
Abstract
Reading relies critically on processing text in foveal vision during brief fixational pauses, and high-quality visual input from foveal text is fundamental to theories of reading. However, the quality of visual input from foveal text that is actually functional for reading and the effects of this input on reading performance are unclear. To investigate these issues, a moving, gaze-contingent foveal filtering technique was developed to display areas of text within foveal vision that provided only coarse, medium, or fine scale visual input during each fixational pause during reading. Normal reading times were unaffected when foveal text up to three characters wide at the point of fixation provided any one visual input (coarse, medium, or fine). Wider areas of coarse visual input lengthened reading times, but reading still occurred, and normal reading times were completely unaffected when only medium or fine visual input extended across the entire fovea. Further analyses revealed that each visual input had no effect on the number of fixations made when normal text was read, that adjusting fixation durations helped preserve reading efficiency for different visual inputs, and that each visual input had virtually no effect on normal saccades. These findings indicate that, despite the resolving power of foveal vision and the emphasis placed on high-quality foveal visual input by theories of reading, normal reading functions with similar success using a range of restricted visual inputs from foveal text, even at the point of fixation. Some implications of these findings for theories of reading are discussed.
Similar content being viewed by others


Due to the nature of the human visual system, foveal vision provides the highest quality of visual input across an area approximately 2° wide centered at the point of fixation (e.g., Green, 1970; Riggs, 1965). As a consequence, when we look around our visual world, our fixations shift so that different objects can be brought into foveal vision and perceived using high-quality foveal input to provide the richest, most complete, and most detailed visual information we can see.
Perhaps unsurprisingly, high-quality foveal input is widely regarded as essential for a range of complex visual tasks, especially reading (for a review, see Rayner, 2009). Indeed, it is well-known that normal reading along a line of text relies on saccadic eye movements to make shifts in fixation location so that areas of text previously located away from the fovea can be brought into foveal vision. In this way, each fixation is well-suited to provide high-quality visual input from text within foveal vision, which may then contribute to the efficiency with which words are processed during reading (e.g., McConkie, Kerr, Reddix, & Zola, 1988; O’Regan, Lévy-Schoen, Pynte, & Brugaillère, 1984; Stevens & Grainger, 2003).
The role of high-quality foveal input in reading seems intuitively obvious. Indeed, a long-standing view is that the rich detail provided by high-quality foveal input is crucially important for word recognition (e.g., McConkie et al., 1988; O’Regan et al., 1984; Stevens & Grainger, 2003), and models of reading emphasize greatly the importance of high-quality visual input from text falling within foveal vision during each fixational pause (e.g., Engbert, Nuthmann, Richter, & Kliegl, 2005; Pollatsek, Reichle, & Rayner, 2006; Reichle, Pollatsek, Fisher, & Rayner, 1998; Reichle, Rayner, & Pollatsek, 2003). Moreover, substantial impairments in reading are known to occur when sections of foveal vision are completely obliterated, either by experimental manipulation (e.g., Fine & Rubin, 1999; Rayner & Bertera, 1979; Rayner, Inhoff, Morrison, Slowiaczek, & Bertera, 1981) or by natural degeneration (e.g., Faye, 1984; Legge, Rubin, Pelli, & Schleske, 1985; Owsley, 2011; Whittaker & Lovie-Kitchin, 1993), and impairments occur even when words in foveal vision are simply made faint (e.g., Drieghe, 2008; Reingold & Rayner, 2006).
But despite the clear importance of foveal vision for reading, the extent to which normal reading actually relies on high-quality foveal input is uncertain. Of particular importance is that since foveal vision provides the highest quality input, reading may suffer considerably when this high-quality input is not available for text brought into foveal vision at each fixational pause. Indeed, the resolving power of foveal vision is at its maximum at the point of fixation, even higher than in the remainder of the fovea (e.g., Green, 1970; Riggs, 1965), and so reliance on high-quality visual input for reading may be particularly great at the exact location of each fixation.
