
Abstract
Growing evidence indicates that a domain-general executive control supports semantic memory retrieval, yet the nature of this interaction remains elusive. To shed light on such control mechanisms, we conducted two dual-task experiments loading distinct executive capacities (working memory maintenance, monitoring, and switching), while participants carried out automatic (free-associative) and controlled (dissociative) word retrieval tasks. We found that these forms of executive load interfered with retrieval fluency in both tasks, but these negative effects were more pronounced for the dissociative performance. Together, these findings indicate that the domain-general executive control supports accessing contextually relevant knowledge as well as the inhibition of automatically activated but task-inappropriate retrieval candidates, putatively via an adaptive gating of semantic activation and interference control. Moreover, the processing costs related to retrieval inhibition and switching were negatively correlated, suggesting a trade-off between the ability to constrain semantic activation (i.e., inhibition) and the ability to initiate flexible transitions between semantic sets (i.e., switching), which may thus represent two complementary control functions governing semantic memory retrieval.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
A challenge in research on human cognition is to understand the mechanisms enabling fluid retrieval of knowledge that is suitable for the current context and situational demands. Contemporary neurocognitive models identify two functionally interacting systems underpinning two distinct modes of knowledge retrieval (Jefferies et al., 2020; Lambon Ralph et al., 2017). The system for semantic representation supports the integration of knowledge distilled through experience and mediates automatic retrieval (i.e., a bottom-up activation of semantic representations driven by environmental cues or spontaneous thought; Marron et al., 2020). On the other hand, the system for semantic control is employed to exert an executive manipulation of the activations within the representational system and mediates controlled retrieval (i.e., a top-down regulation of semantic processing, which is recruited when the automatic retrieval generates outputs that are not adequate for the task at hand; Badre & Wagner, 2007). This control of semantic retrieval is implemented by putative semantic control mechanisms, such as the inhibition of typical but inappropriate associates (Allen et al., 2008; Collette et al., 2001; Marko, Cimrová, et al., 2019a) and flexible switching between semantic sets (Marko, Michalko, et al., 2019b; Marko & Riečanský, 2021b; Troyer et al., 1997).
One of the most fundamental yet unresolved questions is how the semantic (domain-specific) systems interact with the multiple-demand (domain-general) executive control system. Regardless of the type of representation (e.g., visual, motor, semantic), any nontrivial task, especially novel or demanding, is encoded in working memory (WM) and supported by executive attention (EA; Kane & Engle, 2002), which enables a functional coupling between the domain-general control and semantic processes (Chai et al., 2016; Fedorenko & Thompson-Schill, 2014). The putative contributions of WM and EA to semantic memory retrieval have been previously investigated in experiments using verbal fluency tasks. These experiments have shown that individuals with high executive capacity are generally able to name more category exemplars, retrieve larger semantic clusters, and produce fewer perseverations compared with individuals with low executive capacity (Rende et al., 2002; Rosen & Engle, 1997; Unsworth et al., 2013). Interestingly, these advantages can be eliminated by concurrent WM/EA load (Rende et al., 2002; Rosen & Engle, 1997), or by providing the low-capacity individuals with retrieval cues (Unsworth et al., 2013), which has been taken as evidence that WM and EA contribute to semantic processing and strategic knowledge retrieval (Hills et al., 2013; Hills et al., 2015; Hirshorn & Thompson-Schill, 2006; Mayr & Kliegl, 2000; Troyer et al., 1998; Ulrich Mayr, 2002). A principal limitation of these findings, however, concerns the construct of verbal fluency, which is rather ambiguous regarding the involvement of automatic versus controlled processing and the reliance on knowledge/language versus executive capacity (Henry & Crawford, 2004; Shao et al., 2014; Whiteside et al., 2016). Moreover, we (Michalko et al., 2022) have recently demonstrated that the relative involvement of domain-general control in semantic fluency may change as a function of time performing the task (also see Crowe, 1998; Demetriou & Holtzer, 2017)—that is, as typical and easily available retrieval candidates are depleted. Hence, due to these shortcomings, the previous experiments using verbal fluency can provide only limited conclusions on the putative interaction between the domain-general (executive) and semantic processes.
Taken together, converging evidence indicates that domain-general executive capacity plays a role in semantic memory retrieval, but the nature of this involvement remains poorly understood. Here we addressed this issue in two experiments incorporating dual-task and task-switching interference paradigms (Strobach et al., 2018) that manipulated WM (Experiment 1) and EA (Experiment 2) load while participants continuously retrieved words from semantic memory. This dual-task technique was adopted as it inherently depletes multiple-demand executive resources (Baddeley, 2012; Szameitat et al., 2002), making this approach suitable to study the contribution of the respective executive capacities in semantic memory retrieval.
Importantly, in contrast to the previous research relying on the relatively ambiguous verbal fluency measures, we used a novel word-generation paradigm (Marko, Cimrová, et al., 2019a; Marko, Michalko, et al., 2019b) that employs distinct retrieval tasks, separately assessing the automatic (associative) and the controlled (dissociative) processes involved in semantic retrieval. This distinction enabled us to shed light on the specific contributions of WM and EF in these retrieval modes and the specific processes they inherently involve.
General method
A group of participants completed two experiments within a single session in a counterbalanced order, including a break between them. The required sample size was determined using an a priori power analysis (5% Type I error rate, 20% Type II error rate, and expected effect size \({\eta}_p^2\) > .10 for repeated measures [RM] analysis of variance [ANOVA] main effects and interactions; Faul et al., 2007). In total, 64 young, healthy adults were recruited to participate in the study, however, one subject was excluded due to multivariate outliers (>3 SD) in both experiments. The final sample thus included 63 participants (25 males; mean age = 22.89 years, SD = 2.84, range: 19–31 years; two participants were left-handed). None of the participants reported a history of psychiatric disorder, neurological condition, learning/reading disability, or recent medication. The research was conducted in accordance with the Declaration of Helsinki (World Medical Association, 2013) and approved by the institutional review board. All procedures and methods were carried out in accordance with the relevant guidelines and regulations. All participants gave written informed consent and received a financial reward for their participation.
Experiment 1: Working memory load
The first experiment addressed the contribution of WM to automatic and controlled semantic retrieval using concurrent WM load manipulation. To this aim, we manipulated the dual-task interference targeting verbal working memory (no, low, high WM load) while participants performed an automatic or a controlled retrieval task. We expected that the performance in both retrieval tasks would be disrupted as a function of WM load, but the controlled (dissociative) retrieval would be more impaired than the automatic (free-associative) memory retrieval. More precisely, we hypothesized that the maintenance cost (i.e., retrieval slowing due to the WM load) will be larger in the dissociative than the associative condition, and relatedly, that the inhibition cost will increase linearly with the WM load (see Supplementary Information for more details on these custom contrasts/hypotheses). Notably, given that limited WM resources can be allocated either to the primary or secondary task within the experiment (Hegarty et al., 2000), higher demands on WM during dissociative versus associative retrieval could also, or alternatively, manifest in impaired WM performance in the secondary task. Therefore, either of these effect (i.e., higher maintenance/inhibition cost or worse WM performance) would suggest that generating dissociates exhausts more WM resources than the free-associative retrieval. Finally, we performed a robust correlational analysis aimed to explore the links between the retrieval measures from the experiment and tasks assessing individual working memory capacity (WMC) as well as the maintenance cost.
