
Abstract
Avoidance of a target flavor can be produced by providing rats with a highly nutritious solution of 20 % maltodextrin (20 %Malto) in some sessions and a 3 % maltodextrin (3 %Malto) solution containing the target flavor in intermixed sessions. Since 20 %Malto is both more nutritious and more palatable than 3 %Malto, flavor avoidance could arise because the flavor signals either a reduction in calories or reduced palatability, or both. Pilot testing established that rats strongly preferred 3 %Malto plus 0.1 % saccharin to both unflavored 3 %Malto and unflavored 20 %Malto. The two main experiments tested whether the palatability difference, which the pilot data had suggested was larger than the difference between 20 %Malto and 3 %Malto, could produce flavor avoidance. In both experiments, one group of rats were given 3 %Malto plus 0.1 % saccharin on some days, intermixed with other days on which this group was given 3 %Malto plus the target flavor, almond. Neither when trained and tested under conditions of food deprivation (Experiment 1) nor when trained and tested sated (Experiment 2) did palatability reduction produce almond avoidance. In contrast, calorie reduction produced almond avoidance under both conditions. These results suggest that flavor avoidance can be produced by intermixed training involving solutions that differ in nutritious value and palatability, but not when they differ only in palatability.
Similar content being viewed by others
A dilute solution of the nonnutritious chemical saccharin is highly palatable to rats, as it is to humans. After rats have on several occasions drunk a saccharin solution to which has been added some relatively novel flavor, such as the almond used in the present experiments, they show increased preference for the flavor (e.g., Fanselow & Birk, 1982; Harris, Gorissen, Bailey, & Westbrook, 2000; Holman, 1975). Such results indicate that flavor preference learning can be based on flavor–flavor associations alone. Another method for producing an increase in preference for a flavor is to pair its ingestion with intragastric infusion of a nutrient, such as the hydrolyzed starch maltodextrin, also used in the present experiments (e.g., Sclafani, 1991; Sclafani & Nissenbaum, 1988; Yiin, Ackroff, & Sclafani, 2005). Since in these experiments the animals cannot taste the nutrient, flavor preference learning in this case must be based solely on associating the flavor with nutritional consequences—that is, on the acquisition of what are termed flavor–calorie or flavor–nutrient associations.
The experiments reported here were prompted by the question of what is learned when rats develop avoidance of—rather than preference for—a flavor. Previously we had found that, when hungry rats had been given training that intermixed “good” and “bad” sessions, in subsequent two-bottle choice tests they showed avoidance of the flavor that had been present on the bad sessions. The good sessions in those experiments were ones in which the rats were given a concentrated maltodextrin solution (either 16 % or 20 %), while in the bad sessions they were given a dilute maltodextrin solution (either 2 % or 3 %) to which either almond essence or sodium chloride had been added (Boakes, Colagiuri, & Mahon, 2010). We provisionally applied the label missing-calorie effect to the flavor avoidance learning produced by such training. This label was based on the assumption that these hungry rats avoided the target flavor because they had learned that this flavor signaled the absence of calories that would otherwise be available in the experimental context.
An alternative possibility is suggested by the fact that rats find concentrated maltodextrin to be more palatable than dilute maltodextrin. Although humans find the taste of this complex mixture of polysaccharides to be increasingly unpleasant as concentration is increased, rats and other rodents find the taste of maltodextrin to be very palatable (Feigin, Sclafani, & Sunday, 1987). Nondeprived rats were found to drink more maltodextrin (Polycose) than sucrose solutions in both 30 min single-bottle and two-bottle tests over a concentration range of 1 % to 4 % (Sclafani & Clyne, 1987). Furthermore, rates of licking for low concentrations over the first 3 min of these tests were higher for maltodextrin than for sucrose, and they increased monotonically with concentration over the range 1 %–32 %, a finding that supports the assumption that high intakes of maltodextrin reflect its greater palatability.
Initial lick rate has been one of several measures obtained in microstructural analyses of licking patterns. Davis (1996) applied a full analysis of this kind to maltodextrin solutions varying in concentration from 0.6 % to 40 %, again using nondeprived rats. His study confirmed that initial lick rates increased monotonically over this range, and he also found a similarly monotonic function for the mean size of lick clusters (bouts), another measure of palatability. Subsequently, Dwyer (2008, Exp. 1) extended to food-deprived rats such a microstructural analysis of licking for maltodextrin ranging in concentration from 2 % to 16 % and found a steep increase in cluster size as concentration increased beyond 4 %. In a second experiment, cluster size for a flavor that had been added to 16 % maltodextrin during an initial training phase was higher than for a second flavor that had been added to 2 % maltodextrin during the training phase, thus indicating that such training can increase for food-deprived rats the palatability of a previously near-neutral flavor (Dwyer, 2008, Exp. 2).