Reliance on high-quality foveal input would certainly fit the views of current theories of word recognition and reading. But although words viewed within foveal vision normally appear perfect, complete, and fully resolved to the reader, foveal vision can actually provide an array of different types of visual input, ranging from coarse scale visual input that may be useful for determining the overall size, shape, and location of words to fine scale visual input that can specify the exact form of distinct letter fragments and the precise appearance of individual letters (e.g., Allen, Smith, Lien, Kaut, & Canfield, 2009; Blakemore & Campbell, 1969; Boden & Giaschi, 2009; Jordan, 1990, 1995; Patching & Jordan, 2005a, b; Robson, 1966; see also Young, Liversedge, Love, Myers, & Smithson, 2011). Consequently, although high-quality foveal input is generally available and many believe this input is crucial for normal reading, it remains to be seen which types of visual input acquired from foveal text during fixational pauses actually contribute to normal reading and how these different types of visual input affect reading performance. Accordingly, the purpose of the present study was to combine established eye movement procedures with psychophysical manipulations to selectively restrict the high-quality visual input normally available from foveal text during fixational pauses and to determine the effect of each restricted foveal input on normal reading performance.
To investigate these issues, the widely adopted moving mask technique (e.g., Duchowski, 2007; McConkie & Rayner, 1975; Rayner & Bertera, 1979; Rayner, Liversedge, & White, 2006) was adapted to produce a novel foveal filtering paradigm in which skilled adult readers read lines of text that were presented normally, except for a virtual filter within foveal vision that was centered at the location of each fixational pause. The location of the filter was yoked to the direction of the reader’s gaze so that, when the reader’s eyes moved to fixate a new location along a line of text, the filter moved invisibly in synchrony with these eye movements and was present at the new fixation location. For each fixational pause, text outside each filtered area was displayed as normal, but text within each filtered area provided only coarse, medium, or fine visual input, corresponding to three bands of spatial frequencies (see the Method section). The spatial frequency content of these bands differed substantially but are known to be influential in single-word recognition (e.g., Patching & Jordan, 2005a, b) and so were well-suited to reveal differences in the use of foveal visual content during reading. The width of the filtered area (and so the amount of foveal text that was filtered during each fixational pause) ranged from one character (the character fixated) to nine characters (approximating the entire width of the fovea). The phenomenological experience of all these displays was that each filtered area moved in perfect synchrony with the eyes during reading.
This is the first study in which the effects of restricting the spatial frequency content of foveal input on normal reading performance have been specifically investigated, and theories of reading currently lack the explanatory power to drive fine-grained predictions about these effects. Consequently, aspects of this work were necessarily exploratory, and making too many predictions would be overspeculative. Hence, the logic of the research was kept as straightforward as possible. If high-quality foveal input during fixational pauses is crucial for reading, as many theories assume, reading should be severely disrupted when this high-quality input is not available. Indeed, the disruptive influence of removing high-quality foveal input should be particularly apparent at the location of each fixation, since visual resolution here will be maximal. However, if reading is more tolerant of the quality of foveal input than is currently assumed, the nature and extent of this tolerance should become apparent when only filtered visual input is available. To provide a comprehensive measure of the influence of foveal input, reading performance was assessed by recording overall reading time, mean fixation durations (the average length of fixational pauses during reading), total number of fixations (the number of these fixational pauses), regressive saccade count (the number of backward movements in the text), and the length of progressive and regressive saccades.
Method
Participants
Sixteen participants, 18–25 years of age, were recruited from the local university community. All participants were native speakers of English and had normal or corrected-to-normal vision, as determined by Bailey–Lovie (Bailey & Lovie, 1980), ETDRS (Ferris & Bailey, 1996), and Pelli–Robson (Pelli, Robson, & Wilkins, 1988) assessments (see Jordan, McGowan, & Paterson, 2011).
Design and materials
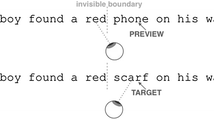
One hundred sixty sentences were displayed either entirely as normal or using a moving filter one, three, five, seven, or nine characters wide, centered at each point of fixation. These widths were chosen to provide filtered areas ranging from just one character wide to the width of the fovea. Text outside each filtered area was normal, and text within each filtered area was altered using MATLAB to leave one of three different, 1-octave wide bands of spatial frequency content with peak frequencies of 3.5, 6.7, and 11.1 cycles per degree (cpd) and low-pass and high-pass cutoff frequencies of 2.6–5.2, 5.0–10.0, and 8.3–16.6 cpd, for coarse, medium, and fine input, respectively (see Patching & Jordan, 2005a, b). These manipulations were achieved by point-wise multiplication in the frequency domain using fourth-order high- and low-pass Butterworth filters, which provide a mathematically tractable filter that avoids problems of ringing associated with other filters. Crucially, these manipulations did not eliminate completely the visual composition of foveal text but selectively restricted its spatial frequency content and so selectively degraded the normal high-quality nature of foveal visual input. The resulting sentence displays were randomized and chosen using a Latin square design, so that each participant saw an equal number of sentences in each display condition but saw each sentence only once. This enabled all sentences to be shown equally often in each display condition across participants but avoided repetition of any sentence for any participant. Figure 1 provides indications of the appearance of the displays that were used.