Methods
Semantic retrieval measures
Semantic retrieval was assessed using a modified version of the associative chain test (Marko, Cimrová, et al., 2019a; Marko, Michalko, et al., 2019b), in which participants continuously generated word chains following two specific retrieval conditions (a within-subject factor retrieval). In the associative retrieval condition, participants were instructed to produce chains of nonrepeated words (nouns) so that each new response in the chain was semantically associated with the previous one (e.g., “Tree–Garden–House–Door…”). In this task, the participants were encouraged to respond with the first (free) association that comes to their mind, which is considered to involve predominantly automatic (i.e., spontaneous, bottom-up) semantic processing with little executive effort (Marko, Cimrová, et al., 2019a; Marko, Michalko, et al., 2019b; Marron et al., 2020). In the dissociative retrieval condition, participants were instructed to produce a chain of semantically unrelated words (nouns), in which each new response does not relate to the previous one (e.g., “Night–Hockey–Dog–Computer…”), and informed that delivering a related word or repetitions would count as an error. Notably, retrieving unrelated words was repeatedly shown to require more time than retrieving free associations, which was attributed to additional controlled (i.e., effortful, top-down) inhibitory demands (Allen et al., 2008; Collette et al., 2001; Marko, Cimrová, et al., 2019a; Marko, Michalko, et al., 2019b). Thus, the difference in latency between dissociative and associative retrieval—that is, inhibition cost (ΔRT dissociative – associative)—is considered a measure reflecting the ability to suppress automatic (stimulus-driven) semantic activation and/or responses (i.e., high inhibition cost indicates less efficient semantic inhibition).
The retrieval experiment included a total of 21 trials (chains), nine trials for the associative and 12 trials for the dissociative retrieval condition presented in random order (different number of trials were included to account for the expected differences in the mean retrieval latency between the conditions observed in the previous studies, see Marko et al., 2022). In each trial, a stimulus word appeared on the computer screen and participants started generating a chain of words following the respective rule (associative/dissociative) for 20 seconds. Participants entered the words via keyboard and were instructed to keep fluent production, ignore grammatical or typing errors, and not repeat words within the same chain. Each response was assessed for response time (RT)—that is, the latency of entering the first letter of the word. As in the previous studies (Marko et al., 2022; Marko & Riečanský, 2021a), response words were screened for errors by two trained and blinded raters, and those responses marked as error by both raters were excluded prior to statistical analyses. The split-half reliability for the associative and the dissociative retrieval measure was sufficiently high (r = .742 and .804, respectively).
Working-memory load manipulation
The manipulation of WM load included three conditions (a within-subject factor WM Load): The no-load condition was a control condition without any WM load, the low load included a three-digit load, and the high load included a six-digit load. All three conditions were presented at random and were balanced across the retrieval conditions. In the trials including the load (low/high), a set of randomly generated unique digits (three digits/six digits) was presented for a short time (4 seconds/8 seconds) before the stimulus word for the retrieval task appeared. During this short interval, participants were instructed to remember the digits in the correct order (encoding). They were then required to keep the digits in memory while performing the retrieval task (i.e., 20 seconds; maintenance) and write them down using a computer keyboard once the retrieval task had finished (recall; see Fig. 1 for a graphical description of an experimental trial). The participants were reminded that a rapid retrieval of words (primary task) is crucial also in the trials including WM load, but the words would not count if the exact recall of digits (secondary task) is not correct. Furthermore, a measure indicating the change in retrieval performance due to WM load—that is, maintenance cost (ΔRT high load – no load)—was computed for each participant (high values indicate more impaired retrieval performance due to WM load). Participants tried several practice trials for all retrieval and load conditions before the main experiment.

Semantic retrieval under working-memory load. The figure depicts an example trial where participants continuously retrieved words under a concurrent WM load (six digits). Each trial started with an alerting signal, which was followed by a WM load manipulation (4 s for the low-load condition including three digits, or 8 s for the high-load condition including 6 digits; note that the procedures and instructions indicated in blue color were not present in the condition with no load). After the encoding of the digits, participants performed the primary retrieval task (associative or dissociative retrieval condition) while maintaining the digits in memory for 20 s. At the end of the retrieval task, participants were requested to recall the remembered digits in the correct order. (Color figure online)
Working-memory capacity assessment
Before the main experimental procedure, participants were assessed for WMC using two distinct tasks. In the backward digit span task (D-Span), participants were instructed to remember a series consisting of unique digits (from 0 to 9) and recall them in the reverse order. In the operation span (O-Span), participants remembered a series of unique consonants and recall them in the same order. However, the presentation of each consonant was followed by a simple equation (e.g., 17 + 11 = 32), for which participants had to indicate whether it is correct or not using a key press. Both WM tasks started with a few practice trials at a span set to two. The practice was then followed by the main task. In each trial, the digits/letters were presented for 400 ms, intermitted by 250 ms intervals (ISI = 650 ms). The span was increased by 1 after each correct recall or decreased by 1 after an error. The main task continued until a total of 4 incorrect recalls were made. The overall capacity in the respective task was calculated as the average span of the four incorrect trials minus 1. Notably, the capacity measures for D-Span as well as O-Span showed high reliability, which was estimated using McDonald’s ω (ω = .872 and .863, respectively). The performance scores in the two tasks were summed to indicate the overall WMC of each individual.
Statistical analysis
The data were processed in R (R Core Team, 2021) and JASP (JASP Team, 2022). Retrieval responses with large RTs (>15 s; <0.2% of responses) as well as incorrect responses (indicated by two independent raters; <4% of responses) were removed prior to statistical analyses. Subsequently, the retained RT values were winsorized (10% quantile two-sided trimming) and averaged for each individual, separately by the experimental conditions. Semantic memory retrieval performance (RT) was then modeled using a 2 × 3 repeated-measures (RM) ANOVA, including the main effect of retrieval (associative, dissociative), WM load (no, low, high), and their interaction (Greenhouse–Geisser sphericity correction was applied where appropriate). Simple main effects for WM load (i.e., separately for the associative and dissociative retrieval task) were also assessed. Effect sizes for the RM ANOVA were estimated using \({\eta}_p^2\).
In addition to the omnibus RM ANOVAs, a custom contrast reflecting the hypothesized interaction effect was specified and then tested (for more details, see Table S1 in the Supplementary file). Then, the generalized order-restricted information criterion approximation (GORICA) approach was used to comprehensively assess the specified informative hypothesis (i.e., that the maintenance cost is larger in dissociative versus associative condition) against the null model (i.e., that the maintenance cost is not different between the retrieval conditions; for more details, see Supplementary file). Furthermore, Bayes factor (BF) was computed to complement the respective frequentists statistical tests. Using the default JASP priors, BF10 was assessed to indicate the strength of evidence favoring the alternative hypothesis (H1) against the null hypothesis (H0), as well as to evaluate how likely a null finding reflects evidence of absence rather than absence of evidence (i.e., lack of statistical power). Finally, the measures from the first experiment were subjected to a robust analysis of association using the percentage bend correlation (rpb) that takes into account the overall structure of the data and protects the estimations against marginal distribution outliers (Mair and Wilcox, 2019).
Results
Working memory load
The RM ANOVA for retrieval RTs revealed significant effects of retrieval, F(1, 60) = 90.97, p < .001, \({\eta}_p^2\)= .603, and WM load, F(2, 120) = 29.55, p < .001, \({\eta}_p^2\)= .330, whereas their interaction did not reach the significance level, F(1.82, 109.2) = 3.02, p = .058, \({\eta}_p^2\)= .048 (see Fig. 2A). The complementary Bayesian RM ANOVA indicated very strong evidence for the effect of retrieval, BF10 = 6.44×1010, as well as WM load, BF10 = 9.65×106, but inconclusive evidence regarding their interaction, BF10 = 1.17. Overall, the RTs were substantially longer in the dissociative than in the associative condition and increased linearly as a function of WM load in both retrieval conditions. However, the analysis of simple main effects showed that the effect of WM load was considerably stronger in the dissociative retrieval, F(2, 120) = 20.89, p < .001, \({\eta}_p^2\)= .258, and BF10 = 6.69×105, compared with the associative retrieval, F(2, 120) = 7.569, p < .001, \({\eta}_p^2\)= .112, and BF10 = 30.90 (see Fig. 2B).

