforth et al. (2020) combine WorldView-2 and LiDAR to detect dieback of New Zealand kauri trees, utilizing spectral indices, such as NDVI and red-green ratio. Including LiDAR structural metrics improved the correlation with graded stress levels but incurs significant additional expense, and steep terrain can cause spatial misalignment with optical data. However, the use of LiDAR to identify and segment individual tree crowns can often be important for the analysis of high-resolution optical data. Recent and next-generation satellite sensors, including Sentinel-2, WorldView-2 and WorldView-3, and GEDI, increase the capacity for monitoring of defoliation across larger spatial scales (Meng et al. 2018).
Pollution can also impact forest health, particularly air pollution from ground-level ozone and excessive nitrogen or sulfur deposition. Remote sensing approaches, including near-infrared aerial photography, airborne hyperspectral systems, and satellite observations, have long been used to monitor forest decline attributed to pollutants including acid rain. For example, Rock et al. (1988) showed a shift in the red-edge location of spruce and fir tree spectra to shorter wavelengths (a so-called blue shift) from airborne hyperspectral data in the presence of forest decline in the American and German mountain sites, believed to be due to pollutants including trace metals. Rees and Williams (1997) monitored the effects of air pollution on terrestrial ecosystems using Landsat-MSS images from 1978 to 1992, to study the impact of sulfur dioxide emissions on boreal forest, while Diem (2002) showed foliar injury related to ozone exposure could be detected from Landsat-MSS vegetation indices. However, the pollution-related decline is often compounded by other stress factors, including insect or disease outbreaks. Remote sensing also has an additional role to play in this area through monitoring of atmospheric conditions and pollutants, including nitrogen dioxide and sulfur dioxide (Martin 2008). For example, ground-level ozone formation can be studied from measurements of precursor compounds (e.g., formaldehyde and nitrogen dioxide) from satellite observations by the ozone monitoring instrument onboard NASA’s Aura satellite (Jin et al. 2017).
11.5 Remote Sensing of Climate Smartness According to the Forest Productivity
11.5.1 Defining Forest Productivity in the Context of Climate Smartness
Forest productivity, i.e., the potential of a particular forest stand to produce aboveground wood volume, is affected by climate change to differing extents, according to geographic area, species, stand composition, tree age, soil water retention capacity, and the interactions between these factors (Gao et al. 2019; Ammer 2019; Paquette et al. 2018). Some changes in productivity may be short term, and transitory and previous levels of productivity may be restored once carbon sinks become saturated (Hedin 2015) and water availability scarce. However, in areas where the water is not a limiting factor, there may be an initial increase in growth if there is less waterlogging. Similar reactions have been noted for carbon dioxide (Ollinger et al. 2008) and nitrogen fertilization (LeBauer and Treseder 2008) and increased temperatures (Reich and Oleksyn 2008). While in most temperate areas, forest productivity has been found to increase with higher temperature, which is probably due to carbon dioxide fertilization, in tropical areas, the productivity declines when carbon dioxide saturation is reached (Hubau et al. 2020), probably due to water deficits over extended periods.
The indicators of productive functions of forests aim to assess the quantity and the values of the produced goods and marketed services, as well as to make sure that this productivity is sustainable, using multipurpose management (FOREST EUROPE 2015). The balance between net annual increment and annual felling is the indicator used to understand the forest’s potential for wood production and the conditions it provides for biodiversity, health, recreation, and other forest functions. The assessment of the quantity and market values of roundwood is important for wood supply, particularly for marginalized rural areas, where the wood energy chain has been suggested as a means to reactivate forest management and improve the value of forest stands (Vacchiano et al. 2018).
11.5.2 Appropriate Remote Sensing Methods for the Monitoring of the Forest Productivity Indicators
Although remote sensing cannot itself directly measure indicators such as the balance between the net annual increment and annual felling of wood, or the quantity and market values of roundwood available for wood chain supply, or the proportion of forest under a management plan, it is worth underlining that the computation of increment assumes the knowledge of the volume of standing trees, living or dead, which is the growing stock whose estimation using remote sensing was already considered in Sect. 11.3.2.
Net primary production (NPP) can be considered an indicator of productivity, quantifiable using remote sensing data (e.g., Coops 2015): according to this approach, top-of-atmosphere measurements of solar radiance from satellite observation are used to estimate the incoming photosynthetically active radiation (PAR), and, successively, LUE modelling techniques are used to estimate the gross primary production (GPP) and the NPP. It appears clear that PAR can be measured and thus GPP can be estimated, but going from GPP to NPP requires that radiation is modelled or makes assumptions regarding LUE, which depend on field observations. Besides, the application of these models at a higher level of detail requires the availability of high spatial and temporal resolution maps of a series of model drivers (e.g., meteorology, land use) frequently unavailable for mountain areas (Yang et al. 2020). Furthermore, there are few good and complete GPP observations globally, and NPP estimates are typically multi-year averages and cannot be better than field observations (Šímová and Storch 2017). Even the amount of foliage of stands, measured as the LAI, is a key indicator of forest productivity, principally due to its importance for photosynthesis, transpiration, evapotranspiration, and, in turn, GPP. Remote sensing estimation of LAI has been undertaken using several approaches reported in Coops (2015), but LAI measurements based on light absorption have been shown to saturate for values bigger than 4 (Waring et al. 2010); therefore, this aspect limits the value of this product. Remote sensing estimates of LAI and fraction of absorbed PAR can be integrated into process-based forest growth models, such as 3-PG SPATIAL, to model productivity at stand to regional scales and growth variables such as site index (SI), the most common means for quantifying forest stand-level potential productivity, and to estimate impacts of climate change on future productivity (Coops et al. 2011). LAI estimates are routinely available from MODIS satellite data and have become widely used since their release in 2000 (De Kauwe et al. 2011). SI has been compared with GPP estimates obtained from 3-PG SPATIAL using climate variables and MODIS (Weiskittel et al. 2011). Results indicated that a nonparametric model with two climate-related predictor variables explained over 68% and 76% of the variation in SI and GPP, respectively. The relationship between GPP and SI was limited (determination coefficient of 36–56%), while the relationship between GPP and climate (determination coefficient of 76–91%) was stronger than the one between SI and climate (determination coefficient of 68–78%).
In terms of harvesting activity, Smith and Askne (2001) used ERS SAR interferograms over 3 years to detect clearcut areas of a minimum size of 0.4 ha in northern Sweden. Saksa et al. (2003) compared the applicability of Landsat satellite imagery and high-altitude panchromatic aerial orthophotos using digital change detection methods in detecting clearcut areas in a boreal forest. MODIS images were used by Bucha and Stibig (2008) for the detection and monitoring of forest clearcuts in the boreal forest in north-west Russia, while Lambert et al. (2015) proposed the usage of MODIS NDVI time series in south-western Massif Central Mountains in France to detect clearfelling. Panagiotidis et al. (2019) detected fallen logs from high-resolution UAV images at plot level (i.e., 2500 m2 plot size) in flat topographic conditions and open canopy cover.
Climatic changes affect the treeline location, causing shifts, and identification and quantification of treeline dynamics are critical. An approach for the identification of shifts in the treeline altitudes for a period of four decades based on NDVI, land surface temperature (LST) data, air temperature data, and forest stand maps has been developed for the mountain forest of Cehennemdere in Turkey, showing a geographical expansion of the treeline in both the highest altitudes and the lowest altitudes (Arekhi et al. 2018). The integration of in situ dendrometric data with analyses from dendrochronological samples, high-resolution 3D UAV photos, and new satellite images has been found a solution to study the dynamics and underlying causal mechanisms of any treeline movement and growth changes in mountain forests located in Central and East Asia (Cazzolla Gatti et al. 2019).
11.6 Remote Sensing of Climate Smartness According to the Forest Biological Diversity
11.6.1 Defining Forest Biological Diversity in the Context of Climate Smartness
Different species have individual climatic ranges, in which they remain competitive with other plant species, can adapt to environmental change, and respond to increased insect attacks, disease, and adverse environmental conditions and anthropogenic influences. Some species will adapt better than others to changing conditions, which will lead to changes in the composition of forests, instead of geographic shifts in forest types (Breshears et al. 2008). In general, tree species are likely to move to higher latitudes or altitudes due to global warming (Rosenzweig et al. 2007; Breshears et al. 2008).
Climate change drives phenological changes, i.e., changes in seasonal timing of life-history events, in many tree species observed in phenological gardens (Seppälä et al. 2009). A phenological garden contains a selection of species and a selection of clonal strains among each species. In order to minimize non-climatic influences on plant development, a network of phenological gardens with the same species and the same clones should be set up as to constitute a network of plants. The observers at the gardens have detailed, illustrated instructions that describe exactly the phenophases to be reported. Phenological gardens, therefore, assure a maximum of exact observations with controlled internal plant conditions. An adjacent meteorological station is essential for later correlations among the results from a phenological garden and for possible physiological modelling (Schnelle and Volkert 1974). The highest number of changes and the most significant changes were noted in phenological gardens located at higher latitudes (Seppälä et al. 2009). There is evidence that part of this phenological change, such as dates of spring bud break and flowering, which can affect productivity and carbon sequestration potential, and autumnal foliar coloration, may be due to increasing atmospheric carbon dioxide concentrations as well as warming (Seppälä et al. 2009).
