
Abstract
Both empirical and anecdotal evidence supports the idea that choice is preferred by humans. Previous research has demonstrated that this preference extends to nonhuman animals, but it remains largely unknown whether animals will actively seek out or prefer opportunities to choose. Here we explored the issue of whether capuchin and rhesus monkeys choose to choose. We used a modified version of the SELECT task—a computer program in which monkeys can choose the order of completion of various psychomotor and cognitive tasks. In the present experiments, each trial began with a choice between two icons, one of which allowed the monkey to select the order of task completion, and the other of which led to the assignment of a task order by the computer. In either case, subjects still had to complete the same number of tasks and the same number of task trials. The tasks were relatively easy, and the monkeys responded correctly on most trials. Thus, global reinforcement rates were approximately equated across conditions. The only difference was whether the monkey chose the task order or it was assigned, thus isolating the act of choosing. Given sufficient experience with the task icons, all monkeys showed a significant preference for choice when the alternative was a randomly assigned order of tasks. To a lesser extent, some of the monkeys maintained a preference for choice over a preferred, but computer-assigned, task order that was yoked to their own previous choice selection. The results indicated that monkeys prefer to choose when all other aspects of the task are equated.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Humans have long pondered the nature of choice and its relevance to freedom, control, and life in general. Popular belief is that having more choice and control over one’s environment is positive and preferred (Iyengar & Lepper, 2000). A body of literature supports the idea that choice has some benefits (e.g., Greenberger, Strasser, Cummings, & Dunham, 1989; Winocur, Moscovitch, & Freedman, 1987; Zuckerman, Porac, Lathin, Smith, & Deci, 1978), and there is considerable empirical support that not having control of one’s environment leads to behavioral and physiological problems (e.g., Mineka & Hendersen, 1985), such as learned helplessness (Overmier & Seligman, 1967). Even when choice does not have an explicit benefit, research suggests that humans prefer having a choice for its own sake—or choice qua choice—and it has been argued that this desire for choice is a biological imperative (see Leotti, Iyengar, & Oschner, 2010, for a review). Psychological and contextual factors may strongly influence and interact with the suggested biological drive for choice, and one possible way to begin to understand their complex interplay is to study preference for choice in nonhuman animals.
Some research has supported that animals also may prefer having choice. Voss and Homzie (1970) found that rats selected a path in a maze that offered a choice of routes over a direct one, even though both led to the same outcome. Research in pigeons has yielded a similar outcome—animals prefer choice for the sake of choice. In a concurrent chain procedure in which an initial choice between keys will lead to either a free choice of keys to peck or a forced choice, subjects prefer the option that leads to the free choice scenario (Catania, 1975; Catania & Sagvolden, 1980). Catania and Sagvolden emphasized the relevance of a specific issue in testing preference for choice in animals. Free-choice versus forced-choice conditions depend on the availability of those alternatives; thus, there is a natural confound between more “choice” and number of stimuli, novelty, variety, and so forth (Catania & Sagvolden, 1980). As an example, if one wanted to test whether an animal preferred a set of one food type (e.g., a bowl of bananas) versus the choice of food types in another set (e.g., a bowl with bananas and apples), the “choice” set would also be more variable, more perceptually distinct, and so forth. The selection of the “choice” between food items might actually reflect a preference for one of the other attributes that are associated with choice, and not the act of choosing itself. To address this potential issue, Catania and Sagvolden presented pigeons with two initial keys. A peck to one of these keys led to one of the following terminal key arrangements: (1) three green keys that yielded fixed interval reinforcement and one red extinction key, or (2) three red extinction keys and one green key that yielded fixed interval reinforcement. Thus the stimulus number (four keys), variety (three keys of one color and one of another) and information availability (two colors) were controlled for in the terminal keys. However, pigeons continued to show a preference for free choice even with these variables controlled. The authors concluded that this preference may be linked to learning or an evolutionary explanation in which organisms that prefer more options and choice would have a selective advantage.
Another study of pigeons used a self-control task and found that the birds preferred the choice between a larger-later and smaller-sooner response if the smaller-sooner link presented a reasonable outcome. If the smaller-sooner option was not a viable alternative, subjects did not choose to have choice, but instead preferred a direct link to the larger-later option without choosing (Hayes, Kapust, Leonard, & Rosenfarb, 1981). Using a similar concurrent chain-link experiment to the earlier pigeon work (Catania & Sogvolden, 1980), Suzuki (1999) found that monkeys (Macaca fascicularis) preferred choice, rather than choosing “not to choose,” as Hayes’s pigeons had done in the self-control task. In other words, monkeys did not prefer the terminal link that only gave them access to the preferred alternative. Instead, they chose the free-choice option more as the number of more preferred alternatives increased. Suzuki’s work involved manipulations to the probability of reinforcement, but other factors may also influence choice behavior. For example, the choice of different task types that must be completed to potentially receive reinforcement.
