
Abstract
In three experiments, we showed that animate entities are remembered better than inanimate entities. Experiment 1 revealed better recall for words denoting animate than inanimate items. Experiment 2 replicated this finding with the use of pictures. In Experiment 3, we found better recognition for animate than for inanimate words. Importantly, we also found a higher recall rate of “remember” than of “know” responses for animates, whereas the recall rates were similar for the two types of responses for inanimate items. This finding suggests that animacy enhances not only the quantity but also the quality of memory traces, through the recall of contextual details of previous experiences (i.e., episodic memory). Finally, in Experiment 4, we tested whether the animacy effect was due to animate items being richer in terms of sensory features than inanimate items. The findings provide further evidence for the functionalist view of memory championed by Nairne and coworkers (Nairne, 2010; Nairne & Pandeirada, Cognitive Psychology, 61:1–22, 2010a, 2010b).
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Several lines of evidence suggest that animate entities have a privileged processing status over inanimate objects—in other words, that animates have priority over inanimates. The animate–inanimate distinction parallels the distinction between “living” and “nonliving” things that has been postulated to account for selective deficits in patients (for a review, see Capitani, Laiacona, Mahon, & Caramazza, 2003). Animates belong to the general category of living things. Gelman and Spelke (1981) identified the following fundamental differences between animate and inanimate objects: (1) Animates can act, whereas inanimates move only when something/someone initiates the action; (2) animates grow and reproduce; (3) animates can know, perceive, emote, learn, and think; and (4) animates are made of biological structures that maintain life and allow reproduction.
In several domains of cognitive science, a growing body of evidence supports the view that animates are given processing priority over inanimates. Animate stimuli are thought to attract more attention than inanimates because it was important for the survival of our human ancestors to identify potentially dangerous entities quickly. To illustrate this, it has been shown that animate stimuli (e.g., animals, human faces) attract more attention than inanimate stimuli, not only those that arouse fear (Öhman, Flykt, & Esteves, 2001; Öhman, Lundqvist, & Esteves, 2001; but see Brosch & Sharma, 2005) but also neutral stimuli (Lipp, Derakshan, Waters, & Logies, 2004). It has been shown that individuals detect changes to humans and animals more quickly and accurately than changes to inanimate objects (New, Cosmides, & Tooby, 2007; see also Kirchner & Thorpe, 2006, for evidence of faster ocular saccades in response to animals than to other objects). Recently, Yang, Wang, Yan, Zhu, Chen, and Wang (2012) tracked participants’ eye movements while they viewed pictures with animals and inanimate images as focal objects. These pictures had either negative or neutral emotional valence, and either human body parts or nonhuman parts were near the focal objects (i.e., context). The picture’s valence, arousal, position, size, and most of the low-level visual features were controlled for across categories. Their findings showed that nonhuman animals were more likely to be attended to (and to be attended to for longer) than inanimate objects. In a visual search paradigm, Abrams and Christ (2003) showed that onset of motion (i.e., an object that has just started to move), but not motion per se, was important to capture attention. According to those authors, motion onset is indicative of animacy. They postulated that the reason why motion onset captures attention is that it may signal a biologically significant event, because objects that undergo motion onset must have their own internal energy source. Since survival may depend on the rapid detection of nearby predators and prey, one can easily imagine why a visual system in which motion onset captures attention is very useful to protect ourselves from predators. Finally, Pratt, Radulescu, Guo, and Abrams (2010) tested the hypothesis that animate motion captures visual attention by comparing the time taken to detect targets involving objects that were moving predictably as a result of collision (inanimate motion) with the time taken to detect targets involving objects that were moving unpredictably, not as a result of collision (animate motion). They found that participants responded more rapidly to targets involving objects undergoing animate than inanimate motion. They postulated that our evolutionary past, when detecting animate objects was critical for survival, has had a profound impact on the way in which we extract information from the visual field.
A novel and important finding has recently been reported in memory research, which we shall refer to as the animacy effect (Nairne, VanArsdall, Pandeirada, Cogdill, & LeBreton, in press; VanArsdall, Nairne, Pandeirada, & Blunt, 2013). The animacy effect refers to the observation that animate stimuli are remembered better than inanimate stimuli. This finding is in line with the functionalist view of memory put forward by Nairne and coworkers (Nairne, 2010; Nairne & Pandeirada, 2008, 2010a, 2010b), whereby the human memory system has been tuned to solve critical adaptive problems (e.g., finding food and water, protection from predators, finding a mate for reproduction). This finding is particularly important since it reinforces the view that human cognitive systems are tuned to detect and remember animate things. A number of studies comparing survival encoding scenarios with various deep encoding contexts (e.g., Nairne & Pandeirada, 2008, 2010a; Nairne, Pandeirada, Gregory, & Van Arsdall, 2009; Nairne, Pandeirada, & Thompson, 2008; Nairne, Thompson, & Pandeirada, 2007) have provided evidence supporting the view that memory systems have evolved to solve fitness problems. In their seminal study, Nairne et al. (2007) demonstrated that encoding words during a survival scenario (rating whether a word denotes something that could be useful in a situation in which the participant has to find food, water, and protection against predators), as compared to several control scenarios (e.g., rating words according to pleasantness), led to better long-term retention. Many different control scenarios have been used to test the survival processing advantage (Nairne & Pandeirada, 2008). The survival effect has been found in experiments carried out in different countries (e.g., the United States, Japan, or Germany) and with different populations (mostly young adults but also older adults and children). Although the boundaries of this effect have been disputed (Klein, 2013; Otgaar et al., 2011), as have the proximate mechanisms that underpin it (e.g., Burns, Burns, & Hwang, 2011; Nouchi, 2013), this effect has attracted considerable attention in the research community.
It is important to stress that in most studies investigating the survival processing advantage, the survival effect has mostly been obtained by varying the encoding contexts and not by directly using words more-or-less related to adaptive problems (e.g., water, knife, fire). Recently, however, Nairne and colleagues (Nairne et al., in press; VanArsdall et al., 2013) have used animate and inanimate stimuli that are clearly related to adaptive issues. VanArsdall et al. presented participants with nonwords together with properties that were characteristic of either living (animate) or nonliving (inanimate) things. Participants had to rate the likelihood that each nonword/property pair corresponded to either a living or a nonliving thing. They were then given a recognition memory test for the nonwords (Exp. 1) or a free-recall test (Exp. 2). In both experiments, the nonwords paired with properties of living things were remembered better than those paired with properties of nonliving things. In two other studies conducted with word stimuli (Nairne et al., in press), it was found that animate words were more likely to be recalled than inanimate words, even after the stimulus classes had been controlled for other mnemonically relevant dimensions (e.g., imageability and meaningfulness) either statistically (Study 1) or methodologically (Study 2).
