
Abstract
Background: Naringin is a major antioxidant in Citrus fruits and herbs. To clarify molecular forms distributed to various tissues, we investigated tissue distribution of naringin and relevant metabolites in rats after repeated dosing.
Methods: Male Sprague-Dawley rats were orally administered naringin (210 mg/kg) twice daily for eight days. At 6 h post the 17th dose, various tissues including liver, kidney, heart, spleen and brain were collected and analyzed by HPLC method before and after hydrolysis with β-glucuronidase and sulfatase, individually.
Results: The free forms of naringin and naringenin were not detected in all the tissues assayed. Liver contained the highest concentration of naringenin sulfates, followed by spleen, heart, brain and kidney. Naringenin glucuronides were present in liver and kidney, but not in spleen, brain and heart.
Conclusion: The bioavailability of naringenin glucuronides and sulfates supported its application for personalized medicine.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1. Introduction
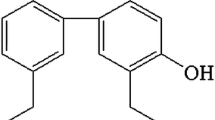
Flavonoids, major class of antioxidants, prove interesting to pharmacologists due to various beneficial bioactivities [1,2]. They also draw attention from pharmaceutical researchers via capacity for modulating P-glycoprotein (P-gp) and CYP 3A4 [3-6], both prominent in pharmacokinetics [7, 8]. Naringin (4’, 5, 7-trihydroxyflavanone 7-rhamnoglucoside) is one major flavonoid distributed in Citrus fruits like grapefruit (C. paradisi) or pomelo (C. grandis) and in Chinese herbs like C. aurantium and C. maxima [9]. Numerous in vitro studies report naringin and naringenin, the aglycon of naringin, (structures in Fig. 1) exhibiting benefits: e.g., anticancer [10,11], superoxide scavenging, antioxidation [12,13], antimicrobial effects [14]. Naringin-containing nutraceuticals are increasingly used as dietary supplements, yet based on our prior studies reporting pharmacokinetics of naringin in rabbits and rats, n aringenin sulfates and glucuronides appeared predominately in the blood-stream, with no trace of naringin or naringenin detected [15,16]. Whether bioactivities of naringin and naringenin cited by earlier in vitro study can be extrapolated to in vivo effects remained unanswered [17]. Regarding major forms in various organs, previous studies report tissue distribution of naringin or naringenin in rats [18-20], yet these followed single dose via intravenous or oral route, findings less than consistent. We analyzed distribution of naringin and relevant metabolites in rat tissue after repeated dosing.
2. Materials and methods
2.1. Chemicals
Naringin (purity 95%), (±)-naringenin (purity 95%), β-glucuronidase (Type B-1, 666,400 units/g, from bovine liver), sulfatase (Type H-1, 20,000 units/g, from Helix pomotia, containing 498,800 units/g of β-glucuronidase) and vanillin were purchased from Sigma Chemical Co. (St. Louis, MO); 5,7-Dimethoxycoumarin (99%) from Aldrich (Milwaukee, WI); acetonitrile, methanol and ethyl acetate of HPLC grade from Mallinckrodt Baker, Inc. (Phillipsburg, NJ); L(+)-Ascorbic acid from RdH Laborchemikalien GmbH & Co. KG (Seelze, Germany). Other reagents were of analytical grade, Milli-Q plus water (Millipore, Bedford, MA) used throughout.

Chemical structures of naringin and naringenin.
2.2. Instrumentation and HPLC conditions
The apparatus was equipped with a pump (LC-10AT, Shimadzu, Japan), UV detector (SPD-10A, Shimadzu, Japan), automatic injector (SIL-10A, Shimadzu, Japan) and Cosmosil C18 column (5 μm, 150×4.6mm, Waters, MA). Detection wavelength was set at 288 nm. Mobile phase consisted of acetonitrile: 0.1% phosphoric acid (36:64, v/v), flow rate 1.0 mL/min for naringenin assay in serum and tissue homogenate.
2.3. Drug administration and collection of blood and organs
Six male Sprague-Dawley rats weighing 300-350 g were purchased from the National Science Council, Taipei, and maintained in the China Medical University Animal Center. Naringin was dispersed in warm water for oral administration (210 mg/kg twice daily) for 17 doses via gastric gavage. Finally, rats were fasted overnight before 17th dose, blood and various organs were collected at 6 h after dosing, based on peak time of naringenin conjugates observed by our previous pharmacokinetic study [16]. Immediately after blood collection, systemic perfusion was conducted by pumping normal saline to wash out blood. The organs including liver, kidney, spleen, heart and brain were dissected, blotted dry, accurately weighed, and frozen at -80℃. Animal study adhered to Guidebook for the Care and Use of Laboratory Animals (2002) published by The Chinese Society for the Laboratory Animal Science, Taiwan.
