
Abstract
From their finding that the substantial magnitude of the Stroop interference that occurs when a participant’s initial fixation is directed at the optimal viewing position is eliminated when the initial fixation is directed at the end of a word, Perret and Ducrot (2010) concluded that initial fixation at the latter position likely prevents reading. In the present study, we further examined this interpretation. To this end, the two conflict dimensions (semantic vs. response) that were confounded in the original work were separated within a semantically based Stroop paradigm (Neely & Kahan, 2001) that was administered with vocal (instead of manual) responses. In line with past findings showing greater interference in the vocal task, the reported results indicated that standard Stroop interference was reduced, but not eliminated, thus making the initial interpretation in terms of reading suppression unlikely. This conclusion is further strengthened by the presence of isolated semantic interference, the magnitude of which remained significant and was unaffected by viewing position. In sum, these results show that initial fixation of the end of a word simply reduces (nonsemantic) response competition.
Similar content being viewed by others
Research investigating eye movements in reading has identified that the optimal viewing position (hereafter, OVP)—located near the word center—facilitates word recognition. Indeed, both naming and lexical decision latencies and gaze durations are shorter when the eyes initially fixate the OVP rather than other viewing positions, such as the beginning or the end of the word (see, e.g., O’Regan & Jacobs, 1992; O’Regan, Lévy-Schoen, Pynte, & Brugaillère, 1984). Because there is a rapid fall in visual acuity with retinal eccentricity, such effects mainly arise from the fact that more letters can be extracted from a word when the eyes are near the word’s center (Nazir, Jacobs, & O’Regan, 1998; see also, e.g., Brysbaert & Nazir, 2005; Yao-N’Dré, Castet, & Vitu, 2013, for further explanations of this effect). In this article, we examine the influence of viewing positions on Stroop interference.
The Stroop (1935) task requires participants to identify the color in which a target word is printed as quickly and accurately as possible. Participants’ identification times are longer when the word designates a color different from the color in which it is printed (e.g., the word BLUE displayed in green) than when a color-neutral trial is presented (e.g., DEAL / #### displayed in green). This type of interference is often considered to be fundamental to the claim that word recognition is automatic. In this view, since they are skilled readers, participants cannot refrain from reading Stroop words and processing their meanings (i.e., computing their lexical and semantic representations; see, e.g., Brown, Gore, & Carr, 2002; Lachter, Ruthruff, Lien, & McCann, 2008).Footnote 1
The recent use of an eyetracking device in the color-naming Stroop task has shown that, although people do not read Stroop words intentionally, their first eye fixations are still systematically biased toward the OVP (Smilek, Solman, Murawski, & Carriere, 2009). The latter finding, combined with those outlined above, led Perret and Ducrot (2010) to predict that the substantial Stroop interference observed when participants’ eyes initially fixate the OVP should be reduced when the initial fixation is on other viewing positions.
In order to test this prediction, Perret and Ducrot (2010) asked their participants to identify the colors of incongruent (e.g., BLUE green), congruent (e.g., BLUE blue), or neutral (e.g., #####green) displays while the initial fixation position was experimentally controlled (at the OVP vs. the end of the display). The results showed that the substantial Stroop interference (e.g., BLUE green – ####green) observed at the OVP was statistically eliminated when both adult (Exp. 1) and fifth grader (Exp. 2) participants first fixated the end-letter position. The stability of this result led Perret and Ducrot to conclude that “low-level oculomotor processes mediate performance on the Stroop task” (p. 553), such that “fixating to the right of the word’s center is likely to reduce the overall letter visibility and prevent participants from reading the word” (p. 554, italics added).
However, a closer inspection of the critical end-letter position shows that at least some interference still occurred (i.e., 16 ms in both experiments), even though it fell below the conventional level of significance (i.e., p = .10 in Exp. 1 and p = .16 in Exp. 2). These results tend to contradict the idea that this viewing position prevents participants from reading Stroop words, and suggests that the influence of position on Stroop interference might be more complex than was implied above. Consequently, the goal of this article was to further examine this issue.