Retrieval latency under working memory load. A The main effects WM load on associative and dissociative retrieval. B Maintenance cost in the retrieval conditions. C Inhibition cost across the WM load condition. Note. Increasing WM load resulted in a linear prolongation of RT across both retrieval conditions, but the estimated maintenance cost (ΔRT high load – no load) was larger for the dissociative than the associative task. Relatedly, the inhibition cost (ΔRT dissociative – associative) was significantly higher under the high-load as compared with the no-load condition. Error bars represent ± SE. ***p < .001, **p < .01, *p < .05
More importantly, the direct test for the specific hypothesis (a priori defined custom contrast that the maintenance cost is higher in the dissociative than in the associative retrieval condition) was statistically significant, t(120) = 2.451, p = .016, confirming this expected pattern. Notably, the maintenance cost in the dissociative retrieval was larger by 0.370 s, 95% CI [0.071,0.670], and thus roughly twice as large as the cost in the associative condition (see Fig. 2B). Moreover, the GORICA analysis indicated that the proposed custom hypothesis predicted the data approximately 35 times better than the null hypothesis (i.e., that the maintenance costs is not statistically different between the two retrieval conditions; see Supplementary Table S2 for more details).
Relatedly, the inhibition cost (i.e., ΔRT dissociative – associative) showed an upward trend with increasing load, whereas the inhibition cost was significantly higher under high load as compared with the no-load condition, one-tailed t(60) = 2.847, p = .003, d = 0.37 (see Fig. 2C). The Bayesian paired-sample t test indicated strong evidence for the difference in the inhibition cost under high load versus no load, BF10 = 10.77. Relatedly, the GORICA analysis indicated that the hypothesis of linearly increasing inhibition cost due to WM load predicted the data approximately 29 times better than the null hypothesis (i.e., that the inhibition cost does not differ as a function of WM load; see Supplementary Table S3 for more details).
Finally, the performance in the secondary WM task showed a ceiling effect (average recall accuracy >90%) and was not significantly different between the associative and dissociative condition, two-tailed t(60) = 0.707, p = .759, d = 0.09, BF10 = 0.178.
Correlations
Exploratory robust correlation analysis showed that WMC (the composite score from D-Span and O-Span task) was negatively correlated with retrieval latency across all conditions (overall rpb = −.264). However, only the correlation between WMC and associative retrieval without WM load remained statistically significant after applying Holm correction (rpb = −.431, p = .003). This correlation was significantly stronger compared with the corresponding correlation between WMC and dissociative retrieval at no WM load (rpb = −.133, p = .311; Δrpb = −.298, p = .020; see Table S4 in the Supplementary Information file for more details). Furthermore, WMC was not significantly related to inhibition cost under no WM load. Finally, at high WM load, inhibition cost correlated more strongly with maintenance cost in the dissociative (rpb = .578, p < .001) than in the associative condition (rpb = −.101, p = .880; Δrpb = −.679, p < .001; see Table S5 in the Supplementary Information file for more details).
Discussion
The first experiment addressed the role of WM in semantic cognition. For this purpose, we manipulated WM load using a dual-task interference paradigm while participants simultaneously carried out retrieval tasks employing either automatic (associative) or controlled (dissociative) semantic processes. We hypothesized that WM load would disrupt both retrieval processes but expected a larger impairment in the dissociative task, as it engages controlled resources pertaining to inhibition (Allen et al., 2008; Collette et al., 2001). This additional demand (i.e., the inhibition cost) was reflected in our data showing substantially longer latencies for dissociative than associative conditions, which concurs with our previous findings (Marko, Cimrová, et al., 2019a; Marko, Michalko, et al., 2019b; Marko & Riečanský, 2021b). In line with our hypothesis, loading verbal WM slowed down retrieval requiring associative as well as dissociative responding. This finding converges with previous studies on verbal fluency (Rende et al., 2002; Rosen & Engle, 1997; Unsworth et al., 2013) but importantly extends those findings by showing that working memory is engaged in both the automatic and the controlled mode of semantic retrieval. Notably, the direct tests for the hypothesis (custom contrast and informative hypothesis) indicated that WM load disrupted controlled (dissociative) retrieval more than the free-associative retrieval, which was also supported by the related finding that increasing WM load prolonged the time required for inhibition. These findings suggest that WM supports a process that is shared by both retrieval tasks, such as maintaining task goals or semantic cues/features of the stimuli, which facilitates the search for suitable candidates (either semantically related or unrelated). An intriguing account for such a utility would be that, by sustaining and updating retrieval cues and search criteria, WM regulates (amplifies and attenuates) the activations within the semantic network, shaping semantic search according to the goals (cues, features) it actively maintains (Diamond, 2013). However, this WM-mediated regulation of semantic activation seems to be more relevant when individuals attempt to disentangle from the currently active set of retrieval candidates (i.e., during the dissociative retrieval), as suggested by the elevated inhibition cost under high WM load (Fig. 2C). Notably, such an account is consistent both with the so-called cue-maintenance (Abwender et al., 2001; Hills et al., 2013; Hills et al., 2015) as well as the global-slowing (Mayr & Kliegl, 2000; Ulrich Mayr, 2002) models of verbal fluency, positing that the maintenance and dynamic updating of search criterion is required for rapid retrieval and optimal exhaustion of information in semantic memory. In line with these theoretical models, our experiment suggests that disturbing WM makes semantic retrieval less goal-oriented/focused, resulting in slowed initiation of likely associates and even poorer control over inappropriate responses.
The exploratory analyses indicated that individual WMC and retrieval latency are overall negatively correlated. Interestingly, however, a closer look suggested that this coupling was considerably stronger in the associative condition without any load as compared with other retrieval conditions. This is in line with the idea that WM facilitates the search through associative memory, but as WM store becomes unavailable (due to the load), memory retrieval can no longer benefit from this domain-general capacity and is therefore slowed (Hills et al., 2013; Rosen & Engle, 1997; Unsworth et al., 2013). Since this effect was present only in the associative condition, one could speculate that the means via which WM supports dissociative retrieval (or inhibition) may be of a different kind. Although we cannot explicitly evaluate this hypothesis based on the present data, we found that the slowing of dissociative performance due to WM load (i.e., maintenance cost) was coupled with inhibition cost, particularly under high WM load. This may suggest that individuals employ the limited WM resources in the dissociative task to prevent uncontrollable triggering of automatic but inappropriate associates (Baror & Bar, 2016; Diamond, 2013). Further research is needed to address and critically evaluate this tentative explanation formulated upon the exploratory correlation analyses, and to inspect whether WM interacts with automatic (associative) and inhibitory (dissociative) semantic processing via the same or distinct mechanism.
Experiment 2: Executive attention load
The second experiment addressed the role of EA in semantic retrieval. To this aim, we introduced two distinct manipulations using dual-task interference to target attentional monitoring and switching. The manipulation of attentional monitoring required participants to perform a (secondary) letter-matching task during the (primary) semantic retrieval task. The monitoring task was selected since attentional monitoring represents an essential EA function of the general-purpose control system (Baddeley, 2012; Diamond, 2013), which has been considered important for semantic memory retrieval (Marko et al., 2022). In particular, monitoring has been implied to regulate semantic search and response output to prevent errors and repetitions (Unsworth et al., 2011), which we hypothesize to be more taxed during the dissociative than the free–associative retrieval task.
