
Abstract
Studies using retrospective memory tasks have revealed that animates/living beings are better remembered than are inanimates/nonliving things (the animacy effect). However, considering that memory is foremost future oriented, we hypothesized that the animacy effect would also occur in prospective memory (i.e., memory for future intentions). Using standard prospective memory (PM) procedures, we explored this hypothesis by manipulating the animacy status of the PM targets. Study 1a reports data collected from an American sample; these results were then replicated with a Portuguese sample (Study 1b). Study 2 employed a new procedure, and data were collected from a broader English-speaking sample. In these three studies, animate (vs. inanimate) targets consistently led to a better PM performance, revealing, for the first time, that the animacy advantage extends to PM. These results strengthen the adaptive approach to memory and stress the need to consider animacy as an important variable in memory studies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Evolutionary psychology postulates that human cognition (e.g., memory) evolved to help solve adaptive problems, such as finding food and shelter (Cosmides & Tooby, 1992; Nairne et al., 2017). Thus, researchers have hypothesized that there is a memory tuning for fitness-relevant information (i.e., information that enhances our odds of survival and/or reproduction). An example of fitness-relevant information relates to animacy, as animates are fitness-relevant in many ways (e.g., they can represent predators, prey, sexual mates, among others; Nairne et al., 2017). Animacy has been operationalized in many ways (e.g., as a synonym of agency and livingness; for an overview, see Félix et al., 2023). According to VanArsdall and Blunt (2022), the livingness construct loads highly onto the animacy factor; thus, animacy will be conceived here as livingness (as in most memory research; e.g., Nairne et al., 2013), or the distinction between living beings (e.g., humans and nonhuman animals) and nonliving things (e.g., objects). Indeed, people tend to remember animates better than inanimates, a phenomenon called the “animacy effect.”
Since the first report showing that animacy is one of the best predictors of free recall (Nairne et al., 2013), the animacy effect has proven to be robust in retrospective memory tasks (i.e., memory for past events); it has been reported using a variety of procedures, types of to-be-encoded stimuli, and in different languages (e.g., free recall, with French words and pictures as the to-be-remembered stimuli: Bonin et al., 2014; metamemory/judgements of learning in English: DeYoung & Serra, 2021; implicit memory in Spanish: Laurino & Kaczer, 2019; working memory: Daley et al., 2020; directed forgetting in English: Murphy & Castel, 2022). There are, however, some circumstances in which evidence is less clear (e.g., recognition: Bonin et al., 2014; Leding, 2020; cued recall: Popp & Serra, 2016; but see VanArsdall et al., 2015).
Allied with the importance of retrieving information from the past, some authors have suggested that our memory is foremost future oriented: One of our memory’s main function is to store information from the past in order to help solve problems in the present and predict/get prepared for future events, which is crucial for survival (Ingvar, 1985; Klein, 2013; Nairne & Pandeirada, 2008; Schacter et al., 2007). This relates directly with prospective memory (PM), which is the memory for upcoming plans, events, actions or intentions to be performed in the future (Einstein & McDaniel, 1990). Everyday examples of PM tasks are to remember to deliver a message to a friend when he/she is encountered, to take the pills after lunch, or to remember to return books at the library the following day. Importantly, prospective memory tasks mostly involve other people (i.e., animates), and PM successes or failures can impact individuals themselves, as well as their relations with others, thus having clear adaptive consequences. For instance, there would be a benefit conferred by remembering to avoid cheaters in future encounters; likewise, remembering to maintain positive interactions with cooperators in the future would also be advantageous (Schaper et al., 2022). Failure to remember to pick up the kids from school or to remove a clamp from the patient’s abdomen (Brandimonte & Ferrante, 2008; Dembitzer & Lai, 2003) also illustrate this point. In the present work, we aimed to combine these two adaptive elements—the animacy variable and prospective memory—and explore whether animates also confer an advantage in PM performance.
Prospective memory has been studied in the laboratory using several types of tasks (see Kvavilashvili & Ellis, 1996, for further details). Our work focused on event-based tasks, in which the moment to perform the intention is signaled by the presence of a specific event–the PM target (e.g., whenever you see John [target], give him a message [PM response]). Laboratory PM studies usually employ a dual-task paradigm, that is, the PM response occurs while another task is ongoing. In a typical procedure, participants first respond to the ongoing task (e.g., a lexical decision task), which provides a baseline to their performance on that task alone; these are called the baseline trials. Then, the PM instructions are presented: Participants are tasked to provide an alternative response (the PM response) whenever specific targets appear (e.g., press F1 whenever the syllable “TOR” appears; McDaniel & Einstein, 2000) while performing the ongoing task. The trials involving the target words are the target trials, whereas those regarding the ongoing task are now named the filler trials. Although animacy has never been systematically manipulated or analyzed in PM (i.e., was not an independent variable in such studies), animates (e.g., animals) and/or inanimates (e.g., clothing, furniture) have been used as PM targets in event-based PM tasks (e.g., Chen et al., 2014; Marsh et al., 2009). For example, in one of the seminal works on PM (Einstein et al., 2005), among the targets we find an animate (tortoise), an inanimate (dormitory) and an ambiguous word (tornado; although classified as an inanimate, it can be perceived as an animate, due to the sense of self-propelled motion and agency; Lowder & Gordon, 2015). Other studies (e.g., Moyes et al., 2019) have asked participants to provide the PM response whenever targets from a specific category were presented, including categories of animate items (e.g., four-footed animals), ambiguous words (e.g., flowers, fruits), and inanimate words (e.g., metals); once again, no information was provided regarding the influence of animacy on PM performance.