2.4. Preparation and quantitation of tissue samples
All tissue samples were lyophilized, then chopped into small pieces and milled with normal saline (300 μL/g tissue) by Potter-Elvehjem tissue grinder (Kontes Glass Co.; Vineland, NJ). Homogenates (0.5 mL) were deproteinized with 3-fold methanol. After centrifuge, supernatant was evaporated to dryness under vacuum, then dissolved with 0.5 mL of pH 5 acetate buffer to afford tissue extract. To quantify naringin, tissue extract was analyzed before and after hydrolysis with β-glucuronidase and sulfatase, individually. To quantify naringin conjugated metabolites, tissue extract (100 μL) was mixed with 100 μL of glucuronidase (1000 units/mL in pH 5 buffer) and/or sulfatase (containing 1000 units/mL of sulfatase and 24,940 units/mL of glucuronidase in pH 5 buffer), 100 μL of ascorbic acid (150 mg/mL) and incubated at 37°C for 2 h, mixture added with 900 μL of acetonitrile (containing 6 μg/mL of vanillin). For free form determination of naringin, homogenate was treated with pH 5 acetate buffer without incubation with glucuronidase or sulfatase and processed as per procedure detailed above. Acetonitrile layer evaporated under N2 gas to dryness was reconstituted with appropriate volume of acetonitrile, 20 μL subjected to HPLC analysis. To quantify free form naringenin, tissue extract was determined before hydrolysis with sulfatase or β-glucuronidase. Briefly, 100 μL of the deproteinized tissue extract was acidified with 100 μL of 0.1 N HCl and partitioned with 500 μL of ethyl acetate (containing 2.0 μg/mL of 5,7-dimethoxycoumarin as the internal standard). The ethyl acetate layer was evaporated under N2 gas to dryness and 20 μL was subject to HPLC analysis.
To quantify naringenin glucuronides, 100 μL of buffer containing tissue extract was mixed with 100 μL of β-glucuronidase (1000 units/mL in pH 5 buffer), 100 μL of ascorbic acid (150 mg/mL) and incubated at 37°C for 2 h. To quantify naringenin sulfates/glucuronides, 100 μL of the buffer containing tissue extract were mixed with 100 μL of sulfatase (containing 1000 units/mL of sulfatase and 24,940 units/mL of glucuronidase in pH 5 buffer), 100 μL of ascorbic acid (150 mg/mL), and incubated at 37°C for 1 h. After hydrolysis, procedure was the same as described above for free form naringenin. For calibrator preparation, 100 μL of tissue standards with various concentrations of naringenin were spiked with 100 μL of pH 5 acetate buffer, 100 μL of ascorbic acid (150 mg/mL), then added to 100 μL of 0.1 N HCl. Later procedure followed that described above. Calibration graph was plotted by linear regression of peak area ratios (naringenin to internal standard) against concentrations of naringenin.
2.5. Data analysis
Concentrations of naringenin glucuronides and naringenin sulfates in each tissue were expressed in μmol per gram of tissues (μmol/g). Ratios (mL/g) of concentrations of naringin glucuronides and naringin sulfates in various organs (μmol/g) to those in serum (μmol/mL) were calculated, although units differed.
3. Results
Quantitation method of naringenin in each tissue was established and optimized. Good linearity was obtained at concentration 0.4-50.0 μg/mL of naringenin in each tissue. Validation of assay indicated all coefficients of variation (CVs) and relative errors of intra-run and inter-run analysis below 2.2% and 18.1%, respectively. The LLOQ and LOD of naringenin were 0.40 and 0.02 μg/mL, respectively, in all tissues. Results indicated neither naringin nor naringenin in liver, spleen, heart, brain and kidney, molecular forms in tissues as naringenin sulfates and naringenin glucuronides. Table 1 plots concentrations of naringenin sulfate and naringenin glucuronide in serum and various organs 6 h post 17th dose. Results indicate the liver with highest ratio of naringenin sulfates, followed by spleen, heart brain, and kidney. Naringenin glucuronides were present in liver and kidney, not in spleen, brain, or heart.