Washburn, Hopkins, and Rumbaugh (1991) studied exactly this situation, in which monkeys either chose or were assigned the tasks that they would perform in a test session. Washburn et al. reported that rhesus macaques (Macaca mulatta), when given a choice of which tasks to perform in a computerized test battery called the SELECT battery, established reliable and stable choice patterns. In addition, performance was significantly better on these tasks when the monkeys chose to complete the task than when the task was presented without having been chosen, suggesting a benefit of choosing. A similar study was conducted with capuchin monkeys (Cebus apella); however, in this version of the SELECT paradigm subjects had to complete all tasks, but were given control over the order in which tasks were completed (Beran, Klein, Evans, Antworth, & Chan, 2007). In that version, four icons representing tasks were presented on the screen. Monkeys selected a task by touching it with the cursor. Once the trials were completed for that task, the remaining task icons appeared on the screen and the subject could select the next task to complete. Beran et al. found significant improvements on some aspects of some tasks when monkeys were given a choice over task order as compared to a yoked or random condition. Therefore, providing more opportunity for control and choice on a computerized task appears to have at least some positive impact on performance and motivation in nonhuman primates. However, the question of whether an organism actually prefers to have choice in this type of situation has not been thoroughly investigated. So the question remains as to whether monkeys will actively choose an option that lets them choose their task order.
To address this question, we added an additional step to the SELECT task and presented it to rhesus macaques and capuchin monkeys. Prior to selection of the tasks, subjects were presented with two icons on the computer screen. One icon allowed the subject subsequently to choose the order of task completion (i.e., the free choice option), and the other icon presented the subject with the tasks in a predetermined order. We created two versions of the latter option; the tasks could be presented in a randomized order (i.e., the random option) or in an order yoked to the subject’s previous order selection (i.e., the yoked option). In the yoked condition, we isolated the act of choosing from all other aspects of the task, given that both icons yielded a preferred task order. The only difference was whether the subject would select that order in real time. In all conditions, subjects still had to move the cursor to touch the different task icons in order for the tasks to actually begin. Additionally, the probability of reinforcement was the same in all conditions, and all four tasks had to be completed regardless of whether the subject chose the order or it was selected by the computer. The tasks were well trained and relatively easy for the monkeys, so they almost always responded correctly. Therefore, unlike any previous study of choice behavior, the only difference between conditions once a monkey selected the free choice or random/yoked icon was whether the monkeys could then choose the order in which they completed the available tasks. If they preferred the icon that subsequently let them choose the task order, this would indicate that monkeys will choose to choose, despite all other factors being held equal, and that finding a way to generate choice options is a valuable outcome for monkeys.
General method
Participants
We tested six capuchin monkeys (four males, two females; ages 4–15 years) and five rhesus monkeys (all males; ages 9–19 years) housed at the Language Research Center (LRC). All monkeys had previous experience with the SELECT paradigm in which they were able to choose the order in which they performed a set of computerized tasks. Capuchin monkeys were group housed but separated for testing. Rhesus monkeys were individually housed with constant visual and auditory access to other monkeys. All monkeys were fed manufactured chow and various fruits and vegetables daily between 1600 and 1800 h. This study complied with protocols approved by the Georgia State University IACUC. All procedures were performed in full accordance with the USDA Animal Welfare Act and conformed to the “Guide for the Care and Use of Laboratory Animals.”
Materials
The monkeys were tested using the LRC’s Computerized Test System comprising a personal computer, digital joystick, color monitor, and pellet dispenser (Evans, Beran, Chan, Klein, & Menzel, 2008; Richardson, Washburn, Hopkins, Savage-Rumbaugh, & Rumbaugh, 1990). Monkeys manipulated the joystick to produce isomorphic movements of a computer-graphic cursor on the screen. Contacting appropriate computer-generated stimuli with the cursor brought them a 45-mg (capuchins) or 94-mg (macaques) banana-flavored chow pellet (Bio-Serv, Frenchtown, NJ) using a pellet dispenser interfaced to the computer through a digital I/O board (PDISO8A; Keithley Instruments, Cleveland, OH). All monkeys had previously participated in multiple psychological experiments involving this computerized test system.