The animacy effect found in memory tasks is important because it provides further empirical support for a functional view of memory. However, before any firm conclusion can be drawn from this memory effect, and in view of the novelty of this finding, we thought further evidence was required. The goal of our first experiment was to replicate in an incidental encoding task the animacy effect found in English by Nairne et al. (in press) with words in an intentional memory task. In Experiment 2, pictures instead of words were used, and again, the animacy effect was tested in free recall. Thus far, no studies have examined whether the advantage of animate over inanimate objects in memory also holds for picture stimuli. If memory has evolved to favor the processing of fitness-relevant information, it seems likely that the animacy effect would emerge with rich visual stimuli such as pictures. In Experiment 3, the animacy effect was tested with word stimuli in a recognition task using the remember/know paradigm (Gardiner, 1988) to evaluate not only the quantity but also the quality of recognition. In effect, the act of “remembering” involves recalling contextual episodic details of previous events and experiences, whereas “knowing” does not involve remembering as such, but is more a general sense of familiarity about more abstract knowledge. Finally, in Experiment 4, we collected sensory experience ratings (Juhasz, Yap, Dicke, Taylor, & Gullick, 2011), which are thought to reflect the extent to which a word evokes a sensory and/or perceptual experience (Juhasz & Yap, 2013). This enabled us to test whether the animacy effect could be attributable to the fact that animate items are richer in terms of sensory and/or perceptual features than inanimate items. More precise rationale of each experiment is provided below.
To sum up, although the main goal of our study was to provide further evidence for animacy effects in long-term memory, it is worth stressing that its originality lies in the fact that we investigated animacy effects (1) in recall tasks with (a) incidental learning and (b) pictures, and (2) in recollection tasks with the remember–know paradigm, and (3) we investigated whether these effects might be underpinned by different weights in terms of sensory features of animate versus inanimate items.
Experiment 1: Animacy effect in free recall with words
The goal of this experiment was to replicate Nairne et al.’s (in press) finding of better free recall of words denoting animate than inanimate items. In contrast to their experiment, we used an incidental rather than an intentional encoding memory task. The participants were first presented with words and had to categorize each one as animate or inanimate. Categorization times were recorded. The categorization task was followed by two interference tasks. Finally, the participants were given an (unexpected) free-recall task. The distinction between incidental and intentional memory involves differences in self-initiated strategies while encoding episodic information (Kirchhoff, 2009). Explicitly instructing participants to encode information enables them to initiate encoding strategies (i.e., intentional encoding). Although both incidental and intentional learning occur in everyday life, the former is more prevalent, in that people generally observe events and activities without explicit instructions to remember them.
Method
Participants
A group of 40 students (mean age 23.3 years) at the University of Bourgogne took part in the experiment in exchange for course credits. The participants were native French speakers and reported having normal or corrected-to-normal vision. None were taking medication known to affect the central nervous system.
Stimuli
A total of 56 nouns were selected from Snodgrass and Vanderwart’s (1980) and Bonin, Peereman, Malardier, Méot, and Chalard’s (2003) databases. Each word referred to either an animate or an inanimate object. The words were divided into two sets of 28 items matched for the surface variables of number of letters and bigram frequency; the lexical variables of book frequency, subtitle frequency, age of acquisition, number of orthographic neighbors, and orthographic uniqueness point; and the semantic variables of conceptual familiarity, imageability, image variability, concreteness, and emotional valence. The statistical characteristics of the controlled variables are presented in Table 1.
Since pictures corresponding to the words were used in Experiment 2, the two types of stimuli were also matched on variables pertaining to the pictures. These will be described in the Stimuli section of Experiment 2.
Apparatus
The words were presented on a Macintosh (iMac) computer running the PsyScope version 1.2.5 software (Cohen, MacWhinney, Flatt, & Provost, 1993).
Procedure
The participants were tested individually, seated comfortably in a quiet room. First, they carried out an animate-inanimate categorization task. They were given a brief definition of what is meant by “animate” versus “inanimate” (e.g., an animate item can move autonomously whereas an inanimate item cannot). They were told that for each presented word they should decide as quickly as possible whether it referred to an animate or inanimate item, indicating their choice by pressing a different key. Each trial began with a fixation point (+) displayed in the middle of the screen for 500 ms. A word was then displayed in the middle of the screen and remained there until the participant responded. The words were presented in random order. Response times were recorded. After the categorization task, the participants performed two interference tasks commonly used to evaluate executive functions, which lasted 5 min. The first was the “X–O” letter-comparison task (Salthouse, Toth, Hancock, & Woodard, 1997), and the second was the “plus–minus” task (from Jersild, 1927, and Spector & Biederman, 1976). In the letter-comparison task, the participants had to decide, as quickly as possible, whether or not pairs of letters were identical. For the plus–minus task, the participants were told to add 3 to each number on the first board, subtract 3 from each number on the second board, and alternate between adding and subtracting 3 from the numbers on the third board. After 5 min, the participants were given a surprise free-recall task.
Results and discussion
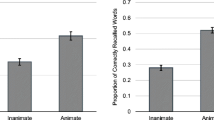
Animate words were categorized reliably faster (M = 960 ms, SD = 219) than inanimate words (M = 1,064 ms, SD = 234), t(39) = 5.58, p < .001. More animate words were correctly recalled (M = 4.78, SD = 2.08) than inanimate words (M = 2.08, SD = 1.40), yielding a reliable main effect of the Type of Word factor, t(39) = 7.50, p < .001. With regard to extralist intrusions, animate words (M = 1.05, SD = 1.08) did not yield more intrusions than inanimate ones (M = 1.10, SD = 0.90), t(39) = 0.24.
These findings replicate in French the animacy effect in correct free-recall initially reported by Nairne et al. (in press) in English in an intentional memory task. The effect cannot be attributable to the fact that animate words were processed for a longer time than inanimate words because we found that animate words were categorized faster than inanimate words.
It might be asked whether the animacy effect was driven by congruity. For instance, it could be that individuals relied on the animate category to make decisions about the category the item belonged to (e.g., “is this a living thing?”) and to cue performance during retrieval. However, we do not think the congruency account likely because the participants were instructed to decide whether a given word referred to an animate or an inanimate thing, each decision requiring a specific response, pressing a different key. It is important to stress that the participants were given a brief explanation about what is meant by animate and inanimate before starting the categorization task. Thus, the animate category was not defined in a more positive way than the inanimate category. This was also the case in VanArsdall et al.’s (2013) study in which the participants had to use a six-point scale anchored at one end by an object and at the other by a living thing. Moreover, the animacy effect was replicated many times with intentional learning in that study, in which attention was not drawn to the animacy dimension.
It is important to stress that the animacy effect in long-term memory was found after controlling for a large number of dimensions between the two types of words. In the next experiment, we tested whether the animacy effect would be found with picture stimuli.
Experiment 2: Animacy effect in free recall with pictures
The animacy effect in long-term memory has thus far been obtained with nonword (VanArsdall et al., 2013) and word (Nairne et al., in press, and Exp. 1 of the present study) stimuli. One unexplored issue is whether the animacy effect is also obtained with pictures. It is well known that information is more likely to be recollected when it is presented in pictures rather than in words (Paivio, 1971; Rajaram, 1996). Because processing pictures (i.e., imagery) preceded the processing of language (e.g., words) in the evolution of human memory (Paivio, 2007), on the basis of a functionalist view of human memory (Nairne, 2010), whereby memory has evolved to favor the processing of fitness-relevant information, we expected to find that pictures of animate items would yield better recall than pictures of inanimate items. Examining this hypothesis would provide valuable information regarding the robustness of the animacy recall advantage.
Method
Participants
A group of 30 adults (mean age 23.63 years) taken from the same pool as in Experiment 1 and having the same characteristics took part in the experiment.