Here, in a series of three studies, we explored the animacy effect in PM. We expected higher PM performance when the target was an animate (e.g., “horse”), comparatively to when it was inanimate (e.g., “shirt”). No strong predictions were made about the animacy effect for the baseline and filler trials. However, as it has been suggested that animates capture attention more automatically than inanimates (e.g., Bugaiska et al., 2019), the former would capture participants’ attention and divert it from the ongoing task. As a result, it would be reasonable to anticipate a decline in the ongoing task when trials (baseline and filler trials) presented animate words compared with inanimate ones.
Again, our main interest was to explore the animacy effect on the PM target trials. Three factors explain the prediction of an animacy advantage in PM: First, both PM and the animacy variable entail adaptive value. Second, people tend to judge animates as more memorable than inanimates for a future memory test (e.g., DeYoung & Serra, 2021); considering that there is a correlation between those judgements of learning and the actual PM performance (Schnitzspahn et al., 2011), it is conceavable that an interplay among metacognitive judgments, animacy, and prospective memory may occur leading to an animacy advantage in PM. Third, most theories on PM were developed based on knowledge about retrospective memory functioning (McDaniel & Einstein, 2000). Consistently, several variables known to influence retrospective memory also influence prospective memory. For instance, emotional words/targets, as compared with neutral ones, enhance both retrospective (e.g., Dewhurst & Parry, 2000) and PM performance (Hostler et al., 2018; May et al., 2015). Also, Smith (2003) found that distinctive words (i.e., targets with a distinctive orthography, such as sphinx), as compared with common orthography words, improved both prospective memory and recognition performance. Given that animates (vs. inanimates) are better remembered in retrospective memory tasks (e.g., free recall: Nairne et al., 2013; working memory: Daley et al., 2020), one could expect the same advantage to occur in PM (e.g., see the relation between working memory and PM; Brewer et al., 2010).
Study 1a
This study used a well-known PM procedure: while performing an ongoing color-matching task (Smith & Hunt, 2014), participants were required to provide an alternative response (PM response) whenever either of two predefined target words (one animate and one inanimate) appeared. Across studies, we included a baseline phase (color-matching task only) and a PM phase (ongoing color-matching task with an embedded PM task). Of particular interest will be the results regarding the PM performance.
Method
Participants
Using G*Power (Version 3.1.9.7; Faul et al., 2007), we predetermined that a sample size of 109 participants was needed to obtain a small-medium effect size, dz = 0.35, with α = .05 and power = .95. A total of 351 Purdue University undergraduate students participated in exchange for course credits. From those, 175 participants were excluded from the analyses: 51 participants were non-English native speakers; 54 did not provide any PM response; 42 reported having cheated and/or not paid attention to the study or had extremely long survey durations (>7.4 hours; which may reveal low engagement with the task and/or low attention, or a start-and-stop behavior throughout the task despite the instruction to respond to the task in just one sitting); 13 participants did not recognize one (or both) target word(s) and did not provide any PM response to those target trials; 10 had more than 50% missing responses to the ongoing task; four participants had low performance on the filler trials/ongoing task (<Grand Mean – 3 SD); and another participant was underaged. See Supplemental Materials for additional information about the sample.
The final sample was composed of 176 participants (31.3% females and 68.8% males; mean age = 19.43 years, SD = 1.17). They were all English native speakers or bi-/multilingual [being proficient in English and other(s) language(s)]. Forty to 46 participants were allocated to each version of the task (see Procedure).
Material
Animate and inanimate words were selected from VanArsdall (2016), which reports animacy norms for a large set of words. Sixteen words were selected for the baseline phase. For the PM phase, a new set of 24 filler and two pairs of target words were selected to increase the generalizability of the results. Two additional words were selected for the practice trials. In all cases, half the words were animate and the other half were inanimate (see Supplemental Materials). The animate and inanimate words were matched along a number of relevant mnemonic variables (see Table 1).
Procedure
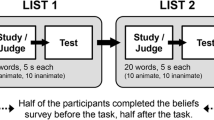
Data were collected online using Qualtrics, in sessions lasting, on average, 25 minutes. The procedure was similar to that used by Smith and Hunt (2014), except that words were presented in a fixed order to every participant (their order was pseudorandomized, ensuring that each quarter of the list had a balanced number of animates and inanimates), and we used fewer trials, aiming for a shorter task; however, we used the same proportion of target trials during the PM phase as in their work (~8%). There were two predetermined presentation orders in the PM phase to ensure that, in each position of the list, an animate and an inanimate item was presented an equal number of times across participants. We also used one out of two sets of PM targets (dancer and bottle / nurse and phone) in each of these versions; their presentation order was predetermined within the list of items and counterbalanced across participants such that, animate and inanimate targets appeared equally in each target position. Therefore, there were four versions of the task to which participants were randomly assigned.
After consenting to participate, participants received the instructions for the ongoing task. Specifically, they were told that six colored squares would be presented, one at a time, each one in a different color (red, yellow, blue, green, pink, orange, or gray). Then, a word would be presented in a colored font. Participants had to decide whether the color font of the word matched the color of any of the just-presented squares by pressing the Y (yes) or the N (no) keys (see Fig. 1 for an illustration of the procedure). Participants started by responding to two practice trials to get familiar with the task. Then, they were reminded of the ongoing task instructions and performed the baseline phase (16 trials; color-match ongoing task only). Throughout the experiment, half of the trials (animate and inanimate) were match-trials (i.e., the color font matched the color of a square), and the other half were nonmatch-trials.