Returning to the question of what is learned in the intermixed training procedure described above, research showing that for rats the palatability of maltodextrin increases with concentration indicates that on “bad” days the solution has a less pleasant taste, as well as containing fewer calories. Thus, learned avoidance of the flavor present on bad days might be a “disappointing taste” effect instead of, or as well as, a “missing-calorie” effect.
The aim of the present experiments was to test whether training involving two solutions differing in palatability, but with the same nutritional value, would produce avoidance of the target flavor. In both experiments, we used a procedure in which a highly palatable solution of low nutritional value—that is, 3 % maltodextrin (3 %Malto) plus saccharin—was given on good days, and a less palatable solution containing the same calories—that is, 3 %Malto to which the target flavor almond had been added—was given on bad days.
Sclafani, Einberg, and Nissenbaum (1987) reported that adding 0.125 % saccharin to either a 3 % or a 32 % Polycose solution made these solutions more palatable to nondeprived rats, in terms of preference in two-bottle tests that included the equivalent plain Polycose solutions. Polycose is a particular type of corn-derived maltodextrin produced in the United States that is very similar to the Australian-produced maltodextrin used in the present series of experiments. In an unpublished experiment, we failed to detect any difference between Polycose and the maltodextrin used here in flavor preference learning based on the two carbohydrates. Since we wanted the palatability difference in the present experiments to be at least as great as between 20 % maltodextrin (20 %Malto) and 3 %Malto, we undertook two substantial pilot experiments that examined preferences when rats were given a choice between strong but unflavored maltodextrin solutions and weak maltodextrin solutions to which various concentrations of saccharin had been added.
The aim of the first pilot experiment was to determine what concentration of saccharin we needed to add to a low concentration of maltodextrin in order for rats to be indifferent between this combination and unflavored 20 %Malto in a two-bottle choice test. A total of 15 rats were first exposed to both high concentrations of unflavored maltodextrin and low-maltodextrin solutions with progressively less saccharin added. Subsequent tests established that, when 0.025 % saccharin was added to 3 %Malto, rats whose weights had been reduced to 80 % of their ad-libitum weights by restricting food access showed a preference (69 %) for 20 %Malto over the saccharin-flavored solution, whereas rats that were not food-deprived were indifferent between the two solutions (48 %).
In a further two-bottle test, the same rats were given a choice between unflavored 3 %Malto and the same solution to which 0.025 % saccharin had been added. This confirmed a strong preference for the saccharin-added solution that was greater in rats that were food-deprived (83 %) than in nondeprived rats (70 %). In summary, the main results of this first pilot experiment were, first, that the normal strong preference that rats show for 20 %Malto over 3 %Malto can be reduced considerably by adding 0.025 % saccharin to the weaker solution and, second, that food deprivation increases preference for 20 %Malto and—to a lesser degree—for 0.025 % saccharin.
The aim of the second pilot experiment was to measure preferences when 0.1 % saccharin (i.e., four times stronger than the solution in the first pilot experiment) was added to 3 %Malto. This combination was used as the high-palatability solution for the saccharin groups in the present Experiments 1 and 2. Also, the level of food deprivation was identical to the one that would be used in Experiment 1. A group of 47 rats had previously served in an experiment in which they had been given 20 %Malto, as well as lower concentrations of maltodextrin and almond flavor, but had no exposure to saccharin. Access to food was restricted so as to maintain them at 85 % of their adjusted ad-lib body weight both during that experiment and throughout the pilot testing described next. First, in order to reduce any neophobia to saccharin, the rats were initially given 3 %Malto plus 0.1 % saccharin for several 10 min sessions. These sessions were followed by a series of 10 min two-bottle tests. The first provided a choice between unflavored 3 %Malto and 3 %Malto plus 0.1 % saccharin. The second provided a choice between unflavored 20 %Malto and 3 %Malto plus 0.1 % saccharin.
Since food-deprived rats in the first pilot experiment had strongly preferred 3 %Malto plus 0.025 % saccharin to unflavored 3 %Malto, it was predicted that the first test in this second pilot experiment would reveal a preference that was at least as strong for the 0.1 % saccharin solution. This was confirmed by the finding of an 82 % preference for 3 %Malto plus 0.1 % saccharin over unflavored 3 %Malto. Even though in the first pilot experiment food-deprived rats had shown a slight preference for 20 %Malto over 3 %Malto plus 0.025 % saccharin, it was predicted that increasing the saccharin concentration to 0.1 % would reverse this preference. This prediction was confirmed, in that the final test revealed a preference (81 %) for the 3 %Malto plus 0.1 % saccharin over 20 %Malto.