Stimuli
The pictures corresponding to the words used in Experiment 1 were used. They were taken from two databases (Bonin et al., 2003b; Snodgrass & Vanderwart, 1980). In addition to the controls described in Experiment 1, animate and inanimate stimuli were also matched on variables pertaining to the pictures. Thus, the name agreement, image agreement, and visual complexity of the pictures were controlled for (see Table 1). Name agreement is the degree to which individuals agree on a name for a particular pictured object. One widely used measure is the percentage of participants who provide the most common name. Image agreement is the degree to which the mental images formed by participants in response to an object name match the object’s appearance; it is measured using a Likert scale. The visual complexity of pictures involves the number of lines and details in the drawing. Participants rate the degree of visual complexity of each picture on a 5-point scale, ranging from 1 for a visually simple picture to 5 for a visually very complex picture.
Apparatus
The same apparatus was used as in Experiment 1.
Procedure
We followed the same procedure as in Experiment 1, except that pictures were used instead of words. Although the name agreement scores were controlled for between animate and inanimate stimuli, the written words were displayed with the pictures to ensure that the participants used the intended names for the target pictures.
Results and discussion
Animate pictures were categorized significantly faster (M = 1,156 ms, SD = 377) than inanimate pictures (M = 1,279 ms, SD = 439), t(29) = 3.28, p < .01. More animate pictures were correctly recalled (M = 6.63, SD = 2.63) than inanimate pictures (M = 3.70, SD = 2.31), yielding a reliable main effect of the Type of Picture factor, t(29) = 3.28, p < .01. With regard to extralist intrusions, animate stimuli (M = 0.83, SD = 1.02) did not yield more intrusions than did inanimate ones (M = 0.70, SD = 1.09), t(29) = 0.47.
In this experiment, we tested whether the animacy effect in memory obtained with words would also be found with pictures. Taking a functionalist view of human memory (Nairne, 2010), this effect was clearly anticipated, particularly as the processing of pictures preceded that of language in human evolution. In effect, according to the adaptive memory view, our memory systems have evolved to favor the processing of fitness-relevant information. Therefore, whatever the format of presentation of animate versus inanimate entities, we predicted that the former would be remembered better than the latter; this prediction was clearly borne out. In line with previous studies (Paivio, 1971; Rajaram, 1996), comparison of the findings of Experiments 1 and 2 revealed better memory performance with pictures than with words. The first two experiments support previous findings (Nairne et al., in press; VanArsdall et al., 2013) of the robustness of the animacy recall advantage.
However, given that the pictures were accompanied by words, one might wonder whether the better memory performance with pictures than with words was due to the picture format itself or to the fact that the word format comprised a single stimulus whereas the picture format included an additional redundant stimulus. Thus, the issue is whether the advantage of animates over inanimates would have been found had the pictures not been labeled. As is explained above, we decided to present each picture with its corresponding written name because although the name agreement of the pictures was matched across animates and inanimates and was relatively high (>80 %), it was not equal to 100 %. However, we believe that the better performance with pictures than with words observed in the present study was driven by picture processing and not by the fact that redundant stimuli were presented (the picture plus its written name), for the following reason. According to the Paivio’s dual-code theory (Paivio & Csapo, 1973), the picture superiority effect in memory can be explained by the fact that during study, pictures elicit their verbal label, so that two representations or codes are stored in memory. By contrast, words do not automatically elicit a picture and have a relatively impoverished memory representation. The redundant representation for pictures favors their retrieval relative to words. Taking this view, in our experiment, the presentation of a picture would lead to both an imagery code and a verbal code, the latter being given additional activation by the accompanying written word. According to the redundancy account, the better performance for “pictures + words” in Experiment 2 than for “words only” in Experiment 1 would essentially be due to the fact that the name of the picture was effectively presented twice, and therefore, more activated. However, Paivio and Csapo showed that although pictures accompanied by words (PW) are better remembered than the same picture presented alone (P), they are not recalled as well as the word presented twice (WW). Although this issue might well be reexamined in future studies, it is likely that the superiority of recall with pictures than with words observed in the present study was driven by the contribution of imaginal memory codes more than by the redundancy of verbal memory codes.
Experiment 3: The animacy effect in recognition with words
In this experiment, we tested whether the animacy effect obtained in free recall would also be found in recognition using the word stimuli of Experiment 1. Nairne et al. (in press) did not test the animacy effect obtained with words in a word recognition task. From an empirical point of view, it would thus be interesting to assess whether the animacy effect found in free recall can be observed in recognition, and more precisely, in the quality of recognition.
A number of studies in recognition memory suggest that people experience at least two quite distinct states of awareness related to the quality of recognition. The first state—remembering—involves recalling contextual details of previous events and experiences, including awareness of one’s self, usually at a particular time and in a particular place. The second state—knowing—does not involve remembering an event as such, but is a more abstract awareness of general knowledge. According to Tulving (1985), these two states of awareness reflect autonoetic and noetic consciousness, respectively, which in turn reflect two mind/brain systems, namely episodic and semantic memory (see also Wheeler, Stuss, & Tulving, 1997). It is assumed that these two states of awareness can be measured with the remember–know paradigm (Gardiner, 1988) in which participants are given a recognition memory test and are instructed to assign their positive recognition decisions to either “remember” responses (R-responses) whenever they can consciously recollect the item’s study presentation, or “know” responses (K-responses) when they recognize the item on the basis of familiarity but are not able to consciously recollect its study presentation. Thus, remembering occurs when an individual has a sense or feeling of the self in the past (autonoetic consciousness; Tulving, 1985). Images (often visual ones), feelings, thoughts, sensations and verbal statements directly related to the recalled episode also often come to mind during the recollective experience. The first two experiments showed that animacy enhanced the quantity of recalled information. In the third experiment, we tested the quality of the mnemonic trace by investigating whether items were recalled on the basis of remembering (R-responses) or knowing (K-responses). This would indicate whether the animacy effect on recognition involves R- and/or K-responses and would provide a better understanding of the animacy effect in long-term memory.
Method
Participants
A group of 33 students (mean age 20.12 years) at the University of Bourgogne participated in the study in exchange for course credits. None were taking any medication known to affect the central nervous system.
Stimuli
The same word stimuli as in Experiment 1 were used here. In addition, 56 new words (half animate and half inanimate) were added for the “new” responses in the recognition task. These new words (“new”) matched the initial experimental words (“old”) on objective word frequency (“old,” M = 12.43, SD = 24.22, min–max = 0.03–115.3; “new,” M = 12.73, SD = 25.94, min–max = 0.03–144.3; t < 1), age of acquisition (“old,” M = 2.64, SD = 0.73, min–max = 1.15–4.2; “new,” M = 2.73, SD = 0.83, min–max = 1.35–4.62; t < 1), and number of letters (“old,” M = 6.55, SD = 1.89, min–max = 3–10; “new,” M = 6.80, SD = 2.27, min–max = 3–15; t < 1).
Apparatus
The same apparatus was used as in the previous experiments.
Procedure
The procedure was the same as in Experiment 1 except that a recognition test was performed after the distraction tasks. For this test, 112 words (56 targets and 56 fillers) were presented one at a time on the computer screen in random order and with no time limit. The participants had to say aloud if they recognized each word from the previous lists. For every word they recognized, participants had to indicate whether their recognition was based on remembering (R-response), knowing (K-response), or guessing (G-response; Gardiner, Ramponi, & Richardson-Klavehn, 1999). They were instructed to give an R-response when the word evoked a specific recollection of the learning sequence. Examples given included remembering a word because it brought to mind a particular association, image, or some other personal experience, or because something about its appearance or position could be recalled. A K-response was to be given for any words that the participants felt confident about recognizing but that did not evoke any specific conscious memory of the learning sequence. G-responses were to be used when they were not sure whether they had seen the word in the study list or not and were included to enhance the quality of K-responses. After the recognition phase, the participants had to explain at least two of their Remember and two of their know judgments to ensure that they used the two types of response correctly.