Example of a match-trial, and representation of the presentation times of each stimulus (Study 1a). Note. In this example, the colors of the squares are presented in the following order: red, yellow, orange, blue, gray, and green. The word “COUSIN” is in a red-colored font; the correct response for this trial would be Y (yes). (Color figure online)
After the baseline phase, participants read the PM task instructions which informed them that they were to memorize two new words (an animate and an inanimate word; PM targets). Also, participants were told they would need to press the SPACEBAR (PM response), instead of Y/N, whenever any of these words appeared during the color-matching task. The target words were then displayed simultaneously for one minute. A 2-min distractor task followed (a 3D mental rotation task; Ganis & Kievit, 2015) to prevent participants from rehearsing the PM instructions.
Next, the PM phase began without further reminders of the PM instructions. In this phase, participants were presented with two target and 24 filler words (half animate, half inanimate). To increase the number of PM target trials during the task, each target was presented twice. The filler words were also repeated to prevent target words from becoming distinctive (Smith, 2003); they appeared in a different color each time, once in a match and once in a nonmatch-trial, totalizing 48 filler trials. The PM target trials were presented in the 11th, 24th, 36th, and 51st trials/positions of the list.
Upon completion of the PM phase, participants were asked to recall the instructions they received for the task. They also performed a target recognition test: participants were presented with a short series of six words, one at a time, and asked whether each word corresponded to a PM target (yes/no); both the targets presented in the task and four lures (half animates and half inanimates) from the PM phase were presented. A color-naming task followed. These words and colors were presented one at a time, in a random order for each participant. Finally, participants provided sociodemographic information (age, gender, and native language); also, they responded to “honesty questions” regarding whether they paid attention and answered honestly to the task (Rouse, 2015). They were asked to provide optional feedback regarding the study, were thanked, and debriefed.
Data analyses
Data from all the three studies were analyzed using SPSS 28 (IBM Corp, 2021). The main dependent variable was the proportion of correct responses (i.e., press the correct key—Y, N, or SPACEBAR—in match, nonmatch and target trials, respectively). We conducted a 2 (Animacy: animates vs. inanimates) × 3 (Type of Trial: baseline vs. filler vs. target) repeated-measures analysis of variance (ANOVA; we report the Greenhouse–Geisser corrected data, as the sphericity assumption was violated in all analyses). We used additional paired-sample t tests with Bonferroni corrections (p < .05/3) to clarify some results. Supplemental Materials present additional analyses, namely: on response times, excluded participants, false alarms, as well as data confirming that the overall results here reported hold when we consider the different sets of PM targets used across experiments.
Results
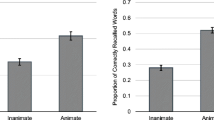
Results are presented in Fig. 2. A significant Animacy main effect was observed, F(1, 175) = 12.48, p = .001, ηp2 = .067, revealing that performance was better for the animate than the inanimate stimuli. The main effect of Type of Trial also reached significance, F(1.29, 224.89) = 38.87, p < .001, ηp2 = .166. A significant Animacy × Type of Trial interaction was also obtained, F(1.20, 209.31) = 18.36, p < .001, ηp2 = .095. The follow-up paired t tests performed on each type of trial revealed that the animacy advantage was significant only on the target trials, t(175) = 4.39, p < .001, dz = 0.33. No animacy advantage was obtained for the baseline, t(175) = −1.27, p = .206, or filler trials, t(175) = −1.08, p = .281.

Mean performance obtained in baseline, filler (ongoing task) and target trials (PM Task), in Study 1a. Error bars represent standard errors of the mean
Study 1b
Study 1a was the first study reporting an animacy advantage in PM: Animate targets elicited better PM performance than the inanimate targets did. As with any first discovery, more empirical evidence is needed for the effect to be considered reliable. Study 1b aimed to replicate the findings from Study 1a with a group of participants from another country and language. The same procedure was employed, except that participants in this study were Portuguese, and a new set of stimuli was selected from existing norms for European Portuguese.
Method
Participants
Using G*Power (Version 3.1.9.7), N was set as 76 participants (α = .05, power = .85) to achieve a small to medium effect size (dz = 0.35). A convenience sample of 163 university students participated in exchange for course credits or a prize draw. From those, 85 participants were excluded from the data analysis: 38 did not provide any PM response; 18 were nonnaïve as they took part in other PM studies from our lab; 14 participants stated not having paid attention, having cheated during the experiment, or had extremely long survey durations (>3.7 hours); 10 were non-European Portuguese native speakers or did not reveal their native language; two did not recognize one target and did not provide PM responses to those target trials; one gave no responses to more than 50% of the ongoing task trials; another was excluded due to a technical problem with the stimuli presentation; and another was underaged. Additional information is available in the Supplemental Materials.
The final sample was composed of 78 European Portuguese native speakers (mean age = 21.60 years, SD = 4.39; one participant was a Portuguese–English bilingual). Of those, 82.1% identified themselves as females, 16.7% as males, and 1.3% preferred not to reveal their gender. Each version of the task had 17 to 22 participants.