Biodiversity remains an important topic for forest policy and management in Europe (FOREST EUROPE 2015). Forest biological diversity criterion encompasses indicators referring to tree species composition, introduced tree species, and threatened forest species. It also includes indicators that quantify the area of forests by the class of naturalness and fragmentation, protected forests, forests for the conservation and utilization of tree genetic resources, seed production, and new forests. The average volume of deadwood, both standing and lying, is also an indicator of biological diversity.
11.6.2 Appropriate Remote Sensing Methods for the Monitoring of Biological Diversity Indicators
The assessment of biological diversity can benefit from the availability of different sources of remotely sensed data and from the opportunities for automated forest interpretation at the tree level, i.e., the possibility to delineate and classify tree species. Whatever approach is used for tree detection, crown area delineation, and species classification, remote sensing offers strong potentiality for the assessment of tree species composition and introduced tree species. Various experiences of tree crown detection, delineation, and species classification had applied digital aerial images (e.g., Brandtberg 2002; Haara and Haarala 2002; Erikson 2004; Korpela 2004), but LiDAR represents the most effective source of data for detecting and delineating trees (e.g., Heinzel and Koch 2012; Maltamo et al. 2009). The integration of ALS data with aerial high-resolution multispectral or hyperspectral images has been tested for tree crown delineation and tree species classification (e.g., Zarea and Mohammadzadeh 2016; Dalponte et al. 2019; Weinstein et al. 2019) as well as the integration of ALS data with high-resolution aerial near-infrared images (Persson and Holmgren 2004). Satellite imagery, e.g., high spatial resolution 8-Band WorldView-2 and 5-band RapidEye, has proven to be valid in species classification (Immitzer et al. 2012).
There are many studies that have modelled patterns in spectral diversity and species richness, paralleling those of biochemical diversity, demonstrating a linkage between the taxonomic and remotely sensed properties of forest canopies (e.g., Asner et al. 2009; Warren et al. 2014; Roth et al. 2015). The approach, ushered in Asner and Martin (2008) and called “spectranomics,” resulting from the combination of science and technology, emerged from aspects of established remote sensing research with new ideas to causally link the biochemistry, spectroscopy, taxonomy, and community ecology of canopies. A review of the history of remote sensing approaches for biodiversity estimation, specifically focused on relating spectral diversity to biodiversity at different scales, with the summarization of the pros and cons of different methods in remote sensing of plant biodiversity can be found in Wang and Gamon (2018). The step from individual trees to the area of forest classified by the number of tree species occurring or to the area of forest land dominated by introduced tree species is quite short.
Despite progress in tree crown delineation and species classification, the quantification of indicators, such as the number of threatened forest species, classified according to the International Union for Conservation of Nature (IUCN) Red List categories in relation to the total number of forest species, could be challenging. Indeed, although the application of techniques – such as support vector machines and Gaussian maximum likelihood with leave-one-out-covariance algorithm classifiers – in hyperspectral imaging and LiDAR data allows classifying classes of species (Dalponte et al. 2008), the classification of all single species contained in the IUCN Red List categories is not yet operationally applied as in the case of species categories.
The amount and variability of deadwood in a forest stand are important indicators of forest biodiversity because deadwood provides critical habitat for thousands of species in forests (Bater et al. 2009; Sandström et al. 2019). The current situation and new perspectives of remote sensing application for deadwood identification and characterization have been summarised by Marchi et al. (2018a, b). Infrared aerial photos are suitable for mapping and quantifying single standing dead trees, i.e., snags (Bütler et al. 2004; Bütler and Schlaepfer 2004). The identification of standing deadwood using LiDAR data has started to be addressed around 10 years ago, due to the increase in high-quality and high-density data availability, as well as the availability of segmentation methods required to work directly with the point cloud. The moderate capacity of LiDAR remote sensing to estimate the distribution of standing dead tree classes in forest stands has been demonstrated (correlation coefficient of 0.61; Bater et al. 2009), as well as the good capacity of high-spatial resolution aerial photos taken from a UAV to survey fallen trees in deciduous broadleaved forests (trees with a diameter bigger than 30 cm or longer than 10 m were identified with a rate equal to 80%, but many trees that were narrower or shorter were missed; Inoue et al. 2014). In general, failure in identifying snags, logs, and stumps may be due to the similarity of fallen trees to trunks and branches of standing trees or masking by standing trees. The noise near the soil also affects the detection performance of the trees lying in the ground.
Indicators of regeneration, naturalness, landscape patterns, protected forests, and genetic resources are related to a landscape scale of analysis instead of a single-tree scale, and they can be assessed using approaches of classification applied to remote sensing data, both to produce maps or to obtain statistics. The emitted and/or reflected radiance from the canopy of the regenerated forest, which is related to the biophysical properties of the vegetation, such as leaf and wood biomass, can be used to identify the forest regenerative stage after, for example, a clearance (Lucas et al. 2000) or wildfire (Morresi et al. 2019) using satellite imagery. Multispectral aerial images in the green, red, and near-infrared spectral bandwidths have been used to monitor regenerating forests using an automated tree detection-delineation algorithm by Pouliot et al. (2005). Sentinel-2 imagery with ALS data was integrated for inventorying regeneration stands by the estimation of sapling density (Landry et al. 2020). The application of optical UAV-based imagery for the inventory of natural regeneration in post-disturbed forests has been tested by Röder et al. (2018). The assessment of the status of forest regeneration using aerial photogrammetry and UAV was carried out by Goodbody et al. (2018). Moreover, young and advanced regenerating forests were studied using UAV-based hyperspectral imagery and photogrammetric point clouds (Imangholiloo et al. 2019) and multispectral ALS data (Imangholiloo et al. 2020), both under leaf-off and leaf-on conditions in boreal forests. Similarly, UAV data have been applied for regeneration assessments in seedling stands (Puliti et al. 2019; Castilla et al. 2020; Green and Burkhart, 2020).
Landsat satellite images constitute a major data source for spatial patterns of fragmentation that allow disturbances in protected areas to be identified (Nagendra et al. 2013). In particular, multitemporal Landsat data resulted in valid satellite imagery to analyze patterns of forest fragmentation by the extraction of landscape metrics, which convey significant information on biophysical changes associated with forest fragmentation at broad scales (Fuller 2001). ALS and SAR data, especially when used synergistically with optical data, allow the detection of changes in the three-dimensional structure of forests, facilitating their classification in different classes of naturalness (Hirschmugl et al. 2007).
Protected forests to conserve biodiversity are a land-use and not a land cover category because the protection of an area is given by the humans according to the use they want to do and by the law. For this reason, remote sensing can be used for mapping large intact forest areas or adjusting the boundaries of areas already protected or detecting major changes in land cover within protected areas. This aspect is relevant to underpin the development of a general strategy for nature conservation at the global and regional scales (Willis 2015). For example, Potapov et al. (2008) introduced a new approach for mapping large intact forest landscapes using existing fine-scale maps and global coverage of high-spatial resolution satellite imagery. Gillespie and Willis (2015) reviewed advances and limitations in spaceborne remote sensing that can be applied to all terrestrial protected areas around the world for baseline vegetation mapping, land cover classifications, invasive species, and degradation identification, monitoring forest ecosystems and land cover dynamics using time series data.
The same consideration can be made for the areas managed for conservation and utilization of forest tree genetic resources or managed for seed production, which are land-use rather than land cover categories that require additional data, to which remote sensing can potentially contribute. Indeed, remote sensing cannot do genotyping, but it can contribute to phenotyping, therefore providing indicators of genetic resilience, e.g., variations in stress responses. This is an emerging area for remote sensing. Most of the studies have been carried out in tree nurseries, experimental plots, or crop trees rather than in the field. For example, Ludovisi et al. (2017) used thermal imaging acquired by UAV for high-throughput field phenotyping of black poplar response to drought, enabling highly precise and efficient, nondestructive screening of genotype performance in large plots. Phenotyping of individual trees in situ in planted forests helping to track genetic performance was conducted by Dungey et al. (2018). But there have been “phenotyping platforms,” which involve the phenotyping of whole forests eventually down to the individual tree level, proposed for whole-forest phenotyping to better quantify the key drivers of forest productivity to inform and optimize future breeding and deployment programs (Dungey 2016). Improved UAV-based measurements of tree health (fluorescence, canopy temperature, structure traits from LiDAR, etc.) could have much potential for linking genetic and phenotypic traits in terms of stress responses to climate change.
11.7 Remote Sensing of Climate Smartness According to the Forest Protective Function
11.7.1 Defining Forest Protective Function in the Context of Climate Smartness
The contribution of forests to water and soil protection has long been recognized. Forests can play a vital role in preventing soil erosion and protecting water supplies (FOREST EUROPE 2015). In addition, a wide variety of man-made infrastructure relies on the protection provided by forests. Such protective functions are mostly found in mountainous areas or areas subject to extreme climatic conditions. The importance of forests with protective functions has increased in the last decades due to settlement pressure, climate change, and the high vulnerability of society in mountain regions (Bauerhansl et al. 2010). However, foresters and hydrologists still debate the nature of the influence that forests have on water regulation. Climate change may make the role of forests in water regulation and soil protection more important, but the capacity of forests to fulfill this role may also be affected. Reductions in rainy season flows and increases in dry season flows are of little value when total annual rainfall is low and significant quantities of water are lost through evapotranspiration and are consumed by forests. Unmanaged forests supply high levels of climate regulation and erosion regulation, while best practice management slightly improves water regulation (Seidl et al. 2019).