Results and discussion
Inanimate words took longer to categorize (M = 1,202 ms, SD = 276) than animate words (M = 1,117 ms, SD = 274), t(32) = 3.66, p < .001.
Overall recognition
More animate words (M = 22.30, SD = 3.55) were recognized than inanimate words (M = 20.33, SD = 5.32), t(32) = 2.54, p = .016. Animacy did not have a significant effect on false alarms (animate words, M = 4.03, SD = 3.18; inanimate words, M = 4.06, SD = 3.49), t(32) = 0.06.
“Remember” and “know” responses
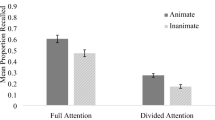
The means and standard errors for R- and K-responses are presented in Fig. 1. For R-responses, t test comparisons revealed a main effect of type of word, t(32) = 2.84, p < .01, with animate words yielding higher R-responses than did inanimate words. For K-responses, we found no reliable effect of the type of word, t(32) = 0.07. The analyses revealed a significant difference between R- and K-responses for animate words, t(32) = 3.66, p < .01, with more R- than K-responses. In contrast, no reliable difference was apparent between the two types of responses for inanimate words, t(32) = 1.25, n.s. For G-responses, no reliable difference emerged between animate (M = 2.72, SD = 2.40) and inanimate (M = 2.85, SD = 2.12) words, t(32) = 0.30. Finally, for false alarms, no reliable differences were observed between animate and inanimate words on R-responses (animate, M = 0.57, SD = 0.97; inanimate, M = 0.76, SD = 1.68), t(32) = 0.83, K-responses (animate, M = 1.36, SD = 1.82; inanimate, M = 1.03, SD = 1.88), t(32) = 1.13, or G-responses (animate, M = 2.09, SD = 2.74; inanimate, M = 2.27, SD = 2.54), t(32) = 0.57.

Mean numbers and standard errors of “remember” and “know” responses as a function of animacy (animate vs. inanimate stimuli)
Although Experiment 1 of VanArsdall et al.’s (2013) study used recognition memory for nonwords, the animacy effect has previously only been reported with words in free-recall tasks (Nairne et al., in press). The aim of Experiment 3 was therefore to assess whether it would also appear in a recognition task. More importantly, we collected “remember” and “know” responses for each item judged as “recognized” since such responses are thought to reflect the kind of memory trace formed during encoding. Two important findings emerged from the analyses. First of all, the animacy property enhanced the quantity of recognized words, as more animate than inanimate words were recognized. Secondly, animate words yielded higher R-responses than inanimate words, indicating greater conscious awareness of encoding these stimuli, suggesting that animacy enhances the quality of memory traces leading to greater episodic retrieval. This finding suggests that the participants were spontaneously engaged in elaborative encoding for animate words. These findings also support the idea that the mnemonic advantage of animate words is provided by the recollection component. From an evolutionary perspective, the key uses of episodic memory are to maintain a sense of self-continuity, to ensure successful social interaction, and to direct future behavior on the basis of information about past events (Raby & Clayton, 2012). Given the functions of episodic memory and the properties of animate objects (e.g., animates can act whereas inanimates move only when something/someone acts on them; animates can know, perceive, emote, learn and think; Gelman & Spelke, 1981), it is likely that animate objects will be recollected with more episodic details than inanimate objects.
In Experiment 4, we assessed the merits of this analysis and tested whether the hypothesis that animate items have better memory traces than inanimate items could be related to the former being richer in terms of sensory (and/or perceptual) experiences (Juhasz & Yap, 2013). Images (often visual), feelings, thoughts, sensations and verbal statements directly related to the recalled episode often come to mind during the recollection process (Tulving, 1985) and could be influenced by sensorial experiences.
Experiment 4: Sensory experience ratings and animacy
In the following experiment, we assessed whether the animacy advantage in memory could be related to a specific property of animate entities, and more specifically, whether animate stimuli are richer in terms of sensory experiences than inanimate stimuli. A number of studies suggest that semantically rich stimuli are remembered better than those that are less rich (e.g., Hargreaves, Pexman, Johnson, & Zdrazilova, 2012). Hargreaves et al. found that words for concepts with a large number of semantic features were recalled better than those with few semantic features in both expected and unexpected memory tasks. In a recent study, Hoffman and Lambon Ralph (2013) used a more direct approach in which subjects rated their knowledge of objects in each sensory-motor modality separately. Compared with these ratings, feature listing over-estimated the importance of visual form and functional knowledge and under-estimated the contributions of other sensory channels. An item’s sensory rating was found to be a better predictor of lexical-semantic processing speed than the number of features it possessed, suggesting that such ratings better capture the overall quantity of sensory information associated with a concept. Importantly, the findings of Hoffman and Lambon Ralph also strongly suggest that animate stimuli are richer in terms of sensory knowledge than inanimate stimuli.
In the following experiment, we used sensory experience ratings (SER) to capture the richness of the animate versus inanimate items used in the previous experiments. According to Juhasz and Yap (2013), SER are thought to reflect the extent to which a word evokes a sensory and/or perceptual experience. SER are obtained by asking participants to rate on a Likert scale the degree to which any given word evokes a sensory experience. More precisely, participants are asked to judge the ability of a word to evoke an actual sensation (taste, touch, sight, sound, or smell) when they are reading the word. The sensory experience variable is therefore not limited to a single sensation and it has the potential to index the links between lexical-semantics and all sensory/perceptual modalities. If the animacy effect in memory is driven by the different weights of sensory experiences of animate versus inanimate items, we should find that SER are higher for animate than inanimate items. Therefore, in Experiment 4, we tested the hypothesis that the SER variable, which is related to semantic richness, could account for the animacy effect found in the first three experiments. We also added the French translation of the English words used by Nairne et al. (in press) since they did not use this measure in their study.
Method
Participants
A group of 56 adults (mean age = 43 years) participated in the study. They were taken from a larger pool of participants (mostly university students but also people from surrounding areas) than in the previous experiments.
Stimuli
The word stimuli used in Experiment 1 were again used here. We also added the French translation of the words used by Nairne et al. (in press).
Procedure
The participants were asked to rate the sensory experience associated with words presented in a list using a 7-point scale ranging from 1 = no sensorial experience to 7 = high sensorial experience. Some examples of words with different types of ratings were given before starting the experiment. We followed closely the procedure used in Juhasz and Yap’s (2013) study.
Results and discussion
In our study, animate words were given SER (M = 3.00, SD = 0.81) that did not differ reliably from inanimate words (M = 3.08, SD = 0.67), t < 1. It was also the case in the Nairne et al. (in press) study (M = 3.04, SD = 0.82, vs. M = 3.38, SD = 0.46), t < 1.
In Experiment 4, we tested whether the animacy effect in memory could be driven by the fact that animate entities are richer in terms of sensory experience than inanimate items. To do this, we collected SER for the words used in Experiments 1 and 2 plus the French translation of the words used in Nairne et al.’s (in press) Experiment 2.
