
Abstract
In three experiments, rats of different ages were trained in a circular pool to find a hidden platform whose location was defined in terms of a single landmark, a cylinder outside the pool. Following training, two main components of the landmark, its shape and pattern, were tested individually. Experiment 1 was performed by adolescent and adult rats (Exp. 1a, males; Exp. 1b, females). Adult rats always learned faster than the adolescent animals. On test trials, interesting tendencies were found—mainly, one favoring males on the shape test trial, and another favoring females on the pattern test trial. Experiment 2 was conducted only with adolescent rats, and these males and females did not differ when learning the task. However, on test trials the males learned more about the landmark shape component than about the landmark pattern component, while the females learned equally about the two components of the landmark. Finally, Experiment 3 was conducted only with adult rats, and again the males and females did not differ when learning the task. However, on test trials the males learned equally about the two components of the landmark (shape and pattern), but the females learned more about the landmark pattern component than about the landmark shape component. This set of experiments supports the claim that male and female rats can learn rather different things about a landmark that signals the location of the platform, with age being a critical variable.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
There is evidence that males and females often show a predisposition to pay attention to different aspects of the environment. An early demonstration in chicks is the study by Vallortigara (1996). Vallortigara trained young male and female chicks to discriminate between two boxes. The correct box was indicated by both a color cue and a position cue. Following training, the chicks were retrained to discriminate on the basis of either color (irrespective of position) or position (irrespective of color). During retraining, females performed better on the color-learning task, and males performed better on the position-learning task. Working with rodents, there have also been many demonstrations showing that male and female rats often rely on different cues to solve spatial problems (Hawley, Grissom, Barratt, Conrad, & Dohanich, 2012; Keeley, Tyndall, Scott, & Saucier, 2013; Rodríguez, Torres, Mackintosh, & Chamizo, 2010; Roof & Stein, 1999; Williams, Barnett, & Meck, 1990): Males are more likely to rely on the geometry of the environment (or apparatus), while females more likely rely on local cues. The same claim has also been made when working with humans, using a variety of tasks (Dabbs, Chang, Strong, & Milun, 1998; Galea & Kimura, 1993; Jones & Healy, 2006; Sandstrom, Kaufman, & Huettel, 1998; Saucier et al., 2002).
Equally interesting is the finding that age may have different effects in males and females when solving a spatial task (in rats, see Kanit et al., 2000; Krasnoff & Weston, 1976; Rodríguez, Chamizo, & Mackintosh, 2013; in humans performing virtual navigation, see Bohbot et al., 2012). The work by Krasnoff and Weston addressed sex differences in spontaneous wheel running and maze performance both pre- and postpuberty (adolescent and adults, respectively). No sex differences were found with adolescents in either task, whereas in adulthood, females performed better than males in wheel running, and males performed better than females in maze performance. In the Kanit et al. (2000) study (see also Kanit et al., 1998), adolescent and adult rats were trained in a Morris pool. After an acquisition phase, a test trial was conducted in which the platform was moved to another quadrant, and the time the animals spent in the quadrant where the platform had been during training as well as the time to reach the visible platform in the new place were both measured. The results showed that adult female rats went directly to the newly positioned visible platform, whereas adult males searched for the platform in its old location, as did both male and female adolescent rats. In the Rodríguez et al. (2013) study, rats were trained in a triangular-shaped pool to find a hidden platform, whose location was defined in terms of two sources of information, a landmark outside the pool and a particular corner of the pool. After acquisition, the rats were given test trials without the platform. On one test trial the two sources of information were put into opposition. Two other trials were single-cue test trials. In the test trials, males and females differed. The differences observed were mainly in their preferred mode of solution, with adult, but not adolescent, females using the landmark in preference to geometry, and males using geometry regardless of age. Also important and relevant for the present study was that males (both adolescents and adults) performed more accurately than did females on the single-cue geometry test trial; for the same result, see also Rodríguez et al. (2010). In both Kanit’s et al. (2000) and Rodríguez’s et al. (2013) work, adult female rats solved the task differently than did both adult male rats and adolescent rats, whether male or female. Therefore, a clear “age effect” in the strategies used by female rats to solve a navigation task was found in the two studies.
If the results reported by Kanit et al. (2000) and Rodríguez et al. (2013) represent a more general manifestation of the response patterns from male and female rats of different ages to geometrical and nongeometrical visual cues, similar results should also be found with respect to other, more restrictive tasks—for example, tasks combining geometrical and nongeometrical visual features or components of a single landmark. Experiment 1 (Exp. 1a, males; Exp. 1b, females) reports such a test of the possibility that age may have different effects on landmark learning—specifically, on what is learned about the two main components of a landmark, its shape and pattern (i.e., geometrical and nongeometrical visual information, respectively). In two subsequent experiments, Experiments 2 and 3, a direct comparison between male and female rats (Exp. 2, adolescents; Exp. 3, adults) was provided. No differences between the groups were expected in Experiment 1a and Experiment 2; however, differences were expected in Experiment 1b as well as in Experiment 3. Would that be the case?