In this context, information on the spatial distribution of protective forests designated to prevent soil erosion, preserve water resources, protect infrastructure, and manage natural resources against natural hazards (i.e., avalanche, snow gliding, rockfall, landslide, flood, water erosion, wind erosion, karstification) becomes essential.
11.7.2 Appropriate Remote Sensing Methods for the Monitoring of Protective Function Indicators
To assess the forest protective function against natural hazards, model simulations are often used. Variables used in the models range from topographic features (e.g., altitude, slope and slope gradient, orientation to wind and orientation to the sun, aspect, plan curvature) to forest characteristics (e.g., crown cover, stem per hectare, gap width) to individual tree features (e.g., tree species, tree diameter, and height) (Bigot et al. 2009). In this context, remote sensing techniques are an indispensable supplement of data, which can be integrated with inventory field data for the identification of protective effects on large areas (Bauerhansl et al. 2010), having in mind that the protective function strongly depends on small-scale local conditions, such as terrain and stand characteristics (Teich and Bebi 2009).
Since the 2000s, Bebi et al. (2001) and Teich and Bebi (2009) used aerial photographs for assessing structures in mountain forests (e.g., canopy density, crown closure, trees in clusters) as a basis for investigating the forests’ protective function. Dorren et al. (2006) assessed protection forest structure with ALS in mountain terrain, and, according to their results, ALS provides excellent input data for 3D natural hazard simulation models, even in steep terrain. Monnet et al. (2010) assessed the potential of ALS for estimating stand parameters required as input data for rockfall simulation models or more generally for quantifying the rockfall protection function of forests. In the case of falling rocks, the topography determines its occurrence, the direction, as well as the velocity of a falling rock; therefore, DEM is the most relevant dataset for simulation studies (Dorren et al. 2004). Maroschek et al. (2015) combined remote sensing data (i.e., a LiDAR-based normalized crown model and a volume map) and inventories to generate realistic fine-grained forest landscapes with single-tree level information as input to the spatially explicit hybrid model used to assess the protective effect of the vegetation. Brožová et al. (2020) determined forest parameters for avalanche simulation using remote sensing data, specifically photogrammetry-based vegetation height model and LiDAR-based vegetation height model .
Hydrological models are commonly used to study both the flow and the quality of water. Forest cover, tree height, LAI, and sky view factor are the four main structural parameters used by hydrological models of forests (Varhola and Coops 2013). Varhola and Coops (2013) estimated these four watershed-level distributed forest structure metrics using LiDAR and Landsat data. They found a high correlation between forest spectral indices and structural metrics, and they successfully modelled the four metrics by Landsa-LiDAR calibrations. Satellite-derived land cover data, as well as data on snow cover, LAI, evapotranspiration, and surface soil moisture, are widely integrated into catchment and regional- and global-scale hydrological modelling (Xu et al. 2014). Concerning the role of forest in preventing soil erosion, de Asis and Omasa (2007) estimated vegetation parameters for modelling soil erosion using Landsat ETM data in a linear spectral mixture analysis at a catchment scale.
11.8 Remote Sensing of Climate Smartness According to Socioeconomic Function
The impacts of climate change on the forest sector will increase in strength, but they will vary in space and over time, depending on the geographical region and the crop type (Viccaro et al. 2019). According to the expected future scenarios for Europe, in the Mediterranean area, there will be a reduction of the forest capital, caused by a reduced water supply, while in the North of Europe, there will be an expansion of forests, both in terms of surface and species and an extended growing season due to more favorable soil temperature and moisture conditions and the higher supply of carbon dioxide for photosynthesis (Viccaro et al. 2019). The expected global increase in wood production together with provisional and local increased availability of wood in the market due to windstorms could lead to lower prices, which would benefit consumers. However, lower prices and regionally differentiated impacts on productivity will have varying effects on incomes and employment derived from timber.
The socioeconomic function criterion assesses the economic value of forests (FOREST EUROPE 2015); indicators related to wood consumption, trade in wood (i.e., import and export), and the share of wood energy in total primary energy supply are contemplated together with accessibility for recreation (i.e., right of access, provision of facilities, and intensity of use). The first three indicators are not directly measurable with remote sensing techniques as wood consumption cannot be estimated from areas felled due to the international market in timber. But, where trees are grown specifically for a local market, e.g., as a biomass energy crop such as short rotation willow, remote sensing can potentially provide input layers for field trial scale analyses and modelling (Castaño-Díaz et al. 2017). The last indicator refers to the right to access the forests for recreational purposes, which is related to the forest holdings: remote sensing technique cannot support the assessment of this indicator.
11.9 Conclusion
The potentiality of the remote sensing as a source of data for the assessment of climate-smart criteria and indicators in mountain forests has been demonstrated for many of the indicators examined in the above sections. Indicators of forest resources, such as the spatial extent of forest cover that can be used to assess the spatial dynamics of that cover, can be monitored using data from optical sensors, which may provide information on the amount of foliage and its biochemical properties, with optical wavelengths both absorbed and scattered by leaves, needles, and branches that make up a forest canopy. Vegetation indices extracted from optical satellite imagery have been used to model carbon stock in aboveground biomass for many years. More recently, the capacity emerged to assess the 3D forest structure, using data acquired from LiDAR and SAR sensors, due to the ability of the waves to penetrate through the canopy and backscatter from branches and stems and so to provide a geometrical description of the forest. LiDAR allows the estimation of the volume of living trees and of carbon stock by models that assume relationships between growing and carbon stock and LiDAR-derived canopy metrics. The measurements provided by optical and radar sensors are different due to divergences in wavelength, imaging geometry, and technical implementation, but, when combined, they can be complementary when the sensitivity to individual 3D forest structure components is considered. This combination of sensors and observation platforms holds a lot of promise for the future.
Indicators of health and vitality can be monitored using data from sensors able to acquire plant reflectance in the visible to shortwave-infrared regions, which provide information in the biochemical properties of foliage and plant fluorescence, which are indicative of the capacity and functioning of the photosynthetic apparatus. Through multisensor data and consideration of spatial and temporal patterns of changes, abiotic and biotic causes of disturbance or stress can be identified.
The specific indicators of productive function, such as increment and felling and roundwood, are not directly measurable. NPP, as an indicator globally considered an expression of forest productivity, is modelled using optical satellite imagery data. These models have been successfully used on a global and continental scale to estimate daily and annual NPP from satellite data with low ground resolutions, but the availability of maps at high level of detail related to model drivers, such as precipitation, temperature, and land use, is required to produce accurate estimation through these models. Indicators of the productive function of forest can be quantified using data acquired with microwave sensors that can provide information on woody biomass.
Indicators of biological diversity criterion, such as tree composition, deadwood, etc., can be assessed with optical, LiDAR, RADAR, and SAR. Although hyperspectral holds a lot of promise, few applications, using its full potential, have so far emerged. Spectral diversity is also increasingly considered as a measure of biodiversity at a range of scales, but tree species mapping over large area is not still an operational procedure mostly due to costs and limited extent of airborne hyperspectral.
LiDAR provides the most suitable data to be inputted in models that quantify the protective function of forests against, for example, rockfall. Data extracted from optical satellite imagery can also be used to assess the effect of natural disturbances on the functionality of direct protection forests.
In the context of an increasing amount of remote sensing data generated by multiple airborne and spaceborne platforms, integrated multisensor frameworks will be required to move beyond the state of the art in forest monitoring (Lehmann et al. 2015). This kind of approach allows balancing the needs for fine-scale monitoring of distribution patterns and satisfactory coverage at broad scales (He et al. 2019). For example, with specific reference to the assessment of the forest health and vitality, Lausch et al. (2018) underline that it is becoming evident that no existing monitoring approach, technique, model , or platform is sufficient on its own to monitor, model , forecast, or assess forest health and its resilience. Hence, a multisource forest health monitoring network for the twenty-first century, which couples different monitoring approaches, is a viable strategy that has to be supported. In addition to multisensor and multiplatform approaches, recently, a significant amount of work has been carried out in the field of multimode sensors and multifunctional sensors (Majumder et al. 2019). Such approaches are increasingly coupled with new computational and analytical approaches, such as cloud computing, machine learning, and deep learning, able to handle big datasets and diverse data sources. From this perspective, monitoring the effects of climate-smart forest management interventions requires smart deployment and the use of remote sensing technologies, which can open up new opportunities in the assessment of the capabilities of the forests to transform, adapt to, and mitigate climate-induced changes throughout the selected indicators.