Indications of the displays used in the experiment. The figure shows a sentence displayed as normal and with a nine-character foveal filter providing only coarse, medium, or fine visual input. The visual appearance of the input shown in the figure is approximate due to variations in display resolution and print medium
Apparatus and procedure
Eye movements were recorded using an Eyelink 2K eyetracker with a spatial resolution of 0.01°. At the viewing distance of approximately 85 cm, nine letters subtended approximately 2.25°. Sentences were displayed on a 19-in. monitor at 100 Hz, and eye position was sampled at 1000 Hz using corneal reflection and pupil tracking. Custom software ensured that the filter moved in close synchrony with eye movements, and display changes were made within 8–12 ms. On each trial, participants fixated a location on the left of the screen, and a sentence was then presented, with its first letter at the fixation location. Participants were instructed to read normally and for comprehension and answered a comprehension question after each sentence.
Results
Participants showed high levels of comprehension (99 % correct responses to questions) and no differences between normal displays and displays using coarse, medium, or fine visual input, for any filtered area (all Fs < 1.60). Reading times are shown in Fig. 2, and fixation durations, number of fixations, number of regressions, progressive saccade length, and regressive saccade length are reported in Table 1. For each of these measures, the purpose of the experiment was to determine the effect of each visual input on the performance observed for normal displays. Accordingly, for each measure, a one-way within-subjects analysis of variance compared performance for each visual input and filtered area with performance for normal displays. Post hoc comparisons were performed using Bonferroni-corrected Tukey tests.

Mean sentence reading times (in milliseconds) for different visual input and filter sizes
Reading time
For coarse visual input, F(5, 75) = 19.05, p < .001, η 2p = .56; reading times were no different from normal for filtered areas one and three characters wide (ps > .90) but were longer than normal for filtered areas five, seven, and nine characters wide (ps < .05). For medium and fine visual input, reading times were no different from normal for any filtered area (Fs < 1).
Fixation duration
For coarse visual input, F(5, 75) = 29.89, p < .001, η 2p = .67; fixation durations were longer than normal for all filtered areas (ps < .05). For medium visual input, fixation durations were no different from normal for any filtered area (F < 1). For fine visual input, F(5, 75) = 8.75, p < .001, η 2p = .37; fixation durations were no different from normal for filtered areas one and three characters wide (ps > .05) but were longer than normal for filtered areas five, seven, and nine characters wide (ps < .05).
Number of fixations
Number of fixations was no different from normal for any visual input for any filtered area (Fs < 1).
Number of regressions
For coarse visual input, F(5, 75) = 8.18, p < .001, η 2p = .35; slightly more regressions than normal were made for filtered areas nine characters wide (p < .05). For medium and fine visual input, number of regressions was no different from normal for any filtered area (Fs < 1).
Progressive saccade length
Length of progressive saccades was no different from normal for any visual input for any filtered area (Fs < 1).
Regressive saccade length
Length of regressive saccades was no different from normal for any visual input for any filtered area (Fs < 1).
Discussion
Despite the resolving power of the fovea, reading performance was remarkably tolerant of substantial restrictions to the visual input available from text in foveal vision. Indeed, normal reading times were unaffected when text up to three characters wide at the point of fixation provided any one visual input (coarse, medium, or fine), despite the highly restricted visual content of each visual input and the substantial differences in visual content that each input provided. Moreover, larger areas of coarse visual input lengthened reading times, but reading still occurred, and presenting only medium or fine visual input across the entire fovea had no effect at all on the speed with which sentences were normally read. The numbers of fixations produced by each visual input were also very similar to the numbers observed for normal text, indicating that the restriction of visual input across the fovea did not disrupt the normal amount of fixations required to obtain information for reading.