Experiment 1
As was discussed in the introduction, avoidance of almond produced by intermixed training in which rats were given 20 %Malto in some sessions and 3 %Malto plus almond in intermixed sessions (Boakes et al. 2010) could have resulted at least partly from the palatability difference between the two solutions. In the present experiment, we tested whether almond avoidance would be produced by training in which rats received almond mixed with 3 %Malto in some sessions but in intermixed sessions received a far more palatable solution, 3 %Malto plus 0.1 % saccharin, that did not contain almond. Since the pilot data had indicated that rats prefer 3 %Malto plus 0.1 % saccharin to 20 %Malto, it seemed likely that the palatability difference between 3 %Malto plus 0.1 % saccharin and 3 %Malto plus almond would be greater than that between 20 %Malto and 3 %Malto plus almond.
To confirm that the conditions were ones that could produce almond avoidance on the basis of calorie reduction, the group given training using 0.1 % saccharin was compared with one given calorie-based intermixed training in which the more preferred solution was 20 %Malto. A further two groups were given a variant of the blocked control procedure used in Boakes et al. (2010), to provide equivalent exposure to both the good and bad solutions without intermixing them. In the present experiment, the blocked condition was one in which rats were given a block of bad sessions followed by a block of good sessions. This control had proved satisfactory in a previous experiment involving flavor preference learning by fluid-deprived rats, in which the group given this condition had shown an almond preference in a subsequent two-bottle test of almond versus water of about 40 %, a similar value to that shown by a group given simple exposure to almond (Albertella & Boakes, 2006, Exp. 1).
The present experiment used a 2 × 2 factorial design, with Training (intermixed vs. blocked) and Nutrient (saccharin vs. maltodextrin) as factors. The resulting four groups were labeled Intermixed-Sacch, Blocked-Sacch, Intermixed-Malto, and Blocked-Malto. Some minor procedural changes were made to the method used by Boakes et al. (2010) that had produced avoidance of the target flavor following intermixed training. These changes included giving two days of habituation to the drinking chambers before training started, giving only six sessions of training, and giving rats in the blocked condition their bad sessions prior to their good sessions, whereas the reverse sequence had been used for the blocked condition in Boakes et al. (2010).
Method
Subjects
A group of 32 experimentally naïve hooded Wistar rats from the University of Adelaide were approximately 60 days old at the start of the experiment, with a mean weight of 321 g, range 269 – 400 g. They were housed communally, with eight rats to a large cage, and were maintained on a feeding schedule whereby they were given sufficient food following a session or on rest days to maintain them at 85 % of their initial ad-lib bodyweights. Water was always available in the home cages, except for 2 h before a session.
Rats were allocated to the four groups (each n = 8) such that the first and fourth squads each contained four rats from Group Intermixed-Sacch and four from Group Intermixed-Malto, while the second and third squads contained four rats from Group Blocked-Sacch and four from Group Blocked-Malto. Allocation to drinking chambers was also counterbalanced.
Apparatus and solutions
Eight acrylic drinking cages, measuring 33 × 21 × 19 cm, were covered with wire lids; into each of these cages two 100-ml plastic bottles, with stainless steel spouts containing two ball bearings, were inserted in a central position, with the tips of the spouts approximately 7 cm apart. The floors were covered with pellets of paper. The maltodextrin consisted of hydrolyzed corn starch containing 1.2 % monosaccharides, 5.4 % disaccharides, and the remainder higher polysaccharides (Myopure, Australia; www.myopure.com.au). The solutions in tap water were 20 %Malto, 3 %Malto plus 0.1 % saccharin, and 3 %Malto plus 1 % almond essence (Queens Fine Foods Pty. Ltd., Queensland) in the training phase and 6 %Malto and 6 %Malto plus 1 % almond in the test phase.
Procedure
Experimental sessions were normally conducted seven days per week. For two days prior to the start of training, the rats were placed in the drinking chambers for 10 min without access to any fluid or any other kind of treatment. During training, only one bottle of solution was given to a rat, and fluid access was restricted to approximately 12 ml. Because in previous experiments of this kind some rats drank very little if the initial sessions were of short duration, following what has since become our standard practice, the first two sessions were longer than the rest. Thus, session duration here started at 20 min for Session 1, 15 min for Session 2, and 10 min for every session thereafter.
All of the rats in the intermixed condition were given 3 %Malto plus almond on “bad” days (Sessions 3, 4, and 6). On “good” days (Sessions 1, 2, and 5), Group Intermixed-Sacch was given 3 %Malto plus saccharin and Group Intermixed-Malto was given 20 %Malto. Rats in both blocked conditions were first given 3 %Malto plus almond on three consecutive bad days (Sessions 1, 2, and 3). On the three good days that followed (Sessions 4, 5, and 6), Group Blocked-Sacch was given 3 %Malto plus saccharin and Group Blocked-Malto was given 20 %Malto.