Importantly, because it has recently been found that the specific characteristics of a landmark cue can play a crucial role in females’ preferences when solving a spatial task (Torres, Rodríguez, Chamizo, & Mackintosh, 2014; see also Chamizo, Rodríguez, Torres, Torres, & Mackintosh, 2014), in the present set of experiments the landmark used was carefully chosen (i.e., it was always a white cylinder entirely covered with black dots). The main requirement in choosing the landmark was that it would have the same appearance when approached from different directions. This is important when rats are trained to find a hidden platform in a pool, since they are put into the pool at different points on its circumference on different trials and, once in the pool, are free to swim anywhere. It seems very probable that they would approach the platform from different directions on different trials.
General method
We first describe the apparatus and general procedures common to all experiments.
Apparatus
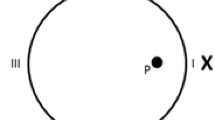
The apparatus was a circular swimming pool made of plastic and fiberglass and modeled after that used by R. G. M. Morris (1981). It measured 1.58 m in diameter and 0.4 m deep, and it was filled to a depth of 0.22 m with water rendered opaque by the addition of 1 cL/L of latex. The water temperature was maintained at 22 ± 1 °C. The pool was situated in the middle of a large room and mounted on a wooden platform 0.43 m above the floor. The pool was surrounded by black curtains reaching from the ceiling to the base of the pool and forming a circular enclosure 2.4 m in diameter. A single object, landmark X, was suspended from a black false ceiling inside this enclosure, 35 cm above the surface of the water and with its midline directly above the wall of the pool. For all subjects, landmark X was always a cylinder, 8.5 cm in diameter and 30 cm in height, entirely patterned with 32 black dots mixed up on the white surface (2-cm diameter for each dot). This single landmark was the only cue defining the location of the platform. To ensure that the rats would use this landmark, X, as the source of information to locate the platform, rather than any inadvertently remaining static room cues (such as noises from pipes and air conditioning), the landmark and the platform were semirandomly rotated with respect to the room (90°, 180°, 270°, or 360°) with the restriction that all four positions in the room were used each day. Two additional objects were also used: the same cylinder but plain light brown, and a white drum entirely patterned with black dots mixed up on a white surface, as in the training landmark (2-cm diameter for each dot; the drum was 21 cm in diameter and 10.5 cm in height). A closed-circuit video camera with a wide-angle lens was mounted 1.75 m above the center of the pool inside the false ceiling, and its picture was relayed to recording equipment in an adjacent room. A circular platform, 11 cm in diameter and made of white Perspex, was mounted on a base and could be placed in front of landmark X, 38 cm from the wall, with its top 1 cm below the surface of the water. The hidden platform, P, and landmark, X, were situated as is shown in Fig. 1a.

(a) Schematic representation of the pool and the position of the landmark (X) and the hidden platform (P) during Experiments 1–3. I, II, III, and IV indicate the four starting positions. (b) Schematic representation of the pool and the positions of the landmark (X), the target area (T), and the control area (C) during the test trials. I and II indicate the starting positions.
General procedure
There were four types of trials: pretraining, preexposure, training, and test trials. Pretraining consisted of placing a rat into the circular pool without the landmark but with the hidden platform present. The rat was given 120 s to find the platform, and once the rat had found it, it was allowed to stay on it for 30 s. If it had not found the platform within the 120 s, it was picked up, placed on it, and left there for 30 s. The platform was moved from one trial to the next, and the rat was placed in the pool in a different location on each trial, as far as possible equally often on the same or the opposite side of the pool from the platform, and with the platform to the right or left of where the rat was placed. Rats were given five such pretraining trials over two days, with two trials on Day 1 and three on Day 2. Rats were run in squads of eight and spent the intertrial interval (ITI) in small individual compartments. After pretraining, preexposure trials took place during one day in an open compartment (specifically, a round soup dish 21 cm in diameter and 3 cm in depth), which was always situated in the center of the pool. This compartment was placed on the platform with its base approximately at the water level. The animals were preexposed to each of the three objects used in the present experiments (cylinder with dots—the training landmark X—a brown cylinder, and a drum with dots) twice, individually (i.e., six trials in total; order of presentation of the objects: A, B, C, C, B, A). A preexposure trial consisted of placing a rat in this open compartment and leaving it there for 1 min facing the single object, which was always placed above the edge of the pool. The average ITI of the preexposure trials was 8–10 min.
The procedure for training was similar to that of pretraining, with the exception that the landmark, X, was always present. As in pretraining, the rat was placed in the pool in a different location on each trial, equally often with the platform to the right, to the left, or in front of (near or far) where the rat was placed (at positions I, II, III, and IV in Fig. 1a). Rats were given eight trials per day over three days (a total of 24 trials). These trials had an ITI of 8–10 min, and the platform and the landmark were rotated between trials. Finally, there were two test days, with eight training trials (identical to those in the training phase), followed by one test trial without the platform, 60 s long. Rats received a different test trial each day, counterbalanced, one with the brown cylinder (i.e., the shape component of the training landmark), and the other with the drum with dots (i.e., the pattern component of the training landmark). For purposes of recording the rat’s behavior, on these trials the amounts of time that the rats spent in two different but identically sized areas (target and control) were recorded. These areas were 22 cm in diameter—twice the hidden platform diameter. Each rat was placed in the pool from one specific position (at I and II in Fig. 1b), and rats were, as much as possible, placed equally in the two starting positions. An alpha level of .05 was adopted for all statistical analyses. Only significant results are presented.