These patterns of performance provide important caveats for the widely held view that normal reading depends critically on high-quality foveal input (e.g., Engbert et al., 2005; McConkie et al., 1988; O’Regan et al., 1984; Pollatsek et al., 2006; Reichle et al., 2003; Stevens & Grainger, 2003). In particular, although visual input from foveal text is important, theories of reading should not assume that the input provided reflects the full resolving power of foveal vision or that only high-quality visual input from foveal text can support normal reading. Indeed, this point is emphasized by the similarities in reading times and number of fixations observed when visual input was restricted at and around the actual location of each fixation (i.e., for filters one and three characters wide), where the resolving power of foveal vision is at its maximum, even higher than in the remainder of the fovea (e.g., Green, 1970; Riggs, 1965).
Further analyses indicated that alterations to the duration of fixational pauses often contributed to the tolerance of restricted visual input. While medium visual input produced no change in the fixation durations observed for normal text, longer fixations than normal occurred for coarse visual input for all filter sizes and for fine visual input five, seven, and nine characters wide, but no disruption in normal reading times was apparent for coarse visual input up to three characters wide or for fine visual input in any filter size. Thus, while changes in fixation duration indicate that visual input affected the processing of words in foveal vision, altering the duration of fixational pauses appears to be a process that often allows reading efficiency to be preserved when foveal visual input is restricted. Indeed, this influence of fixation duration may help maintain reading efficiency under the variable viewing conditions that affect the quality of foveal visual input for readers in everyday life. For example, the moment-by-moment quality of foveal visual input is affected continually by slight variations in reading distance, lighting, and print quality, and even by blinking due to visual suppression and the intermittent tear film that alters the optical characteristics of the eyes (e.g., Montés-Micó, 2007; Ridder & Tomlinson, 1993, 1995). Dealing with variable visual quality in foveal input, therefore, may be a common component of reading.Footnote 1
Other factors are likely to contribute to the effectiveness of restricted foveal visual input during reading and to a tolerance of variations in this input. In particular, interactions between visual processing and cognitive processes associated with the syntactic, linguistic, and semantic content of language are normal in reading (see Rayner, 2009), and these interactions are naturally likely to involve foveal input. Indeed, when fixations are made during reading, information is also acquired from locations extending outside the fovea in the direction of reading, and this extrafoveal information is used to preprocess the identity of words before the next saccade is made in their direction (see Rayner, 2009). Consequently, when text is read from left to right, processing extrafoveal words to the right of fixation may be underway before the next fixation takes place, and this preprocessing may facilitate the recognition of foveal words from limited visual input when a fixation brings into foveal vision text seen previously in extrafoveal locations. Indeed, since the visual resolution available in extrafoveal locations is relatively coarse, the use of coarse visual input in foveal vision may help assimilate the processing of words in extrafoveal and foveal locations and so help integrate textual information across saccades during reading.
The findings of this study clearly suggest that a range of visual inputs at and around the point of fixation activate processes of word recognition during reading, and this has further implications for understanding when and where eye movements occur. In particular, these findings show that the quality of foveal visual input does not generally influence eye movement planning or the subsequent processing of words occupying locations outside foveal vision, and this distinction is broadly consistent with the E-Z Reader model (e.g., Pollatsek et al., 2006; Reichle et al., 1998), where access to high-quality foveal input modulates the time spent processing fixated words, but not the subsequent eye movements or processing of words away from fixation (e.g., Reingold & Rayner, 2006; see also Drieghe, 2008). But our findings now show that this distinction in normal reading behaviour is supported by a range of different visual inputs from foveal text and that the distinction cannot be explained by considering access to high-quality foveal input alone. Indeed, while changes in foveal visual input produced clear changes in fixation durations, the pattern of saccades produced by normal displays was essentially unaltered when any one visual input (coarse, medium, or fine) extended across the entire fovea. Accordingly, our findings add weight to the view that decisions about when to move the eyes from fixation during reading and where to move them next along a line of text are made independently (e.g., Rayner, 2009; Reichle, Liversedge, Rayner, & Pollatsek, 2009) but show that this aspect of normal reading is driven by a range of visual inputs from foveal text across a substantial region around the point of fixation when text is read. Indeed, this use of multiple visual inputs in foveal vision may be a powerful component of the efficiency with which visual information supports reading (e.g., Rayner, Liversedge, White, & Vergilino-Perez, 2003).