Material and procedure
The procedure was the same as in Study 1b, except that participants were asked to press S or N, for Yes [Sim] or No [Não] responses, respectively. Animate and inanimate European Portuguese words were selected from Félix et al. (2020) and matched along several variables (Table 1; selected words available as Supplemental Materials). Participants took, on average, 33 minutes to complete the task.
Results
Results are depicted in Fig. 3. The Animacy main effect was significant, F(1, 77) = 10.96, p < .001, ηp2 = .125, as was the Type of Trial main effect, F(1.28, 98.77) = 10.21, p < .001, ηp2 = .117. Furthermore, the Animacy × Type of Trial interaction reached significance, F(1.33, 102.50) = 6.29, p = .002, ηp2 = .076. Follow-up paired t tests revealed, again, a significant animacy advantage only on the target trials, t(77) = 2.93, p = .005, dz = 0.33. No animacy effect was obtained in the filler, t(77) = −1.11, p = .270, or on the baseline trials, t(77) = 2.15, p = .035 (a nonsignificant result considering the Bonferroni correction).

Mean performance obtained in baseline, filler (ongoing task) and target trials (PM Task), in Study 1b. Error bars represent standard errors of the mean
Study 2
Study 1b replicated the findings from Study 1a: The animacy effect was obtained on PM target trials. In Study 1b, a new set of stimuli was used, and the study was conducted with participants from another country and language, allowing the generalizability of the results and revealing that the effect is not language dependent. Looking for more evidence of the animacy effect in PM, Study 2 used a new ongoing task (a visuospatial task) as the main procedure.
Method
Participants
The sample size was calculated as in Study 1b. A total of 130 participants were recruited from Testable Verified Minds (https://www.testable.org/) using the following prescreeners: age (18–40 years old), first language (English), location (USA, UK, Ireland, New Zealand, Canada, or Australia). Following the preregistered exclusion criteria, 51 participants were excluded: 22 did not perform any PM response; 16 had a low performance in the filler trials (<Grand Mean – 3 SD); five participants were non-English native speakers or preferred not to reveal their native language; three participants did not provide any response to 50% or more trials; another three did not recognize one of the targets and did not perform any PM response to those target trials; and, another two failed both attention checks. Additional information is available as Supplemental Materials.
Seventy-nine participants were included in the data analysis (mean age = 30.33 years, SD = 6.52; one participant did not reveal his/her age; 48.1% were females and 51.9% males). Seventeen to 22 participants responded to each version of the task.
Material and procedure
We used the same stimuli and procedure as in Study 1a, except for the ongoing task: Participants now performed a visuospatial task. Also, the distractor task was an even/odd task. Finally, the experiment presented two attention checks: one right after the practice and the other after the baseline phases (“Have you ever walked on Mars?” and “Can you fly with invisible wings?”—yes/no responses), which served to exclude inattentive participants (i.e., those who responded “yes” to both questions; VanArsdall, 2016). On average, the experiment lasted 26 minutes.
In the ongoing task (Fig. 4), inspired by Costa et al. (2013), seven white squares were displayed on the screen. One at a time, six of them turned black, each one in a different location. Then, a word was presented in one of the seven possible square positions. The participants had to decide if the word’s location matched the location where a black square was displayed by pressing Y (yes) or N (no). For the PM phase, participants were instructed to press the SPACEBAR whenever any of the targets (an animate and an inanimate) was presented while performing the ongoing task.

Example of a match-trial, and representation of the presentation times of each stimulus (Study 2)
Results
Results are depicted in Fig. 5. The Animacy main effect did not reach conventional levels of significance, F(1, 78) = 3.51, p = .065, ηp2 = .043, but the main effect of Type of Trial was significant, F(1.32, 103.24) = 19.18, p < .001, ηp2 = .197. The Animacy × Type of Trial interaction was also significant, F(1.22, 95.03) = 11.83, p < .001, ηp2 = .132. This was due to a higher performance for inanimate (vs. the animate) words in the baseline trials, t(78) = −2.99, p = .004, dz = −0.34; and, more importantly, due to a significantly higher PM performance towards animate targets, as compared with the inanimate ones, t(78) = 3.05, p = .003, dz = 0.34. No animacy effect was obtained on the filler trials, t(78) = −0.70, p = .486.

Mean performance obtained in the baseline, filler (ongoing task) and target trials (PM Task), in Study 2. Error bars represent standard errors of the mean
General discussion
The proposal that memory should be tuned to remembering animates/living beings (as compared with inanimates/nonliving things) follows from the assumption that animates typically have a high fitness-relevant value (Nairne et al., 2017). Empirical evidence of the animacy advantage exists in retrospective memory but not in prospective memory. Combining two adaptive features of memory—its tuning toward animates and its future orientation—we predicted that the animacy effect would also occur in PM.
In a series of three studies using typical PM procedures, we reported, for the first time, that PM is also sensitive to the animacy dimension, at least in the type of tasks employed here (event-based tasks; cf. Einstein & McDaniel, 2005). Indeed, PM performance was consistently better in response to animate than to inanimate targets. Most participants had better performance for the animate targets (62% in Study 1a and in Study 2; and 55% in Study 1b), a smaller percentage had better PM performance for the inanimates (20%, 26% and 21% in Studies 1a, 1b and 2, respectively), and there were around 19% ties across studies (i.e., equal performance for animates and inanimates).Footnote 1 We should note that, in each study, we only used two different PM targets (one animate and one inanimate) to prevent a high cognitive load and, consequently, low levels of PM performance (Anderson et al., 2019), while maintaining the usual proportion of target/ongoing trials (Smith & Hunt, 2014). Still, we opted to use different sets of possible targets to increase the generalizability of our results. It is also noteworthy that the present results were obtained using different ongoing tasks and in two languages (Portuguese and English), which further reinforces the relevance and generalizability of the present findings.