Following the six training sessions, all rats were treated identically. In the two sessions between training and test (Sessions 7 and 8), they were given 6 %Malto. This solution was presented in two bottles, roughly 7 cm apart, to accustom rats to the two-bottle procedure to be used during the tests and to check for strong position habits (> 80 %). The groups were then given two-bottle tests for almond avoidance: In Session 9, the almond solution, 6 %Malto plus almond, was on the left, and unflavored 6 %Malto was on the right. The positions of these solutions were reversed on Session 10. The reason for adding the almond to the maltodextrin solution in this test was because previous experience had shown that, when food-deprived, our rats drink very little in a test in which the choice is between almond-in-water and water.
Results and discussion
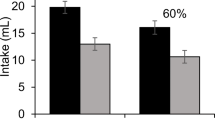
The intakes of the various training solutions are shown in Fig. 1. It may be seen in the top panel that Groups Intermixed-Sacch and Intermixed-Malto drank essentially identical amounts of both the high-palatability solutions (3 %Malto plus saccharin or 20 %Malto, on Days 1, 2, and 5) and the low-palatability solution (3 %Malto plus almond, on Days 3, 4, and 6). As can seen in the lower panel, the two blocked groups also did not differ in consumption of the training solutions, with initial low intakes of 3 %Malto plus almond (Days 1, 2, and 3) followed by high intakes of the highly palatable solutions on Days 4, 5, and 6.

Experiment 1: Fluid intakes during training. The upper panel shows intakes by the two groups given intermixed training, in which they received 3 % maltodextrin plus almond in Sessions 3, 4, and 6 and either 3 % maltodextrin plus 0.1 % saccharin (Group Intermixed-Sacch) or 20 % maltodextrin (Group Intermixed-Malto) in Sessions 1, 2, and 5. The lower panel shows intakes by the two groups given blocked training, who were given 3 % maltodextrin plus almond in Sessions 1–3 and either 3 % maltodextrin plus 0.1 % saccharin (Group Blocked-Sacch) or 20 % maltodextrin (Group Blocked-Malto) in Sessions 4–6
A repeated-measures 2 × 2 ANOVA applied to consumption of 3 %Malto plus almond during training, with Training (intermixed vs. blocked) and Nutrient (saccharin vs. maltodextrin) as factors, revealed an effect of training, F(1, 28) = 15.7, p < .001, with more 3 %Malto plus almond being consumed by rats in the intermixed groups (mean = 7.5 ml) than by rats in the blocked groups (mean = 3.3 ml) but, importantly, no effect of nutrient, F < 1, and no interaction between training and nutrient, F(1, 28) = 1.25, p > .05. There were two probable reasons for the training effect, whereby the blocked group drank less than the intermixed group. One possibility was neophobia, since the 3 %Malto plus almond was essentially the first fluid other than water that the blocked groups had ever drunk, and the other possible factor was an increase in palatability of this solution for intermixed groups due to generalization from the 20 %Malto that these rats had drunk in the previous two sessions.
A similar 2 × 2 ANOVA was applied to the consumption of fluid on the “good” days of training, again with Training and Nutrient as factors. On these days, rats in the maltodextrin group received 20 %Malto, while rats in the saccharin group received 3 %Malto plus saccharin. There was a significant effect of training, whereby rats in the blocked groups drank significantly more fluid than did rats in the intermixed groups, F(1, 28) = 13.5, p < .01. However, no effect of nutrient was found, such that rats drank similar amounts of 20 %Malto and 3 %Malto plus saccharin, regardless of training, F = 1.2, p > .05. The two factors did not interact, F < 1. It may be noted that the absence of differences in consumption on good days between 20 %Malto and 3 %Malto plus saccharin was expected because of the approximate 12-ml limit on the amounts of fluid available. As for the training effect, one possible reason why the blocked groups would drink more than the intermixed groups was the same as one suggested above for the training effect found for intakes of 3 %Malto plus almond: The low intakes of both of the highly palatable solutions by the intermixed groups in Session 1 probably reflect neophobia. On the other hand, there is no obvious reason why the blocked groups would drink so much in Session 4 or why they would drink more than the intermixed groups on Session 5.
As for total fluid intakes over the two test sessions, in which rats had a choice between 6 %Malto plus almond and unflavored 6 %Malto, Group Intermixed-Sacch (M = 13.8 ml, SD = 3.6) drank more than Group Blocked-Sacch (M = 11.5 ml, SD = 2.6), and Group Blocked-Malto (M = 12.5 ml, SD = 4.0) drank more than Group Intermixed-Malto (M = 9.9 ml, SD = 3.8). These differences were confirmed by a 2 × 2 ANOVA with Nutrient and Training as the between-subjects factors, which did not reveal main effects of either nutrient or training, largest F = 1.2, but did show an interaction, F(1, 28) = 10.7, p = .04. Analysis of the simple effects confirmed that both the differences detailed above were significant, ps < .05. These differences should be borne in mind in relation to the analysis of preferences that is reported next.