Experiment 1
The aim of Experiment 1 (Exp. 1a, males; Exp. 1b, females) was to provide a direct comparison between two groups of same-sex rats: adolescents and adults. The experiment was conducted to answer the following questions: Would the rats learn the spatial task equally fast, despite having such different ages? And would the two age groups learn equally about the two components of the landmark, shape and pattern?
Method
Subjects
The subjects were naive Long Evans rats (Rattus norvegicus), ten adolescent and ten adult males in Experiment 1a, and ten adolescent and ten adult females in Experiment 1b. The adolescent rats were approximately one month old at the beginning of the experiments, and the adult rats were approximately three months old at the beginning of the experiments. The animals were housed in standard cages, 25 × 15 × 50 cm, in groups of two (same sex, same age) and were maintained on ad lib food and water, in a colony room with a 12:12-h light:dark cycle. They were tested within the first 8 h of the light cycle.
Results and discussion
All of Experiment 1 (Exps. 1a and 1b) was analyzed together. Latencies (with standard errors of the means [SEMs]) to find the platform decreased over the course of the five initial pretraining trials. Adolescent males decreased from a mean of 113.0 s (6.97) on Trial 1 to a mean of 80.6 s (11.88) on Trial 5; adult males decreased from a mean of 106.2 s (9.94) on Trial 1 to a mean of 54.7 s (11.74) on Trial 5; adolescent females decreased from a mean of 120.0 s (0.00) on Trial 1 to a mean of 84.9 s (11.00) on Trial 5; and adult females decreased from a mean of 103.7 s (6.78) on Trial 1 to a mean of 67.7 s (13.09) on Trial 5. An analysis of variance (ANOVA) conducted on these data—taking into account the variables trials (1–5), sex (males, females), and age (adolescent, adult)—showed that the variables trials, F(4, 144) = 13.24 (p < .001, ηp2 = .27, 95% CIs = 0.14, 0.36; power > .99), and age, F(1, 36) = 22.63 (p < .001, ηp2 = .39, 95% CIs = 0.14, 0.56; power > .99), were significant. No other main effect or interaction was significant, minimum F(4, 144) = 0.36 (p = .835). All rats improved their performance as pretraining trials progressed, and adult rats reached the platform significantly faster than adolescent rats.
Latencies (with SEMs) to find the platform also decreased over the course of the training days (see Fig. 2a for Exp. 1a, and Fig. 2b for Exp. 1b). An ANOVA conducted on these data—taking into account the variables days (1–3), sex (males, females), and age (adolescent, adult)—showed that the variables days, F(2, 72) = 135.88 (p < .001, ηp2 = .79, 95% CIs = 0.70, 0.84; power > .99), and age, F(1, 36) = 50.07 (p < .001, ηp2 = .58, 95% CIs = 0.35, 0.71; power > .99), were again significant. However, the interactions Days × Age, F(2, 72) = 3.11 (p = .051), and Age × Sex, F(1, 36) = 3.15 (p = .084), were nearly significant. No other main effect or interaction was significant, minimum F(2, 72) = 0.46 (p = .633). An ANOVA conducted on the escape trials on the two test days (with the variables days, sex, and age) showed that the three variables were significant: days, F(1, 36) = 9.37 (p = .004, ηp2 = .21, 95% CIs = 0.02, 0.41; power = .85); sex, F(1, 36) = 6.99 (p = .012, ηp2 = .16, 95% CIs = 0.01, 0.37; power = .73); and age, F(1, 36) = 80.04 (p < .001, ηp2 = .69, 95% CIs = 0.49, 0.78; power > .99). However, no interaction was significant, minimum F(1, 36) = 0.19 (p = .664). Although all rats improved their performance as the experiment progressed, adult animals reached the platform faster than adolescents, and males faster than females.


(a) Mean time spent in the target and control areas by the subjects during the two test trials of Experiment 1a (males). Error bars denote standard errors of the means. (b) Mean time spent in the target and control areas by the subjects during the two test trials of Experiment 1b (females). Error bars denote standard errors of the means.
Figure 3 (Fig. 3a for Exp. 1a, and 3b for Exp. 1b) shows the time spent in the target and control areas by the groups during the two test trials (i.e., one with the brown cylinder, the shape component of the training landmark in the absence of the pattern component; and the other with the drum with dots, the pattern component of the training landmark with a different shape). Student’s t tests were used to compare the rats’ performance in each target area with the corresponding control area. On both test trials, all rats spent a significantly longer time in the target than in the control area [minimum t(9) = 3.18, and all ps < .02, d = 1.01, 95% CIs = 0.22, 1.76]. The results above imply that all rats (both males and females, adolescents and adults) had learned about the two components of the landmark (shape and pattern). An ANOVA was conducted on the time spent in the target area on the two test trials, taking into account the variables type of component tested (shape, pattern), sex (males, females), and age (adolescent, adult). The results showed that the age variable was significant, F(1, 36) = 8.94 (p = .005, ηp2 = .20, 95% CIs = 0.02, 0.40; power = .83), as well as the interactions Type of Component Tested × Sex, F(1, 36) = 4.26 (p = .046, ηp2 = .11, 95% CIs = 0.00, 0.30; power = .52), and Type of Component Tested × Age, F(1, 36) = 5.90 (p = .020, ηp2 = .14, 95% CIs = 0.00, 0.34; power = .66). The analysis of the Type of Component Tested × Sex interaction showed only two tendencies: one favoring males on the shape test trial, F(1, 38) = 3.42 (p = .072), and the second one showing that females had better performance on the pattern test trial than on the shape test trial, F(1, 19) = 3.96 (p = .061). The analysis of the Type of Component Tested × Age interaction revealed that adult rats performed better on the pattern test trial than did adolescent rats, F(1, 38) = 14.07 (p = .001, ηp2 = .27, 95% CIs = 0.06, 0.46; power = .96). In addition, adult rats showed a tendency favoring superior performance on the pattern test trial than on the shape test trial, F(1, 19) = 3.32 (p = .084).