References
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH (Ted), Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Ammer C (2019) Diversity and forest productivity in a changing climate. New Phytol 221:50–66. https://doi.org/10.1111/nph.15263
Anderson K, Hancock S, Disney M, Gaston KJ (2015) Is waveform worth it? A comparison of LiDAR approaches for vegetation and landscape characterization. Remote Sens Ecol Conserv 2(1):5–15. https://doi.org/10.1002/rse2.8
Arekhi M, Yesil A, Ozkan UY, Balik Sanli F (2018) Detecting treeline dynamics in response to climate warming using forest stand maps and Landsat data in a temperate forest. For Ecosyst 5:23–36. https://doi.org/10.1186/s40663-018-0141-3
Arias-Rodil M, Diéguez-Aranda U, Álvarez-González JG et al (2018) Modeling diameter distributions in radiata pine plantations in Spain with existing countrywide LiDAR data. Ann For Sci 75:36–47. https://doi.org/10.1007/s13595-018-0712-z
Asner GP, Martin RA (2008) Airborne spectranomics: mapping canopy chemical and taxonomic diversity in tropical forests. Front Ecol Environ 7(5):269–276. https://doi.org/10.1890/070152
Asner GP, Martin RE, Ford AJ, Metcalfe DJ, Liddell MJ (2009) Leaf chemical and spectral diversity in Australian tropical forests. Ecol Appl 19(1):236–253. https://doi.org/10.1890/08-0023.1
Bater CW, Coops NC, Gergel SE et al (2009) Estimation of standing dead tree class distributions in northwest coastal forests using lidar remote sensing. Can J For Res 39:1080–1091. https://doi.org/10.1139/X09-030
Bauerhansl C, Berger F, Dorren L et al (2010) Development of harmonized indicators and estimation procedures for forests with protective functions against natural hazards in the alpine space (PROALP). European Commission, Joint Research Centre, Institute for Environment and Sustainability. Office for Official Publications of the European Communities. © European Communities, 2010. https://doi.org/10.2788/51473
Baumann M, Ozdogan M, Wolter PT et al (2014) Landsat remote sensing of forest windfall disturbance. Remote Sens Environ 143:171–179. https://doi.org/10.1016/j.rse.2013.12.020
Bayat AT, van Gils H, Weir M (2012) Carbon stock of European Beech forest; a case at M. Pizzalto, Italy. APCBEE Procedia 1:159–168. https://doi.org/10.1016/j.apcbee.2012.03.026
Bebi P, Kienast F, Schönenberger W (2001) Assessing structures in mountain forests as a basis for investigating the forests’ dynamics and protective function. For Ecol Manag 145:3–14. https://doi.org/10.1016/S0378-1127(00)00570-3
Bergseng E, Ørka HO, Næsset E, Gobakken T (2015) Assessing forest inventory information obtained from different inventory approaches and remote sensing data sources. Ann For Sci 72:33–45. https://doi.org/10.1007/s13595-014-0389-x
Bigot C, Dorren LKA, Berger F (2009) Quantifying the protective function of a forest against rockfall for past, present and future scenarios using two modelling approaches. Nat Hazard 49:99–111. https://doi.org/10.1007/s11069-008-9280-0
Bowditch E, Santopuoli G, Binder F et al (2020) What is Climate-Smart Forestry? A definition from a multinational collaborative process focused on mountain regions of Europe. Ecosyst Serv 43:101113. https://doi.org/10.1016/j.ecoser.2020.101113
Brandtberg T (2002) Individual tree-based species classification in high spatial resolution aerial images of forests using fuzzy sets. Fuzzy Sets Syst 132:371–387. https://doi.org/10.1016/S0165-0114(02)00049-0
Breshears DD, Huxman TE, Adams HD, Zou CB, Davison JE (2008) Vegetation synchronously leans upslope as climate warms. Proceedings of the National Academy of Sciences 105(33):11591–11592. https://doi.org/10.1073/pnas.0806579105
Brožová N, Fischer JT, Bühler Y et al (2020) Determining forest parameters for avalanche simulation using remote sensing data. Cold Reg Sci Technol 172:102976. https://doi.org/10.1016/j.coldregions.2019.102976
Bucha T, Stibig HJ (2008) Analysis of MODIS imagery for detection of clear cuts in the boreal forest in north-west Russia. Remote Sens Environ 112:2416–2429. https://doi.org/10.1016/j.rse.2007.11.008
Burton PJ, Bergeron Y, Bogdansky BEC, Juday GP et al (2010) Sustainability of boreal forests and forestry in a changing environment. In: Mery G, Katila P, Galloway G, Alfaro R, Kanninen M, Lobovikov M, Varjo J (eds) Forests and society responding to global drivers of change, vol 25. IUFRO World, Series, pp 249–282
Bütler R, Schlaepfer R (2004) Spruce snag quantification by coupling colour infrared aerial photos and a GIS. For Ecol Manag 195:325–339. https://doi.org/10.1016/j.foreco.2004.02.042
Bütler R, Angelstam P, Ekelund P, Schlaepfer R (2004) Dead wood threshold values for the three-toed woodpecker presence in boreal and sub-Alpine forest. Biol Conserv 119:305–318. https://doi.org/10.1016/j.biocon.2003.11.014
Calders K, Jonckheere I, Nightingale J, Vastaranta M (2020) Remote sensing technology applications in forestry and REDD+. Forests 11:10–13. https://doi.org/10.3390/f11020188
Campbell JB, Wynne RH (2011) Introduction to remote sensing, 5th edn. Guilford Press, New York. ISBN:9781609181765
Castaño-Díaz M, Álvarez-Álvarez P, Tobin B, Nieuwenhuis M, Afif-Khouri E, Cámara-Obregón A (2017) Evaluation of the use of low-density LiDAR data to estimate structural attributes and biomass yield in a short-rotation willow coppice: an example in a field trial. Ann For Sci 74:69. https://doi.org/10.1007/s13595-017-0665-7
Castilla G, Filiatrault M, McDermid GJ, Gartrell M (2020) Estimating individual conifer seedling height using drone-based image point clouds. Forests 11:924. https://doi.org/10.3390/f11090924
Cazzolla Gatti R, Callaghan T, Velichevskaya A et al (2019) Accelerating upward treeline shift in the Altai Mountains under last-century climate change. Sci Rep 9:1–13. https://doi.org/10.1038/s41598-019-44188-1
Chen G, Meentemeyer RK (2016) Remote sensing of forest damage by diseases and insects. 145–162. https://doi.org/10.1201/9781315371931-9
Coops NC (2015) Characterizing forest growth and productivity using remotely sensed data. Curr For Rep 1:195–205. https://doi.org/10.1007/s40725-015-0020-x
Coops NC, Gaulton R, Waring RH (2011) Mapping site indices for five Pacific Northwest conifers using a physiologically based model. Appl Veg Sci 14:268–276. https://doi.org/10.1111/j.1654-109X.2010.01109.x
Copernicus Emergency Management Service. https://emergency.copernicus.eu/. Accessed 06/29/2020
Corona P, Fattorini L, Franceschi S et al (2014) Estimation of standing wood volume in forest compartments by exploiting airborne laser scanning information: model-based, design-based, and hybrid perspectives. Can J For Res 44:1303–1311. https://doi.org/10.1139/cjfr-2014-0203
Dalponte M, Coomes DA (2016) Tree-centric mapping of forest carbon density from airborne laser scanning and hyperspectral data. Methods Ecol Evol 7:1236–1245. https://doi.org/10.1111/2041-210X.12575
Dalponte M, Bruzzone L, Gianelle D (2008) Fusion of hyperspectral and LIDAR remote sensing data for classification of complex forest areas. IEEE Trans Geosci Remote Sens 46:1416–1427. https://doi.org/10.1109/TGRS.2008.916480
Dalponte M, Bruzzone L, Dalponte M et al (2009) Analysis on the use of multiple returns LiDAR data for the estimation of tree stems volume. IEEE J Sel Top Appl Earth Obs Remote Sens 2:310–318. https://doi.org/10.1109/JSTARS.2009.2037523
Dalponte M, Frizzera L, Gianelle D (2019) Individual tree crown delineation and tree species classification with hyperspectral and LiDAR data. PeerJ 6:e6227. https://doi.org/10.7717/peerj.6227
de Asis AM, Omasa K (2007) Estimation of vegetation parameter for modeling soil erosion using linear Spectral Mixture Analysis of Landsat ETM data. ISPRS J Photogramm Remote Sens 62:309–324. https://doi.org/10.1016/j.isprsjprs.2007.05.013
De Kauwe MG, Disney MI, Quaife T, Lewis P, Williams M (2011) An assessment of the MODIS collection 5 leaf area index product for a region of mixed coniferous forest. Remote Sens Environ 115(2):767–780. https://doi.org/10.1016/j.rse.2010.11.004
Diem JE (2002) Remote assessment of forest health in southern Arizona, USA: evidence for ozone-induced foliar injury. Environ Manag 29:373–384. https://doi.org/10.1007/s00267-001-0011-5
Dorren LKA, Maier B, Putters US, Seijmonsbergen AC (2004) Combining field and modelling techniques to assess rockfall dynamics on a protection forest hillslope in the European Alps. Geomorphology 57:151–167. https://doi.org/10.1016/S0169-555X(03)00100-4
Dorren L, Maier B, Berger F (2006) Assessing protection forest structure with airborne laser scanning in steep mountainous terrain. Paper presented at the Workshop on 3D Remote Sensing in Forestry, 14th–15th Feb 2006, Vienna 238–242