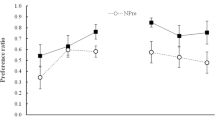
As is shown in Fig. 2, preferences for almond revealed that in the maltodextrin condition, intermixed training reduced almond preference relative to blocked training—that is, a missing-calorie effect—but no such effect was detected in the saccharin condition. This was confirmed by a 2 × 2 ANOVA in which we found no main effect of nutrient or training, both Fs < 1, but a nutrient by training interaction, F(1, 28) = 6.7, p < .05. Analysis of the simple effects confirmed that almond preferences in the maltodextrin condition were greater in rats that had received blocked training than in those given intermixed training, p < .05, but there was no effect of training in the saccharin group, p > .05.

Experiment 1: Almond preferences in the two-bottle test between 6 % maltodextrin plus 1 % almond and unflavored 6 % maltodextrin
As noted above, during training the blocked groups had drunk less 3 %Malto plus almond, but more of the highly palatable solutions, than had the intermixed groups (see Fig. 1), but no effect of nutrient was found. To the extent that a greater intake of the almond solution during training might increase almond preference on test—a mere exposure effect—this should have tended to increase almond preferences in both intermixed groups relative to both blocked groups. Since this was not the pattern of results obtained at test, we concluded that differences in training intakes did not have a major impact on almond preferences.
In summary, the main finding from this experiment was that almond avoidance was greater when this flavor had signaled the absence of a calorie-rich solution, 20 %Malto, than when almond had signaled the absence of an arguably more palatable, but less nutritious, solution, 3 %Malto plus saccharin.
Experiment 2
The previous experiment failed to detect in food-deprived rats any effect of intermixed training based on palatability differences. A study of flavor preference learning in which rats were given an almond–sucrose mixture has suggested that, when trained while food-deprived, they acquired a flavor–calorie association but, when trained while sated, they acquired a flavor–flavor association (Harris, Gorissen, Bailey, & Westbrook, 2000). Consequently, it seemed possible that flavor avoidance learning based on a palatability difference might be detected when rats were sated throughout the experiment. Experiment 2 tested this possibility. In addition, the use of the same design as in Experiment 1 meant that a comparison between Groups Intermixed-Malto and Blocked-Malto could reveal whether a missing-calorie effect—that is, greater avoidance of almond by Group Intermixed-Malto than by Group Blocked-Malto—can be produced even when rats are not food-deprived.
Method
Subjects
A group of 31 experimentally naïve hooded Wistar rats from the University of Adelaide were approximately 70 days old at the start of the experiment, with a mean weight of 358 g, range 273–408 g. (No more rats of about this age were available at the time of the experiment.) They were housed eight to a cage, except for one squad of seven. All of the rats had access to unrestricted food and water, except for the removal of water bottles 2 h before a session. The intention here was to ensure that the rats’ stomachs contained little or no food or water at the start of a session, but nonetheless the rats would be neither hungry nor thirsty, especially since the sessions were held during the light phase.
Rats were allocated to groups such that the first squad contained three rats in Group Intermixed-Sacch and four in Group Intermixed-Malto, the second and third squads each contained four rats in Groups Blocked-Sacch and Blocked-Malto, and the final squad contained four rats in Group Intermixed-Malto and four in Group Intermixed-Sacch. Thus, there were eight rats in three of the groups, but only seven rats in Group Intermixed-Sacch.
Apparatus and solutions
The drinking chambers were those used in the previous experiment. The solutions were also the same as in Experiment 1, except in the test, as noted below.
Procedure
The training procedure was identical to that used in Experiment 1: Six sessions of either intermixed or blocked single-bottle training, followed by two sessions in which rats were given 6 %Malto in each of two bottles. The only change was to the test procedure—namely, to give rats a choice between 3 %Malto plus almond and unflavored 3 %Malto. This change was made because it was possible that combining almond and 6 %Malto in Experiment 1 may have served as a brief counterconditioning procedure and, as such, have increased preference for almond on the second of the two test sessions. As in Experiment 1, in Session 9 the almond solution was on the left, with unflavored 3 %Malto on the right, and the positions of these solutions were reversed in Session 10.