In conclusion, Experiment 1 tentatively supports the claim that, depending on age, male and female rats could learn rather different things about the two components (shape and pattern) of a landmark that signals the location of a hidden platform.
Following this experiment, Experiments 2 and 3 addressed the sex variable in two different age groups (adolescents and adults, respectively), in an attempt to optimize the number of subjects per experiment (Russell & Burch, 1959).
Experiment 2
The aim of Experiment 2 was to provide a direct comparison between two groups of adolescent rats: males and females. The experiment was conducted to answer the following questions: Would the two sexes learn the spatial task equally fast? Would females learn equally about the two components of the training landmark, as Experiment 1b suggested? And would males learn more about the shape component than about the pattern component of the training landmark, as Experiment 1a suggested?
Method
Subjects and apparatus
The subjects were 24 naive Long Evans rats, 12 males and 12 females, approximately one month old at the beginning of the experiment. The animals were kept and maintained as in Experiment 1. The apparatus (a Morris pool) and the objects used in this experiment [the cylinder with black dots (landmark X), the brown cylinder, and the drum with dots] were also exactly the same as in Experiment 1.
Procedure
The general procedure was exactly the same as that used in Experiment 1, with one important exception: In this experiment, only adolescent rats (males and females) were used.
Results and discussion
Latencies (with SEMs) to find the platform decreased over the course of the five initial pretraining trials. Males decreased from a mean of 120.0 s (0.0) on Trial 1 to a mean of 69.1 s (11.50) on Trial 5, and females decreased from a mean of 107.2 s (8.79) on Trial 1 to a mean of 72.7 s (10.48) on Trial 5. An ANOVA conducted on these data, taking into account the variables trials (1–5) and sex (male, female), showed that the only significant variable was trials, F(4, 88) = 5.82 (p < .001, ηp2 = .21, 95% CIs = 0.05, 0.32; power = .98). No other main effect or interaction was significant, minimum F(1, 22) = 0.03 (p = .857). Thus, both males and females improved their performance as pretraining trials progressed, reaching the platform equally fast.
Latencies (with SEMs) to find the platform also decreased over the course of the training days (see Fig. 4a). An ANOVA conducted on these data, taking into account the variables days (1–3) and sex (male, female), showed that the only significant variable was days, F(2, 44) = 132.44 (p < .001, ηp2 = .86, 95% CIs = 0.76, 0.90; power > .99). No other main effect or interaction was significant, minimum F(2, 44) = 0.04 (p = .963). An ANOVA conducted on the escape trials of the two test days (with the variables days and sex) showed that no main effect or interaction was significant, minimum F(1, 22) = 0.02 (p = .893). Both males and females improved their performance as the experiment progressed, reaching the platform equally fast.

Experiment 2. (a) Mean escape latencies for the groups during the training and test days. Error bars denote standard errors of the means. (b) Mean time spent in the target and control areas by the subjects during the two test trials. Error bars denote standard errors of the means.
Figure 4b shows the time spent in the target and control areas by the groups during the two test trials (i.e., one with the brown cylinder and the other with the drum with dots—matching the shape and pattern components of the training landmark, respectively). Student’s t tests were used to compare rats’ performance in each target area with the corresponding control area. On both test trials, male and female adolescent rats spent a significantly longer time in the target than in the control area [minimum t(11) = 4.34, and all ps < .005, d = 1.25, 95% CIs = 0.47, 2.00]. This implies that all rats had learned about both components of the landmark (shape and pattern). An ANOVA conducted on the time spent in the target area on the two test trials, taking into account the variables type of component tested (shape, pattern) and sex (male, female), showed that the variables type of component tested, F(1, 22) = 8.65 (p = .008, ηp2 = .28, 95% CIs = 0.24, 0.52; power = .80), and sex, F(1, 22) = 12.26 (p = .002, ηp2 = .36, 95% CIs = 0.06, 0.57; power = .92) were significant, as well as the Type of Component Tested × Sex interaction, F(1, 22) = 21.67 (p < .001, ηp2 = .50, 95% CIs = 0.17, 0.67; power = .99). The analysis of the interaction Type of Component Tested × Sex revealed that males performed better than females in the shape test trial, F(1, 22) = 22.38 (p < .001, ηp2 = .50, 95% CIs = 0.18, 0.68; power > .99). Moreover, male rats performed better in the shape than in the pattern test trial, F(1, 11) = 21.84 (p < .001, ηp2 = .67, 95% CIs = 0.21, 0.81; power = .99). These results show that male rats had performed more accurately in the shape than in the pattern test trial, while female rats performed equally well in both test trials.