Considering that knowledge about PM has been derived from retrospective memory theories, we consider how the two main accounts that have been proposed to explain the animacy effect in that context might explain the current results: the attention-based and the richness of encoding accounts. The richness of encoding account suggests that animates tend to be better recalled because they naturally lead to the generation of more ideas and/or have more features than inanimates (e.g., Meinhardt et al., 2020; Rawlinson & Kelley, 2021). Those ideas, features, or associates potentially work as retrieval cues and might improve performance (for animates) in free recall. When the existence of multiple cues is irrelevant to the task at hand, such as in cued recall, there is sometimes no animacy effect (e.g., Popp & Serra, 2016). In the case of PM, having multiple cues associated with the target could hinder the access to the PM intention memory trace (association: target–intention), thus impairing the PM performance for the animate targets (McDaniel & Einstein, 2000). In the same vein, a previous study has shown that when the PM target is paired with other words/associates in a study phase, PM performance decreases; also, the more associates are paired with the PM target, the lower the PM performance is (Cook et al., 2006). All together, these data, along with the present findings, suggest that the richness of encoding is unlikely to explain the animacy advantage reported here. As the main aim of this work was to explore, for the first time, a possible animacy advantage in PM, further studies using procedures designed specifically to disentangle the potential mechanisms are needed.
The attentional account posits that animates tend to be better recalled because they recruit attentional resources in a more automatic manner, thus requiring lower activation thresholds to be detected (e.g., Bugaiska et al., 2019). In our studies, the monitoring of the animate targets during the PM task might have benefited from this automatic-attention capture; that is, their detection would be facilitated as compared with the inanimate items, promoting more correct PM responses.
Following this latter account, one could also speculate about possible effects of animacy on the baseline and filler trials performance. In particular, the automatic attention captured by animates could impair performance in these trials as compared with the inanimate ones. Such a prediction was confirmed only in our Study 2, whereas no effect of animacy was observed in neither Study 1a or 1b. Moreover, the response time data has also been used as an indicator of the attentional mechanisms associated with animacy. For example, the response times in a color-naming Stroop task were longer when words refered to animates than to inanimates (e.g., Bugaiska et al., 2019). In our case, no effect of animacy was found on the response times of the baseline and filler trials (see Supplemental Materials). In sum, the predictions based on this account are not consistent with our results (see also Rawlinson & Kelley, 2021), revealing that the animacy effect in PM may not be explained solely by the attention-prioritization account. Other studies manipulating the characteristics of the target and the baseline/filler words, for example, in terms of emotionality, have found similar results: an enhancement of the PM performance for the emotional (target) words, as compared with the neutral ones, but no difference between them on the ongoing task (filler trials; May et al., 2012).
All in all, the present work reinforces the importance of animacy in memory functioning and adds PM to the list of processes that benefit from animacy. Additionally, not considering such variable might lead to disparate results. For example, emotionally valenced items are more likely to involve animates than neutral ones (e.g., May et al., 2012, 2015). Prospective memory research has also used materials that are ambiguous with respect to animacy (Félix et al., 2023; Lowder & Gordon, 2015). These include categories such as fruits, plants, body parts, and natural forces (e.g., Guynn, 2003; Moyes et al., 2019; Thomas & McBride, 2016). At this point, we cannot inform if and how this animacy category affects PM. Finally, we would encourage researchers to consider the variable of animacy when selecting their research materials, as is usually done for other variables (e.g., arousal, word frequency; May et al., 2012). Recent work has reported some differences on animacy ratings depending on the participants’ language and age; thus, specific language and age-group norms should be used (Félix et al., 2023).
Besides the theoretical relevance of the animacy effect in PM, one can speculate about the potential interest of these results to more applied contexts. Considering that PM is crucial to maintain a functional and independent life, one needs (and uses) PM ubiquitously. At the same time, most of our daily memory failures are PM-related (Cockburn, 1995), which can have severe consequences (e.g., Dembitzer & Lai, 2003). Thus, it is crucial to find the best tools to improve PM functioning. Our results suggest that using the naturally existing mnemonic tuning toward animates might be one such tool.
Data availability
The datasets generated during and/or analyzed during the current study are available in the OSF repository (https://osf.io/g6uqt/?view_only=None).
Code availability (software application or custom code)
Not applicable.
Notes
For these analyses, participants performing at 100% on the PM task were excluded, as no room existed for a possible effect to occur (59 participants in Study 1a, 20 in Study 1b, and 45 in Study 2).