This complex influence is due in part to the complexity of Stroop interference itself. It should be remembered that functional analyses of the Stroop task have suggested that in standard incongruent trials (e.g., BLUE green), the target (i.e., green) and the distractor (i.e., blue) mismatch at both the stimulus and response levels (see, e.g., De Houwer, 2003). Thus, contrary to (classic) approaches attempting to explain Stroop interference in terms of a single locus (see, e.g., MacLeod, 1991, for a review), such analyses necessarily imply that the interference has at least two possible loci (i.e., stimulus and response level).
Applying this general logic, we previously argued that the interference occurring at the stimulus level is due to semantic competition (see also, e.g., Klein, 1964; Luo, 1999; Neely & Kahan, 2001). In this view, Stroop interference appears because processing of the color dimension interferes with (automatic) processing of the word dimension, with the result that the semantic information computed from the target (e.g., green) conflicts with the semantic information automatically computed from the distractor (e.g., BLUE). Precisely because participants cannot refrain from processing the distractor (see above),Footnote 2 we have also argued, and repeatedly demonstrated, that this component of Stroop interference is automatic, in the senses that it (a) is independent of attentional resources (Augustinova & Ferrand, 2007; Augustinova, Flaudias, & Ferrand, 2010; see also Augustinova & Ferrand, 2012a) and (b) cannot be controlled or prevented (see, e.g., Augustinova & Ferrand, 2012a, 2012b, 2013).Footnote 3
We have also argued that an additional component of Stroop interference arises at the response level, because both the target and the distractor prime motor responses that compete and dominate overt response activity (see, e.g., De Houwer, 2003; Neely & Kahan, 2001; Schmidt & Cheesman, 2005; van Veen & Carter, 2005). Indeed, because both correct (primed by the target) and incorrect (primed by the distractor) motor responses share the same response set, in a manual task for instance (i.e., the one used by Perret & Ducrot, 2010), considerable competition takes place about which key to press, since blue is assigned to one key and green to another. Even though this contribution to overall Stroop interference is far from being fully understood, our own work mentioned above has also unambiguously shown that this component is extremely flexible and can be eliminated (unlike the aforementioned semantic component).
This elimination might give the impression that standard Stroop interference is eliminated in a manual task, leading researchers to the (erroneous) conclusion that word reading is suppressed (see above). Indeed, manual versions of the Stroop task are known to produce smaller interference than do Stroop tasks with vocal responses (see, e.g., MacLeod, 1991; Sharma & McKenna, 1998). This is due, at least in part, to the fact that in manual-response variants, the contribution of semantic interference is small (see, e.g., Augustinova & Ferrand, 2013; Sharma & McKenna, 1998; Schmidt & Cheesman, 2005) and may therefore not be detected by traditional chronometric measures (see, e.g., Sharma & McKenna, 1998). Thus, the immediate solution would be to examine whether any elimination demonstrated in a manual task persists with vocal responses (see Brown, Joneleit, Robinson, & Brown, 2002, who initiated this idea), a case in which semantic interference has been consistently detected (see, e.g., Augustinova & Ferrand, 2013).
In light of this idea, the first specific goal of this article was to replicate the original study of Perret and Ducrot (2010) and extend this work to the vocal response modality. We expected that in this particular response modality, the initial fixation of the end-letter position (as opposed to the OVP) would simply reduce (but not eliminate) standard Stroop interference. Given this a priori prediction, the next goal was to examine the level at which this reduction would occur.
Because the two components of the overall interference mentioned above (i.e., the contributions resulting from semantic conflict and response conflict) are confounded within the standard Stroop task, as described above, the study reported below was conducted using a semantic variant of the Stroop paradigm (see, e.g., Neely & Kahan, 2001; see also, e.g., Klein, 1964). The main advantage of the so-called semantically based Stroop paradigm is that it isolates the semantic conflict from the response conflict without substantially modifying the original Stroop paradigm. Indeed, it simply supplements standard incongruent trials (e.g., BLUE green) with trials on which the words are associated with an incongruent color (e.g., SKY green). Thus, it allows for the simultaneous computation of both standard (e.g., BLUE green – DEAL / ####green) and semantic (e.g., SKY green – DEAL / ####green) Stroop interference.