In conclusion, when the two landmark components (shape and pattern) were tested one by one, Experiment 2 showed that males learned more about the shape than about the pattern component (as Exp. 1a had suggested), and females learned equally about the two landmark components (as Exp. 1b had suggested). In addition, a clear male advantage was found when learning the shape component, but the two sexes did not differ when learning the pattern component.
Experiment 3
The aim of Experiment 3 was to provide a direct comparison between two groups of adult rats: males and females. The experiment was conducted to answer the following questions. Would the two sexes learn the spatial task equally fast? Would males learn equally about the two components of the training landmark, shape and pattern, as Experiment 1a suggested? And would females learn more about the pattern than about the shape component of the training landmark, as Experiment 1b suggested?
Method
Subjects and apparatus
The subjects were 36 naive Long Evans rats, 18 males and 18 females, approximately three months old at the beginning of the experiment. The animals were kept and maintained as in the previous experiments. The apparatus (a Morris pool) and the objects used in this experiment [the cylinder with black dots (landmark X), the brown cylinder, and the drum with dots] were exactly the same as in Experiments 1 and 2. [Initially, this experiment was carried out with groups of 12 rats (Replication 1). However, on the test trials without the platform, although the main variables sex and component were significant, the interaction did not reach significance (p = .11). Then six more rats per group were added (Replication 2), to see whether the interaction might be significant, as was the case.]
Procedure
The general procedure was the same as in Experiments 1 and 2, although with two important exceptions: In this experiment, only adult rats (males and females) were used, and second, this experiment was conducted in two replications.
Results and discussion
All statistical analyses were carried out with the variable replication. However, because this variable was neither significant nor interacted with another variable in either the acquisition or test trials, the results and statistics for this experiment are reported without this variable, in order to make them less cumbersome.
Latencies (with SEMs) to find the platform decreased over the course of the five initial pretraining trials. Males decreased from a mean of 79.9 s (10.13) on Trial 1 to a mean of 46.5 s (10.17) on Trial 5, while females decreased from a mean of 99.7 s (8.44) on Trial 1 to a mean of 57.9 s (10.32) on Trial 5. An ANOVA conducted on these data, taking into account the variables trials (1–5) and sex (male, female), showed that the only significant variable was trials, F(4, 136) = 4.60 (p = .002, ηp2 = .12, 95% CIs = 0.02, 0.20; power = .94). No other main effect or interaction was significant, minimum F(4, 136) = 0.13 (p = .973). Both males and females improved their performance as the pretraining trials progressed, reaching the platform equally fast.
Latencies (with SEMs) to find the platform also decreased over the course of the training days (see Fig. 5a). An ANOVA conducted on these data, taking into account the variables days (1–3) and sex (male, female), showed that the only significant variable was days, F(2, 68) = 27.52 (p < .001, ηp2 = .45, 95% CIs = 0.26, 0.57; power > .99). No other main effect or interaction was significant, minimum F(2, 68) = 0.29 (p = .746). An ANOVA conducted on the escape trials of the two test days (with the variables days and sex) showed that the only significant variable was days, F(1, 34) = 8.02 (p = .008, ηp2 = .19, 95% CIs = 0.01, 0.40; power = .79). No other main effect or interaction was significant, minimum F(1, 34) = 1.19 (p = .283). Both males and females improved their performance as the experiment progressed, reaching the platform equally fast.

Experiment 3. (a) Mean escape latencies for the groups during the training and test days. Error bars denote standard errors of the means. (b) Mean time spent in the target and control areas by the subjects during the two test trials. Error bars denote standard errors of the means.
Figure 5b shows the time spent in the target and control areas by the groups during the two test trials (i.e., one with the brown cylinder and the other with the drum with dots—matching the shape and pattern components of the training landmark, respectively). Student’s t tests were used to compare the rats’ performance in each target area with the corresponding control area. The time in the target areas differed significantly from that in the control areas on all test trials [minimum t(17) = 6.46, and all ps < .001, d = 1.52, 95% CIs = 0.83, 2.20], with the exception of females on the shape test trial, t(17) = 1.41 (p = .178). This implies that all rats had learned about both components of the landmark (shape and pattern), except for the females, who only learned about the pattern component. An ANOVA conducted on the time spent in the target area on the two test trials, taking into account the variables type of component tested (shape, pattern) and sex (male, female), showed that both the variables type of component tested, F(1, 34) = 18.87 (p < .001, ηp2 = .36, 95% CIs = 0.11, 0.54; power = .99), and sex, F(1, 34) = 8.23 (p = .007, ηp2 = .20, 95% CIs = 0.02, 0.40; power = .80), were significant, as well as the Type of Component Tested × Sex interaction, F(1, 34) = 7.75 (p = .009, ηp2 = .19, 95% CIs = 0.01, 0.39; power = .77). The analysis of the interaction Type of Component Tested × Sex revealed that males performed better than females in the shape test trial, F(1, 34) = 15.40 (p < .001, ηp2 = .31, 95% CIs = 0.08, 0.51; power = .97). Moreover, females performed better in the pattern test trial than in the shape test trial, F(1, 17) = 19.23 (p < .001, ηp2 = .53, 95% CIs = 0.16, 0.71; power = .99).