Based on studies suggesting that items that are semantically rich are better remembered than items that are less rich (Hargreaves et al., 2012), and given the findings of Hoffman and Lambon Ralph (2013) suggesting greater sensory knowledge for animate than inanimate items, it was not unreasonable to hypothesize that the animacy effect could be driven by this variable. The findings were clear: No reliable difference was observed between the SER for animate and inanimate words in either the present study or the one by Nairne et al. (in press), and if anything, the difference in SER was descriptively very small for each set and ran counter to the prediction than animates are richer in sensory experience than inanimates.Footnote 1 Thus, the animacy effects in memory cannot be easily ascribed to differences in the richness of sensory and perceptual features. However, it could be argued that testing the hypothesis that the animacy effect was due to the richness of encoding was rather odd in that our stimuli were controlled for imageability. In effect, this variable is typically assumed to index the richness of semantic representations (Evans, Lambon Ralph, & Woollams, 2012), with the result that words that are rated high in imagery are thought to be semantically richer than those that are rated as low in imagery. However, Connell and Lynott (2012) have recently shown that imageability is visually biased at the expense of other perceptual modalities. Therefore, the findings of Experiment 4 are particularly interesting in that they clearly show that the animacy effect is not underpinned by differences at the level of sensory features.
General discussion
A growing body of evidence supports the view that our memory systems have evolved to reflect specific selection pressures that belonged to our ancestral past. A core assumption of evolutionary psychologists is that the human mind is composed of cognitive modules that fulfill specific functions (Bell & Buchner, 2012). Therefore, as claimed by Nairne (2012), just as the heart serves the function of pumping the blood to the organs, and the different “apps” of our iPhones have specific functions (e.g., to indicate where we are and where to go; to provide information about train times), there is no reason why our memory systems would not have specific functions. Indeed, several studies have shown a memory advantage of processing information in terms of fitness value (Nairne et al., 2009; Nairne et al., 2008; Nairne et al., 2007). More recently, it has been found that processing items along an animacy dimension led to better retention of animate than inanimate items (Nairne et al., in press; VanArsdall et al., 2013). In the present study, we aimed to provide further evidence for the animacy effect in long-term memory. Four experiments were conducted and the findings can be easily summarized.
In Experiment 1, we found that incidental encoding led to better recall of words referring to animate than inanimate items. In Experiment 2, we found that the animacy effect found in long-term memory with words also extended to picture stimuli. This was of particular interest because, according to the adaptive memory view, our memory systems have evolved to favor the processing of fitness-relevant information, whatever the format in which animate versus inanimate entities are encountered.
In Experiment 3, we used a recognition paradigm to test for the animacy effect in memory. Importantly, in order to gain a better understanding of this effect, we also investigated “remember” and “know” responses for each recognized item. We found that animate words were better recognized than inanimate words and that R-responses outnumbered K-responses for animate stimuli, whereas no difference emerged between R- and K-response for inanimate stimuli. Moreover, the findings showed that the mnemonic advantage of animate words was produced by the recollection component. This suggests that animate words benefit from more elaboration processes at encoding. It should be remembered that from an evolutionary perspective, episodic memory allows individuals to maintain a sense of self-continuity, to be successful in social interactions, and to use information from past events to direct future behavior (Raby & Clayton, 2012). This novel finding is in line with the functionalist view of memory put forward by Nairne and coworkers (Nairne, 2010; Nairne & Pandeirada, 2008, 2010a, 2010b) whereby the human memory system has been tuned to solve critical adaptive problems (e.g., finding food and water, protection from predators, finding a mate for reproduction). More particularly, the observation of more R-responses for animate than for inanimate stimuli suggests that they possess better quality and more stable traces in memory.
The advantage of animate over inanimate words and pictures was found in Experiments 1–3 in the context of an animate–inanimate categorization task. It might be asked whether the same effects would be found if the categorization task involved another dimension of the stimuli (e.g., their size). In effect, we did not design an experiment in which the orienting task was changed to draw attention to another dimension of the stimuli. However, Nairne et al. (in press) used intentional memory tasks in which no explicit mention was made of the animate–inanimate distinction in the stimuli, and the animacy effects on memory performance were still reliable. It could be argued that if the animacy effect does indeed support the functionalist view of memory, spontaneous encoding must be shown to occur along the animacy dimension. In fact there are clear empirical grounds to support this assumption, as reviewed above in the introduction.
Finally, in Experiment 4, we tested whether the hypothesis that the better quality of memory traces for animate than inanimate items might be related to the former being richer in terms of perceptual and sensory features. However, neither the sensory experience ratings for our stimuli nor for those in Nairne et al.’s (in press) Experiment 2, yielded any reliable difference between animate and inanimate stimuli. Therefore, the animacy effect in memory is not attributable to differences in the richness of sensory/perceptual features or of semantic features in general. The functional view of memory assumes that what matters for the animacy effect is the status in terms of fitness of the things that we process. Because animate stimuli have a greater fitness value than inanimate stimuli, they are given processing priority. It should be noted however that the categorization times were reliably shorter for animate than inanimate stimuli in all experiments, in line with previous findings in the literature. However, there is still a possibility that animate stimuli have “something special” that renders them easier to remember than inanimate stimuli, something that has nothing to do with their animacy status. But what exactly?
Is the animacy effect really due to “animacy” or did we miss something when investigating this effect?
As far as the animacy effects in long-term memory are concerned, the possibility still exists that we are victims of confounding variables—namely, that we think we have shown a genuine animacy effect, when this effect is in fact driven by (an)other, uncontrolled variable(s).
First and foremost, it should be remembered that VanArsdall et al. (2013) initially found that animate items were remembered better than inanimate items using nonwords. The use of nonwords was justified by the authors by the fact that using words would require matching the stimuli on numerous dimensions and also because “demonstrating that people are more likely to remember animals than household objects might not be seen as particularly convincing by the community of memory researchers” (p. 173). Since the same nonwords were used (with different participants) with animate and inanimate properties in VanArsdall et al.’s (2013) study, it is difficult to assume that the animacy effect was due to uncontrolled properties of the stimuli. Somewhat paradoxically, in a further publication (Nairne et al., in press), the authors investigated the animacy effect in long-term memory using words (and thus contrary to their claim that finding an animacy effect with words would not be accepted by the research community).
Second, and more generally, an important and recurring issue in experimental psychology is to ensure that an effect on a behavioral outcome is genuinely attributable to the manipulation carried out and not to another potential (but uncontrolled) variable. This concern applies here as one cannot definitively ascertain that the animacy effect in memory is not attributable to another influential variable. In other words, the animacy effect could be due to (an)other characteristic(s) of the words that is (are) correlated with the animate–inanimate distinction. This concern is particularly relevant in psycholinguistic studies in which researchers aim to establish which specific characteristics of the words play a role in lexical processing. Controlling for stimuli in psycholinguistic experiments has been said to be a difficult challenge (Cutler, 1981). To give an example, a lively debate in the literature on object and word naming has concerned whether the effect of the frequency of encountering words is actually a genuine effect of word frequency or is due to another factor such as age of acquisition (AoA). Some researchers have claimed that word frequency effects in object naming are in fact AoA effects (Bonin, Fayol, & Chalard, 2001), but subsequent studies using better word frequency measures have found effects of both variables (e.g., Bonin et al., 2003b).