Dotzler S, Hill J, Buddenbaum H, Stoffels J (2015) The potential of EnMAP and sentinel-2 data for detecting drought stress phenomena in deciduous forest communities. Remote Sens 7:14227–14258. https://doi.org/10.3390/rs71014227
Dubayah R, Blair JB, Goetz S et al (2020) The global ecosystem dynamics investigation: high-resolution laser ranging of the Earth’s forests and topography. Sci Remote Sens 1:100002. https://doi.org/10.1016/j.srs.2020.100002
Dungey H (2016) Forest genetics for productivity – the next generation. New Zeal J For Sci 46:40490. https://doi.org/10.1186/s40490-016-0081-z
Dungey HS, Dash JP, Pont D et al (2018) Phenotyping whole forests will help to track genetic performance. Trends Plant Sci 23:854–864. https://doi.org/10.1016/j.tplants.2018.08.005
Einzmann K, Immitzer M, Böck S et al (2017) Windthrow detection in european forests with very high-resolution optical data. Forests 8:1–26. https://doi.org/10.3390/f8010021
Erikson M (2004) Species classification of individually segmented tree crowns in high-resolution aerial images using radiometric and morphologic image measures. Remote Sens Environ 91:469–477. https://doi.org/10.1016/j.rse.2004.04.006
Falkowski MJ, Wulder MA, White JC, Gillis MD (2009) Supporting large-area, sample-based forest inventories with very high spatial resolution satellite imagery. Prog Phys Geogr 33:403–423. https://doi.org/10.1177/0309133309342643
FAO (2015) Knowledge reference for national forest assessments. http://www.fao.org/3/a-i4822e.pdf
Fassnacht FE, Latifi H, Ghosh A et al (2014) Assessing the potential of hyperspectral imagery to map bark beetle-induced tree mortality. Remote Sens Environ 140:533–548. https://doi.org/10.1016/j.rse.2013.09.014
Filipescu CN, Groot A, Maclsaac DA et al (2012) Prediction of diameter using height and crown attributes: a case study. West J Appl For 27:30–35. https://doi.org/10.1093/wjaf/27.1.30
Fischlin A, Ayres M, Karnosky D, Kellomäki S, Louman B, Ong C, Plattner C, Santoso H, Thompson I, Booth T, Marcar N, Scholes B, Swanston C, Zamolodchikov D (2009) Future environmental impacts and vulnerabilities. In: Seppala R, Buck A, Katila P (eds) Adaptation of forests and people to climate change. IUFRO World Series 22. Geist and Lambin, 2002
FOREST EUROPE (2015) State of Europe’s Forests:2015
Fragoso-Campón L, Quirós E, Mora J, Gutiérrez Gallego JA, Durán-Barroso P (2020) Overstory-understory land cover mapping at the watershed scale: accuracy enhancement by multitemporal remote sensing analysis and LiDAR. Environ Sci Pollut Res 27:75–88. https://doi.org/10.1007/s11356-019-04520-8
Friedlaender H, Koch B (2000) First experience in the application of laser scanner data for the assessment of vertical and horizontal forest structures. Int Arch Photogramm Remote Sensing XXXIII(Part B7, ISPRS Congr XXXIII):693–700
Fuller DO (2001) Forest fragmentation in Loudoun County, Virginia, USA evaluated with multitemporal Landsat imagery. Landscape Ecology 16:627–642. https://link.springer.com/article/10.1023/A:1013140101134
Gallaun H, Zanchi G, Nabuurs GJ et al (2010) EU-wide maps of growing stock and above-ground biomass in forests based on remote sensing and field measurements. For Ecol Manag 260:252–261. https://doi.org/10.1016/j.foreco.2009.10.011
Gamon JA, Serrano L, Surfus JS (1997) The photochemical reflectance index: an optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 112:492–501. https://doi.org/10.1007/s004420050337
Gamon JA, Huemmrich KF, Wong CYS et al (2016) A remotely sensed pigment index reveals photosynthetic phenology in evergreen conifers. Proc Natl Acad Sci USA 113:13087–13092. https://doi.org/10.1073/pnas.1606162113
Gao BC (1996) NDWI – a normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens Environ 58:257–266. https://doi.org/10.1016/S0034-4257(96)00067-3
Gao WQ, Lei XD, Fu LY (2019) Impacts of climate change on the potential forest productivity based on a climate-driven biophysical model in northeastern China. J For Res. https://doi.org/10.1007/s11676-019-00999-6
Ghaffarian S, Kerle N, Filatova T (2018) Remote sensing-based proxies for urban disaster risk management and resilience: a review. Remote Sens 10:1760–1789. https://doi.org/10.3390/rs10111760
Gillespie TW, Willis KS, Ostermann-Kelm S (2015) Spaceborne remote sensing of the world’s protected areas. Prog Phys Geogr 39:388–404. https://doi.org/10.1177/0309133314561648
Gitelson AA, Zur Y, Chivkunova OB, Merzlyak MN (2002) Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem Photobiol 75:272. https://doi.org/10.1562/0031-8655(2002)0750272ACCIPL2.0.CO2
Goodbody TRH, Coops NC, Hermosilla T et al (2018) Assessing the status of forest regeneration using digital aerial photogrammetry and unmanned aerial systems. Int J Remote Sens 39:5246–5264. https://doi.org/10.1080/01431161.2017.1402387
Green PC, Burkhart HE (2020) Plantation Loblolly pine seedling counts with unmanned aerial vehicle imagery: a case study. J For 118(5):487–500. https://doi.org/10.1093/jofore/fvaa020
Haara A, Haarala M (2002) Tree species classification using semi-automatic delineation of trees on aerial images. Scand J For Res 17:556–565. https://doi.org/10.1080/02827580260417215
Haines-Young R, Potschin MB (2018) Common international classification of ecosystem services (CICES) V5.1 and guidance on the application of the revised structure. Fabis Consulting Ltd. The Paddocks, Chestnut Lane, Barton in Fabis, Nottingham, NG11 0AE, UK. Available from www.cices.eu
Hamraz H, Contreras MA, Zhang J (2017) Vertical stratification of forest canopy for segmentation of understory trees within small-footprint airborne LiDAR point clouds. ISPRS J Photogramm Remote Sens 130:385–392. https://doi.org/10.1016/j.isprsjprs.2017.07.001
Hasenauer H, Neumann M, Moreno A, Running S (2017) Assessing the resources and mitigation potential of European forests. Energy Procedia 125:372–378. https://doi.org/10.1016/j.egypro.2017.08.052
He Y, Chen G, Potter C, Meentemeyer RK (2019) Integrating multi-sensor remote sensing and species distribution modeling to map the spread of emerging forest disease and tree mortality. Remote Sens Environ 231:111238. https://doi.org/10.1016/j.rse.2019.111238
Hedin LO (2015) Signs of saturation in the tropical carbon sink Hot on the trail of temperature processing. Nature 519:295–296. https://doi.org/10.1038/519295a
Heinzel J, Koch B (2012) Investigating multiple data sources for tree species classification in temperate forest and use for single tree delineation. Int J Appl Earth Obs Geoinf 18:101–110. https://doi.org/10.1016/j.jag.2012.01.025
Hermosilla T, Wulder MA, White JC et al (2015) Regional detection, characterization, and attribution of annual forest change from 1984 to 2012 using Landsat-derived time-series metrics. Remote Sens Environ 170:121–132. https://doi.org/10.1016/j.rse.2015.09.004
Hernández-Clemente R, Navarro-Cerrillo RM, Suárez L et al (2011) Assessing structural effects on PRI for stress detection in conifer forests. Remote Sens Environ 115:2360–2375. https://doi.org/10.1016/j.rse.2011.04.036
Hilker T, Wulder MA, Coops NC (2008) Update of forest inventory data with lidar and high spatial resolution satellite imagery. Can J Remote Sens 34:5–12. https://doi.org/10.5589/m08-004
Hilker T, Coops NC, Gaulton R, Wulder MA, Cranston J, Stenhouse GB (2011) Biweekly disturbance capture and attribution: case study in western Alberta grizzly bear habitat. J Appl Remote Sens 5(1). https://doi.org/10.1117/1.3664342
Hirschmugl M, Ofner M, Raggam J, Schardt M (2007) Single tree detection in very high resolution remote sensing data. Remote Sens Environ 110:533–544. https://doi.org/10.1016/j.rse.2007.02.029
Huang C-Y, Anderegg WRL, Asner GP (2019) Remote sensing of forest die-off in the Anthropocene: from plant ecophysiology to canopy structure. Remote Sens Environ 231:111233. https://doi.org/10.1016/j.rse.2019.111233
Hubau W, Lewis SL, Phillips OL et al (2020) Asynchronous carbon sink saturation in African and Amazonian tropical forests. Nature 579:80–87. https://doi.org/10.1038/s41586-020-2035-0
Hunt ER, Rock BN (1989) Detection of changes in leaf water content using Near- and Middle-Infrared reflectances. Remote Sens Environ 30:43–54. https://doi.org/10.1016/0034-4257(89)90046-1
Imangholiloo M, Saarinen N, Markelin L et al (2019) Characterizing seedling stands using leaf-off and leaf-on photogrammetric point clouds and hyperspectral imagery acquired from unmanned aerial vehicle. Forests 10:1–17. https://doi.org/10.3390/f10050415
Imangholiloo M, Saarinen N, Holopainen M, Yu X, Hyyppä J, Vastaranta M (2020) Using leaf-off and leaf-on multispectral airborne laser scanning data to characterize seedling stands. Remote Sens 12:3328. https://doi.org/10.3390/rs12203328