In conclusion, when the two landmark components (shape and pattern) were tested one by one, Experiment 3 showed that males learned equally about the two landmark components, shape and pattern (as had been suggested in Exp. 1a), and females learned more about the pattern than about the shape component (as had been suggested in Exp. 1b). In addition, as in Experiment 2, a clear male advantage was found when learning the shape component, but the two sexes did not differ when learning the pattern component.
General discussion
In all the present experiments, a submerged platform was located near the wall of a circular pool, in front of a landmark. Circular black curtains surrounded the pool, which was rotated from trial to trial, along with the landmark and the platform, so that the only indicator of the location of the platform was the landmark. Once the rats had learned to find the platform, they were given two test trials without the platform, on which the two main components of the landmark (its shape and pattern) were individually presented: On one, the landmark had a different pattern (thus testing shape, the geometric component of the training landmark), and on the other, the pattern was presented on a different shape (thus testing pattern, the nongeometric component of the training landmark). The present study showed that adult males performed equally well when tested on both components, whereas adult females performed much less accurately when tested with the geometric component of the training landmark. However, adolescent males performed much less accurately when tested with the pattern component of the training landmark, whereas adolescent females behaved like adult males (i.e., they learned equally about the two components of the landmark), but unlike adult females, when searching for the platform in the Morris pool.
It is worth noting that the present experiments with adolescent rats (Exps. 1a, 1b, and 2) took place when the rats were between 31 and 39 days old, before the period of vaginal opening (around the 42nd day). Moreover, in relation to hormonal changes that appear at the onset of puberty, there is reason to believe (see Pleil & Williams, 2010; Williams & Meck, 1991) that the critical factor is the rats’ hormonal state during acquisition rather than at the time of test. The literature on sex differences in spatial navigation (and, for humans, in mental rotation) has largely focused on differences in speed of learning or accuracy of performance. There was no suggestion of any such differences in the present experiments when males and females were directly compared (Exps. 2 and 3). Male and female rats did not differ during the course of initial training (for the same result, see also Rodríguez et al., 2013). However they differed on test trials. In both adolescence (Exp. 2) and adulthood (Exp. 3), a sex difference was found when landmark learning. In males, selective learning of the shape component was evident only in adolescence, while in females selective learning of the pattern component was observed only in adulthood. Therefore, age seems to determine selective learning of one or the other component of the training landmark in both males and females. We are forced to conclude that the two sexes, both when adolescent and when adults, differed in the amounts learned about the two main components (shape and pattern) of a landmark that signaled the location of a platform (for related results, see Kanit et al., 2000; Rodríguez et al., 2013). Most importantly, the present set of experiments demonstrated for the first time that the differential response patterns from males and females of different ages can apply to the geometrical and nongeometrical components of a single landmark (its shape and pattern).
When rats are trained in a circular pool to find a hidden platform whose location is defined in terms of a single landmark outside the pool, Chamizo et al. (2014) have shown that adult male and female rats do not seem to learn the same things about the landmark (for a similar result, see also Torres et al., 2014). Chamizo et al. found that the nature of the landmark affected only the behavior of females, and not that of males. In this study, two identical cylinders were used as landmarks, one plain white and the other divided into four vertical segments, each patterned differently. Adult male rats learned about the four-pattern cylinder as rapidly as about the cylinder with a single pattern. However, that was not the case with the females. Adult female rats learned to swim to the platform more rapidly with the plain white cylinder than with the four-pattern cylinder. Chamizo et al. concluded that male rats, since they are less likely than females to rely on landmarks to solve a spatial problem, do not pay very close attention to the specific features of any landmark—that they simply learned that the platform was to be found next to the only large object suspended above the pool. The test results in the present study with adult male rats contrast sharply with this previous conclusion. They show that adult male rats pay attention to the main components (shape, pattern) of a single landmark. Other studies have indeed shown that, depending on the salience of the cues, adult male rats may in some circumstances prefer to use the visual features of a landmark rather than its spatial location (e.g., Arain & Cohen, 2013) or rather than the local geometry provided by the apparatus (Mesa, Osorio, Ballesta, Marimon, & Chamizo, 2017).
What is the explanation of the changes in the behavior of rats, both males and females, as they grow older? It could be argued that the two sexes differ in cognitive styles when adolescent and when adults, thus learning differently about the two main components (shape and pattern) of a landmark that signaled the location of a platform. Selective landmark learning was found in the two sexes, but at different ages—in males, in adolescence; but in females, in adulthood. In males (but not females), a natural predisposition to geometry could explain the results of Experiment 2 (Izard, Pica, Dehaene, Hinchey, & Spelke, 2011). If this is correct, then Experiment 3 reveals that such a predisposition seems to disappear with age. In any case, it is clear that the present results offer little support for the claim that animals recognize objects primarily on the basis of their shapes, regardless of the task demands (Izard et al., 2011). In females, the obvious answer, suggested by the observation that ovariectomized females behave like younger rather than like older rats (Rodríguez et al., 2013), is that the hormonal changes associated with the onset of puberty change the females’ learning and attention. In both males and females, age could capture different aspects of the landmark, although we do not know exactly why. Sex differences in cognitive patterns probably arose because they proved to be evolutionarily advantageous (for reviews in spatial cognition, see Halpern, 2012; Jones, Braithwaite, & Healy, 2003; Mackintosh, 2011).