Procedure
The general task was a modified version of the SELECT task that had been presented to monkeys in previous studies conducted at the LRC (e.g., Beran et al., 2007; Washburn et al., 1991). The original SELECT paradigm allowed monkeys to choose between a number of unique psychomotor and cognitive tasks that the monkeys had previous experience with using a menu format. At the beginning of a session, monkeys selected one of several task icons on the computer screen to initiate a series of test trials for a particular task. Each task icon was visually unique and represented one and only one task type. Monkeys were never directly rewarded for selecting these icons—the only contingency for doing so was the presentation of the appropriate computer task assigned to that particular icon. Once selected, each icon was removed from the menu at its next presentation, until the monkey had selected and completed trial blocks of all tasks represented in the menu. Thus, all tasks had to be completed within a block before any individual task could be completed again.
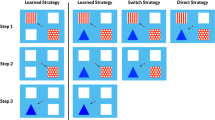
In the present study, we added an extra layer of choice to the original SELECT task. Each session began with a new menu screen in which the monkey chose between two visually unique icons on a gray background (Fig. 1a). Selection of one of these icons (hereafter referred to as the choice icon) resulted in the immediate presentation of the SELECT menu, consisting of four task icons presented on a black background (Fig. 1b). Here, the monkey could choose task icons in any order; selection of each icon resulted in its removal from the menu, until all four tasks (described below) had been completed (cf. Fig. 1b, d, f, and h).

Example test block. This illustration shows the sequence of responses involved in a block in which a monkey moves the cursor (small dot) into contact with the free choice icon (chosen in panel a), and then chooses to contact the four SELECT icons in the following order: CHASE (selected in panel b, with the CHASE task itself in panel c), learning set (LS; selected in panel d, task in panel e), delayed matching to sample (DMTS; selected in panel f, task in panel g), and matching to sample (MTS; selected in panel h, task in panel i)
The second icon represented a situation in which the task order would be automatically chosen for the monkey by the program (but the monkey still had to move its cursor into contact with the predetermined task icon—the only visible icon—before task trials would be presented). The order of tasks was determined in one of two ways, depending on the experiment: yoked or random. The selection of the yoked icon caused the program to order the tasks in a way that matched the monkey’s most recent choice order. Thus, the yoked icon presumably resulted in a preferred task order, given that subjects had just selected the same tasks in that same order. The selection of the random icon caused the program to present the tasks in a truly random order. It is critical to note that, regardless of which icon was selected, the monkey always had to complete all four trials of each task before contacting the next task icon and beginning the next task, and the monkey always had to exhaust all four tasks in a test block before those tasks would again be made available. Up to four pellets could be earned from each task (i.e., one pellet per trial). Monkeys were tested in sessions lasting from 4 to 24 h, during which they could complete as many test blocks (i.e., groupings of the four tasks) as they wanted within the allotted time (a 120-s interval separated test blocks, and during the interval the computer screen was blank). The computer program automatically recorded all responses and allowed monkeys to begin a session where they had left off on a previous day.
The four tasks included in the SELECT menu in this study were CHASE, learning set (LS), matching to sample (MTS), and delayed matching to sample (DMTS; see Washburn, Hopkins, & Rumbaugh, 1989, 1991), and each of these tasks was represented by a unique stimulus that was consistent for the entire study (Fig. 1b). These tasks were designed to be relatively easy for the monkeys, in an effort to ensure motivation and keep performance high, regardless of condition. Monkeys completed four trials of each task against a white background, and trials were separated by 1-s intertrial intervals. In CHASE, the monkey had to bring the cursor into contact with a moving circular target in order to receive a pellet (Fig. 1c). The target began in a randomly selected position and started out moving diagonally down and rightward at a speed slightly slower than the cursor. If the target came into contact with the screen edge, it appeared to deflect off the edge and then continued moving in a new direction. The angle of deflection depended on the angle of contact.
In the LS task, monkeys used the cursor to choose between two stimuli positioned in the top two corners of the screen (Fig. 1e), one of which provided a single pellet (S+) and the other of which provided a 10-s time-out penalty (S–). Monkeys had to track the S+ across the block of four trials, since the S+ and S– were randomly repositioned in the corners between trials. New LS stimuli were used for each new test block.
In the MTS task, monkeys were presented simultaneously with one sample stimulus (positioned centrally on the screen) and two comparison stimuli (randomly positioned in two of the four corners of the computer screen; Fig. 1i). Monkeys first had to move the cursor into contact with the sample stimulus, and then they could select a comparison stimulus. One comparison stimulus was identical to the sample stimulus and, when contacted, resulted in a single pellet; the other stimulus was different from the sample stimulus and resulted in a 10-s time-out penalty.
DMTS was similar to MTS, except that the sample stimulus disappeared upon contact and the comparison stimuli were presented after a delay interval that ranged from 100 ms to 12 s (randomly determined by the program on a trial-by-trial basis; Fig. 1g).