Regarding analysis of serum collected 6 h post 17th dose, quantiation showed naringenin glucuronides and naringenin sulfates as major forms, whereas naringin and naringenin were not present, which was consistent with previous studies [15, 16]. Fig. 2 shows ratios of naringenin glucuronides and naringenin sulfates in each tissue to those in serum. Among assayed organs, liver had the highest naringenin sulfate level, about 10.7-fold; spleen, heart and brain had higher concentrations of naringin sulfates than serum by 386, 100, and 57 %, respectively. In regard to distribution of naringenin glucuronides, conversely, liver and kidney contained lower concentrations than serum by 30 and 76%, respectively.
4. Discussion
Quantitation method of naringenin glucuronides and sulfates in various tissues was established and validated. Due to considerable amount of β-glucuronidase in sulfatase (Type H-1), this enzyme hydrolyzed both sulfates and glucuronides simultaneously. Results of both hydrolysis with sulfatase and glucuronidase could yield individual naringenin sulfate and naringenin glucuronide concentrations in each tissue specimen. Quantitation of serum and collected tissues showed that after repeated dosing, free forms of naringin and naringenin were not detected in both bloodstream and all assayed organs; major molecular forms were naringenin sulfates and naringenin glucuronides. In serum, naringenin glucuronides were predominant; in liver, spleen, heart and brain, narigenin sulfate was the major form. Findings imply naringenin glucuronides deglucuronidated and then sulfated in these organs.

Relative mean (±S.E.) ratios of concentrations of naringenin glucuronides and naringenin sulfates in various tissues to serum at 6 h after repeated doses of naringin (210 mg/kg) twice daily for nine days to six rats.
Among assayed organs, the liver contained higher naringenin sulfate and naringenin glucuronide concentrations than other organs, which concurred with previous studies [19] and indicated these naringenin conjugates have higher protein binding with liver proteins. In the liver, concentration of naringenin sulfates was higher than that of naringenin glucuronides by 240%. In spleen, heart, and brain, naringenin glucuronides were not detected, meaning naringenin released through hydrolysis with sulfatase in these organs was solely from naringenin sulfates. It can thus be assumed that when naringenin glucuronides entered liver, spleen, heart, or brain from circulation, they were hydrolyzed by glucuronidase, then sulfated by sulfotransferase in these organs [21,22]. In sum, liver, spleen, heart and brain contained narigenin sulfates as principal metabolites of naringin. Therefore, bioactivities of naringenin sulfates in liver, spleen, heart and brain warrant more investigations. In the kidney, naringenin glucuronides and naringenin sulfates manifested far lower concentrations than those in serum, indicating that only a little fraction of glucuronides or sulfates had entered the kidney. This finding was not consistent with previous study reporting moderate concentrations of naringenin glucuronides detected in kidney, liver and brain at 2 h post oral dose of naringenin in rats [18]; future studies must clarify.
Our study revealing absence of naringenin in all assayed tissues was not in good agreement with prior research detecting naringenin in tissues after single dose of naringin [19]. This discrepancy might arise from variant dosage or detection method: i.e., 17 doses of naringin might modulate expression of UDP-glucuronosyltransferase or sulfotransferase and result in more extensive metabolism [23]. Repeated dosing of naringin to rats yielded wide distribution of naringenin sulfates to various organs, while naringin and naringenin reached no organs. We identified chemical nature and concentration of naringenin conjugates in tissue; these may disclose pharmacological roles of putative active metabolites after chronic dosing of naringin.
Acknowledgements
The work was in part supported by National Science Council (NSC 102-2320-B-039-014-MY2, NSC 102-2320- B-039-008), and China Medical University, Taiwan, R.O.C. (CMU101- N2-08, CMU102-S-16).
Declaration of Interest: Authors declare no conflicts of interest for this work.
References
CooK NC, Samman S. Flavonoids—Chemistry, metabolism, cardioprotective effects, and dietary sources. J Nutr Biochem 1996; 7: 66–76.
Middleton E, Jr., Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Rev 2000; 52: 673-751.
Miniscalco A, Lundahl J, Regardh CG, Edgar B, Eriksson UG. Inhibition of dihydropyridine metabolism in rat and human liver microsomes by flavonoids found in grapefruit juice. J Pharmacol Exp Ther 1992; 261: 1195–1199.
Critchfield JW, Welsh CJ, Phang JM, Yeh GC. Modulation of adriamycin accumulation and efflux by flavonoids in HCT-15 colon cells. Activation of P-glycoprotein as a putative mechanism. Biochem Pharmacol 1994; 48: 1437–1445.
Scambia G, Ranelletti FO, Panici PB, De Vincenzo R, Bonanno G, Ferrandina G, et al. Quercetin potentiates the effect of adriamycin in a multidrug-resistant MCF-7 human breast-cancer cell line: P-glycoprotein as a possible target. Cancer Chemother Pharmacol 1994; 34: 459–464.