Results and discussion
Fluid intakes during training are shown in Fig. 3, with the intermixed groups shown in the upper panel and the blocked groups in the lower panel. It may be noted that the general patterns of these training data are very similar to those in the training data from the previous experiment, shown in Fig. 1. In particular, the saccharin and maltodextrin groups drank very similar amounts, whereas the intermixed groups drank more 3 %Malto plus almond (M = 7.9 ml) than did the blocked groups (M = 5.0 ml). This summary was confirmed by a 2 × 2 ANOVA with Training (intermixed vs. blocked) and Nutrient (saccharin vs. maltodextrin) as factors. As in Experiment 1, no main effect of nutrient emerged, F < 1, but a significant effect of training was found, F(1, 27) = 10.7, p < .05, that did not interact with nutrient, p > .05. As suggested for the similar difference observed in Experiment 1, the lower intakes of 3 %Malto plus almond in the blocked groups probably reflected greater neophobia toward this solution when it was given to these groups in Sessions 1–3 than when it was given to the Intermixed groups in Sessions 3, 4, and 6.

Experiment 2: Fluid intakes during training. The upper panel shows intakes by the two groups given intermixed training, in which they received 3 % maltodextrin plus almond in Sessions 3, 4, and 6 and either 3 % maltodextrin plus 0.1 % saccharin (Group Intermixed-Sacch) or 20 % maltodextrin (Group Intermixed-Malto) in Sessions 1, 2, and 5. The lower panel shows intakes by the two groups given blocked training, who were given 3 % maltodextrin plus almond in Sessions 1–3 and either 3 % maltodextrin plus 0.1 % saccharin (Group Blocked-Sacch) or 20 % maltodextrin (Group Blocked-Malto) in Sessions 4–6
The only major difference between these training data and those from Experiment 1 was that in the blocked conditions (lower panels of Figs. 1 and 3) the sated rats in the present experiment drank less of the palatable solutions, 20 %Malto and 3 %Malto plus saccharin, when these were first introduced in Session 4 than had the food-deprived rats in Experiment 1. This is consistent with the pilot data reported in the introduction. These results suggested that the palatability of both 20 %Malto and 3 %Malto plus saccharin is greater under conditions of food deprivation (as in Exp. 1) than when rats are sated, as in the present experiment. A 2 × 2 ANOVA applied to fluid intakes on the “good” days of training, when the maltodextrin groups received 20 %Malto and the saccharin groups received 3 %Malto plus saccharin, found no significant effects of either training or nutrient, largest F = 1.5, p > .05, and no interaction, F < 1.
Over the two test sessions, the blocked groups drank more (10.0 ml) than did the intermixed groups (8.1 ml). This difference was confirmed by a 2 × 2 ANOVA, with Nutrient and Training as the between-subjects factors, that revealed a main effect of training, F(1, 27) = 12.5, p < .05, but no main effect of nutrient, F < 1, and no interaction between nutrient and training, F(1, 27) = 1.9, p < .05. Since the intake difference between the blocked groups and the intermixed groups was relatively small, and thus unlikely to bias preferences, a subsequent analysis was performed on almond preferences.
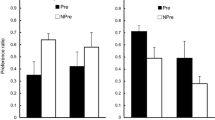
As is shown in Fig. 4, in the maltodextrin condition, intermixed training reduced almond preference relative to blocked training, whereas in the saccharin condition, intermixed training increased almond preference relative to blocked training. This was confirmed by a 2 × 2 ANOVA in which we found a main effect of nutrient, F(1, 27) = 10.0, p < .04, such that almond preferences were higher in the saccharin groups than in the maltodextrin groups, and an interaction between nutrient and training, F(1, 27) = 16.7, p < .01, but no main effect of training, F(1, 27) = 2.6, p < .05. Analysis of the simple effects confirmed that almond preferences in the maltodextrin condition were lower in rats that had received intermixed training than in those given blocked training, p < .05, and that almond preferences in the saccharin condition were higher for rats that had received intermixed training than for those that had received blocked training, p < .01. It may be noted that the same pattern of test data had been obtained in Experiment 1 (see Fig. 2), but in that case, although in the saccharin condition almond preferences were higher in the intermixed rats than in the blocked rats, this difference did not reach significance.

Experiment 2: Almond preferences in the two-bottle test between 3 % maltodextrin plus 1 % almond and unflavored 3 % maltodextrin
As we discussed in relation to the similar pattern of test results from Experiment 1, the somewhat greater consumption of 3 %Malto plus almond by the intermixed groups appears unlikely to have influenced these almond preferences. Although in the saccharin condition these preferences were higher for the intermixed than for the blocked groups—a direction consistent with a mere exposure effect during training—in the maltodextrin condition they were in the opposite direction.