The manipulation of attentional switching required participants to flexibly alternate between the retrieval rules and semantic sets. The attentional switching was selected following the previous studies, which regard switching a core EA function (Friedman & Miyake, 2017; Miyake et al., 2000), and because switching has been postulated a principal process involved in semantic search (e.g., Troyer et al., 1998). Notably, we assume that flexible switching may be particularly important during the dissociative task, which requires individuals to shift between remote semantic clusters encoded in the memory to generate sufficiently remote or unrelated responses (Hills et al., 2012; Marko & Riečanský, 2021b).
We expected that the performance in both retrieval tasks would be disrupted by the attentional loads, but the controlled (dissociative) retrieval would be more affected than the automatic (free-associative) retrieval. More specifically, our prediction was that, if dissociative retrieval exhausts more executive attentional resources than associative retrieval, then either the monitoring load effect should be larger in the dissociative task than the associative task, or the monitoring performance in the secondary task should be worse during the dissociative retrieval than the associative retrieval. Furthermore, using correlation analysis, we also explored the relationship between the retrieval measures and performance in tasks assessing individual domain-general EA (Stroop interference and Response interference).
Methods
Semantic retrieval measures
Semantic retrieval was assessed using the same tasks involving continuous production of associative and dissociative responses, as described in Experiment 1. However, specifically for this experiment, 12 trials (chains) lasting 50 seconds each were utilized. Three distinct attentional conditions were included across the trials in random order: four trials involved a concurrent monitoring task (monitoring load), another four trials involved a rapid switching between the two retrieval rules (switching load), and the remaining four trials involved no secondary attentional load (no load). Participants tried several practice trials for all retrieval and load conditions before the main experiment. The split-half reliability for the associative and the dissociative retrieval measure was sufficiently high (r = .761 and .861, respectively).
Monitoring load manipulation
The monitoring load was implemented in four (two associative and two dissociative) retrieval trials using a secondary task, in which the participants had to continuously monitor a changing stimulus presented concurrently with the primary retrieval task. In this secondary task, centered closely below the line where the retrieved words were typed, two stimuli were displayed next to each other: a fixed (target) stimulus and a changing probe. The target stimulus was a letter (either L, T, or F) in color (either blue or yellow), which was randomly generated at the beginning of each 50-second trial and remained the same (fixed) until the end of the trial. The probe was also a single letter, however, changing its identity (L, T, or F) and/or color (blue or yellow) each 900 ms. The participants were instructed to press a keyboard button each time the probe changed so that it matched the target (both the letter identity and color) as quickly as possible (hits), but not to respond when the two stimuli were not the same (false alarms). The probes were presented continuously during the primary retrieval task in blocks of six (i.e., all combinations of letters and colors) so there were > 50 probes (~8–9 of them that matched the target) in each chain (see Fig. 3A for a graphical description of the monitoring dual-task situation). The participants were reminded that a rapid retrieval of words (primary task) is crucial in the trials including the secondary task, but the words would not count if their monitoring performance were low (monitoring performance was analyzed as a secondary dependent measure).

Semantic retrieval under attention load. A An example trial including monitoring load. Participants continuously retrieved either word associations (depicted) or dissociations for 50 s (primary task) and simultaneously monitored for matches between the letters’ identity and color (secondary task). B An example trial including switching load. Participants continuously retrieved words for 50 s (primary task) but switched the retrieval rule (i.e., from associative to dissociative and vice versa) after each response (secondary task). Note. Procedures indicated in blue color were not present in the condition with no load. (Color figure online)
Switching load manipulation
The switching load was implemented in 4 trials using a secondary task that involved rapid switching between retrieval rules. In contrast to the other conditions where the rule was fixed within each chain, the switching trials required the participants to deliver associations and dissociations in alternation—that is, changing the retrieval rule after each retrieved word (e.g., “Phone–Call–Banana–Monkey…”; see Fig. 3B for a graphical description of the switching-task situation). Note that the switching (associative–dissociative) chains did not include monitoring load as the combination of both attentional loads was overly difficult, as revealed in a pilot testing.
The assessment of executive attention capacity
Before the main experimental procedure including attentional loads, the participant’s capacity of executive attention was assessed using the Stroop test and a response inhibition task. The Stroop test included three blocks of trials with distinct conditions: (1) in the block with neutral trials (N = 84), participants were presented with a series of “XXXXX” strings in color (red, green, blue, or yellow); (2) in congruent trials (N = 48), participants saw a series of color names presented in the corresponding font color; (3) in incongruent trials (N = 48), participants saw a series of color names presented in mismatching font colors (the mismatching name–font pairs were counterbalanced). In all conditions, participants were instructed to respond with a key press corresponding with the stimulus font color as quickly and accurately as possible. Participants practiced on 12 trials for each condition before the main task. The split-half reliability of the performance in the task was high for the congruent condition (r = .880) as well as the incongruent condition (r = .913). The difference in RT between the incongruent and congruent trials was assessed as a domain-general measure of selective attention and interference control. This measure also showed high split-half reliability (r = .740). In the response inhibition task, participants were presented with a series of arrows pointing in four distinct directions. The task included two types of trials presented in random order. In the congruent trials (N = 192), the arrow was presented in white color, indicating that participants were required to press a keyboard arrow in the corresponding direction (i.e., left, right, up, down). In the incongruent trials (N = 96), the arrow was red, indicating that participants were required to press a keyboard arrow in the opposite direction (i.e., to press the left keyboard arrow when seeing a rightward stimulus). In both conditions, participants were required to respond as quickly and accurately as possible. A short practice including 12 trials for each condition was introduced before the main task. The split-half reliability of the performance in the task was high for the congruent condition (r = .979) as well as the incongruent condition (r = .983). The difference in RT between the incongruent and congruent trials was assessed as a domain-general measure of response inhibition. This estimate was also sufficiently reliable (r = .830).
Statistical analysis
The data were processed and analyzed following the same procedures as in the first experiment. In this case, however, semantic retrieval (RT) was modeled using two separate 2 × 2 RM ANOVAs (i.e., the two attentional load effects were analyzed separately as they represent qualitatively distinct manipulations). The first ANOVA included the main effect of retrieval (associative, dissociative) and monitoring load (no load, monitoring), and their interaction. The second ANOVA included the switching load factor (no load, switching) instead. Simple main effects for both loads were also assessed. Notably, the performance in the secondary monitoring task was assessed using the sensitivity measure (d′) with the log-linear correction (as recommended by Hautus, 1995). Finally, the retrieval measures from the second experiment and tasks assessing executive attention capacity (interference control) were subjected to analysis of association using the percentage bend correlation (rpb). Further details regarding the custom contrasts and GORICA analysis of the interaction effects are provided in the Supplementary file (Tables S6–S9).
Results
Monitoring load
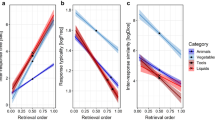
For monitoring load, the RM ANOVA revealed significant effects of retrieval, F(1, 61) = 127.99, p < .001, \({\eta}_p^2\)= .677, and monitoring load, F(1, 61) = 123.36, p < .001, \({\eta}_p^2\)= .669, but their interaction was not statistically significant, F(1, 61) = 0.52, p = .476, \({\eta}_p^2\)= .001. The complementary Bayesian RM ANOVA indicated very strong evidence for the respective main effects, BF10 = 5.72×1013 and BF10 = 9.20×1011, but moderate evidence for absence of the interaction effect, BF01 = 3.90. Overall, the retrieval latency was longer in the dissociative than in the associative condition, while both retrieval conditions were further slowed by the concurrent monitoring task (see Fig. 4A). As suggested by the absence of interaction, the simple main effect of monitoring load was similar in both retrieval conditions [for associative retrieval: F(1, 61) = 54.30, p < .001, \({\eta}_p^2\)= .471, and the related BF10 = 2.87×107; for dissociative retrieval: F(1, 61) = 58.50, p < .001, \({\eta}_p^2\)= .490, and BF10 = 3.43×107].