References
Anderson, F. T., Strube, M. J., & McDaniel, M. A. (2019). Toward a better understanding of costs in prospective memory: A meta-analytic review. Psychological Bulletin, 145, 1053–1081. https://doi.org/10.1037/bul0000208
Bonin, P., Gelin, M., & Bugaiska, A. (2014). Animates are better remembered than inanimates: Further evidence from word and picture stimuli. Memory & Cognition, 42, 370–382. https://doi.org/10.3758/s13421-013-0368-8
Bradley, M. M., & Lang, P. J. (1999). Affective Norms for English Words (ANEW): Instruction manual and affective ratings. https://e-lub.net/media/anew.pdf
Brandimonte, M. A., & Ferrante, D. (2008). The social side of prospective memory. In M. Kliegel, M. A. McDaniel, & G. O. Einstein (Eds.), Prospective memory: Cognitive, neuroscience, developmental, and applied perspectives (pp. 347–366). Taylor & Francis Group.
Brewer, G. A., Knight, J. B., Marsh, R. L., & Unsworth, N. (2010). Individual differences in event-based prospective memory: Evidence for multiple processes supporting cue detection. Memory & Cognition, 38, 304–311. https://doi.org/10.3758/MC.38.3.304
Bugaiska, A., Grégoire, L., Camblats, A.-M., Gelin, M., Méot, A., & Bonin, P. (2019). Animacy and attentional processes: Evidence from the Stroop task. Quarterly Journal of Experimental Psychology, 72, 882–889. https://doi.org/10.1177/1747021818771514
Cameirão, M. L., & Vicente, S. G. (2010). Age-of-acquisition norms for a set of 1749 Portuguese words. Behavior Research Methods, 42, 474–480. https://doi.org/10.3758/BRM.42.2.474
Chen, X., Wang, Y., Liu, L., Shi, H., Wang, J., Cui, J., Shum, D. H. K., & Chan, R. C. K. (2014). The effect and mechanisms of implementation intentions on prospective memory in individuals with and without schizotypal personality features. Memory, 22, 349–359. https://doi.org/10.1080/09658211.2013.792841
Cockburn, J. (1995). Task interruption in prospective memory: A frontal lobe function? Cortex, 31, 87–97. https://doi.org/10.1016/S0010-9452(13)80107-4
Cook, G. I., Marsh, R. L., Hicks, J. L., & Martin, B. A. (2006). Fan effects in event-based prospective memory. Memory, 14, 890–900. https://doi.org/10.1080/09658210600816079
Cortese, M. J., & Khanna, M. M. (2008). Age of acquisition for 3,000 monosyllabic words. Behavior Research Methods, 40, 791–794. https://doi.org/10.3758/BRM.40.3.791
Cosmides, L., & Tooby, J. (1992). Cognitive adaptations for social exchange. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 163–228). Oxford University Press.
Costa, A., Oliveri, M., Barban, F., Bonnì, S., Koch, G., Caltagirone, C., & Carlesimo, G. A. (2013). The right frontopolar cortex is involved in visual-spatial prospective memory. PLOS ONE, 8(2), Article e56039. https://doi.org/10.1371/journal.pone.0056039
Daley, M. J., Andrews, G., & Murphy, K. (2020). Animacy effects extend to working memory: Results from serial order recall tasks. Memory, 28, 157–171. https://doi.org/10.1080/09658211.2019.1699574
Dembitzer, A., & Lai, E. J. (2003). Retained surgical instrument. New England Journal of Medicine, 348(3), 228. https://doi.org/10.1056/NEJMicm020710
Dewhurst, S. A., & Parry, L. A. (2000). Emotionality, distinctiveness, and recollective experience. European Journal of Cognitive Psychology, 12, 541–551. https://doi.org/10.1080/095414400750050222
DeYoung, C. M., & Serra, M. J. (2021). Judgments of learning reflect the animacy advantage for memory, but not beliefs about the effect. Metacognition and Learning. https://doi.org/10.1007/s11409-021-09264-w
Einstein, G. O., & McDaniel, M. A. (1990). Normal aging and prospective memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 717–726. https://doi.org/10.1037/0278-7393.16.4.717
Einstein, G. O., & McDaniel, M. A. (2005). Prospective memory: Multiple retrieval processes. Current Directions in Psychological Science, 14, 286–290. https://doi.org/10.1111/j.0963-7214.2005.00382.x
Einstein, G. O., McDaniel, M. A., Thomas, R., Mayfield, S., Shank, H., Morrisette, N., & Breneiser, J. (2005). Multiple processes in prospective memory retrieval: Factors determining monitoring versus spontaneous retrieval. Journal of Experimental Psychology: General, 134, 327–342. https://doi.org/10.1037/0096-3445.134.3.327
Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007). G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39, 175–191. https://doi.org/10.3758/BF03193146
Félix, S. B., Pandeirada, J. N. S., & Nairne, J. S. (2020). Animacy norms for 224 European Portuguese concrete words. Análise Psicológica, 38, 257–269. https://doi.org/10.14417/ap.1690
Félix, S. B., Poirier, M., & Pandeirada, J. N. S. (2023). Is “earth” an animate thing? Cross-language and inter-age analyses of animacy word ratings in European Portuguese and British English young and older adults. PLOS ONE, 18, Article e0289755. https://doi.org/10.1371/journal.pone.0289755
Ganis, G., & Kievit, R. (2015). A new set of three-dimensional shapes for investigating mental rotation processes: Validation data and stimulus set. Open Journal of Psychology Data, 3, Article e3. https://doi.org/10.5334/jopd.ai
Guynn, M. J. (2003). A two-process model of strategic monitoring in event-based prospective memory: Activation/retrieval mode and checking. International Journal of Psychology, 38, 245–256. https://doi.org/10.1080/00207590344000178
Hostler, T. J., Wood, C., & Armitage, C. J. (2018). The influence of emotional cues on prospective memory: A systematic review with meta-analyses. Cognition and Emotion, 32, 1578–1596. https://doi.org/10.1080/02699931.2017.1423280
IBM Corp. (2021). IBM SPSS Statistics for Windows, Version 28.0. Armonk, NY: IBM Corp.