Experiment 1: Free choice versus yoked
Method
Procedure 1a—Initial test set
In this experiment, monkeys first selected between the free choice icon and the yoked icon at the beginning of a block. A block consisted of the initial choice between icons and the completion of all four tasks (see Fig. 1). After 50 blocks, new icons were introduced to represent the free choice and yoked options. For each pair of free choice and yoked icons, monkeys completed a total of 20 forced blocks in which only one icon was available (ten for each icon) and 30 test blocks involving the choice between these two icons (Table 1). The forced trials were included to ensure that the subjects had sufficient experience with each icon. Because the yoked order was based on the subject’s previous order selection of the task icons, the first block of each session had to be a forced free choice block. For the second block, the yoked icon was presented. Blocks 3–10 consisted of four forced free choice and four forced yoked blocks, presented in a random order. Blocks 11–50 began with a string of five test blocks in which subjects could select either the free choice or the yoked icon. Then, to ensure continued exposure to the outcome of each icon, test blocks alternated between two forced blocks (one of each type presented in random order) and five test blocks, and the set ended with a string of five test blocks. Monkeys completed ten such sets, each involving a unique pair of free choice and yoked icons. This process was repeated ten times (ten sets of 50 blocks per icon pair). We compared performance on these ten sets to random selection of the free choice icon (15 out of 30) using a one-sample t test for each individual.
Procedure 1b—Extended test set
To examine the possibility that 50 blocks did not provide sufficient experience for subjects to learn the consequences of selecting the free choice and yoked icons, we tested a subset of individuals on a phase in which the same icons were used for 500 blocks. This experiment examined the possibility that monkeys might show stronger biases for one icon or the other if they had more exposure to a particular pair of icons. The 500 blocks were completed over multiple test days with the same pair of free choice and yoked icons. The first 50 blocks of this experiment proceeded exactly as they had in Experiment 1a (in terms of the mixture of forced and test blocks). However, Blocks 51–500 consisted of a greater proportion of test blocks, with monkeys completing only one pair of forced blocks at the beginning of each new test day. We hypothesized that monkeys would show a greater bias for the free choice icon here than in Experiment 1a. We compared the number of free choice selections to chance expectation using the cumulative probability that the number of responses would be equal to or greater than chance using a binomial distribution.
Results
In Experiment 1a, no individual selected the free choice icon significantly more often than the yoked icon across the ten sets (all ts < 2, p > .05; see Table 2). On average, capuchin monkeys selected the free choice icon on 53.5 % of trials, and rhesus monkeys selected it on 51.9 % of trials (see Table 2 for the individual breakdown). Thus, subjects did not show a significant preference for either the free choice or yoked icons in this experiment.
In Experiment 1b, no individual selected the free choice icon significantly more than would be expected by chance (cumulative binomial probability p > .05 for all monkeys; see Table 2). Five animals completed this follow-up task, and because of program errors, performed between 431 and 585 trials with the same icons (see Table 2 for the individual breakdown). On average, the two rhesus macaques selected the free choice icon on 47.2% of trials, and the three capuchin monkeys selected it on 52.4 % of trials. Thus, subjects did not show a significant preference for either the free choice or yoked icons in this experiment, and this pattern did not change across blocks.
Discussion
Performance on this task suggested that subjects did not have a preference for choosing the task order or being automatically given a preferred task order (that matched their most recent selection). However, because we observed no explicit difference in the outcomes for selecting the free choice or yoked icon, subjects may have simply viewed these icons as “start” buttons that had no true function and simply began the SELECT task. The fact that subjects did not develop a preference for either of the options supports this possibility. Thus, we implemented a second experiment that made the outcome of the initial choice between icons more salient. In the second experiment, one icon again provided choice, whereas the second icon resulted in an automatically selected random order of tasks, rather than a necessarily preferred order, as had been true in the yoked condition of Experiment 1. This would be more noticeable to the monkeys in terms of the order in which they did the tasks that actually led to food reward, and therefore might evoke some preferences from them.
Experiment 2: Free choice versus random
Method
Procedure 2a—Initial test set
This experiment was identical to Experiment 1a (free choice vs. yoked; 50 blocks per stimulus pair), except that the yoked icon was replaced by the random icon. Monkeys completed ten sets, each involving a unique pair of free choice and random icons (Table 1). When the random icon was selected, the program used a random number generator to select tasks, without replacement, in a random order. We hypothesized that monkeys would show a greater bias for the free choice icon over the random icon than they had for the free choice icon over the yoked icon. We expected this because the yoked condition allowed monkeys to complete the four computer tasks in their already established and preferred order (even though the monkeys did not select them in real time), whereas the random condition did not always yield a preferred order. This should have been a more salient difference to the monkeys when it came time to actually complete the individual tasks, and thus should have placed a greater premium on selecting the free choice icon, so that monkeys could still do the individual tasks in the order that they preferred. We compared performance on these ten sets to random selection of the free choice icon (15 out of 30) using a one-sample t test for each individual.