Retrieval latency under attentional load. A The main effects of monitoring load on associative and dissociative retrieval. B Monitoring accuracy in associative versus dissociative retrieval. C The effects of switching load on associative and dissociative retrieval. Note. Monitoring load affected both retrieval types to the same extent, but the monitoring performance (d′) was significantly worse during the dissociative than associative retrieval condition. Switching load significantly modulated only the dissociative retrieval. Error bars represent ± SE. ***p < .001, ns = nonsignificant
A direct test of the informative hypothesis (a priori defined custom contrast that the monitoring cost is higher in the dissociative than in the associative condition) was not statistically significant, t(61) = 0.718, p = .476, further confirming that the secondary monitoring task affected both retrieval conditions in a similar fashion. In line with this result, the GORICA analysis indicated that the proposed hypothesis predicted the data approximately 1.27 times worse than the null hypothesis (i.e., that maintenance effect is not statistically different between the two retrieval conditions; see Supplementary Table S7 for more details). Likewise, the Bayesian paired-sample t test indicated moderate evidence against the difference in the monitoring cost between the retrieval conditions, BF01 = 3.71.
Finally, we evaluated the attentional monitoring performance (sensitivity) in the secondary monitoring task. This analysis revealed that the monitoring performance was considerably worse during the dissociative (d′ = 1.73) than the associative (d′ = 2.12) retrieval, t(62) = 5.13, p < .001, d = 0.646 (see Fig. 4B). The corresponding Bayesian paired-sample t test indicated very strong evidence for the difference, BF10 = 5.39×103.
Switching load
For switching load, the RM ANOVA revealed significant effects of retrieval, F(1, 61) = 123.22, p < .001, \({\eta}_p^2\)= .669, and switching load, F(1, 61) = 23.14, p < .001, \({\eta}_p^2\)= .275, and their interaction, F(1, 61) = 10.07, p = .002, \({\eta}_p^2\)= .142. The corresponding Bayesian RM ANOVA indicated very strong evidence for the effect of retrieval, BF10 = 2.31×1013, switching load, BF10 = 262.53, as well as their interaction, BF10 = 31.40. Furthermore, the analysis of simple main effects showed that switching load significantly altered the dissociative retrieval, F(1, 61) = 22.91, p < .001, \({\eta}_p^2\) = .273, and BF10 = 1373.6, but not the associative retrieval, F(1, 61) = 2.14, p = .149, \({\eta}_p^2\)= .034, and BF10 = 0.479 (see Fig. 4C). Notably, the switching cost in the dissociative condition was 0.500 s, 95% CI [0.324,0.675], whereas in the associative condition it was only 0.101 s, 95% CI [−0.074, 0.277].
A direct test of the hypothesis (a priori defined custom contrast that the switching cost is higher in the dissociative than in the associative condition) was statistically significant, t(61) = 3.173, p = .002, further confirming the hypothesis. In line with that, the GORICA analysis indicated that our informative hypothesis predicted the data approximately 93 times better than the null hypothesis (see Supplementary Table S9 for more details). Likewise, the Bayesian paired-sample t test indicated strong evidence for the difference in the switching cost between the retrieval conditions, BF01 = 24.53.
Executive attention measures
For the Stroop task and the response inhibition task, RM ANOVAs revealed significant main effects of condition (congruent versus incongruent), F(1, 61) = 116.13, p < .001, \({\eta}_p^2\)= .656 and F(1, 61) = 293.48, p < .001, \({\eta}_p^2\)= .826, respectively. As expected, the incongruent condition was associated with longer latencies as compared with the congruent condition in both tasks. These robust effects were also confirmed by the corresponding Bayesian analysis, which indicated very strong evidence in favor of the differences, BF01 > 1.38×1013.
Correlations
Finally, an exploratory robust correlation analysis was performed to inform on the relationships between the retrieval and the interference measures. Overall, the analysis showed weak positive associations between the retrieval measures (across all conditions) and the Stroop interference (overall rpb = −.192) as well as Response interference (overall rpb = −.267; see Table S10 in the Supplementary Information file for more details). Furthermore, the correlations between retrieval control measures (inhibition cost and switching cost) and the two interference measures were not statistically significant (|rpb| < .152, pholm > .950). Finally, the exploratory analysis also showed a moderately strong negative correlation between the switching cost (the RT difference between the dissociative retrieval under switching load versus no load) and inhibition cost (rpb = −.413 p < .001; see Fig. 1S in the Supplementary Information file).
Discussion
In the second experiment, we investigated the role of EA functions in semantic retrieval. We hypothesized that EA, i.e., monitoring and switching, plays a role in semantic retrieval. Thus, we predicted that attentional load would impair retrieval performance, especially in the dissociative task that requires control over habitual associates (Allen et al., 2008; Collette et al., 2001). As in the first experiment, dissociative production was substantially slower than associative production, reflecting such demands for control (i.e., inhibition cost). In line with our hypothesis, loading attention using the secondary monitoring or switching task slowed down semantic retrieval. Importantly, however, monitoring load impaired both retrieval tasks in a similar fashion, whereas switching load disrupted only the dissociative performance. These results thus indicate that the respective attentional manipulations modulated semantic retrieval via (at least partially) distinct mechanisms.
The uniform impairment of associative and dissociative retrieval by monitoring load corroborates the view that a generic attentional mechanism supports a wide range of computations (Friedman & Miyake, 2017; Hartwigsen, 2018), including the semantic ones. This finding is in line with the current models of semantic search (Mayr & Kliegl, 2000; Ulrich Mayr, 2002) suggesting that executive ability facilitates the overall rate of retrieval independently from the specific semantic processes involved in the two retrieval tasks. Since dual-task interference is known to deplete the executive capacity (Szameitat et al., 2002),Footnote 1 the impaired performance can be explained by a decreased availability of the limited resources that support rapid retrieval (i.e., executive bottleneck). Alternatively, however, impaired performance in dual-task paradigms has also been attributed to a bottleneck at the response selection level (rather than at the executive level; Hegarty et al., 2000)—that is, as both the primary (retrieval) and the secondary (letter matching) task require rapid responding, the slowing of retrieval need not necessarily result from the depleted executive capacity but could be caused by the inability of participants to select appropriate responses for both task at once. However, against this explanation is our finding that more errors were committed during the dissociative than the associative retrieval (see Fig. 4B), especially when considering that there were fewer dissociative than associative responses. The impaired monitoring performance during dissociative retrieval thus likely stems from a strategic trade-off (Hegarty et al., 2000) when participants dedicate maximum processing resources to one task (the word retrieval) at the expense of the other, usually less demanding task (the letter matching). Taken together, the robust difference in monitoring performance between the retrieval tasks indicates that producing dissociates exhausts more executive attentional resources than delivering associates. Given the nature of the dissociative task, it could be concluded that attentional monitoring is engaged to a higher extent when facing the need to evaluate and suppress inappropriate (habitual) associates.