Could the animacy effect be attributable to another factor, namely a by-product of other more general factors? We were careful when selecting our stimuli to control for a large number of important factors that could potentially influence word processing and long-term encoding. Since certain studies have suggested that the quality of memory traces is dependent upon the richness of the stimuli in terms of semantic or motoric features (Hargreaves et al., 2012), and given (1) the findings of our Experiment 3 that suggest that the memory traces of animate stimuli are of better quality than those of inanimate stimuli, and (2) the findings of Hoffman and Lambon Ralph (2013) suggesting that animate stimuli are richer in terms of sensory knowledge than inanimate stimuli, our Experiment 4 tested whether the animacy effect in memory might be related to differences in terms of sensory features. The outcome of this experiment was clear: Neither our stimuli nor those used by Nairne et al. (in press) differed reliably on this dimension. Of course, we cannot definitively rule out the possibility that the animacy effect is due to another potent variable rather than the status of animacy per se. But since we were able to control for a large number of potentially important variables, the weight of evidence reported in the present study, together with recent evidence on words (Nairne et al., in press) and nonwords (VanArsdall et al., 2013), strongly suggests that animacy has a strong influence on long-term memory.
Evidence for a functional view of memory
As we shall discuss now, we think that animacy effects in memory provide further convincing evidence for a functional view of human memory (and it should be stressed that this was the goal of the present study). Nairne and coworkers have defended the view that our memory has been sculpted by evolution as a result of problems faced by our ancestors, based essentially on findings obtained using survival processing scenarios. They found that processing stimuli (and not stimuli obviously related to adaptive problems) in terms of fitness yields better memory performance than processing them in terms of other dimensions corresponding to well-known deep encoding conditions (Nairne et al., 2008). However, the idea that the survival processing effect (e.g., Nairne et al., 2007) is due to the contribution of episodic memory to maximizing fitness in our ancestral past, that is to say that our episodic memory helped our ancestors to solve adaptive problems related to survival (Nairne & Pandeirada, 2010a, 2010b), has been challenged by a number of researchers. They maintain that the survival effect can be accounted for by appealing to more general proximate memory mechanisms such as elaboration (Howe & Derbish, in press; Kroneisen & Erdfelder, 2011), item-specific processing (Burns, Hart, Griffith, & Burns, 2013), or richness of encoding (Kroneisen, Erfelder, & Buchner, 2013). Howe and Otgaar (2013) have claimed that general memory principles provide a better explanation of the adaptive function of memory.
According to Nairne (in press), the criticisms raised against the evolutionary account of the survival memory advantage have come from a misunderstanding between ultimate and proximate explanations. Without going into the details of the line of argument, he defines ultimate explanations as “statements about the function of a trait and ‘why’ it would have been selected by nature during an evolutionary process,” whereas proximate explanations “focus on the mechanisms that produced the trait—that is, they are statements about ‘how’ the trait works and the condition under which the trait is likely to be expressed” (p. 309). Following this distinction, memory evolved because it solved specific problems related to fitness (e.g., remembering information processed for its survival value as indexed by the survival memory paradigm). One consequence of memory system evolution is that information relevant to survival would be afforded special status. However, the proximate mechanisms underpinning the retention advantages may well be elaborative, distinctive or self-related encoding.
The animacy effect in memory can clearly not be disputed on the grounds of its adaptive value. The core of the adaptive view of memory is that not all stimuli are equal when processing them in order to remember them; stimuli that are relevant to fitness are more important than other types of stimuli. Therefore, as claimed above, animacy effects in memory provide further evidence for a functionalist view of memory.
A remaining issue that should be the focus of future studies is whether animacy effects in long-term memory are due to animates being inherently more memorable, all things being equal, or to the fact that they capture attention or lead to some kind of mental simulation that enhances memory. In other words, animacy effects in memory might be explained via attentional mechanisms rather than inherent mnemonic tunings. In their two empirical papers on animacy effects, Nairne and colleagues (Nairne et al., in press; VanArsdall et al., 2013) envisioned such a possibility, stating that “another possibility is that the nonwords processed for animacy were remembered well because animate things are especially likely to capture attention” (VanArsdall et al., 2013, p. 176). It might be tempting from the categorization data of our experiments, consistently showing that animates were categorized faster than inanimates, to suggest that the proximate mechanism underlying animacy effects is attentional capture. However, since our study was not designed to test such an explanation, further research will be required in order to ascertain whether or not animate objects attract more attention than inanimate ones, and whether this differential attention processing results in better memory. Clearly, an avenue has been revealed for testing hypotheses regarding the proximate mechanisms of animacy effects in episodic memory—in other words, the “how” of these effects. In the meantime, the findings of the present study reinforce the claim made earlier (Nairne et al., in press; VanArsdall et al., 2013) that we remember animate things better than inanimate ones. This is a robust findingFootnote 2 and provides further evidence for the adaptive view of memory (Nairne, 2010; Nairne & Pandeirada, 2010a, 2010b).
It must be stressed that the primacy of processing animates over inanimates seems to be a general phenomenon as it has not only been found in perception and memory, but also in language production, showing that animate entities tend to be privileged during syntactic production processes (e.g., Branigan, Pickering, & Tanaka, 2008). In cognitive neuropsychology, the dissociation between animates and inanimates has been reported in brain-damaged patients (e.g., Bi, Han, Shu, & Caramazza, 2007). Furthermore, the brain regions underpinning the processing of animates versus inanimates have been investigated, and certain studies (e.g., Wiggett, Pritchard, & Downing, 2009) suggest that different parts of the brain are differentially involved (ventrolateral visual brain regions for animate stimuli and ventromedial for inanimate ones). Finally, the ability to distinguish between animate and inanimate items has been assumed to underlie the construction of different mental categories in the brain (Gelman & Spelke, 1981).
In conclusion, we believe that the animacy effect in memory provides another type of demonstration that clearly supports the functionalist view of memory put forward by Nairne and colleagues (Nairne, 2010; Nairne & Pandeirada, 2010a, 2010b). Indeed, the only straightforward explanation for this effect is that remembering animate entities is helpful for survival. To paraphrase a well-known saying, we could say that as far as remembering is concerned, “all stimuli are equal but some stimuli (animate ones) are more equal than others.” The next step for opponents of this view, if they want to rule out the hypothesis that the animacy effect in memory is due to our memory system being selectively tuned to processing and remembering animate items, will be to show that animate and inanimate items covary along another, uncontrolled dimension that is genuinely responsible for this effect.
Notes
Since it is always difficult to draw conclusions from null results, we ran an additional Bayesian analysis from the data of Experiment 4, to determine the extent to which the prediction of a reliable animacy effect on SER could be rejected. In effect, according to Masson (2011, p. 679), this kind of analysis “generates a graded level of evidence regarding which model (e.g., effect absent [null hypothesis] vs. effect present [alternative hypothesis]) is more strongly supported by the data.” The analysis revealed positive evidence (Raftery, 1995) in favor of the null hypothesis.
References
Abrams, R. A., & Christ, S. E. (2003). Motion onset captures attention. Psychological Science, 14, 427–432. doi:10.1111/1467-9280.01458
Alario, F.-X., & Ferrand, L. (1999). A set of 400 pictures standardized for French: Norms for name agreement, image agreement, familiarity visual complexity, image variability, and age of acquisition. Behavior Research Methods Instruments & Computers, 31, 531–552.