Procedure 2b—Extended test set
This experiment was identical to Experiment 1b (free choice vs. yoked; 500 blocks) except that the yoked icon was replaced by the random icon. We hypothesized that monkeys would show a greater bias for the free choice icon over the random icon than they had in Experiment 2a (free choice vs. random; 50 blocks per stimulus pair) because they would have more time to learn the contingencies of a particular pair of free choice and random icons. All monkeys were tested in this experiment. We compared the number of free choice selections to chance expectation using the cumulative probability that the number of responses would be equal to or greater than chance using a binomial distribution.
Results
In Experiment 2a, no individual selected the free choice icon significantly more than the random icon across the ten sets (all ts < 2.2, ps > .05; see Table 3). On average, capuchin monkeys selected the free choice icon on 54.53 % of trials, and rhesus monkeys selected it on 52.5 % of trials (see Table 3 for the individual breakdown). Thus, subjects did not show a significant preference for either the free choice or yoked icons in this experiment, and this pattern did not change across blocks.
In Experiment 2b, approximately half of the subjects showed a significant preference for free choice over random when given 500 blocks of the same pair of icons (two macaques and four capuchin monkeys, p < .05; see Table 3). For only the individuals showing a significant preference, the rhesus macaques selected the free choice icon on 65.66 % of trials, and the capuchin monkeys selected it on 68 % of trials.
Discussion
Six monkeys showed a significant preference for choice in Experiment 2b. For this effect to emerge, the animals had to have considerable experience with the icons themselves and had to learn that the other possible outcome involved random presentation of the task order, rather than the monkeys’ own preferred order, as had been the case in Experiment 1. Given these conditions, preference for choice emerged in more than half of the animals. However, we were interested in whether this finding would generalize to new icons, provided that a sufficient number of trials were completed to permit the requisite new learning.
Experiment 3: Generalization of preference for free choice versus random
This experiment was included to determine whether the significant preferences for the free choice icon in Experiment 2b would generalize to new sets of stimuli in the first part of the block. We presented two additional 500-block sets using novel free choice and random icons (Table 1). Furthermore, we were interested in the overall pattern of preference, and whether a preference for free choice would develop more rapidly with the introduction of each set of new icons (i.e., across icon pairs). For the purpose of the analyses, we included the 500-block set from Experiment 2b in addition to the two new sets collected as part of Experiment 3. Each 500-block set included approximately 480 actual test blocks, and these were divided into twelve 40-block intervals. We compared the overall number of free choice selections to chance expectation using the cumulative probability that the number of responses would be equal to or greater than chance using a binomial distribution.
Results
Five out of six capuchin monkeys showed a significant preference for free choice for the first and second sets of icons, and all of the capuchin monkeys showed a significant preference in the third set (p < .05; see Table 4). Two rhesus monkeys showed a significant preference for free choice in the first set; three showed a preference in the second set of icons; and all five showed a significant preference in the third set (p < .05; see Table 4). In no case did any monkey show a preference for the random icon.
The data then were divided into twelve 40-block intervals and averaged across all individuals. A within-subjects ANOVA was conducted with Set and Block as factors. For capuchin monkeys, we observed a significant main effect of set, F(2, 110) = 6.03, p = .019, a significant main effect of block, F(11, 110) = 10.48, p < .001, and no significant interaction, F(22, 110) = 0.512, p = .964 (Fig. 2). For rhesus macaques, a significant main effect of set emerged, F(2, 110) = 5.47, p = .032, but we found no significant main effect of block, F(11, 110) = 1.47, p = .177, and no significant interaction, F(22, 110) = 0.747, p = .778 (Fig. 3).

Capuchin monkey performance across 40-block intervals for the 1st, 2nd, and 3rd sets of free choice and random icons (± SE)

Rhesus monkey performance across 40-block intervals for the 1st, 2nd, and 3rd sets of free choice and random icons (± SE)
Discussion
The results indicated that all subjects showed a significant preference for choice when the alternative was a randomly determined task order. This suggests that monkeys do prefer choice after sufficient exposure, at least in some contexts, even though all other components of the task were the same regardless of whether subjects selected the free choice or random icon. Specifically, the same number of trials had to be completed, the same tasks had to be completed, the same physical movements had to be completed (i.e., touching the task icons with the cursor), and most importantly, performance was highly accurate, and thus the global rate of reinforcement was functionally equivalent. This suggests that something above and beyond the task contingencies drove the observed preference, and it appeared to be a preference to choose a preferred task order. For both species, we observed a significant effect of set, indicating that the preference for choice increased across sets. The main effect of block was only significant for capuchin monkeys. Within a set, the capuchin monkeys’ preference for choice increased significantly across blocks. This was not the case for rhesus monkeys, but as can be seen in Fig. 3, this preference emerged in the very first interval. This finding suggests that subjects were not simply accruing experience with the response contingencies, but may have been seeking out and preferentially selecting the choice icon from the beginning of the set. Given that a large number of trials was necessary for this pattern to emerge and stabilize, we were interested in whether this preference would generalize to the yoked condition if sufficient experience was provided.