Interestingly, the switching load substantially impaired the dissociative performance only. Even though the associative retrieval was also slightly slowed down, this effect was weak and unreliable. A domain-general rule shifting (Diamond, 2013; Ravizza & Carter, 2008) is not a likely account for the specific impairment of the dissociative retrieval since both dissociative and associative responding involved the same number of switches, thus imposing equal demands on such capacity.
Alternatively, the nonuniform effect of switching load could be explained, at least in part, by the conflict-monitoring model proposed by Botvinick et al. (2004). Since free-associative trials do not involve conflict, the recruitment of cognitive control in the subsequent dissociative trial may be decreased, leading to prolonged dissociative latency. Notably, in dissociative chains without switching, the continuous retrieval of unrelated responses may keep the state of cognitive control tonically upregulated, since all trials involve conflict. Such sustained involvement of cognitive control could aid the performance, leading to shorter dissociative latencies in the non-switching than in the switching dissociative condition (see Fig. 4C). Nevertheless, the conflict-monitoring hypothesis predicts that the state of cognitive control varies depending on the level of conflict activation on preceding trials (Botvinick et al., 2001; but also see Schmidt, 2019). Therefore, under the switching load, the conflict in dissociative trials should upregulate the cognitive control and deliberative processing, hence prolonging the latency on the subsequent associative trials. Although a subtle trend in the predicted direction was observed (see Fig. 4C), the associative slowing due to switching was minor and not statistically reliable.
Furthermore, the dissociative retrieval slowing due to switching could also (or alternatively) be due to a residual semantic activation from the preceding trials. Delivering an associated word response (e.g., cat [stimulus] → dog [response]) induces semantic activation within the connected parts of the conceptual network. So activated conceptual associates (e.g., animal, fur, barking, etc.) may carry over to the subsequent dissociative trial (Goschke, 2000), hence inducing a proactive cognitive interference slowing the performance (i.e., the pre-activated concepts in semantic memory impair the ability to find an unrelated response). Notably, the attempt to retrieve an unrelated response (e.g., dog → stone) also activates relevant features/associates within the conceptual network (e.g., rock, weight, wall, etc.), but these activations do not interfere with the subsequent associative task (e.g., stone → rock). This residual semantic activation account can explain the asymmetric switching cost, predicting a slowing of dissociative but not associative latencies (as shown in Fig. 4C). Nevertheless, since both conflict-monitoring (domain-general mechanism) and residual semantic activation (domain-specific mechanism) are in fact consistent with the present findings, experiments to directly test these alternative explanations are warranted.
Finally, the second experiment also showed that the processing costs associated with switching and inhibition are negatively correlated. Together with the selective impairment of dissociative retrieval by switching load, this evidence indicates that flexible transitions between semantic sets (switching) and constraining semantic activation (inhibition) may represent two diverging or interfering modes of controlled semantic search. Notably, such a tradeoff between (restricting) inhibition versus (flexible) switching (Marko & Riečanský, 2021b) may not necessarily arise from a competition between two distinct neurocognitive systems (or their limited resources) but simply because these two functions operate efficiently at different sets of parameters within the same cognitive or neural architecture (Friedman & Miyake, 2017; Marko & Riečanský, 2021a).
Summary and conclusion
Two experiments implementing the dual-task and task-switching interference approach were conducted to investigate the role of domain-general executive capacities in semantic memory retrieval. We employed novel generative tasks, enabling us to differentiate between the automatic (associative) and the controlled (dissociative) retrieval processes, and so elucidate how they interact with WM and EA. From a broader perspective, our results conform with the hypothesis that domain-general executive functions support semantic memory retrieval, which largely concurs with the prevailing models of verbal fluency (Mayr & Kliegl, 2000; Ulrich Mayr, 2002; Unsworth et al., 2012). Nevertheless, the current findings critically extend the previous studies, showing that distinct forms of interference (loads) disrupt automatic and controlled retrieval in a non-unitary way, which provides a deeper insight into the putative mechanisms that govern these interactions.
We observed that WM load impaired both forms of retrieval, suggesting a generic mechanism of action. Generally, it is agreed that representations maintained in WM become globally available for other mental processes (Baars et al., 2013; Kamiński & Rutishauser, 2020), which amplifies the activation of related information encoded in long-term memory and enhances the strategic access to the relevant knowledge (Belke, 2008; Cowan et al., 2012; Unsworth et al., 2013). Thus, a likely account for the current findings is that depleting WM hinders the representation and depth of processing of word stimuli (e.g., their semantic features and relations), rendering semantic retrieval less focused and goal-oriented. Yet, a more detailed analyses regarding the combined effect of load and retrieval type revealed that the time required to suppress habitual but inappropriate responses (inhibition cost) was prolonged with increasing WM load, and relatedly, the retrieval slowing due to WM load (maintenance cost) was considerably higher (approximately twice as large) in the dissociative than in the associative retrieval condition. Thus, together with the model comparison (GORICA and Bayesian) approach, these lines of evidence support the hypothesis that the WM resources are relatively more important for the control of semantic memory retrieval than for the free-associative ability.
Furthermore, monitoring load disrupted both modes of semantic memory retrieval performance by a comparable margin, suggesting that the free associative and controlled retrieval involve a common domain-general attentional mechanism. However, we further revealed that the secondary monitoring performance was substantially worse when individuals generated dissociates than associates, supporting the hypothesis that controlled retrieval involving the suppression of automatic associates is particularly taxing the limited attentional resources. Notably, since our retrieval measures do not manifest the clustering-switching pattern typical for category fluency tasks (Troyer et al., 1997), we can assume that the proposed contribution of executive attention in semantic memory retrieval goes beyond the ability to initiate switches as proposed by Troyer and colleagues (Hirshorn & Thompson-Schill, 2006; Troyer et al., 1998).
Notably, although maintenance and monitoring loads slowed down retrieval fluency in the same direction and similar degree, one should be careful when comparing these two interventions since they differed not only in the underlying process being loaded (i.e., WMC versus attentional monitoring) but also in the modality of the secondary task (i.e., verbal versus visual stimuli). Since both the primary (retrieval) and secondary (maintenance) tasks in Experiment 1 involved the processing of verbal material, one could expect stronger interference, than in Experiment 2, where the primary and secondary monitoring tasks were of different modalities. Further studies are needed to address the possible sources of interference, both at the level of underlying processes and modalities. These studies should also assess which strategies (e.g., subvocal rehearsal or mental imagery) individuals employ in such dual-task settings.
The involvement of executive attention in controlled retrieval was further supported using the switching load, which considerably impaired only the dissociative but not the associative task. This asymmetry could be explained by either (1) varying demands these two tasks exert on a domain-general mechanisms that is responsible for conflict monitoring and implementation of cognitive control (i.e., conflict resolution) or (2) by residual activation in the conceptual networks that carries over to the subsequent trials and so interferes with the domain-specific inhibitory mechanisms. Finally, we would like to note that only the switching load included mixed-task chains (i.e., mixing associative and dissociative trials within the same chain), whereas under the other forms of load individuals retrieved either related or unrelated concepts - i.e., without mixing these two retrieval conditions in the same chain. Although such mixed-tasks design is a well-established switching paradigm in cognitive science (see Kiesel et al., 2010), the comparison between the qualitatively distinct treatments (loads) in this study should be made with caution.