Immitzer M, Atzberger C, Koukal T (2012) Tree species classification with Random forest using very high spatial resolution 8-band worldView-2 satellite data. Remote Sens 4:2661–2693. https://doi.org/10.3390/rs4092661
Inoue T, Nagai S, Yamashita S et al (2014) Unmanned aerial survey of fallen trees in a deciduous broadleaved forest in eastern Japan. PLoS One 9:1–7. https://doi.org/10.1371/journal.pone.0109881
Jin X, Fiore AM, Murray LT et al (2017) Evaluating a space-based indicator of surface ozone-NOx-VOC sensitivity over midlatitude source regions and application to decadal trends. J Geophys Res Atmos 122:10439–10461. https://doi.org/10.1002/2017JD026720
Jucker T, Caspersen J, Chave J et al (2017) Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob Chang Biol 23:177–190. https://doi.org/10.1111/gcb.13388
Kapos V, Rhind J, Edwards M et al (2000) Developing a map of the world’s mountain forests. In: Price MF, Butt N (eds) Forests in sustainable mountain development: a state-of-knowledge report for 2000. CAB International, Wallingford, pp 4–9
Kasischke ES, Bourgeau-Chavez LL, Johnstone JF (2007) Assessing spatial and temporal variations in surface soil moisture in fire-disturbed black spruce forests in Interior Alaska using spaceborne synthetic aperture radar imagery – implications for post-fire tree recruitment. Remote Sens Environ 108:42–58. https://doi.org/10.1016/j.rse.2006.10.020
Kauppi P, Hanewinkel M, Lundmark T, Nabuurs GJ, Peltola H, Trasobares A, Hetemäki L (2018) Climate smart forestry in Europe. European Forest Institute
Kempeneers P, Sedano F, Pekkarinen A, Seebach L, Strobl P, San Miguel-Ayanz J (2012). Pan-European forest maps derived from optical satellite imagery. IEEE Earthzine 5 (2nd quarter theme). https://earthzine.org/pan-european-forest-maps-derived-from-optical-satellite-imagery/
Kennedy RE, Cohen WB, Schroeder TA (2007) Trajectory-based change detection for automated characterization of forest disturbance dynamics. Remote Sens Environ 110:370–386. https://doi.org/10.1016/j.rse.2007.03.010
Kennedy RE, Yang Z, Cohen WB (2010) Detecting trends in forest disturbance and recovery using yearly Landsat time series: 1. LandTrendr – temporal segmentation algorithms. Remote Sens Environ 114:2897–2910. https://doi.org/10.1016/j.rse.2010.07.008
Klein WH (1982) Estimating bark beetle-killed lodgepole pine with high altitude panoramic photography. Photogramm Eng Remote Sens 48:733–737
Knorn J, Rabe A, Radeloff VC et al (2009) Land cover mapping of large areas using chain classification of neighboring Landsat satellite images. Remote Sens Environ 113:957–964. https://doi.org/10.1016/j.rse.2009.01.010
Koch B. (2015). Remote sensing supporting national forest inventories NFA. In Food and Agriculture Organization of the United Nations, Knowledge reference for national forest assessments. Rome: Food and Agriculture Organization of the United Nations (pp. 77–92)
Korpela I (2004) Individual tree measurements by means of digital aerial photogrammetry. In Silva Fennica Monographs 3:93 p
Lambert J, Denux JP, Verbesselt J et al (2015) Detecting clear-cuts and decreases in forest vitality using MODIS NDVI time series. Remote Sens 7:3588–3612. https://doi.org/10.3390/rs70403588
Landry S, St-Laurent M-H, Pelletier G, Villard M-A (2020) The best of both worlds? Integrating Sentinel-2 images and airborne LiDAR to characterize forest regeneration. Remote Sens 2020(12):2440. https://doi.org/10.3390/rs12152440
Lausch A, Borg E, Bumberger J et al (2018) Understanding forest health with remote sensing, Part III: Requirements for a scalable multi-source forest health monitoring network based on data science approaches. Remote Sens 10:1120–1171. https://doi.org/10.3390/rs10071120
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89(2):371–379. https://doi.org/10.1890/06-2057.1
Lefsky MA (2010) A global forest canopy height map from the moderate resolution imaging spectroradiometer and the geoscience laser altimeter system. Geophys Res Lett 37:1–5. https://doi.org/10.1029/2010GL043622
Lehmann EA, Caccetta P, Lowell K et al (2015) SAR and optical remote sensing: Assessment of complementarity and interoperability in the context of a large-scale operational forest monitoring system. Remote Sens Environ 156:335–348. https://doi.org/10.1016/j.rse.2014.09.034
Li X, Zhang L, Weihermuller L et al (2014) Measurement and simulation of topographic effects on passive microwave remote sensing over mountain areas: a case study from the tibetan plateau. IEEE Trans Geosci Remote Sens 52:1489–1501. https://doi.org/10.1109/TGRS.2013.2251887
Lorente M, Gauthier S, Bernier P, Ste-Marie C (2018) Tracking forest changes: Canadian Forest Service indicators of climate change. Clim Change:1–15. https://doi.org/10.1007/s10584-018-2154-x
Lucas RM, Honzák M, Curran PJ et al (2000) Mapping the regional extent of tropical forest regeneration stages in the Brazilian Legal Amazon using NOAA AVHRR data. Int J Remote Sens 21:2855–2881. https://doi.org/10.1080/01431160050121285
Lucier A, Ayres M, Karnosky D, Thompson I, Loehle C, Percy K, Sohngen B (2009) Forest responses and vulnerabilities to recent climate change. In: Seppala R, Buck A, Katila P (eds) Adaptation of forests and people to climate change. IUFRO World Series 22. Geist and Lambin, 2002
Ludovisi R, Tauro F, Salvati R et al (2017) UAV-based thermal imaging for high-throughput field phenotyping of black poplar response to drought. Front Plant Sci 8:1–18. https://doi.org/10.3389/fpls.2017.01681
Majumder BD, Roy JK, Padhee S (2019) Recent advances in multifunctional sensing technology on a perspective of multi-sensor system: a review. IEEE Sens J 19:1204–1214. https://doi.org/10.1109/JSEN.2018.2882239
Maltamo M, Eerikäinen K, Packalén P, Hyyppä J (2006) Estimation of stem volume using laser scanning-based canopy height metrics. Forestry 79:217–229. https://doi.org/10.1093/forestry/cpl007
Maltamo M, Peuhkurinen J, Malinen J et al (2009) Predicting tree attributes and quality characteristics of scots pine using airborne laser scanning data. Silva Fenn 43:507–521. https://doi.org/10.14214/sf.203
Maltamo M, Hauglin M, Næsset E, Gobakken T (2019) Estimating stand level stem diameter distribution utilizing harvester data and airborne laser scanning. Silva Fenn 53:1–19. https://doi.org/10.14214/sf.10075
Marchi N, Pirotti F, Lingua E (2018a) Airborne and terrestrial laser scanning data for the assessment of standing and lying deadwood: current situation and new perspectives. Remote Sens 10:1356–1376. https://doi.org/10.3390/rs10091356
Marchi N, Weisberg P, Greenberg J et al (2018b) Remote sensing application for deadwood identification and characterisation. In: Geophysical Research Abstracts of the 20th EGU General Assembly held 4–13 April, 2018 in Vienna, Austria, vol 20, EGU2018-16440-1, p 16440. https://ui.adsabs.harvard.edu/abs/2018EGUGA..2016440M/abstract
Maroschek M, Rammer W, Lexer MJ (2015) Using a novel assessment framework to evaluate protective functions and timber production in Austrian mountain forests under climate change. Reg Environ Chang 15:1543–1555. https://doi.org/10.1007/s10113-014-0691-z
Martin RV (2008) Satellite remote sensing of surface air quality. Atmos Environ 42:7823–7843. https://doi.org/10.1016/j.atmosenv.2008.07.018
Masek JG, Goward SN, Kennedy RE et al (2013) United states forest disturbance trends observed using landsat time series. Ecosystems 16:1087–1104. https://doi.org/10.1007/s10021-013-9669-9
Matikainen L, Karila K, Litkey P et al (2020) Combining single photon and multispectral airborne laser scanning for land cover classification. ISPRS J Photogramm Remote Sens 164:200–216. https://doi.org/10.1016/j.isprsjprs.2020.04.021
McDowell NG, Coops NC, Beck PSA et al (2015) Global satellite monitoring of climate-induced vegetation disturbances. Trends Plant Sci 20:114–123. https://doi.org/10.1016/j.tplants.2014.10.008
Meiforth JJ, Buddenbaum H, Hill J, Shepherd JD, Dymond JR (2020) Stress Detection in New Zealand Kauri Canopies with WorldView-2 Satellite and LiDAR Data. Remote Sens 12:1906. https://doi.org/10.3390/rs12121906
Meng R, Dennison PE, Zhao F et al (2018) Mapping canopy defoliation by herbivorous insects at the individual tree level using bi-temporal airborne imaging spectroscopy and LiDAR measurements. Remote Sens Environ 215:170–183. https://doi.org/10.1016/j.rse.2018.06.008
Mitchard ETA, Saatchi SS, White LJT et al (2012) Mapping tropical forest biomass with radar and spaceborne LiDAR in Lopé National Park, Gabon: overcoming problems of high biomass and persistent cloud. Biogeosciences 9:179–191. https://doi.org/10.5194/bg-9-179-2012