With regard to the maltodextrin groups, the finding of almond avoidance in sated rats was somewhat unexpected, in the light of flavor preference studies in which motivational conditions have been varied. Several researchers have reported that flavor preferences based on pairing a flavor with sucrose are expressed more strongly at test when rats are food-deprived (e.g., Capaldi, Owens, & Palmer, 1994; Fedorchak & Bolles, 1987; Harris et al. 2000). Furthermore, some of these authors have suggested that the flavor–calorie learning produced when rats drink a flavored sucrose solution depends on the rats being hungry to some extent during training (Capaldi et al. 1994; Harris et al. 2000). On the other hand, flavor preference learning based on pairing a flavor with intragastric infusion of maltodextrin can take place even in sated rats (Yiin et al. 2005). Combined with the present results, the latter finding indicates that rats are able to monitor the energy content of a drink even when they have no current need to do so.
In summary, the main results from this experiment were, first, finding the missing-calorie effect even in rats that were not food-deprived and, second, finding that in the saccharin condition intermixed training increased, rather than decreased, preference for almond.
General discussion
As we explained in the introduction, the main question addressed by these experiments was whether avoidance of a target flavor could be produced by training in which rats were provided with a highly palatable solution on some sessions that were intermixed with sessions in which a less palatable solution was available, in the absence of any nutritional difference between these solutions. No evidence was obtained for such learning. Thus, in both experiments, for rats in Group Intermixed-Sacch the presence of almond in a 3 %Malto solution signaled the absence of the highly palatable solution of 3 %Malto plus 0.1 % saccharin, and yet these rats still developed a preference for almond. This outcome was found under exactly the same conditions in which training that did involve a nutritional difference—namely, 20 %Malto versus 3 %Malto plus almond—produced avoidance of almond.
The failure to find flavor avoidance based on a palatability difference alone has to leave open the possibility that some other method might be more effective than the present one at identifying such a difference. For example, instead of using saccharin to enhance the palatability of a solution without impacting its nutritional value, one might test the effect of intermixing 20 %Malto and 3 %Malto-plus-almond sessions under conditions in which the nutritional difference between the two solutions was neutralized. One possibility would be to introduce a fistula that prevented either solution from reaching a rat’s stomach. A less familiar method would be to introduce a drug, such as acarbose, that prevents absorption of nutrients from maltodextrin solutions in rats (Elizalde & Sclafani, 1988). Until such experiments are undertaken, the present results point to the conclusion that the flavor avoidance produced when two solutions do differ in nutritional value as well as in palatability is probably based mainly, if not entirely, on the nutritional difference.
An unexpected finding from Experiment 2, and marginally from Experiment 1, was that in the saccharin condition, almond preference was greater following intermixed than following blocked training. We tentatively suggest that this finding probably arose as a result of second-order simultaneous conditioning (Rescorla, 1973), since almond is an essentially neutral flavor and 3 %Malto is only moderately palatable, while 0.1 % saccharin is a strong unconditioned stimulus known to support flavor preference learning. Thus, it is possible that intermixed training produces strong first-order conditioning (flavor–flavor learning) that enhances the palatability of 3 %Malto in the three sessions in which saccharin is added to 3 %Malto and second-order conditioning of an almond preference in the three sessions in which almond is added to 3 %Malto. This could not happen in Group Blocked-Sacch because all three 3 %Malto-plus-almond sessions here were given prior to the three 3 %Malto-plus-saccharin sessions. This last arrangement would allow for a possible sensory preconditioning effect, but such effects may be weaker than second-order conditioning. In the absence of relevant flavor-learning research, this account remains entirely speculative.
One implication of this proposal is that in the saccharin condition, a higher preference for almond following intermixed than following blocked training would not have been found if the sequence of conditions in the blocked condition had been reversed. Interestingly, in previous unpublished experiments that have included conditions similar to those given to Groups Intermixed-Sacch and Blocked-Sacch in the present experiments, but in which the control condition introduced a block of “good” sessions prior to the block of “bad” sessions, we did not detect an increased flavor preference in Group Intermixed-Sacch.
While Experiment 1 replicated the effect previously reported by Boakes et al. (2010) and Experiment 2 demonstrated that this effect could be obtained even in nondeprived rats, in both experiments the degree of almond avoidance was small relative to that in the original report. This probably occurred because the training was brief—six sessions—in the present experiments, as compared to the 12 sessions of training that had been used previously. Even though the effect was relatively small, the present results support our previous application of the label missing-calorie effect to this kind of learned flavor avoidance. We should note, however, that this label is not intended to imply that animals will learn to avoid a flavor signaling a reduction in calories, whatever the source. For example, fructose does not normally support flavor–calorie learning, and this appears to be because it fails to generate a postingestive nutrient reward signal (Ackroff, Touzani, Peets, & Sclafani, 2001; Sclafani, Cardieri, Tucker, Blusk, & Ackroff, 1993); consequently, it seems unlikely that intermixed training of the kind used here, but with fructose substituted for maltodextrin, would produce a missing-calorie effect.