In conclusion, we found that exploiting domain-general cognitive abilities using concurrent cognitive loads substantially impairs both the automatic–associative and the controlled–dissociative retrieval. Yet, importantly, a more detailed analysis revealed that while WM and EA monitoring support the access to both related and unrelated conceptual representations, these domain-general capacities are particularly taxed when one needs to disentangle from the currently activated set of concepts in semantic memory. Although the precise nature of this mechanism awaits further investigation, we propose that WM and EA are functionally involved in the adaptive gating of stimulus-driven (i.e., automatic) semantic activation that spreads within the representational network and/or resolving the interference from among competing response candidates, through which these domain-general capacities bias the retrieval towards goal-appropriate representations. More generally, our results also suggest that EA may instantiate a mechanism for a fine-grained regulation between a constraining (inhibitory) versus a flexible (switching) mode of semantic search and retrieval. Further experiments utilizing semantic-specific interference may shed more light on the mechanisms governing efficient retrieval of knowledge and flexible thinking. Measures focusing on qualitative aspects of retrieved responses may also represent a promising way to extend the findings provided in the current study.
Data availability
The data and materials are openly available in the OSF repository (https://osf.io/9v5gb/), and none of the experiments was preregistered.
Notes
In dual-task paradigms, EA is recruited to compensate for overall higher processing demands, coordinate the processing employed in the concurrent tasks, and resolve their mutual interference. The exertion of such attentional control is reflected in higher activation within the large-scale executive brain network that includes bilateral dorsal-lateral and parietal regions.
References
Abwender, D. A., Swan, J. G., Bowerman, J. T., & Connolly, S. W. (2001). Qualitative analysis of verbal fluency output: Review and comparison of several scoring methods. Assessment, 8(3), 323–336. https://doi.org/10.1177/107319110100800308
Allen, P., Mechelli, A., Stephan, K. E., Day, F., Dalton, J., Williams, S., & McGuire, P. K. (2008). Fronto-temporal interactions during overt verbal initiation and suppression. Journal of Cognitive Neuroscience, 20(9), 1656–1669. https://doi.org/10.1162/jocn.2008.20107
Baars, B. J., Franklin, S., & Ramsoy, T. Z. (2013). Global workspace dynamics: Cortical “binding and propagation” enables conscious contents. Frontiers in Psychology, 4. https://doi.org/10.3389/fpsyg.2013.00200
Baddeley, A. (2012). Working memory: Theories, models, and controversies. Annual Review of Psychology, 63(1), 1–29. https://doi.org/10.1146/annurev-psych-120710-100422
Badre, D., & Wagner, A. D. (2007). Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia, 45(13), 2883–2901. https://doi.org/10.1016/j.neuropsychologia.2007.06.015
Baror, S., & Bar, M. (2016). Associative activation and its relation to exploration and exploitation in the brain. Psychological Science, 27(6), 776–789. https://doi.org/10.1177/0956797616634487
Belke, E. (2008). Effects of working memory load on lexical-semantic encoding in language production. Psychonomic Bulletin & Review, 15(2), 357–363. https://doi.org/10.3758/PBR.15.2.357
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652. https://doi.org/10.1037/0033-295X.108.3.624
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict monitoring and anterior cingulate cortex: an update. Trends in Cognitive Sciences, 8(12), 539–546. https://doi.org/10.1016/j.tics.2004.10.003
Chai, L. R., Mattar, M. G., Blank, I. A., Fedorenko, E., & Bassett, D. S. (2016). Functional network dynamics of the language system. Cerebral Cortex, 26(11), 4148–4159. https://doi.org/10.1093/cercor/bhw238
Collette, F., Van der Linden, M., Delfiore, G., Degueldre, C., Luxen, A., & Salmon, E. (2001). The functional anatomy of inhibition processes investigated with the Hayling task. NeuroImage, 14(2), 258–267. https://doi.org/10.1006/nimg.2001.0846
Cowan, N., Rouder, J. N., Blume, C. L., & Saults, J. S. (2012). Models of verbal working memory capacity: What does it take to make them work? Psychological Review, 119(3), 480–499. https://doi.org/10.1037/a0027791
Crowe, S. F. (1998). Decrease in performance on the verbal fluency test as a function of time: Evaluation in a young healthy sample. Journal of Clinical and Experimental Neuropsychology, 20(3), 391–401. https://doi.org/10.1076/jcen.20.3.391.810
Demetriou, E., & Holtzer, R. (2017). Mild cognitive impairments moderate the effect of time on verbal fluency performance. Journal of the International Neuropsychological Society, 23(1), 44–55. https://doi.org/10.1017/S1355617716000825
Diamond, A. (2013). Executive functions. Annual Review of Psychology, 64(1), 135–168. https://doi.org/10.1146/annurev-psych-113011-143750
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191. https://doi.org/10.3758/BF03193146
Fedorenko, E., & Thompson-Schill, S. L. (2014). Reworking the language network. Trends in Cognitive Sciences, 18(3), 120–126. https://doi.org/10.1016/j.tics.2013.12.006
Friedman, N. P., & Miyake, A. (2017). Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex, 86, 186–204. https://doi.org/10.1006/cogp.1999.0734
Goschke, T. (2000). Intentional reconfiguration and involuntary persistence in task set switching. In S. Monsell & J. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 331–355). MIT Press. https://doi.org/10.7551/mitpress/1481.003.0023
Hartwigsen, G. (2018). Flexible redistribution in cognitive networks. Trends in Cognitive Sciences, 22(8), 687–698. https://doi.org/10.1016/j.tics.2018.05.008
Hautus, M. J. (1995). Corrections for extreme proportions and their biasing effects on estimated values of d′. Behavior Research Methods, Instruments, & Computers, 27(1), 46–51. https://doi.org/10.3758/BF03203619
Hegarty, M., Shah, P., & Miyake, A. (2000). Constraints on using the dual-task methodology to specify the degree of central executive involvement in cognitive tasks. Memory & Cognition, 28(3), 376–385. https://doi.org/10.3758/BF03198553
Henry, J. D., & Crawford, J. R. (2004). A meta-analytic review of verbal fluency performance following focal cortical lesions. Neuropsychology, 18(2), 284–295. https://doi.org/10.1037/0894-4105.18.2.284
Hills, T. T., Jones, M. N., & Todd, P. M. (2012). Optimal foraging in semantic memory. Psychological Review, 119(2), 431–440. https://doi.org/10.1037/a0027373
Hills, T. T., Mata, R., Wilke, A., & Samanez-Larkin, G. R. (2013). Mechanisms of age-related decline in memory search across the adult life Span. Developmental Psychology, 49(12), 2396–2404. https://doi.org/10.1037/a0032272
Hills, T. T., Todd, P. M., & Jones, M. N. (2015). Foraging in semantic fields: How we search through memory. Topics in Cognitive Science, 7(3), 513–534. https://doi.org/10.1111/tops.12151
Hirshorn, E. A., & Thompson-Schill, S. L. (2006). Role of the left inferior frontal gyrus in covert word retrieval: Neural correlates of switching during verbal fluency. Neuropsychologia, 44(12), 2547–2557. https://doi.org/10.1016/j.neuropsychologia.2006.03.035
JASP Team. (2022). JASP version 0.16.4 Computer software. https://jasp-stats.org/
Jefferies, E., Thompson, H., Cornelissen, P., & Smallwood, J. (2020). The neurocognitive basis of knowledge about object identity and events: Dissociations reflect opposing effects of semantic coherence and control. Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1791), 20190300. https://doi.org/10.1098/rstb.2019.0300