Finally, while some manipulations of coarse and fine visual input affected aspects of normal reading performance, medium visual input produced no disruption at all for any measure of reading performance for any filtered area. The parsimonious interpretation of this finding is that although medium visual input is just part of the visual composition of foveal text and clearly does not provide high-quality foveal input, medium visual input is all that is required from foveal vision for normal reading to take place. Further research will clarify this role, but the indication already is that the role of foveal input in reading cannot be determined by considering the importance of high-quality visual input alone. Instead, normal reading can clearly function successfully using a range of restricted visual inputs from foveal text during each fixational pause and does not require high-quality input from foveal vision, even at the point of fixation.
Notes
The number of fixations produced by each visual input was very similar to the number observed for normal text. However, fixation number was often slightly (although never significantly) higher with normal text, suggesting that dealing with variable visual quality in foveal input may sometimes involve small reductions in fixation numbers.
References
Allen, P. A., Smith, A. F., Lien, M. C., Kaut, K. P., & Canfield, A. (2009). A multi-stream model of visual word recognition. Attention, Perception, & Psychophysics, 71, 281–296.
Bailey, I. L., & Lovie, J. E. (1980). The design and use of a new near-vision chart. American Journal of Optometry and Physiological Optics, 57, 378–387.
Blakemore, C., & Campbell, F. W. (1969). Adaptation to spatial stimuli. The Journal of Physiology, 2000, 11–13.
Boden, C., & Giaschi, D. (2009). The role of low-spatial frequencies in lexical decision and masked priming. Brain and Cognition, 69, 580–591.
Drieghe, D. (2008). Foveal processing and word skipping during reading. Psychonomic Bulletin & Review, 15, 856–860.
Duchowski, A. T. (2007). Eye tracking methodology: Theory and practice. London: Springer.
Engbert, R., Nuthmann, A., Richter, E., & Kliegl, R. (2005). SWIFT: A dynamical model of saccade generation during reading. Psychological Review, 112, 777–813.
Faye, E. E. (1984). Clinical Low Vision. Boston: Little, Brown & Co.
Ferris, F. L., & Bailey, I. L. (1996). Standardizing the measurement of visual acuity for clinical research studies. Ophthalmology, 103, 181–182.
Fine, E. M., & Rubin, G. S. (1999). Reading with a central field loss: Number of letters masked is more important than the size of the mask in degrees. Vision Research, 39, 747–756.
Green, D. G. (1970). Regional variations in the visual acuity for interference fringes on the retina. The Journal of Physiology, 207, 351–356.
Jordan, T. R. (1990). Presenting words without interior letters: Superiority over single letters and influence of postmask boundaries. Journal of Experimental Psychology. Human Perception and Performance, 16, 893–909.
Jordan, T. R. (1995). Perceiving exterior letters of words: Differential influences of letter-fragment and nonletter fragment masks. Journal of Experimental Psychology. Human Perception and Performance, 21, 512–530.
Jordan, T. R., McGowan, V. A., & Paterson, K. B. (2011). Out of sight, out of mind: The rarity of assessing and reporting participants’ visual abilities when studying perception of linguistic stimuli. Perception, 40, 873–876.
Legge, G. E., Rubin, G. S., Pelli, D. G., & Schleske, M. M. (1985). Psychophysics of reading. II. Low vision. Vision Research, 25, 253–266.
McConkie, G. W., Kerr, P. W., Reddix, M. D., & Zola, D. (1988). Eye movement control during reading: I. The location of initial fixations in words. Vision Research, 28, 1107–1118.
McConkie, G. W., & Rayner, K. (1975). The span of the effective stimulus during a fixation in reading. Perception & Psychophysics, 17, 578–586.
Montés-Micó, R. (2007). Role of the tear film in the optical quality of the human eye. Journal of Cataract and Refractive Surgery, 33, 1631–1635.
O’Regan, J. K., Lévy-Schoen, A., Pynte, J., & Brugaillère, B. (1984). Convenient fixation location within isolated words of different length and structure. Journal of Experimental Psychology. Human Perception and Performance, 10, 250–257.