Moghaddam M, Saatchi S, Cuenca RH (2000) Estimating subcanopy soil moisture with radar. J Geophys Res Atmos 105:14899–14911. https://doi.org/10.1029/2000JD900058
Monnet J, Mermin E, Chanussot J, Berger F (2010) Using airborne laser scanning to assess forest protection function against rockfall. Interpraevent International Symposium in Pacific Rim, April 2010, Taipei, Taiwan, pp 586–594. hal-00504706
Morresi D, Vitali A, Urbinati C, Garbarino M (2019) Forest spectral recovery and regeneration dynamics in stand-replacing wildfires of central Apennines derived from Landsat time series. Remote Sens 11:308–325. https://doi.org/10.3390/rs11030308
Mullen K, Yuan F, Mitchell M (2018) The mountain pine beetle epidemic in the Black Hills, South Dakota: the consequences of long term fire policy, climate change and the use of remote sensing to enhance mitigation. J Geogr Geol 10:69. https://doi.org/10.5539/jgg.v10n1p69
Mura M, Bottalico F, Giannetti F et al (2018) Exploiting the capabilities of the Sentinel-2 multi spectral instrument for predicting growing stock volume in forest ecosystems. Int J Appl Earth Obs Geoinf 66:126–134. https://doi.org/10.1016/j.jag.2017.11.013
Nabuurs GJ, Delacote P, Ellison D et al (2017) By 2050 the mitigation effects of EU forests could nearly double through climate smart forestry. Forests 8:1–14. https://doi.org/10.3390/f8120484
Næsset E (1997) Estimating timber volume of forest stands using airborne laser scanner data. Remote Sens Environ 61:246–253. https://doi.org/10.1016/S0034-4257(97)00041-2
Nagendra H, Lucas R, Honrado JP et al (2013) Remote sensing for conservation monitoring: assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol Indic 33:45–59. https://doi.org/10.1016/j.ecolind.2012.09.014
Newnham GJ, Armston JD, Calders K et al (2015) Terrestrial laser scanning for plot-scale forest measurement. Curr For Rep 1:239–251. https://doi.org/10.1007/s40725-015-0025-5
Ningthoujam RK, Balzter H, Tansey K et al (2016) Airborne S-band SAR for forest biophysical retrieval in temperate mixed forests of the UK. Remote Sens 8:1–22. https://doi.org/10.3390/rs8070609
Ollinger SV, Goodale CL, Hayhoe K, Jenkins JP (2008) Potential effects of climate change and rising CO2 on ecosystem processes in northeastern U.S. forests. Mitig Adapt Strateg Glob Chang 13:467–485. https://doi.org/10.1007/s11027-007-9128-z
Olsson PO, Lindström J, Eklundh L (2016) Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI. Remote Sens Environ 181:42–53. https://doi.org/10.1016/j.rse.2016.03.040
Ottosen T-B, Petch G, Hanson M, Skjøth CA (2020) Tree cover mapping based on Sentinel-2 images demonstrate high thematic accuracy in Europe. Int J Appl Earth Obs Geoinf 84:101947. https://doi.org/10.1016/j.jag.2019.101947
Pablos M, González-Zamora Á, Sánchez N, Martínez-Fernández J (2018) Assessment of root zone soil moisture estimations from SMAP, SMOS and MODIS observations. Remote Sens 10:981–1000. https://doi.org/10.3390/rs10070981
Päivinen R, Van Brusselen J, Schuck A (2009) The growing stock of European forests using remote sensing and forest inventory data. Forestry 82:479–490. https://doi.org/10.1093/forestry/cpp017
Panagiotidis D, Abdollahnejad A, Surový P, Kuželka K (2019) Detection of fallen logs from high-resolution UAV images. New Zeal J For Sci 49. https://doi.org/10.33494/nzjfs492019x26x
Paquette A, Vayreda J, Coll L et al (2018) Climate change could negate positive tree diversity effects on forest productivity: a study across five climate types in Spain and Canada. Ecosystems 21:960–970. https://doi.org/10.1007/s10021-017-0196-y
Paul-Limoges E, Damm A, Hueni A et al (2018) Effect of environmental conditions on sun-induced fluorescence in a mixed forest and a cropland. Remote Sens Environ 219:310–323. https://doi.org/10.1016/j.rse.2018.10.018
Peña MA, Altmann SH (2009) Use of satellite-derived hyperspectral indices to identify stress symptoms in an Austrocedrus chilensis forest infested by the aphid Cinara cupressi. Int J Pest Manag 55:197–206. https://doi.org/10.1080/09670870902725809
Persson Å, Holmgren J (2004) Tree species classification of individual trees in Sweden by combining high resolution laser data with high resolution near-infrared digital images. Int Arch Photogramm Remote Sens Spat Inf Sci 36:204–207
Peuhkurinen J, Tokola T, Plevak K, Sirparanta S, Kedrov A, Pyankov S (2018) Predicting Tree Diameter Distributions from Airborne Laser Scanning, SPOT 5 Satellite, and Field Sample Data in the Perm Region, Russia. Forests 9:639. https://doi.org/10.3390/f9100639
Pontius J, Schaberg P, Hanavan R (2020) Remote sensing for early, detailed, and accurate detection of forest disturbance and decline for protection of biodiversity. In: Cavender-Bares J, Gamon JA, Townsend PA (eds) Remote sensing of plant biodiversity. Springer, Cham
Potapov P, Yaroshenko A, Turubanova S et al (2008) Mapping the world’s intact forest landscapes by remote sensing. Ecol Soc 13(2):51–66. https://doi.org/10.5751/ES-02670-130251
Pouliot DA, King DJ, Pitt DG (2005) Development and evaluation of an automated tree detection-delineation algorithm for monitoring regenerating coniferous forests. Can J For Res 35:2332–2345. https://doi.org/10.1139/x05-145
Puliti S, Solberg S, Granhus A (2019) Use of UAV photogrammetric data for estimation of biophysical properties in forest stands under regeneration. Remote Sens 11:233. https://doi.org/10.3390/rs11030233
Quegan S, Le Toan T, Chave J et al (2019) The European Space Agency BIOMASS mission: Measuring forest above-ground biomass from space. Remote Sens Environ 227:44–60. https://doi.org/10.1016/j.rse.2019.03.032
Rao K, Anderegg WRL, Sala A et al (2019) Satellite-based vegetation optical depth as an indicator of drought-driven tree mortality. Remote Sens Environ 227:125–136. https://doi.org/10.1016/j.rse.2019.03.026
Rasel SMM, Groen TA, Hussin YA, Diti IJ (2017) Proxies for soil organic carbon derived from remote sensing. Int J Appl Earth Obs Geoinf 59:157–166. https://doi.org/10.1016/j.jag.2017.03.004
Rees WG, Williams M (1997) Monitoring changes in land cover induced by atmospheric pollution in the Kola Peninsula, Russia, using Landsat-MSS data. Int J Remote Sens 18:1703–1723. https://doi.org/10.1080/014311697218061
Reich PB, Oleksyn J (2008) Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol Lett 11(6):588–597
Rock BN, Hoshizaki T, Miller JR (1988) Comparison of in situ and airborne spectral measurements of the blue shift associated with forest decline. Remote Sens Environ 24:109–127. https://doi.org/10.1016/0034-4257(88)90008-9
Röder M, Latifi H, Hill S, Wild J, Svoboda M, Brůna J, Macek M, Nováková MH, Gülch E, Heurich M (2018) Application of optical unmanned aerial vehicle-based imagery for the inventory of natural regeneration and standing deadwood in post-disturbed spruce forests. International Journal of Remote Sensing 39(15-16):5288–5309. https://doi.org/10.1080/01431161.2018.1441568
Rosenzweig C, Casassa G, Karoly DJ et al (2007) Assessment of observed changes and responses in natural and managed systems. In: Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE (eds) Contribution of working group II to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, pp 79–131
Roth KL, Roberts DA, Dennison PE, Alonzo M, Peterson SH, Beland M (2015) Differentiating plant species within and across diverse ecosystems with imaging spectroscopy. Remote Sens Environ 167:135–151. https://doi.org/10.1016/j.rse.2015.05.007
Saksa T, Uuttera J, Kolström T et al (2003) Clear-cut detection in boreal forest aided by remote sensing. Scand J For Res 18:537–546. https://doi.org/10.1080/02827580310016881
Salas C, Ene L, Gregoire TG et al (2010) Modelling tree diameter from airborne laser scanning derived variables: a comparison of spatial statistical models. Remote Sens Environ 114:1277–1285. https://doi.org/10.1016/j.rse.2010.01.020
Sánchez-Ruiz S, Chiesi M, Maselli F, Gilabert MA (2016) Mapping growing stock at 1-km spatial resolution for Spanish forest areas from ground forest inventory data and GLAS canopy height. In: Proceedings of SPIE 10005, Earth Resources and Environmental Remote Sensing/GIS Applications VII, 100051I (18 October 2016). https://doi.org/10.1117/12.2241166
Sandström J, Bernes C, Junninen K et al (2019) Impacts of dead wood manipulation on the biodiversity of temperate and boreal forests. A systematic review. J Appl Ecol 56:1770–1781. https://doi.org/10.1111/1365-2664.13395