Bell, R., & Buchner, A. (2012). How adaptive is memory for cheaters? Current Directions in Psychological Science, 21, 403–408. doi:10.1177/0963721412458525
Bi, Y., Han, Z., Shu, H., & Caramazza, A. (2007). Nouns, verbs, objects, actions, and the animate/inanimate effect. Cognitive Neuropsychology, 24, 485–504. doi:10.1080/02643290701502391
Bonin, P., Fayol, M., & Chalard, M. (2001). Age of acquisition and word frequency in written picture naming. Quarterly Journal of Experimental Psychology, 54A, 469–489. doi:10.1080/02724980042000219
Bonin, P., Méot, A., Aubert, L., Malardier, N., Niedenthal, P., & Capelle-Toczek, M.-C. (2003a). Normes de concrétude, de valeur d’imagerie, de fréquence subjective et de valence émotionnelle pour 866 mots [Concreteness, imageability, subjective frequency and emotional valence norms for 866 words]. L’Année Psychologique, 104, 655–694.
Bonin, P., Peereman, R., Malardier, N., Méot, A., & Chalard, M. (2003b). A new set of 299 pictures for psycholinguistic studies: French norms for name agreement, image agreement, conceptual familiarity, visual complexity, image variability, age of acquisition, and naming latencies. Behavior Research Methods, Instruments, & Computers, 35, 158–167. doi:10.3758/BF03195507
Branigan, H. P., Pickering, M. J., & Tanaka, M. (2008). Contributions of animacy to grammatical function assignment and word order during production. Lingua, 118, 172–189. doi:10.1016/j.lingua.2007.02.003
Brosch, T., & Sharma, D. (2005). The role of fear-relevant stimuli in visual search: A comparison of phylogenetic and ontogenetic stimuli. Emotion, 5, 360–364. doi:10.1037/1528-3542.5.3.360
Burns, D. J., Burns, S. A., & Hwang, A. J. (2011). Adaptive memory: Determining the proximate mechanisms responsible for the memorial advantages of survival processing. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37, 206–218. doi:10.1037/a0021325
Burns, D. J., Hart, J., Griffith, S. E., & Burns, A. D. (2013). Adaptive memory: The survival scenario enhances item-specific processing relative to a moving scenario. Memory, 21, 695–706. doi:10.1080/09658211.2012.752506
Capitani, E., Laiacona, M., Mahon, B., & Caramazza, A. (2003). What are the facts of semantic category-specific deficits? A critical review of the clinical evidence. Cognitive Neuropsychology, 20, 213–261. doi:10.1080/02643290244000266
Cohen, J. D., MacWhinney, B., Flatt, M., & Provost, J. (1993). PsyScope: A new graphic interactive environment for designing psychology experiments. Behavioral Research Methods, Instruments, & Computers, 25, 257–271. doi:10.3758/BF03204507
Connell, L., & Lynott, D. (2012). Strength of perceptual experience predicts word processing performance better than concreteness or imageability. Cognition, 125, 452–465. doi:10.1016/j.cognition.2012.07.010
Cutler, A. (1981). Making up materials is a confounded nuisance, or: Will we be able to run any psycholinguistic experiments at all in 1990? Cognition, 10, 65–70. doi:10.1016/0010-0277(81)90026-3
Evans, G., Lambon Ralph, M., & Woollams, A. (2012). What’s in a word? A parametric study of semantic influences on visual word recognition. Psychonomic Bulletin & Review, 19, 325–31. doi:10.3758/s13423-011-0213-7
Gardiner, J. M. (1988). Functional aspects of recollective experience. Memory & Cognition, 16, 309–313. doi:10.3758/BF03197041
Gardiner, J. M., Ramponi, C., & Richardson-Klavehn, A. (1999). Response deadline and subjective awareness in recognition memory. Consciousness and Cognition, 8, 484–496. doi:10.1006/ccog.1999.0409
Gelman, R., & Spelke, E. (1981). The development of thoughts about animate and inanimate objects: Implications for research on social cognition. In J. H. Flavell & L. Ross (Eds.), Social cognitive development: Frontiers and possible futures (pp. 43–66). Cambridge, UK: Cambridge University Press.
Hargreaves, I. S., Pexman, P. M., Johnson, J. C., & Zdrazilova, L. (2012). Richer concepts are better remembered: Number of features effects in free recall. Frontiers in Human Neuroscience, 6, 73. doi:10.3389/fnhum.2012.00073
Hoffman, P., & Lambon Ralph, M. A. (2013). Shapes, scents and sounds: Quantifying the full multi-sensory basis of conceptual knowledge. Neuropsychologia, 51, 14–25. doi:10.1016/j.neuropsychologia.2012.11.009
Howe, M. L., & Derbish, M. H. (in press). Adaptive memory: Survival processing, ancestral relevance, and the role of elaboration. In B. L. Schwartz, M. L. Howe, M. P. Toglia, & H. Otgaar (Eds.), What is adaptive about adaptive memory? New York, NY: Oxford University Press.
Howe, M. L., & Otgaar, H. (2013). Proximate mechanisms and the development of adaptive memory. Current Directions in Psychological Science, 22, 16–22. doi:10.1177/0963721412469397
Jersild, A. T. (1927). Mental set and shift. Archives of Psychology, 89, 5–82.
Juhasz, B. J., & Yap, M. J. (2013). Sensory experience ratings for over 5,000 mono- and disyllabic words. Behavior Research Methods, 45, 160–168. doi:10.3758/s13428-012-0242-9
Juhasz, B., Yap, M. J., Dicke, J., Taylor, S. C., & Gullick, M. M. (2011). Tangible words are recognized faster: The grounding of meaning in sensory and perceptual systems. Quarterly Journal of Experimental Psychology, 64, 1683–1691. doi:10.1080/17470218.2011.605150
Kirchhoff, B. A. (2009). Individual differences in episodic memory: The role of self-initiated encoding strategies. Neuroscientist, 15, 166–179. doi:10.1177/1073858408329507
Kirchner, H., & Thorpe, S. (2006). Ultra-rapid object detection with saccadic eye movements: Visual processing speed revisited. Vision Research, 46, 1762–1776. doi:10.1016/j.visres.2005.10.002
Klein, S. T. (2013). Does optimal recall performance in the adaptive memory paradigm require the encoding context to encourage thoughts about the environment of evolutionary adaptation? Memory & Cognition, 41, 49–59. doi:10.3758/s13421-012-0239-8
Kroneisen, M., & Erdfelder, E. (2011). On the plasticity of the survival processing effect. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37, 1553–1562. doi:10.1037/a0024493
Kroneisen, M., Erdfelder, E., & Buchner, A. (2013). The proximate memory mechanism underlying the survival processing effect: Richness of encoding or interactive imagery? Memory, 21, 494–502. doi:10.1080/09658211.2012.741603
Lipp, O. V., Derakshan, N., Waters, A. M., & Logies, S. (2004). Snakes and cats in the flower bed: Fast detection is not specific to pictures of fear-relevant animals. Emotion, 4, 233–250. doi:10.1037/1528-3542.4.3.233
Masson, M. E. J. (2011). A tutorial on a practical Bayesian alternative to null hypothesis significance testing. Behavior Research Methods, 43, 679–690. doi:10.3758/s13428-010-0049-5
Nairne, J. S. (2010). Adaptive memory: Evolutionary constraints on remembering. In B. H. Ross (Ed.), The psychology of learning and motivation (Vol. 53, pp. 1–32). Burlington, VT: Academic Press.