In summary, the present study points to the conclusion that in the context of foods of high nutritional value—apart from possible exceptions such as fructose—animals tend to avoid flavors that they have learned signal food of relatively low nutritional value. It would be of interest to examine whether the same learning process is involved in relation to specific appetites, such as the selection of calcium-rich foods by diabetic rats (Tordoff, 2002).
References
Ackroff, K., Touzani, K., Peets, T. K., & Sclafani, A. (2001). Flavor preferences conditioned by intragastric fructose and glucose: Differences in reinforcement potency. Physiology and Behavior, 72, 691–703.
Albertella, L., & Boakes, R. A. (2006). Persistence of conditioned flavor preferences is not due to inadvertent food reinforcement. Journal of Experimental Psychology. Animal Behavior Processes, 32, 386–395.
Boakes, R. A., Colagiuri, B., & Mahon, M. (2010). Learned avoidance of flavors signaling reduction in a nutrient. Journal of Experimental Psychology. Animal Behavior Processes, 36, 117–125.
Capaldi, E. D., Owens, J., & Palmer, K. A. (1994). Effects of food deprivation on learning and expression of flavor preferences conditioned by saccharin or sucrose. Animal Learning and Behavior, 22, 173–180.
Davis, J. D. (1996). Microstructural analysis of the ingestive behavior of the rat ingesting polycose. Physiology and Behavior, 60, 1557–1563. doi:10.1016/S0031-9384(96)00321-6
Dwyer, D. M. (2008). Microstructural analysis of conditioned and unconditioned responses to maltodextrin. Learning & Behavior, 36, 149–158.
Elizalde, G., & Sclafani, A. (1988). Starch-based conditioned flavor preferences in rats: Influence of taste, calories and CS-US delay. Appetite, 11, 179–200.
Fanselow, M. S., & Birk, J. (1982). Flavor–flavor associations induce hedonic shifts in taste preference. Animal Learning and Behavior, 10, 223–228.
Fedorchak, P. M., & Bolles, R. C. (1987). Hunger enhances the expression of calorie- but not taste-mediated conditioned flavor preferences. Journal of Experimental Psychology. Animal Behavior Processes, 13, 73–79.
Feigin, M. B., Sclafani, A., & Sunday, S. R. (1987). Species differences in polysaccharide and sugar taste preferences. Neuroscience and Biobehavioral Reviews, 11, 231–240.
Harris, J. A., Gorissen, M. C., Bailey, G. K., & Westbrook, R. F. (2000). Motivational state regulates the content of learned flavor preferences. Journal of Experimental Psychology. Animal Behavior Processes, 26, 15–30.
Holman, E. W. (1975). Immediate and delayed reinforcers for flavor preferences in rats. Learning and Motivation, 6, 91–100.
Rescorla, R. A. (1973). Second order conditioning: Implications for theories of learning. In F. J. McGuigan & D. Lumsden (Eds.), Contemporary approaches to learning and conditioning (pp. 127–150). New York: Winston.
Sclafani, A. (1991). Conditioned food preferences. Bulletin of the Psychonomic Society, 29, 256–260.
Sclafani, A., Cardieri, C., Tucker, K., Blusk, D., & Ackroff, K. (1993). Intragastric glucose but not fructose conditions robust flavor preferences in rats. American Journal of Physiology, 265, R320–R325.
Sclafani, A., & Clyne, A. E. (1987). Hedonic responses of rats to polysaccharide and sugar solutions. Neuroscience and Biobehavioral Reviews, 11, 173–180.
Sclafani, A., Einberg, L. T., & Nissenbaum, J. W. (1987). Influence of saccharin on Polycose, sucrose, and glucose intake and preference in rats. Neuroscience and Biobehavioral Reviews, 11, 223–229.
Sclafani, A., & Nissenbaum, J. W. (1988). Robust conditioned flavor preferences produced by intragastric starch infusions in rats. American Journal of Physiology, 255, R672–R675.
Tordoff, M. G. (2002). Intragastric calcium infusions support flavor preference learning by calcium-deprived rats. Physiology and Behavior, 76, 521–529.
Yiin, Y.-M., Ackroff, K., & Sclafani, A. (2005). Flavor preferences conditioned by intragastric nutrient infusions in food restricted and free-feeding rats. Physiology and Behavior, 84, 217–231.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Boakes, R.A., Patterson, A.E. & Kwok, D.W.S. Flavor avoidance learning based on missing calories but not on palatability reduction. Learn Behav 40, 542–550 (2012). https://doi.org/10.3758/s13420-012-0074-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-012-0074-6