Owsley, C. (2011). Aging and vision. Vision Research, 51, 1610–22.
Patching, G. R., & Jordan, T. R. (2005a). Assessing the role of different spatial frequencies in word perception by good and poor readers. Memory & Cognition, 33, 961–971.
Patching, G. R., & Jordan, T. R. (2005b). Spatial frequency sensitivity differences between adults of good and poor reading ability. Investigative Ophthalmology & Visual Science, 46, 2219–2224.
Pelli, D. G., Robson, J. G., & Wilkins, A. J. (1988). The design of a new letter chart for measuring contrast sensitivity. Clinical Vision Science, 2, 187–199.
Pollatsek, A., Reichle, E. D., & Rayner, K. (2006). Tests of the E-Z Reader model: Exploring the interface between cognition and eye-movement control. Cognitive Psychology, 52, 1–56.
Rayner, K. (2009). Eye movements and attention in reading, scene perception, and visual search. The Quarterly Journal of Experimental Psychology, 62, 1457–1506.
Rayner, K., & Bertera, J. H. (1979). Reading without a fovea. Science, 206, 468–469.
Rayner, K., Inhoff, A. W., Morrison, R. E., Slowiaczek, M. L., & Bertera, J. H. (1981). Masking of foveal and parafoveal vision during eye fixations in reading. Journal of Experimental Psychology. Human Perception and Performance, 7, 167–179.
Rayner, K., Liversedge, S. P., & White, S. J. (2006). Eye movements when reading disappearing text: The importance of the word to the right of fixation. Vision Research, 46, 310–323.
Rayner, K., Liversedge, S. P., White, S. J., & Vergilino-Perez, D. (2003). Reading disappearing text: Cognitive control of eye movements. Psychological Science, 14, 385–388.
Reichle, E. D., Liversedge, S. P., Rayner, K., & Pollatsek, A. (2009). Encoding multiple words simultaneously in reading is implausible. Trends in Cognitive Sciences, 13, 115–119.
Reichle, E. D., Pollatsek, A., Fisher, D. L., & Rayner, K. (1998). Toward a model of eye movement control in reading. Psychological Review, 105, 125–157.
Reichle, E. D., Rayner, K., & Pollatsek, A. (2003). The E-Z Reader model of eye-movement control in reading: Comparisons to other models. The Behavioral and Brain Sciences, 26, 445–526.
Reingold, E. M., & Rayner, K. (2006). Examining the word identification stages hypothesized by the E - Z reader model. Psychological Science, 17, 742–746.
Ridder, W. H., & Tomlinson, A. (1993). Suppression of contrast sensitivity during eyelid blinks. Vision Research, 33, 1795–1802.
Ridder, W. H., & Tomlinson, A. (1995). Spectral characteristics of blink suppression in normal observers. Vision Research, 35, 2569–2578.
Riggs, L. A. (1965). Visual acuity. In C. H. Graham (Ed.), Vision and visual perception (pp. 321–349). New York: Wiley.
Robson, J. G. (1966). Spatial and temporal contrast sensitivity functions of the visual system. Journal of the Optical Society of America, 56, 141–142.
Stevens, M., & Grainger, J. (2003). Letter visibility and the viewing position effect in visual word recognition. Perception & Psychophysics, 65, 133–151.
Whittaker, S. G., & Lovie-Kitchin, J. (1993). Visual requirements for reading. Optometry and Vision Science, 70, 54–65.
Young, L. K., Liversedge, S. P., Love, G. D., Myers, R. M., & Smithson, H. E. (2011). Reading impairment depends on aberration type and magnitude. Journal of Vision, 11(13), 1–19. 20.
Author Notes
This research was supported by the Ulverscroft Foundation and by a Professorial Fellowship from the Economic Research Foundation awarded to Tim Jordan and a Mid-Career Fellowship from the British Academy awarded to Kevin Paterson.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jordan, T.R., McGowan, V.A. & Paterson, K.B. Reading with a filtered fovea: The influence of visual quality at the point of fixation during reading. Psychon Bull Rev 19, 1078–1084 (2012). https://doi.org/10.3758/s13423-012-0307-x
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-012-0307-x