Nairne, J. S. (2012, March). Adaptive memory: Evolutionary constraints on remembering. Paper presented at the 54. Tagung experimentell arbeitender Psychologen (TeaP), Mannheim, Germany.
Nairne, J. S. (in press). Adaptive memory: Controversies and future directions. In B. L. Schwartz, M. L. Howe, M. P. Toglia, & H. Otgaar (Eds.), What is adaptive about adaptive memory? New York, NY: Oxford University Press.
Nairne, J. S., & Pandeirada, J. N. S. (2008). Adaptive memory: Remembering with a stone-age brain. Current Directions in Psychological Science, 17, 239–243. doi:10.1111/j.1467-8721.2008.00582.x
Nairne, J. S., & Pandeirada, J. N. S. (2010a). Adaptive memory: Ancestral priorities and the mnemonic value of survival processing. Cognitive Psychology, 61, 1–22. doi:10.1016/j.cogpsych.2010.01.005
Nairne, J. S., Pandeirada, J. N. S. (2010b). Memory functions. In The Corsini encyclopedia of psychology and behavioral science (4th ed., Vol. 3, pp. 977–979). Hoboken, NJ: Wiley.
Nairne, J. S., Pandeirada, J. N. S., Gregory, K. J., & Van Arsdall, J. E. (2009). Adaptive memory: Fitness-relevance and the hunter-gatherer mind. Psychological Science, 20, 740–746. doi:10.1111/j.1467-9280.2009.02356.x
Nairne, J. S., Pandeirada, J. N. S., & Thompson, S. R. (2008). Adaptive memory: The comparative value of survival processing. Psychological Science, 19, 176–180. doi:10.1111/j.1467-9280.2008.02064.x
Nairne, J. S., Thompson, S. R., & Pandeirada, J. N. S. (2007). Adaptive memory: Survival processing enhances retention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33, 263–273. doi:10.1037/0278-7393.33.2.263
Nairne, J. S., VanArsdall, J. E., Pandeirada, J. N. S., Cogdill, M., & LeBreton, J. M. (in press). Adaptive memory: The mnemonic value of animacy. Psychological Science. doi:10.1177/0956797613480803
New, J., Cosmides, L., & Tooby, J. (2007). Category-specific attention for animals reflects ancestral priorities, not expertise. Proceedings of the National Academy of Sciences, 104, 16598–16603. doi:10.1073/pnas.0703913104
New, B., Pallier, C., Brysbaert, M., & Ferrand, L. (2004). Lexique 2: A new French lexical database. Behavior Research Methods, Instruments, & Computers, 36, 516–524. doi:10.3758/BF03195598
Nouchi, R. (2013). Can the memory enhancement of the survival judgment task be explained by the elaboration hypothesis? Evidence from a memory load paradigm. Japanese Psychological Research, 55, 58–71. doi:10.1111/j.1468-5884.2012.00531.x
Öhman, A., Flykt, A., & Esteves, F. (2001a). Emotion drives attention: Detecting the snake in the grass. Journal of Experimental Psychology: General, 130, 466–478. doi:10.1037/0096-3445.130.3.466
Öhman, A., Lundqvist, D., & Esteves, F. (2001b). The face in the crowd revisited: A threat advantage with schematic stimuli. Journal of Personality and Social Psychology, 80, 381–396. doi:10.1037/0022-3514.80.3.381
Otgaar, H., Smeets, T., Merckelbach, H., Jelicic, M., Verschuere, B., Galliot, A. M., & van Riel, L. (2011). Adaptive memory: Stereotype activation is not enough. Memory & Cognition, 39, 1033–1041. doi:10.3758/s13421-011-0091-2
Paivio, A. (1971). Imagery and verbal processes. New York, NY: Holt, Rinehart & Winston.
Paivio, A. (2007). Mind and its evolution: A dual coding theoretical approach. Mahwah, NJ: Erlbaum.
Paivio, A., & Csapo, K. (1973). Picture superiority in free recall: Imagery or dual coding? Cognitive Psychology, 5, 176–206. doi:10.1016/0010-0285(73)90032-7
Pratt, J., Radulescu, P. V., Guo, R. M., & Abrams, R. A. (2010). It’s alive! animate motion captures visual attention. Psychological Science, 21, 1724–1730. doi:10.1177/0956797610387440
Raby, C. R., & Clayton, N. S. (2012). Episodic memory and planning. In T. Schackleford & J. Vonk (Eds.), The Oxford handbook of comparative evolutionary psychology (pp. 217–235). Oxford, UK: Oxford University Press.
Raftery, A. E. (1995). Bayesian model selection in social research. In P. V. Marsden (Ed.), Sociological methodology (pp. 111–196). Oxford, UK: Blackwell.
Rajaram, S. (1996). Perceptual effects on remembering: Recollective processes in picture recognition memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 22, 365–377. doi:10.1037/0278-7393.22.2.365
Salthouse, T. A., Toth, J. P., Hancock, H. E., & Woodard, J. L. (1997). Controlled and automatic forms of memory and attention: Process purity and the uniqueness of age-related influences. Journals of Gerontology, 52B, 216–P228.
Snodgrass, J. C., & Vanderwart, M. (1980). A standardized set of 260 pictures: Norms for names agreement, image agreement, familiarity, and visual complexity. Journal of Experimental Psychology: Human Learning and Memory, 6, 174–215. doi:10.1037/0278-7393.6.2.174
Spector, A., & Biederman, I. (1976). Mental set and mental shift revisited. American Journal of Psychology, 89, 669–679. doi:10.2307/1421465
Tulving, E. (1985). Memory and consciousness. Canadian Psychologist, 26, 1–12. doi:10.1037/h0080017
VanArsdall, J. E., Nairne, J. S., Pandeirada, J. N. S., & Blunt, J. R. (2013). Adaptive memory: Animacy processing produces mnemonic advantages. Experimental Psychology, 60, 172–178. doi:10.1027/1618-3169/a000186
Wheeler, M. A., Stuss, D. T., & Tulving, E. (1997). Toward a theory of episodic memory: The frontal lobes and autonoetic consciousness. Psychological Bulletin, 121, 331–354. doi:10.1037/0033-2909.121.3.331
Wiggett, A., Pritchard, I. C., & Downing, P. E. (2009). Animate and inanimate objects in human visual cortex: Evidence for task-independent category effects. Neuropsychologia, 47, 3111–3117. doi:10.1016/j.neuropsychologia.2009.07.008
Yang, J., Wang, A., Yan, M., Zhu, Z., Chen, C., & Wang, Y. (2012). Distinct processing for pictures of animals and objects: Evidence from eye movements. Emotion, 12, 540–551. doi:10.1037/a0026848
Author Note
Part of this work was presented at the 16 meeting of the European Society for Cognitive Psychology, Budapest, Hungary (August 2013). This work was supported by a grant from the Institut universitaire de France to the first author and by a grant from the Conseil Régional de Bourgogne to the third author. The authors thank James Nairne, Alice Healy, and Reed Hunt for their very constructive comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bonin, P., Gelin, M. & Bugaiska, A. Animates are better remembered than inanimates: further evidence from word and picture stimuli. Mem Cogn 42, 370–382 (2014). https://doi.org/10.3758/s13421-013-0368-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13421-013-0368-8