Ingvar, D. H. (1985). “Memory of the future”: An essay on the temporal organization of conscious awareness. Human Neurobiology, 4(3), 127–136.
Klein, S. B. (2013). The temporal orientation of memory: It’s time for a change of direction. Journal of Applied Research in Memory and Cognition, 2, 222–234. https://doi.org/10.1016/j.jarmac.2013.08.001
Kučera, H., & Francis, W. N. (1967). Computational analysis of present-day American English. Providence, RI: Brown University Press.
Kvavilashvili, L., & Ellis, J. (1996). Varieties of intention: Some distinctions and classifications. In M. A. Brandimonte, G. O. Einstein, & M. A. McDaniel (Eds.), Prospective memory: Theory and appliations (pp. 23–51). Psychology Press.
Laurino, J., & Kaczer, L. (2019). Animacy as a memory enhancer during novel word learning: Evidence from orthographic and semantic memory tasks. Memory, 27, 820–828. https://doi.org/10.1080/09658211.2019.1572195
Leding, J. K. (2020). Animacy and threat in recognition memory. Memory & Cognition, 48, 788–799. https://doi.org/10.3758/s13421-020-01017-5
Leitão, J., Figueira, A., & Almeida, A. (2010). Normas de imaginabilidade, familiaridade e idade de aquisição para 252 nomes comuns [Imaginability, familiarity, and age of acquisition norms for 252 common nouns]. Laboratório de Psicologia, 8, 101–119. https://doi.org/10.14417/lp.651
Lowder, M. W., & Gordon, P. C. (2015). Natural forces as agents: Reconceptualizing the animate–inanimate distinction. Cognition, 136, 85–90. https://doi.org/10.1016/j.cognition.2014.11.021
Marsh, R. L., Brewer, G. A., Jameson, J. P., Cook, G. I., Amir, N., & Hicks, J. L. (2009). Treat-related processing supports prospective memory retrieval for people with obsessive tendencies. Memory, 17, 679–686. https://doi.org/10.1080/09658210903032762
May, C. P., Manning, M., Einstein, G. O., Becker, L., & Owens, M. (2015). The best of both worlds: Emotional cues improve prospective memory execution and reduce repetition errors. Aging, Neuropsychology, and Cognition, 22, 357–375. https://doi.org/10.1080/13825585.2014.952263
May, C., Owens, M., & Einstein, G. O. (2012). The impact of emotion on prospective memory and monitoring: No pain, big gain. Psychonomic Bulletin and Review, 19, 1165–1171. https://doi.org/10.3758/s13423-012-0301-3
McDaniel, M. A., & Einstein, G. O. (2000). Strategic and automatic processes in prospective memory retrieval: A multiprocess framework. Applied Cognitive Psychology, 14, S127–S144. https://doi.org/10.1002/acp.775
Meinhardt, M. J., Bell, R., Buchner, A., & Röer, J. P. (2020). Adaptive memory: Is the animacy effect on memory due to richness of encoding? Journal of Experimental Psychology: Learning, Memory, and Cognition, 46, 416–426. https://doi.org/10.1037/xlm0000733
Moyes, J., Sari-Sarraf, N., & Gilbert, S. J. (2019). Characterising monitoring processes in event-based prospective memory: Evidence from pupillometry. Cognition, 184, 83–95. https://doi.org/10.1016/j.cognition.2018.12.007
Murphy, D. H., & Castel, A. D. (2022). Selective remembering and directed forgetting are influenced by similar stimulus properties. Memory, 30, 1130–1147. https://doi.org/10.1080/09658211.2022.2092152
Nairne, J. S., & Pandeirada, J. N. S. (2008). Remembering with a stone-age brain. Current Directions in Psychological Science, 17, 239–243. https://doi.org/10.1111/j.1467-8721.2008.00582.x
Nairne, J. S., VanArsdall, J. E., & Cogdill, M. (2017). Remembering the living: Episodic memory is tuned to animacy. Current Directions in Psychological Science, 26, 22–27. https://doi.org/10.1177/0963721416667711
Nairne, J. S., VanArsdall, J. E., Pandeirada, J. N. S., Cogdill, M., & LeBreton, J. (2013). Adaptive memory: The mnemonic value of animacy. Psychological Science, 24, 2099–2105. https://doi.org/10.1177/0956797613480803
Popp, E. Y., & Serra, M. J. (2016). Adaptive memory: Animacy enhances free recall but impairs cued recall. Journal of Experimental Psychology: Learning, Memory, and Cognition, 42, 186–201. https://doi.org/10.1037/xlm0000174
Rawlinson, H. C., & Kelley, C. M. (2021). In search of the proximal cause of the animacy effect on memory: Attentional resource allocation and semantic representations. Memory & Cognition, 49, 1137–1152. https://doi.org/10.3758/s13421-021-01154-5
Rouse, S. V. (2015). A reliability analysis of Mechanical Turk data. Computers in Human Behavior, 43, 304–307. https://doi.org/10.1016/j.chb.2014.11.004
Schacter, D. L., Addis, D. R., & Buckner, R. L. (2007). Remembering the past to imagine the future: The prospective brain. Nature Reviews Neuroscience, 8, 657–661. https://doi.org/10.1038/nrn2213
Schaper, M. L., Horn, S. S., Bayen, U. J., Buchner, A., & Bell, R. (2022). Adaptive prospective memory for faces of cheaters and cooperators. Journal of Experimental Psychology: General, 151, 1358–1376. https://doi.org/10.1037/xge0001128
Schnitzspahn, K. M., Zeintl, M., Jäger, T., & Kliegel, M. (2011). Metacognition in prospective memory: Are performance predictions accurate? Canadian Journal of Experimental Psychology, 65(1), 19–26. https://doi.org/10.1037/a0022842