In the literature on spatial abilities, it has been repeatedly said that sex differences, in both rodents and humans, appear after puberty (for reviews, see Kimura, 1999; Voyer, Voyer, & Briden, 1995). However, recent research carried out with babies using mental rotation tasks with 3-D objects (Lauer, Udelson, Jeon, & Lourenco, 2015; D. S. Moore & Johnson, 2008; Quinn & Liben, 2008, 2014) has shown that the previous statement is inaccurate. Undoubtedly, more research will be needed to understand how spatial tasks (or other tasks with spatial components) interact with sex differences throughout the life cycle. For example, the different natural selection pressures exerted over centuries on males and females of different species are a subject that will have to be addressed more in the future, given its many implications (Lauer & Lourenco, 2016).
According to Mackintosh (1975), the attention paid to a cue increases if it is a better predictor of the outcome than all other cues present on a trial, and decreases if it is no better a predictor of the outcome than all the other cues present on a trial. Thus, cues that are good predictors of subsequent events will enjoy an increase in their salience (i.e., an increase in their attention). Irrelevant cues that are poor predictors of subsequent events, however, suffer a reduction in their attention (see also Esber & Haselgrove, 2011; Le Pelley, 2004). We believe that the previous analysis can also be applied to the two main components (shape and pattern) of a landmark. Most importantly, both sex and age seem able to determine the gains or losses in the attention given to these components.
Author note
This research was supported by a grant from the Spanish Ministerio de Ciencia e Innovación (Ref. PSI2013-47430-P) to V.D.C.
References
Arain, M., & Cohen, J. (2013). Hierarchical use of cues in the missing object recognition task by rats. Behavioural Processes, 97, 41–52.
Bohbot, V. D., McKenzie, S., Konishi, K., Fouquet, C., Kurdi, V., Schachar, R., . . . Robaey, P. (2012). Virtual navigation strategies from childhood to senescence: Evidence for changes across the life span. Frontiers in Aging Neuroscience, 4, 28. doi:https://doi.org/10.3389/fnagi.2012.00028
Chamizo, V. D., Rodríguez, C. A., Torres, I., Torres, M. N., & Mackintosh, N. J. (2014). What makes a landmark effective? Sex differences in a navigation task. Learning & Behavior, 42, 348–356. doi:https://doi.org/10.3758/s13420-014-0152-z
Dabbs, J. M., Chang, E. L., Strong, R. A., & Milun, R. (1998). Spatial ability, navigation strategy, and geographic knowledge among men and women. Evolution and Human Behavior, 19, 89–98.
Esber, G. R., & Haselgrove, M. (2011). Reconciling the influence of predictiveness and uncertainty on stimulus salience: A model of attention in associative learning. Proceedings of the Royal Society B, 278, 2553–2561. doi:https://doi.org/10.1098/rspb.2011.0836
Galea, L. A. M., & Kimura, D. (1993). Sex differences in route learning. Personality and Individual Differences, 14, 53–65.
Halpern, D. F. (2012). Sex differences in cognitive abilities (4th ed.). Hillsdale, NJ: Erlbaum.
Hawley, W. R., Grissom, E. M., Barratt, H. E., Conrad, T. S., & Dohanich, G. P. (2012). The effects of biological sex and gonadal hormones on learning strategy in adult rats. Physiology & Behavior, 105, 1014–1020. doi:https://doi.org/10.1016/j.physbeh.2011.11.021
Izard, V., Pica, P., Dehaene, S., Hinchey, D., & Spelke, E. (2011). Geometry as a universal mental construction. In S. Dehaene & E. M. Brannon (Eds.), Space, time and number in the brain: Searching for the foundations of mathematical thought (pp. 319–332). San Diego, CA: Academic Press.
Jones, C. M., & Healy, S. D. (2006). Differences in cue use and spatial memory in men and women. Proceedings of the Royal Society B, 273, 2241–2247.
Jones, C. M., Braithwaite, V. A., & Healy, S. D. (2003). The evolution of sex differences in spatial ability. Behavioral Neuroscience, 117, 403–411.
Kanit, L., Taskiran, D., Furedy, J. J., Kulali, B., McDonald, R., & Pöğün, S. (1998). Nicotine interacts with sex in affecting rat choice between “look-out” and “navigational” cognitive styles in the Morris water maze place learning task. Brain Research Bulletin, 46, 441–445.
Kanit, L., Taskiran, D., Yilmaz, Ö. A., Balkan, B., Demirgören, S., Furedy, J. J., & Pögün, S. (2000). Sexually dimorphic cognitive style in rats emerges after puberty. Brain Research Bulletin, 52, 243–248.