Experiment 4: Generalization of preference for free choice versus yoked
Here, we repeated Experiment 1b (free choice vs. yoked, 500 blocks) in order to assess whether any of the differences in performance seen between Experiments 1b and 3 were due to differences in experience with the overall task (see Table 1). We hypothesized that monkeys’ preference for the free choice icon would return to the levels exhibited in Experiment 1b and that their greater preference for free choice over random would not be explainable by experience. We also repeated this procedure for two additional sets of icons (except in the case of one capuchin monkey that was not available for testing). We compared the number of free choice selections to chance expectation using the cumulative probability that the number of responses would be equal to or greater than chance using a binomial distribution. It is important to note that, although we found some differences in preference for task order completion across monkeys, the preferred order of task completion within a monkey remained highly consistent in the free choice condition. Thus, we can assume that the order in the yoked condition was a preferred order for each subject.
Results
For the first 500-block set of icons, only one out of 11 monkeys showed a significant overall preference for the free choice over the yoked icon (p < .05; see Table 5). For the second set, two out of five rhesus monkeys showed a significant preference for free choice (p < .05; Table 5). For the third set, four out of five rhesus monkeys showed a significant preference for free choice. Only two of five capuchin monkeys showed a significant preference for free choice, and they did so only once each, with the second set of stimuli. In no case did any monkey show a preference for the yoked icon over the free choice icon.
Discussion
The results of Experiment 4 indicate that with sufficient exposure, some individuals maintain a preference for choice even when the alternative gives a preferred task order that had been previously selected by the individual. However, this pattern only emerged for some of the individuals, and it was not as robust as when the alternative to choice produced a random task order. In Experiment 4, we provided the monkeys with the same number of test blocks (500) to learn each new set of stimuli. This was sufficient for a universal preference for free choice to emerge by the third set of Experiment 3, and so we assume that this number of test blocks provided sufficient experience for the monkeys to learn the icons. Thus, although the preference was weaker in Experiment 4, some monkeys continued to prefer the act of choosing, and no monkeys reliably preferred “no choice,” even when every aspect of “choice” versus “no choice” was matched. In other words, this preference existed even when monkeys were completing tasks in the same order, reinforcement rates were matched and spread evenly across sessions, and the task demands completely overlapped, with the exception of whether or not the monkeys chose the order or a preferred order was determined for them.
General discussion
In animal research, it is hard to disentangle a preference for choice from various aspects of the task that co-vary with the option of having more choice (e.g., greater overall quantity, more variability, perceptual distinctiveness). Here we took advantage of an existing computerized task paradigm to allow capuchin and rhesus monkeys to choose whether they selected the order in which tasks were completed or the order was assigned by the computer. In both cases, subjects still had to complete the same four tasks to earn food pellets, and thus task demands and overall reinforcement rates were controlled across the choice and no choice conditions. In the no-choice conditions, subjects were either randomly assigned a task order (random) or it was yoked to the previous order selection (yoked), thus yielding a preferred order every time.
In the free choice versus random condition, subjects showed a significant and reliable preference for choice when given sufficient experience with the two icons. In this comparison, even though overall reinforcement rates were the same, it is possible that reinforcement was frontloaded within a test block given that preferred tasks (ostensibly faster to complete) occurred earlier in a block. Thus, reinforcement distribution could in part account for the preference for the free choice icon. Yet performance with the third set of icons provides support against a molecular, learning-based account of performance. The preference emerged more rapidly and reached higher levels across sets, producing an apparent “learning set” (Harlow, 1949) for the concept of choice. As is shown in Figs. 2 and 3, the preference for choice emerged within the very first interval of the third set, before subjects could have had much experience with the subtle, within-block shifts in the reinforcement distribution that might underlie a purely learning-based preference for choice. However, it remains a possibility that this factor might have influenced performance. In contrast, for the free choice versus yoked condition, the same task order almost always occurred for subjects, thus reinforcement was equated at a molar and molecular level. In this condition, some subjects continued to prefer choice, although to a lesser extent than in the random condition. This outcome is especially relevant because we have isolated the act of choosing from all other aspects of the task, and found that a preference for choice appears in nonhuman primates.