Zhang S, Morris ME. Effects of the flavonoids biochanin A, morin, phloretin, and silymarin on P-glycoproteinmediated transport. J Pharmacol Exp Ther 2003; 304: 1258–1267.
Lin JH, Yamazaki M. Role of P-glycoprotein in pharmacokinetics: clinical implications. Clin Pharmacokinet 2003; 42: 59–98.
Estudante M, Morais JG, Soveral G, Benet LZ. Intestinal drug transporters: an overview. Adv Drug Deliv Rev 2013; 65: 1340–1356.
Lu Y, Zhang C, Bucheli P, Wei D. Citrus flavonoids in fruitand traditional Chinese medicinal food ingredients in China. Plant Foods Hum Nutr 2006; 61: 57–65.
Frydoonfar HR, McGrath DR, Spigelman AD. The variable effect on proliferation of a colon cancer cell line by the citrus fruit flavonoid Naringenin. Colorectal Dis 2003; 5: 149–152.
Harmon AW, Patel YM. Naringenin inhibits glucose uptake in MCF-7 breast cancer cells: a mechanism for impaired cellular proliferation. Breast Cancer Res Treat 2004; 85: 103–110.
Heo HJ, Kim DO, Shin SC, Kim MJ, Kim BG, Shin DH. Effect of antioxidant flavanone, naringenin, from Citrus junoson neuroprotection. J Agric Food Chem 2004; 52: 1520–1525.
Cavia-Saiz M, Busto MD, Pilar-Izquierdo MC, Ortega N, Perez-Mateos M, Muñiz P. Antioxidant properties, radical scavenging activity and biomolecule protection capacity of flavonoid naringenin and its glycoside naringin: a comparative study. J Sci Food Agric 2010; 90: 1238–1244.
Celiz G, Daz M, Audisio MC. Antibacterial activity of naringin derivatives against pathogenic strains. J Appl Microbiol 2011; 111: 731–738.
Hsiu SL, Huang TY, Hou YC, Chin DH, Chao PD. Comparison of metabolic pharmacokinetics of naringin and naringenin in rabbits. Life Sci 2002; 70: 1481–1489.
Wang MJ, Chao PD, Hou YC, Hsiu SL, Wen KC, Tsai SY. Pharmacokinetics and conjugation metabolism of naringin and naringenin in rats after single dose and multiple dose administrations. J Food Drug Anal 2006; 14: 247–253.
Kroon PA, Clifford MN, Crozier A, Day AJ, Donovan JL, Manach C, Williamson G. How should we assess the effects of exposure to dietary polyphenols in vitro? Am J Clin Nutr 2004; 80: 15–21.
El Mohsen MA, Marks J, Kuhnle G, Rice-Evans C, Moore K, Gibson G, et al. The differential tissue distribution of the citrus flavanone naringenin following gastric instillation. Free Radic Res 2004; 38: 1329–1340.
Zou W, Yang C, Liu M, Su W. Tissue distribution study of naringin in rats by liquid chromatography-tandem mass spectrometry. Arzneimittelforschung 2012; 62: 181–186.
Peng HW, Cheng FC, Huang YT, Chen CF, Tsai TH. Determination of naringenin and its glucuronide conjugate in rat plasma and brain tissue by high-performance liquid chromatography. J Chromatogr B Biomed Sci Appl 1998; 714: 369–374.
Slaus K, Coughtrie MW, Sharp S, Vanhaecke T, Vercruysse A, Rogiers V. Influence of culture system and medium enrichment on sulfotransferase and sulfatase expression in male rat hepatocyte cultures. Biochem Pharmacol 2001; 61: 1107–1117.
O’Leary KA, Day AJ, Needs PW, Mellon FA, O’Brien NM, Williamson G. Metabolism of quercetin-7- and quercetin- 3-glucuronides by an in vitro hepatic model: the role of human beta-glucuronidase, sulfotransferase, catechol- O-methyltransferase and multi-resistant protein 2 (MRP2) in flavonoid metabolism. Biochem pharmacol 2003; 65: 479–491.
Moon YJ, Wang X, Morris ME. Dietary flavonoids: effects on xenobiotic and carcinogen metabolism. Toxicol In Vitro 2006; 20: 187–210.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Open Access. This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lin, SP., Hou, YC., Tsai, SY. et al. Tissue distribution of naringenin conjugated metabolites following repeated dosing of naringin to rats. BioMed 4, 16 (2014). https://doi.org/10.7603/s40681-014-0016-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.7603/s40681-014-0016-z