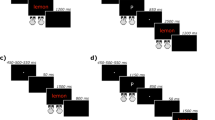
As can be seen in Fig. 1, semantic Stroop interference, just like standard Stroop interference, implies a semantic conflict because the target (e.g., green) and the distractor (e.g., SKY, which is blue) still overlap semantically. However, unlike standard Stroop interference, it is free of response conflict, because the response sets for the target (e.g., green) and the distractor (e.g., SKY) are distinct. Indeed, the color associates do not produce the response linked to the associated color (i.e., blue, associated with SKY; see Schmidt & Cheesman, 2005, for a straightforward demonstration).

Comparison of standard and semantic Stroop interference effects, using data from (Augustinova et al. 2010, Exp. 1; a naming task). SI = Stroop interference. In this example, DEAL, SKY, and BLUE are displayed in green color
In line with our view that this (i.e., semantic) contribution to overall Stroop interference is automatic, we expected semantic Stroop interference to remain significant and to be of the same magnitude, irrespective of whether participants’ initial fixations were directed at the end-letter position or at the OVP, and further expected only standard Stroop interference to be reduced by an initial fixation directed at the end-letter position. Within the “subtractive” logic of the semantic Stroop paradigm (see Fig. 1), and in line with our past work, this result would mean that variations in viewing positions solely influence response conflict.
Method
Participants and design
A group of 34 psychology undergraduates (M age = 21.05 years; min = 18, max = 27 years) at Blaise Pascal University, Clermont-Ferrand, France, took part in this experiment in exchange for course credit. All were native French speakers, had normal or corrected-to-normal vision, and were not color-blind. They were assigned to all conditions in a 2 (initial fixation position: P3 [optimal viewing position] vs. P5 [end letter]) × 4 (type of stimulus: standard incongruent vs. color-associated incongruent vs. standard congruent vs. neutral) within-subjects design.
Stimuli, apparatus, and procedure
The study was identical to Perret and Ducrot (2010), with two exceptions: The response mode was verbal instead of manual, and the original set of stimuli—consisting of four incongruent French color words (bleu, vert, rouge, and jaune; blue, green, red, and yellow, respectively), four congruent color words (bleu, vert, rouge, and jaune), and four neutral signs (“#####” of different lengths)—was supplemented by four color-associated incongruent words (ciel [sky], salade [salad], tomate [tomato], and maïs [corn]).
Participants were tested individually. The stimuli were presented on a 17-in. Dell color monitor connected to a PC running DMDX (Forster & Forster, 2003). The stimuli were displayed in colored uppercase letters on a black background in 24-point Courier New font, with a 640 × 480 resolution. The participants were seated on a fixed chair 60 cm from the screen. At this distance, each letter subtended 1º of visual angle. Each trial consisted of the following sequence of events (see Fig. 1 in Perret & Ducrot, 2010, p. 552). At the beginning of a trial, participants had to fixate a cross (“+”) displayed in the middle of the screen and not to move their eyes. The importance of continuing to focus on this point was stressed repeatedly. After 500 ms, the fixation point was replaced by a stimulus that remained on the screen until the participant responded. The stimulus was displayed off-center—that is, shifted to the side with respect to the fixation point, depending on the position condition. In the P3 position, stimuli were presented in such a way that the participants initially fixated the center of the OVP zone, which was located slightly to the left of center. In the P5 condition, stimuli were presented with their last letters on the central fixation point. To perform this manipulation, each stimulus was divided into five equally wide zones (i.e., 0.8 letters wide for four-letter words, 1.0 letters wide for five-letter words, and 1.2 letters wide for six-letter words).
The participant’s task was to name the color in which each item was printed, as quickly and accurately as possible. Their vocal responses were recorded via a Koss 70-dB microphone headset, and response latencies were measured to the nearest millisecond. Once the participant had responded, the screen was cleared, and a new trial began after a 1,000-ms delay. The order of trials was randomized. A 12-item training phase was held at the beginning of the experiment, followed by a single block of 96 trials.