Schnelle F, Volkert E (1974) International phenological gardens in Europe. The basic network for international phenological observations. In: Lieth H (ed) Phenology and seasonality modeling. Ecological studies (Analysis and synthesis), vol 8. Springer, Berlin/Heidelberg. https://doi.org/10.1007/978-3-642-51863-8_32
Seidl R, Albrich K, Erb K et al (2019) What drives the future supply of regulating ecosystem services in a mountain forest landscape? For Ecol Manag 445:37–47. https://doi.org/10.1016/j.foreco.2019.03.047
Seppälä R, Buck A, Katila P (eds) (2009) Adaptation of forests and people to climate change. A global assessment report. IUFRO World Series Volume 22. Helsinki, 224 p
Shi L, Liu S (2017) Methods of estimating forest biomass: a review. In: Tumuluru JS (ed) Biomass volume estimation and valorization for energy. IntechOpen. https://doi.org/10.5772/65733
Simard M, Pinto N, Fisher JB, Baccini A (2011) Mapping forest canopy height globally with spaceborne lidar. J Geophys Res Biogeosci 116:1–12. https://doi.org/10.1029/2011JG001708
Šímová I, Storch D (2017) The enigma of terrestrial primary productivity: measurements, models, scales and the diversity–productivity relationship. Ecography 40(239–252):2017. https://doi.org/10.1111/ecog.02482
Sinha S, Santra A, Das AK et al (2019) Regression-based integrated Bi-sensor SAR data model to estimate forest carbon stock. J Indian Soc Remote Sens 47:1599–1608. https://doi.org/10.1007/s12524-019-01004-7
Skakun RS, Wulder MA, Franklin SE (2003) Sensitivity of the Thematic Mapper Enhanced Wetness Difference Index (EWDI) to detect mountain pine needle red-attack damage. Remote Sens Environ 86(4):433–443. https://doi.org/10.1016/S0034-4257(03)00112-3
Smigaj M, Gaulton R, Suárez JC, Barr SL (2019) Canopy temperature from an Unmanned Aerial Vehicle as an indicator of tree stress associated with red band needle blight severity. For Ecol Manag 433:699–708. https://doi.org/10.1016/j.foreco.2018.11.032
Smith G, Askne J (2001) Clear-cut detection using ERS interferometry. Int J Remote Sens 22:3651–3664. https://doi.org/10.1080/01431160110040477
Spriggs RA, Coomes DA, Jones TA et al (2017) An alternative approach to using LiDAR remote sensing data to predict stem diameter distributions across a temperate forest landscape. Remote Sens 9:944. https://doi.org/10.3390/rs9090944
Stone C, Mohammed C (2017) Application of remote sensing technologies for assessing planted forests damaged by insect pests and fungal pathogens: a review. Curr For Rep 3:75–92. https://doi.org/10.1007/s40725-017-0056-1
Szpakowski DM, Jensen JLR (2019) A Review of the Applications of Remote Sensing in Fire Ecology. Remote Sens 11:2638. https://doi.org/10.3390/rs11222638
Tang H, Armston J, Hancock S et al (2019) Characterizing global forest canopy cover distribution using spaceborne lidar. Remote Sens Environ 231:111262. https://doi.org/10.1016/j.rse.2019.111262
Teich M, Bebi P (2009) Evaluating the benefit of avalanche protection forest with GIS-based risk analyses – a case study in Switzerland. For Ecol Manag 257:1910–1919. https://doi.org/10.1016/j.foreco.2009.01.046
Thomas V, Oliver RD, Lim K, Woods M (2008) LiDAR and Weibull modeling of diameter and basal area. For Chron 84:866–875. https://doi.org/10.5558/tfc84866-6
Torresan C, Corona P, Scrinzi G, Marsal JV (2016) Using classification trees to predict forest structure types from LiDAR data. Ann For Res 59(2):281–298. https://doi.org/10.15287/afr.2016.423
Vacchiano G, Berretti R, Motta R, Mondino EB (2018) Assessing the availability of forest biomass for bioenergy by publicly available satellite imagery. IForest 11:459–468. https://doi.org/10.3832/ifor2655-011
Varhola A, Coops NC (2013) Estimation of watershed-level distributed forest structure metrics relevant to hydrologic modeling using LiDAR and Landsat. J Hydrol 487:70–86. https://doi.org/10.1016/j.jhydrol.2013.02.032
Vastaranta M, Saarinen N, Yrttimaa T, Kankare V (2020) Monitoring forests in space and time using close-range sensing. https://doi.org/10.20944/preprints202002.0300.v1
Verkerk PJ, Costanza R, Hetemäki L et al (2020) Climate-smart forestry: the missing link. For Policy Econ 115:102164. https://doi.org/10.1016/j.forpol.2020.102164
Viccaro M, Cozzi M, Fanelli L, Romano S (2019) Spatial modelling approach to evaluate the economic impacts of climate change on forests at a local scale. Ecol Indic 106:105523. https://doi.org/10.1016/j.ecolind.2019.105523
Vincent G, Caron F, Sabatier D, Blanc L (2012) LiDAR shows that higher forests have more slender trees. Bois Forets des Tropiques 66:51–56
Vittucci C, Ferrazzoli P, Kerr Y et al (2016) SMOS retrieval over forests: exploitation of optical depth and tests of soil moisture estimates. Remote Sens Environ 180:115–127. https://doi.org/10.1016/j.rse.2016.03.004
Wallace L, Lucieer A, Malenovskỳ Z et al (2016) Assessment of forest structure using two UAV techniques: a comparison of airborne laser scanning and structure from motion (SfM) point clouds. Forests 7:1–16. https://doi.org/10.3390/f7030062
Walsh SJ, Weiss DJ, Butler DR, Malanson GP (2004) An assessment of snow avalanche paths and forest dynamics using Ikonos satellite data. Geocarto Int 19:85–93. https://doi.org/10.1080/10106040408542308
Wang R, Gamon JA (2018) Remote sensing of terrestrial plant biodiversity. Remote Sens Environ 231:111218. https://doi.org/10.1016/j.rse.2019.111218
Wang C-J, Zhang Z-X, Wan J-Z (2019) Vulnerability of global forest ecoregions to future climate change. Glob Ecol Conserv 20:1–10. https://doi.org/10.1016/j.gecco.2019.e00760
Waring RH, Coops NC, Landsberg JJ (2010) Improving predictions of forest growth using the 3-PGS model with observations made by remote sensing. For Ecol Manag 259:1722–1729. https://doi.org/10.1016/j.foreco.2009.05.036
Warren SD, Alt M, Olson KD, Irl SDH, Steinbauer MJ, Jentsch A (2014) The relationship between the spectral diversity of satellite imagery, habitat heterogeneity, and plant species richness. Ecol Informatics 24:160–168. https://doi.org/10.1016/j.ecoinf.2014.08.006
Weatherall A, Nabuurs G-J, Velikova V et al (2021) Defining Climate-Smart Forestry. In: Managing Forest Ecosystems, Vol. 40, Tognetti R, Smith M, Panzacchi P (Eds): Climate-Smart Forestry in Mountain Regions. Springer Nature, Switzerland, AG
Weinstein BG, Marconi S, Bohlman S et al (2019) Individual tree-crown detection in rgb imagery using semi-supervised deep learning neural networks. Remote Sens 11:1–13. https://doi.org/10.3390/rs11111309
Weiskittel AR, Crookston NL, Radtke PJ (2011) Linking climate, gross primary productivity, and site index across forests of the western United States. Can J For Res 41(8). https://doi.org/10.1139/x11-086
Weiss DJ, Walsh SJ (2009) Remote sensing of mountain environments. Geogr Compass 3:1–21. https://doi.org/10.1111/j.1749-8198.2008.00200.x
Wigneron JP, Jackson TJ, O’Neill P et al (2017) Modelling the passive microwave signature from land surfaces: a review of recent results and application to the L-band SMOS & SMAP soil moisture retrieval algorithms. Remote Sens Environ 192:238–262. https://doi.org/10.1016/j.rse.2017.01.024
Willis KS (2015) Remote sensing change detection for ecological monitoring in United States protected areas. Biol Conserv 182:233–242. https://doi.org/10.1016/j.biocon.2014.12.006
Xu X, Li J, Tolson BA (2014) Progress in integrating remote sensing data and hydrologic modelling. Prog Phys Geogr 38(4):464–498. https://doi.org/10.1177/0309133314536583
Yang H, Hu D, Xu H, Zhing X (2020) Assessing the spatiotemporal variation of NPP and its response to driving factors in Anhui province, China. Environ Sci Pollut Res 27:14915–14932. https://doi.org/10.1007/s11356-020-08006-w
Zarea A, Mohammadzadeh A (2016) A novel building and tree detection method from LiDAR data and aerial images. IEEE J Sel Top Appl Earth Obs Remote Sens 9:1864–1875. https://doi.org/10.1109/JSTARS.2015.2470547
Zimble DA, Evans DL, Carlson GC, Parker RC, Grado SC, Gerard PD (2003) Characterizing vertical forest structure using small-footprint airborne LiDAR. Remote Sens Environ 87(2–3):171–182. https://doi.org/10.1016/S0034-4257(03)00139-1
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2022 The Author(s)
About this chapter

Cite this chapter
Torresan, C., Luyssaert, S., Filippa, G., Imangholiloo, M., Gaulton, R. (2022). Remote Sensing Technologies for Assessing Climate-Smart Criteria in Mountain Forests. In: Tognetti, R., Smith, M., Panzacchi, P. (eds) Climate-Smart Forestry in Mountain Regions. Managing Forest Ecosystems, vol 40. Springer, Cham. https://doi.org/10.1007/978-3-030-80767-2_11
Download citation
DOI: https://doi.org/10.1007/978-3-030-80767-2_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-80766-5
Online ISBN: 978-3-030-80767-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)