Kamiński, J., & Rutishauser, U. (2020). Between persistently active and activity-silent frameworks: novel vistas on the cellular basis of working memory. Annals of the New York Academy of Sciences, 1464(1), 64–75. https://doi.org/10.1111/nyas.14213
Kane, M. J., & Engle, R. W. (2002). The role of prefrontal cortex in working-memory capacity, executive attention, and general fluid intelligence: An individual-differences perspective. Psychonomic Bulletin & Review, 9(4), 637–671. https://doi.org/10.3758/BF03196323
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—A review. Psychological Bulletin, 136(5), 849–874. https://doi.org/10.1037/a0019842
Lambon Ralph, M. A., Jefferies, E., Patterson, K., & Rogers, T. T. (2017). The neural and computational bases of semantic cognition. Nature Reviews Neuroscience, 18(1), 42–55. https://doi.org/10.1038/nrn.2016.150
Mair, P., & Wilcox, R. (2019). Robust statistical methods in R using the WRS2 package. Behavior Research Methods, 52, 464–488. https://doi.org/10.3758/s13428-019-01246-w
Marko, M., Cimrová, B., & Riečanský, I. (2019a). Neural theta oscillations support semantic memory retrieval. Scientific Reports, 9, 17667. https://doi.org/10.1038/s41598-019-53813-y
Marko, M., Michalko, D., Dragašek, J., Vančová, Z., Jarčušková, D., & Riečanský, I. (2022). Assessment of automatic and controlled retrieval using verbal fluency tasks. Assessment, 0(0). https://doi.org/10.1177/10731911221117512
Marko, M., Michalko, D., & Riečanský, I. (2019b). Remote associates test: An empirical proof of concept. Behavior Research Methods, 51(6), 2700–2711. https://doi.org/10.3758/s13428-018-1131-7
Marko, M., & Riečanský, I. (2021a). The left prefrontal cortex supports inhibitory processing during semantic memory retrieval. Cortex, 134, 296–306. https://doi.org/10.1016/j.cortex.2020.11.001
Marko, M., & Riečanský, I. (2021b). The structure of semantic representation shapes controlled semantic retrieval. Memory, 29(4), 538–546. https://doi.org/10.1080/09658211.2021.1906905
Marron, T. R., Berant, E., Axelrod, V., & Faust, M. (2020). Spontaneous cognition and its relationship to human creativity: A functional connectivity study involving a chain free association task. NeuroImage, 220, Article 117064. https://doi.org/10.1016/j.neuroimage.2020.117064
Mayr, U., & Kliegl, R. (2000). Complex semantic processing in old age: Does it stay or does it go? Psychology and Aging, 15(1), 29–43. https://doi.org/10.1037/0882-7974.15.1.29
Michalko, D., Marko, M., & Riečanský, I. (2022). Executive functioning moderates the decline of retrieval fluency in time. Psychological Research. https://doi.org/10.1007/s00426-022-01680-0
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D. (2000). The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cognitive Psychology, 41(1), 49–100. https://doi.org/10.1006/cogp.1999.0734
Ravizza, S. M., & Carter, C. S. (2008). Shifting set about task switching: Behavioral and neural evidence for distinct forms of cognitive flexibility. Neuropsychologia, 46(12), 2924–2935. https://doi.org/10.1016/j.neuropsychologia.2008.06.006
Rende, B., Ramsberger, G., & Miyake, A. (2002). Commonalities and differences in the working memory components underlying letter and category fluency tasks: A dual-task investigation. Neuropsychology, 16(3), 309–321. https://doi.org/10.1037/0894-4105.16.3.309
R Core Team. (2021). R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. https://www.R-project.org/
Rosen, V. M., & Engle, R. W. (1997). The role of working memory capacity in retrieval. Journal of Experimental Psychology: General, 126(3), 211–227. https://doi.org/10.1037/0096-3445.126.3.211
Schmidt, J. R. (2019). Evidence against conflict monitoring and adaptation: An updated review. Psychonomic Bulletin & Review, 26(3), 753–771. https://doi.org/10.3758/s13423-018-1520-z
Shao, Z., Janse, E., Visser, K., & Meyer, A. S. (2014). What do verbal fluency tasks measure? Predictors of verbal fluency performance in older adults. Frontiers in Psychology, 5, 772. https://doi.org/10.3389/fpsyg.2014.00772
Strobach, T., Wendt, M., & Janczyk, M. (2018). Editorial: Multitasking: Executive functioning in dual-task and task switching situations. Frontiers in Psychology, 9(FEB), 1–5. https://doi.org/10.3389/fpsyg.2018.00108
Szameitat, A. J., Schubert, T., Müller, K., & Von Yves Cramon, D. (2002). Localization of executive functions in dual-task performance with fMRI. Journal of Cognitive Neuroscience, 14(8), 1184–1199. https://doi.org/10.1162/089892902760807195
Troyer, A. K., Moscovitch, M., & Winocur, G. (1997). Clustering and switching as two components of verbal fluency: Evidence from younger and older healthy adults. Neuropsychology, 11(1), 138–146. https://doi.org/10.1037/0894-4105.11.1.138
Troyer, A. K., Moscovitch, M., Winocur, G., Alexander, M. P., & Stuss, D. (1998). Clustering and switching on verbal fluency: the effects of focal frontal- and temporal-lobe lesions. Neuropsychologia, 36(6), 499–504. https://doi.org/10.1016/S0028-3932(97)00152-8
Ulrich Mayr, S. (2002). On the dissociation between clustering and switching in verbal fluency: Comment on Troyer, Moscovitch, Winocur, Alexander and Stuss. Neuropsychologia, 40, 562–566.
Unsworth, N., Brewer, G. A., & Spillers, G. J. (2013). Working memory capacity and retrieval from long-term memory: the role of controlled search. Memory & Cognition, 41(2), 242–254. https://doi.org/10.3758/s13421-012-0261-x
Unsworth, N., Spillers, G. J., & Brewer, G. A. (2011). Variation in verbal fluency: A latent variable analysis of clustering, switching, and overall performance. Quarterly Journal of Experimental Psychology, 64(3), 447–466. https://doi.org/10.1080/17470218.2010.505292
Unsworth, N., Spillers, G. J., & Brewer, G. A. (2012). Working memory capacity and retrieval limitations from long-term memory: An examination of differences in accessibility. The Quarterly Journal of Experimental Psychology, 65(12), 2397–2410. https://doi.org/10.1080/17470218.2012.690438
Whiteside, D. M., Kealey, T., Semla, M., Luu, H., Rice, L., Basso, M. R., & Roper, B. (2016). Verbal fluency: Language or executive function measure? Applied Neuropsychology: Adult, 23(1), 29–34. https://doi.org/10.1080/23279095.2015.1004574
World Medical Association. (2013). World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA. Journal of the American Medical Association, 310(20), 2191–2194. https://doi.org/10.1001/jama.2013.281053
Funding
This work was supported by the Scientific Grant Agency of the Ministry of Education, Science, Research and Sport of the Slovak Republic (Project No. VEGA 2/0059/20) and the Slovak Research and Development Agency (Project No. APVV-19-0570).
Author information
Authors and Affiliations
Contributions
M.M. and I.R. wrote the manuscript; M.M. designed the experiment and analyzed the data. Both authors contributed to interpretation of the results and critical discussion. We would like to thank Andrej Zabnikar, Klára Horváthová, and Danijela Topić Vizcaya for help with data collection and data preparation.
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
The authors have no relevant financial or nonfinancial interests to disclose.
Consent and Ethics approval
The research was conducted in accordance with the Declaration of Helsinki and approved by the institutional review board. All procedures and methods were carried out in accordance with the relevant guidelines and regulations. All participants gave written informed consent and received a financial reward for their participation.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 42 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Marko, M., Riečanský, I. Differential effects of executive load on automatic versus controlled semantic memory retrieval. Mem Cogn 51, 1145–1158 (2023). https://doi.org/10.3758/s13421-022-01388-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13421-022-01388-x