Results
Latencies more than three SDs above or below each participant’s mean latency for each condition (accounting for less than 2 % of the total data in this experiment) were excluded from the analyses.
All Stroop-like findings, except for the congruent facilitation effect, were observed (see Table 1 for descriptive statistics).Footnote 4 Thus, the computed magnitudes of both standard (standard color-incongruent trials – color-neutral trials, all ps < .001; see Table 1) and semantic Stroop interference (color-associated incongruent trials – color-neutral trials, all ps < .001; see Table 1) were analyzed in a 2 (type of Stroop interference: standard vs. semantic) × 2 (initial fixation position: P3 [OVP] vs. P5 [end letter]) repeated measures analysis of variance to examine the level at which the initial fixation position affects Stroop interference.
This analysis revealed that the main effects of type of Stroop interference, F(1, 33) = 37.80, p < .001, η p 2 = .53, and of initial fixation position, F(1, 33) = 12.32, p < .001, η p 2 = .27, as well as the Type of Stroop Interference × Initial Fixation Position interaction, F(1, 33) = 29.09, p < .001, η p 2 = .46, were all significant. The decomposition of the latter interaction showed that standard Stroop interference was significantly reduced (but not eliminated) at the end-fixation position (P5), as compared to the OVP position (P3) [109 vs. 170 ms], F(1, 33) = 44.63, p < .001, η p 2 = .57. However, the viewing positions had no effect on the semantic Stroop interference, which remained of the same magnitude whether the initial fixation was directed at the end position (P5) or the OVP (P3) [87 vs. 88 ms], F(1, 33) < 1, p = .93, n.s.
These results are consistent with our initial reasoning that initial fixation at the end-letter position (P5) might substantially reduce response conflict. This conclusion was strengthened when we decomposed the above-mentioned interaction in order to compare the two types of Stroop interference in this condition. The 22-ms difference between the standard and semantic types of Stroop interference (109 vs. 87 ms) was only marginally significant, F(1, 33) = 3.85, p = .058, η p 2 = .11, suggesting that the response conflict was almost eliminated (see Fig. 1) in this condition.
Discussion
As was suggested by previous work (see, e.g., MacLeod, 1991; Sharma & McKenna, 1998), the standard Stroop interference effects observed in our vocal response modality were larger than those observed in the manual response modality used in Perret and Ducrot’s (2010) original study. In all other respects, these effects followed the same general pattern reported in the original study.
As these greater interferences suggest, and also as expected, the substantial standard Stroop interference (standard incongruent – color-neutral stimuli) observed when the first fixation was located at the OVP was only significantly reduced (but not eliminated) when the first fixation was located at the end of the word. Therefore, contrary to Perret and Ducrot’s (2010) claim that “fixating to the right of the word’s center is likely to reduce the overall letter visibility and prevent participants from reading the word” (2010, p. 554; see also Smilek et al. 2009), our findings clearly show that this is not the case. Indeed, one can hardly argue that substantial standard Stroop interference (i.e., 109 ms) can appear without word reading.
This conclusion is supported directly by the results relating to the semantic contribution to overall interference, since the semantic Stroop interference (incongruent color-associated – color-neutral stimuli) remained significant and of exactly the same magnitude in both viewing positions (see also Augustinova et al., 2010, for a similar result). This result is consistent with the findings of Jordan, McGowan, and Paterson (2012) showing that, despite the importance attributed to high-quality foveal input by theories of reading (e.g., O’Regan et al., 1984), word reading can function successfully with restricted foveal inputs. In a similar vein, our results show that variations in overall letter visibility, as reflected by different viewing positions, have no effect on semantic processing.
Moreover, the latter result supports our initial predictions that low-level oculomotor processes would influence response conflict and that this would be substantially reduced (and almost completely eliminated) by initial fixation at the end-letter position. The direct comparison between the original study of Perret and Ducrot (2010), conducted with a manual Stroop variant, and our own does indeed show that the near elimination of response conflict directly demonstrated in our study was the main mechanism responsible for the statistical elimination of standard Stroop interference in Perret and Ducrot’s results. Consequently, the most obvious limitation of the present study is that we do not have any immediate answers to the question of how this influence operates.