It should be noted that although we observed individual differences in the effects reported here, there were numerous reasons to predict that the animals would show no preference whatsoever for the free choice conditions. In our test system, the monkeys always have control over when or whether they work on tasks. Although they receive fruit-flavored chow pellets for completing computer-task trials, they are not deprived of food or reduced in body weight for purposes of testing, and they are fully provisioned each day whether or not they complete the tasks. Within a test session, the animals can choose not to work at all, with no penalty other than the delay in receiving rewards. We have argued that the animals’ psychological well-being and performance benefit from this level of choice, and in the present study (as in Beran et al., 2007; Washburn et al., 1991) any advantage or preference for choice of task order was additive to this other aspect of freedom to choose. This finding has important implications for animal care, and for our understanding of the human desire to choose.
In particular, the present data suggest that animals are sensitive to the benefit of choice and will seek out opportunities for choice when all other aspects of the task are equivalent. One interpretation is that animals “value” choice—either through learned mechanisms or an evolved inherent preference for choice (Voss & Homzie, 1970). Thus, opportunities for increased control and choice should be implemented whenever possible with nonhuman primates. Additionally, increased choice may also directly or indirectly impact performance related and behavioral variables (Bloomsmith, Keeling, & Lambeth, 1990; Bloomsmith & Lambeth, 2000; Bloomsmith, Baker, Lambeth, Ross, & Schapiro, 2000; Hanson, Larson, & Snowdon, 1976; Mineka, & Hendersen, 1985; Washburn et al., 1991). Interestingly, a body of research also suggests that not necessarily control, but the perception of control, is most beneficial to an individual (e.g., Perlmuter & Monty, 1977). Therefore, it may not be necessary to implement changes that facilitate actual choices, but the more relevant goal may be to create environmental conditions in which the belief or perception of choice or control prevails. In other words, even if an individual is not actually controlling a given situation, the belief or perception that one is in control is important (Perlmuter & Monty, 1977).
These results are also relevant to our understanding of human choice. It has been suggested that humans have a biological imperative to prefer choice from an early age (Leotti et al., 2010), and the present work adds to this body of literature. However, the relationship between a preference for choice and its actual benefit is not straightforward in humans. For example, Iyengar and Lepper (2000) found that having fewer options might actually increase motivation and satisfaction with the choice, as compared to having more options. The researchers set up a “jam tasting” table with either six or 24 samples to choose from. More people approached the table when more choices were available; however, they found that participants who were given fewer options were more likely to actually purchase jam. In a subsequent experiment, subjects with fewer options of chocolates reported greater satisfaction and less regret in their decision than did those with more options (Iyengar & Lepper, 2000). These results are consistent with the growing idea that whereas some opportunity for choice over no choice may be beneficial, there can be too much of a good thing, leading to the “tyranny of choice” (Schwartz, 2004) or the “paradox of choice” (Reed, Kaplan, & Brewer, 2012). The present paradigm is well-suited to investigate the boundaries of the preference for choice in nonhumans and to identify similarities and differences to the paradox of choice in humans.
In the wild, a primate would likely be faced with two, three, or maybe four choices between food sources, mates, sleeping sites, and so forth. Rarely can a scenario in the wild be imagined in which an individual would have to make the kinds of choices facing modern humans, for whom the magnitude of available options seems to be growing at an exponential rate, ranging from choosing a college, which car to drive, or which of the many varieties of sweeteners to purchase, and the Internet is quickly magnifying the possibilities. The paradox of choice appears to grow from conflict between a biological imperative to prefer choice and the cultural drive to provide and have access to more choices rather than fewer. An evolutionary perspective can inform this debate and provide a better understanding of choice. Our research supports the idea that—despite the issues that arise in modern society from the seemingly never-ending options available to us—choice, at its core, is indeed beneficial and is preferred by primates.
References
Beran, M. J., Klein, E. D., Evans, T. A., Antworth, R., & Chan, B. (2007). Perceived control, motivation, and task performance in capuchin monkeys. In P. R. Zelick (Ed.), Issues in the psychology of motivation (pp. 171–185). Hauppauge: Nova Science.
Bloomsmith, M. A., Keeling, M. E., & Lambeth, S. P. (1990). Videotapes: Environmental enrichment for singly housed chimpanzees. Lab Animal, 19, 42–46.
Bloomsmith, M. A., & Lambeth, S. P. (2000). Videotapes as enrichment for captive chimpanzees (Pan troglodytes). Zoo Biology, 19, 541–551.
Bloomsmith, M. A., Baker, K. C., Lambeth, S. P., Ross, S. K., & Schapiro, S. J. (2000). Is giving chimpanzees control over environmental enrichment a good idea? In The apes: Challenges for the 21st century (pp. 88–89). Brookfield: Chicago Zoological Society.