Schock, J., Cortese, M. J., Khanna, M. M., & Toppi, S. (2012). Age of acquisition estimates for 3,000 disyllabic words. Behavior Research Methods, 44, 971–977. https://doi.org/10.3758/s13428-012-0209-x
Smith, R. E. (2003). The cost of remembering to remember in event-based prospective memory: Investigating the capacity demands of delayed intention performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29, 347–361. https://doi.org/10.1037/0278-7393.29.3.347
Smith, R. E., & Hunt, R. R. (2014). Prospective memory in young and older adults: The effects of task importance and ongoing task load. Aging, Neuropsychology, and Cognition, 21(4), 411–431. https://doi.org/10.1080/13825585.2013.827150
Soares, A. P., Comesaña, M., Pinheiro, A., Simões, A., & Frade, C. (2012). The adaptation of the Affective Norms for English words (ANEW) for European Portuguese. Behavior Research Methods, 44, 256–269. https://doi.org/10.3758/s13428-011-0131-7
Soares, A. P., Costa, A. S., Machado, J., Comesaña, M., & Oliveira, H. (2017). The Minho word pool: Norms for imageability, concreteness and subjective frequency for 3800 Portuguese words. Behavior Research Methods, 49, 1065–1081. https://doi.org/10.3758/s13428-016-0767-4
Thomas, B. J., & McBride, D. M. (2016). The effect of semantic context on prospective memory performance. Memory, 24, 315–323. https://doi.org/10.1080/09658211.2015.1004351
VanArsdall, J. E. (2016). Exploring animacy as a mnemonic dimension (Doctoral dissertation,. Purdue University. https://docs.lib.purdue.edu/open_access_dissertations/873/
VanArsdall, J. E., & Blunt, J. R. (2022). Analyzing the structure of animacy: Exploring relationships among six new animacy and 15 existing normative dimensions for 1,200 concrete nouns. Memory & Cognition, 50, 997–1012. https://doi.org/10.3758/s13421-021-01266-y
VanArsdall, J. E., Nairne, J. S., Pandeirada, J. N. S., & Cogdill, M. (2015). Adaptive memory: Animacy effects persist in paired-associate learning. Memory, 23, 657–663. https://doi.org/10.1080/09658211.2014.916304
Wilson, M. (1988). MRC Psycholinguistic Database: Machine-usable dictionary, version 2.00. Behavior Research Methods, Instruments, and Computers, 20, 6–10. https://doi.org/10.3758/BF03202594
Acknowledgements
The authors thank Michelle E. Coverdale (Purdue University) and Andronikos Botsas (City, University of London) for their help in programing and reviewing some of the tasks. We also thank Lara Palmeira (Universidade Portucalense) for providing access to some of the sample for Study 1b.
Open practices statement
The data for all studies are publicly accessible via OSF (https://osf.io/g6uqt/?view_only=None). The materials used in all studies are available as Supplemental Materials. At the time Studies 1a and 1b were conducted, preregistration was not yet a common practice. Study 2 was preregistered before starting data collection (available at: https://aspredicted.org/3gq9v.pdf).
Funding
Open access funding provided by FCT|FCCN (b-on). Preparation of this work was supported by the Portuguese Foundation for Science and Technology through a doctoral fellowship to SBF (SFRH/BD/145097/2019), a grant to JNSP (CEECIND/01914/2017), and multiannual funding to the William James Center for Research (UIDB/04810/2020). The City, University of London funded Study 2.
Author information
Authors and Affiliations
Contributions
All authors contributed to the design of the studies, the selection of the materials, and the discussion of the results. SBF programmed the experiments, and collected and analyzed the data with the contribution of JNSP. SBF wrote the first draft of the manuscript. All authors revised, edited, and approved the final version of the manuscript for submission.
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
The authors have no competing interests to declare.
Ethics approval
All studies were conducted in accordance with the ethical standards as laid down in the 1964 Declaration of Helsinki and its later amendments. Study 1a was approved by the Purdue IRB (Ref: 1301013109). Study 1b was approved by the Ethics and Deontology Committee of the University of Aveiro (Ref: 34/2019). Study 2 was approved by the City, University of London Psychology Research Ethics Committee (Ref: ETH2122-0271).
Consent to participate
All individual participants provided their informed written consent prior to take part in the study.
Consent for publication
All individual participants provided their written informed consent regarding publishing their anonymized data.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(PDF 298 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Félix, S.B., Poirier, M., Nairne, J.S. et al. The breadth of animacy in memory: New evidence from prospective memory. Psychon Bull Rev 31, 1323–1334 (2024). https://doi.org/10.3758/s13423-023-02406-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-023-02406-y