Even though response conflict is usually thought to occur late in processing (see, e.g., van Veen & Carter, 2005), Chen, Lei, Ding, Li, and Chen (2013) recently suggested that the reduction of response conflict (rather than of semantic conflict) requires an increased allocation of cognitive resources not only during the late stage of processing, but also during early stimulus processing. Their fMRI data are compatible with our recent event-related potential investigation of the influence of a single-letter coloring/spatial-cuing manipulation that reduces response (but not semantic) conflict in the Stroop task (see, e.g., Flaudias, Silvert, Augustinova, Llorca, & Ferrand, 2013). Although, as predicted, this manipulation had no effect on the N400 amplitude (which is an indicator of semantic processing), it elicited a larger N100 amplitude, thus suggesting that the reduction of the response conflict is contingent on the allocation of adequate attentional resources at an early stage of processing. Thus, further investigation of the effects of viewing positions using this technique, combined with delta-plot analyses (i.e., a time course indicator of inhibition; see, e.g., Sharma, Booth, Brown, & Huguet, 2010), should provide interesting insights into whether low-level oculomotor processes influence response conflict at an early (vs. a later) stage of processing and whether this reduction is due to an improved allocation of cognitive resources (possibly to the target) and/or to more efficient inhibition of the distractor. Moreover, such combinations of indicators would also help us better understand response conflict in general.
Another limitation relates to the sampling procedure used here to define the OVP. To perform the replication, we divided each stimulus into five equally wide zones and sampled Zones 3 and 5, as in Perret and Ducrot (2010). But is Position 3 really the OVP, or actually the center of the word? For example, Parris, Sharma, and Weekes (2007; see also Augustinova et al., 2010) approximated the OVP at the second position (P2) in five-letter words. More importantly, they also sampled the middle letter position (P3) and did not find an interference effect as large as the one observed at P2, approximating the OVP (but see Augustinova et al., 2010). It is therefore possible that the reduction in Stroop interference would be greater if only the actual OVP position were sampled. Since, somewhat contrary to this reasoning, O’Regan and Jacobs (1992, Exp. 2a) reported that the OVP in a word-naming task was at P3 (and not at P2) for high-frequency four-, five- and seven-letter words (i.e., the kinds of words used in the present study), future research should compare these different zones (P2, P3, and P5) more systematically.
Again, even though the stimuli that we used were identical to those of Perret and Ducrot (2010; see also, e.g., Smilek et al., 2009), they were larger than those used in some other studies investigating the OVP in the Stroop task (see, e.g., Augustinova et al., 2010; Parris et al., 2007). Thus, our finding is potentially limited to visually larger Stroop words (as compared to their smaller counterparts).Footnote 5 However, the fact that we found Stroop interference effects of similar magnitude to those obtained in vocal Stroop studies using smaller Stroop words (see, e.g., Augustinova & Ferrand, 2012a) suggests otherwise. In sum, future investigations should address the issue of whether the size of Stroop words directly influences participants’ processing of these words.
Despite these different limitations, the data presented above clearly strengthen our general claim that semantic processing in the Stroop task is indeed automatic (see, e.g., Augustinova & Ferrand, 2007, 2012a, 2012b, 2013; Augustinova et al., 2010; Brown et al., 2002)—a claim that clearly contrasts with some previous findings (see, e.g., Besner, 2001).
Notes
On the basis of the view that visual word recognition is automatic, many studies have supported the idea that visual word recognition does not require spatial attention (e.g., Brown, Gore, Carr 2002 and Brown, Joneleit, Robinson et al. 2002; Lachter et al., 2008; Neely & Kahan, 2001; but see, e.g., Lachter et al., 2004; Waechter, Besner, & Stolz, 2011, for a contrasting view).
The underlying idea here is that lexical–semantic processing typically reflects the default set that participants adopt. Note, however, that this account has been challenged by results suggesting that the activation of lexical–semantic representations can be temporarily controlled (see, e.g., Besner, 2001; Risko, Stolz, & Besner, 2005).
For the sake of brevity, we do not report the overall analyses on either response times or percentages of errors. However, the data showed no signs of a speed–accuracy trade-off.