Catania, A. C. (1975). Freedom and knowledge: An experimental analysis of preference in pigeons. Journal of the Experimental Analysis of Behavior, 24, 89–106.
Catania, A. C., & Sagvolden, T. (1980). Preference for free choice over forced choice in pigeons. Journal of the Experimental Analysis of Behavior, 34, 77–86.
Evans, T. A., Beran, M. J., Chan, B., Klein, E. D., & Menzel, C. R. (2008). An efficient computerized testing method for the capuchin monkey (Cebus apella): Adaptation of the LRC-CTS to a socially housed nonhuman primate species. Behavior Research Methods, Instruments, & Computers, 40, 590–596.
Greenberger, D. B., Strasser, S., Cummings, L. L., & Dunham, R. B. (1989). The impact of personal control on performance and satisfaction. Organizational Behavior and Human Decision Processes, 43, 29–51.
Hanson, J. D., Larson, M. E., & Snowdon, C. T. (1976). The effects of control over high intensity noise on plasma cortisol levels in rhesus monkeys. Behavioral Biology, 16, 333–340.
Harlow, H. F. (1949). The formation of learning sets. Psychological Review, 56, 51–65.
Hayes, S. C., Kapust, J., Leonard, S. R., & Rosenfarb, I. (1981). Escape from freedom: Choosing not to choose in pigeons. Journal of the Experimental Analysis of Behavior, 36, 1–7.
Iyengar, S. S., & Lepper, M. R. (2000). When choice is demotivating: Can one desire too much of a good thing? Journal of Personality and Social Psychology, 79, 995–1006.
Leotti, L. A., Iyengar, S. S., & Ochsner, K. N. (2010). Born to choose: The origins and value of the need for control. Trends in Cognitive Science, 14, 457–463.
Mineka, S., & Hendersen, R. W. (1985). Controllability and predictability in acquired motivation. Annual Review of Psychology, 36, 495–529. doi:10.1146/annurev.ps.36.020185.002431
Overmier, J. B., & Seligman, M. E. (1967). Effects of inescapable shock upon subsequent escape and avoidance responding. Journal of Comparative and Physiological Psychology, 63, 28–33.
Perlmuter, L. C., & Monty, L. A. (1977). The importance of perceived control: Fact or fantasy? Experiments with both humans and animals indicate that the mere illusion of control significantly improves performance in a variety of situations. American Scientist, 65, 759–765.
Reed, D. D., Kaplan, B. A., & Brewer, A. T. (2012). Discounting the freedom to choose: Implications for the paradox of choice. Behavioural Processes, 90, 424–427.
Richardson, W. K., Washburn, D. A., Hopkins, W. D., Savage-Rumbaugh, S. E., & Rumbaugh, D. M. (1990). The NASA/LRC computerized test system. Behavior Research Methods, Instruments, & Computers, 22, 127–131.
Schwartz, B. (2004). The tyranny of choice. Scientific American, 290, 70–76.
Suzuki, S. (1999). Choice between forced- and free-choice by monkeys (Macaca fascicularis). Perceptual and Motor Skills, 88, 242–250.
Voss, S. C., & Homzie, M. J. (1970). choice as a value. Psychological Reports, 26, 912–914.
Washburn, D. A., Hopkins, W. D., & Rumbaugh, D. M. (1989). Automation of learning-set testing: The video-task paradigm. Behavior Research Methods, Instruments, & Computers, 21, 281–284.
Washburn, D. A., Hopkins, W. D., & Rumbaugh, D. M. (1991). Perceived control in rhesus monkeys (Macaca mulatta): Enhanced video-task performance. Journal of Experimental Psychology: Animal Behavior Processes, 17, 123–129.
Winocur, G., Moscovitch, M., & Freedman, J. (1987). An investigation of cognitive function in relation to psychosocial variables in institutionalized old people. Canadian Journal of Psychology, 41, 257–269.
Zuckerman, M., Porac, J., Lathin, D., Smith, R., & Deci, E. L. (1978). On the importance of self-determination for intrinsically motivated behavior. Personality and Social Psychology Bulletin, 4, 443–446.
Author note
This work was supported in part by the Duane M. Rumbaugh Fellowship, by funding from NICHD (Grant No. HD060563), and by Georgia State University. We greatly appreciate the efforts of the staff at the Language Research Center of Georgia State University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Perdue, B.M., Evans, T.A., Washburn, D.A. et al. Do monkeys choose to choose?. Learn Behav 42, 164–175 (2014). https://doi.org/10.3758/s13420-014-0135-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-014-0135-0