Keeley, R. J., Tyndall, A. V., Scott, G. A., & Saucier, D. M. (2013). Sex difference in cue strategy in a modified version of the Morris water task: correlations between brain and behaviour. PLoS ONE, 8, e69727. doi:https://doi.org/10.1371/journal.pone.0069727
Kimura, D. (1999). Sex and cognition. Cambridge, MA: MIT Press.
Krasnoff, A., & Weston, L. M. (1976). Puberal status and sex differences: activity and maze behavior in rats. Developmental Psychobiology, 9, 261–269.
Lauer, J. E., Udelson, H. B., Jeon, S. O., & Lourenco, S. F. (2015). An early sex difference in the relation between mental rotation and object preference. Frontiers in Psychology, 6, 558. doi:https://doi.org/10.3389/fpsyg.2015.00558
Lauer, J. E., & Lourenco, S. F. (2016). Spatial processing in infancy predicts both spatial and mathematical aptitude in childhood. Psychological Science, 27, 1291–1298.
Le Pelley, M. E. (2004). The role of associative history in models of associative learning: A selective review and a hybrid model. Quarterly Journal of Experimental Psychology, 57B, 193–243.
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological Review, 82, 276–298. doi:https://doi.org/10.1037/h0076778
Mackintosh, N. J. (2011). IQ and human intelligence (2nd ed.). Oxford, UK: Oxford University Press.
Mesa, V., Osorio, A., Ballesta, S., Marimon, J. M., & Chamizo, V. D. (2017). Geometric vs. non-geometric information: Explaining male rats’ selective preferences in a navigation task. Learning and Motivation, 60, 23–33.
Moore, D. S., & Johnson, S. P. (2008). Mental rotation in human infants: A sex difference. Psychological Science, 19, 1063–1066. doi:https://doi.org/10.1111/j.1467-9280.2008.02200.x
Morris, R. G. M. (1981). Spatial localization does not require the presence of local cues. Learning and Motivation, 12, 239–260.
Pleil, K. E., & Williams, C. L. (2010). The development and stability of estrogen-modulated spatial navigation strategies in female rats. Hormones and Behavior, 57, 360–367.
Quinn, P. C., & Liben, L. S. (2008). A sex difference in mental rotation in young infants. Psychological Science, 19, 1067–1070.
Quinn, P. C., & Liben, L. S. (2014). A sex difference in mental rotation in infants: Convergent evidence. Infancy, 19, 103–116.
Rodríguez, C. A., Chamizo, V. D., & Mackintosh, N. J. (2013). Do hormonal changes that appear at the onset of puberty determine the strategies used by female rats when solving a navigation task? Hormones and Behavior, 64, 122–135.
Rodríguez, C. A., Torres, A., Mackintosh, N. J., & Chamizo, V. D. (2010). Sex differences in the strategies used by rats to solve a navigation task. Journal of Experimental Psychology: Animal Behavior Processes, 36, 395–401.
Roof, R. L., & Stein, D. G. (1999). Gender differences in Morris water maze performance depend on task parameters. Physiology & Behavior, 68, 81–86.
Russell, W. M. S., & Burch, R. L. (1959). The principles of humane experimental technique (1992 reprint). Wheathampstead, UK: Universities Federation for Animal Welfare.
Sandstrom, N. J., Kaufman, J., & Huettel, S. A. (1998). Males and females use different distal cues in a virtual environment navigation task. Cognitive Brain Research, 6, 351–360.
Saucier, D. M., Green, S. M., Leason, J., MacFadden, A., Bell, S., & Elias, L. J. (2002). Are sex differences in navigation caused by sexually dimorphic strategies or by differences in the ability to use the strategies? Behavioral Neuroscience, 116, 403–410.
Torres, M. N., Rodríguez, C. A., Chamizo, V. D., & Mackintosh, N. J. (2014). Landmark vs. geometry learning: Explaining female rats’ selective preference for a landmark. Psicológica, 35, 81–100.
Vallortigara, G. (1996). Learning of colour and position cues in domestic chicks: Males are better at position, females at colour. Behavioural Processes, 36, 289–296.
Voyer, D., Voyer, S., & Bryden, M. P. (1995). Magnitude of sex differences in spatial abilities: A meta-analysis and consideration of critical variables. Psychological Bulletin, 117, 250–270. doi:https://doi.org/10.1037/0033-2909.117.2.250
Williams, C. L., Barnett, A. M., & Meck, W. H. (1990). Organizational effects of early gonadal secretions on sexual differentiation in spatial memory. Behavioral Neuroscience, 104, 84–97.
Williams, C. L., & Meck, W. H. (1991). The organizational effects of gonadal steroids on sexually dimorphic spatial ability. Psychoneuroendocrinology, 16, 155–176.
Author information
Authors and Affiliations
Corresponding author
Additional information
Professor Mackintosh has died
Electronic supplementary material
ESM 1
(XLSX 30 kb)
Rights and permissions
About this article

Cite this article
Chamizo, V.D., Torres, M.N., Rodríguez, C.A. et al. What makes a landmark effective in adolescent and adult rats? Sex and age differences in a navigation task. Learn Behav 47, 156–165 (2019). https://doi.org/10.3758/s13420-018-0364-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-018-0364-8