We are grateful to one of the reviewers, Benjamin Parris, for raising this issue and the one that has just been discussed.
References
Augustinova, M., & Ferrand, L. (2007). Influence de la présentation bicolore des mots sur l’effet Stroop [First-letter coloring and the Stroop effect]. L’Année Psychologique, 107, 163–179.
Augustinova, M., & Ferrand, L. (2012a). The influence of mere social presence on Stroop interference: New evidence from the semantically-based Stroop task. Journal of Experimental Social Psychology, 48, 1213–1216.
Augustinova, M., & Ferrand, L. (2012b). Suggestion does not de-automatize word reading: Evidence from the semantically based Stroop task. Psychonomic Bulletin & Review, 19, 521–527. doi:10.3758/s13423-012-0217-y
Augustinova, M., & Ferrand, L. (2013). Social priming of dyslexia and reduction of the Stroop effect: What component of the Stroop effect is actually reduced? Revised manuscript under review.
Augustinova, M., Flaudias, V., & Ferrand, L. (2010). Single-letter coloring and spatial cuing do not eliminate or reduce a semantic contribution to the Stroop effect. Psychonomic Bulletin & Review, 17, 827–833. doi:10.3758/PBR.17.6.827
Besner, D. (2001). The myth of ballistic processing: Evidence from Stroop’s paradigm. Psychonomic Bulletin & Review, 8, 324–330. doi:10.3758/BF03196168
Brown, T. L., Gore, C. L., & Carr, T. H. (2002a). Visual attention and word recognition in Stroop color naming: Is word recognition “automatic”? Journal of Experimental Psychology: General, 131, 220–240. doi:10.1037/0096-3445.131.2.220
Brown, T. L., Joneleit, K., Robinson, C. S., & Brown, C. R. (2002b). Automaticity in reading and the Stroop task: Testing the limits of involuntary word processing. American Journal of Psychology, 115, 515–543.
Brysbaert, M., & Nazir, T. (2005). Visual constraints in written word recognition: Evidence from the optimal viewing position. Journal of Research in Reading, 28, 216–228.
Chen, Z., Lei, X., Ding, C., Li, H., & Chen, A. (2013). The neural mechanisms of semantic and response conflicts: An fMRI study of practice-related effects in the Stroop task. NeuroImage, 66, 577–584.
Cohen, J. (1988). Statistical power analysis for behavioral science (2nd ed.). Hillsdale, NJ: Erlbaum.
De Houwer, J. (2003). On the role of stimulus–response and stimulus–stimulus compatibility in the Stroop effect. Memory & Cognition, 31, 353–359. doi:10.3758/BF03194393
Flaudias, V., Silvert, L., Augustinova, M., Llorca, P.-M., & Ferrand, L. (2013). An electrophysiological investigation of single-letter coloring and spatial cuing in the Stroop task. Submitted manuscript.
Forster, K. I., & Forster, J. C. (2003). DMDX: A Windows display program with millisecond accuracy. Behavior Research Methods, Instruments, & Computers, 35, 116–124. doi:10.3758/BF03195503
Jordan, T. R., McGowan, V. A., & Paterson, K. B. (2012). Reading with a filtered fovea: The influence of visual quality at the point of fixation during reading. Psychonomic Bulletin & Review, 19, 1078–1084. doi:10.3758/s13423-012-0307-x
Klein, G. S. (1964). Semantic power measured through the effect of words with color-naming. American Journal of Psychology, 77, 576–588.
Lachter, J., Forster, K. I., & Ruthruff, E. (2004). Forty-five years after Broadbent (1958): Still no identification without attention. Psychological Review, 111, 880–913. doi:10.1037/0033-295X.111.4.880
Lachter, J., Ruthruff, E., Lien, M.-C., & McCann, R. S. (2008). Is attention needed for word identification? Evidence from the Stroop paradigm. Psychonomic Bulletin & Review, 15, 950–955. doi:10.3758/PBR.15.5.950
Luo, C. R. (1999). Semantic competition as the basis of the Stroop interference: Evidence from color-word matching tasks. Psychological Science, 10, 35–40.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review. Psychological Bulletin, 109, 163–203. doi:10.1037/0033-2909.109.2.163
Nazir, T. A., Jacobs, A. M., & O’Regan, J. K. (1998). Letter legibility and visual word recognition. Memory & Cognition, 26, 810–821.
Neely, J. H., & Kahan, T. (2001). Is semantic activation automatic? A critical re-evaluation. In H. L. Roediger III, J. S. Nairne, I. Neath, & A. M. Surprenant (Eds.), The nature of remembering: Essays in honor of Robert G. Crowder (pp. 69–93). Washington, DC: American Psychological Association.
O’Regan, J. K., & Jacobs, A. M. (1992). Optimal viewing position effect in word recognition: A challenge to current theory. Journal of Experimental Psychology: Human Perception and Performance, 18, 185–197. doi:10.1037/0096-1523.18.1.185
O’Regan, J. K., Lévy-Schoen, A., Pynte, J., & Brugaillère, B. (1984). Convenient fixation location within isolated words of different length and structure. Journal of Experimental Psychology: Human Perception and Performance, 10, 250–257. doi:10.1037/0096-1523.10.2.250
Parris, B. A., Sharma, D., & Weekes, B. (2007). An optimal viewing position effect in the Stroop task when only one letter is the color carrier. Experimental Psychology, 54, 273–280.
Perret, P., & Ducrot, S. (2010). Viewing-position effects in the Stroop task: Initial fixation position modulates Stroop effects in fully colored words. Psychonomic Bulletin & Review, 17, 550–555. doi:10.3758/PBR.17.4.550
Risko, E. F., Stolz, J. A., & Besner, D. (2005). Basic processes in reading: Is visual word recognition obligatory? Psychonomic Bulletin & Review, 12, 119–124. doi:10.3758/BF03196356
Roberts, M. A., & Besner, D. (2005). Stroop dilution revisited: Evidence for domain-specific, limited-capacity processing. Journal of Experimental Psychology: Human Perception and Performance, 31, 3–13. doi:10.1037/0096-1523.31.1.3
Schmidt, J. R., & Cheesman, J. (2005). Dissociating stimulus–stimulus and response–response effects in the Stroop task. Canadian Journal of Experimental Psychology, 59, 132–138.
Sharma, D., Booth, R., Brown, R., & Huguet, P. (2010). Exploring the temporal dynamics of social facilitation in the Stroop task. Psychonomic Bulletin & Review, 17, 52–58. doi:10.3758/PBR.17.1.52
Sharma, D., & McKenna, F. P. (1998). Differential components of the manual and vocal Stroop tasks. Memory & Cognition, 26, 1033–1040. doi:10.3758/BF03201181
Smilek, D., Solman, G. J. F., Murawski, P., & Carriere, J. S. A. (2009). The eyes fixate the optimal viewing position of task-irrelevant words. Psychonomic Bulletin & Review, 16, 57–61. doi:10.3758/PBR.16.1.57
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18, 643–662. doi:10.1037/0096-3445.121.1.15
van Veen, V., & Carter, C. S. (2005). Separating semantic conflict and response conflict in the Stroop task: A functional MRI study. NeuroImage, 27, 497–504. doi:10.1016/j.neuroimage.2005.04.042
Waechter, S., Besner, D., & Stolz, J. A. (2011). Basic processes in reading: Spatial attention as a necessary preliminary to orthographic and semantic processing. Visual Cognition, 19, 171–202.
Yao-N’Dré, M., Castet, E., & Vitu, F. (2013). The optimal viewing position effect in the lower visual field. Vision Research, 76, 114–123.
Author Note
Both authors thank Stéphanie Ducrot for sharing the computer program used in their original study, Johanna Paul for running the experiment, and Melvin Yap, Derek Besner, Benjamin Parris, and one anonymous reviewer for their helpful advice, comments, and suggestions on previous drafts of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ferrand, L., Augustinova, M. Differential effects of viewing positions on standard versus semantic Stroop interference. Psychon Bull Rev 21, 425–431 (2014). https://doi.org/10.3758/s13423-013-0507-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-013-0507-z