
Abstract
The present study investigated the automaticity of top-down instructions in visual search when the instruction was no longer actively implemented. To do so, we exploited the Priming of Pop-out (PoP) effect, a selection history phenomenon that reflects faster responses when the target and distractor colors are repeated than switched across trials of singleton search. We then had participants perform a color singleton search task where they implemented the instruction of imagining the opposite color of the previous target, which put the target colors underlying PoP and the imagery instruction in opposition. To assess automaticity, on some trials participants were instructed to stop implementing the imagery instruction. When the imagery instruction was implemented, responses were faster when the target and distractor colors switched (i.e., imagery congruent) than repeated (i.e., imagery incongruent) across search displays – a pattern of results opposite to the PoP effect. When participants were to not implement this instruction, the PoP effect was absent, indicating the imagery instruction had a lingering influence on visual search. This remained true even when participants reported successfully not implementing the instruction, and only when the imagery abandonment instruction was supplanted by a different top-down task was the lingering influence removed such that the PoP effect returned. Overall, the present study demonstrates that top-down instructions can continue to influence visual search despite the will of the observer.
Similar content being viewed by others
Introduction
When it comes to visual search, how it has been performed in the past can influence how it is performed in the present (e.g., Chun & Jiang, 1998; Maljkovic & Nakayama, 1994, 1996; Wang & Theeuwes, 2018). At the same time, visual search can also be influenced by the current goals of the observer (e.g., Found & Müller, 1996; Wolfe et al., 1989, 2003). While whether the top-down system influences visual search has been a historically contentious issue, in recent times it has become widely accepted that this system does indeed have an influence on visual search (Awh et al., 2012; Theeuwes, 2019). The caveat here is that the influence of the top-down system is often purported to be quite weak relative to that of the selection history system. In simple terms, the top-down system is characterized as effortful and deliberate and the selection history system is characterized as passive and automatic (Jonides, 1981; Maljkovic & Nakayama, 1994; Van der Stigchel et al., 2009).
A product of the selection history system that operates in a passive and automatic manner in visual search is the Priming of Pop-out (PoP) effect (Maljkovic & Nakayama, 1994). That is, if the observer must find the oddball-colored object amongst an array of homogeneously colored distractor objects then indicate whether a notch is missing out of the left or right side, responses are faster when the target and distractor colors repeat than switch across trials of the search task. Maljkovic and Nakayama (1994, 2000) theorized that the PoP effect reflects the passive maintenance of an attentional control setting that incorporates the relevant features of the search target, which then automatically guide attention to these features if repeated shortly thereafter (for a review, see Kristjánsson & Campana, 2010).
A pertinent issue concerns whether the automaticity of the PoP effect can endure top-down intervention. This issue was first explored by Maljkovic and Nakayama (1994) in two experiments. In Experiment 2, singleton color search was performed when the likelihood of a target and distractor color switch was varied across the blocks of trials. The rationale underlying this experiment was that if top-down expectancies can influence the processes underlying PoP, responses should be fastest when color switches and repeats were predictable (i.e., 100% switches or 100% repeats), and slowest when they were not (i.e., 50% switches/repeats). This was not what Maljkovic and Nakayama found – instead, response speed decreased with increased switch likelihood in an approximately linear manner, suggesting that top-down expectancy did not influence PoP. In Experiment 4, the target and distractor color switches and repeats varied in a predictable manner (e.g., red, red, green, green, red, red, etc.) both when the observers were aware of the target color sequence (i.e., the passive condition) and when they had to additionally subvocalize the upcoming target color prior to its onset (i.e., the active condition). It was revealed that the PoP effect was present throughout, and that its magnitude did not differ across the passive and active conditions – again suggesting that the top-down system does not affect PoP.
Some studies have since challenged these findings of Maljkovic and Nakayama (1994). Shurygina et al. (2019) demonstrated that the magnitude of the PoP effect was attenuated when the target and distractor color switches were predictable compared with when they were unpredictable across trials of color singleton search. Similarly, Pascucci et al. (2012) demonstrated that singleton color search targets were more accurately identified when briefly presented and followed by a metacontrast mask when the target and distractor color switches were predictable than when they were unpredictable. Thomson et al. (2013) demonstrated that the PoP effect in color singleton search was attenuated when the target and distractor colors switched 80% relative to 20% of the time; however, this result could have been due to the build-up of priming across consecutive trials when target and distractor color repetitions were more than less likely (see Cochrane & Pratt, 2020). In a replication of Maljkovic and Nakayama’s Experiment 4, we found that having participants say aloud the upcoming target color did significantly attenuate the PoP effect relative to when participants either had passive knowledge of the predictable color sequence or when the target and distractor color switches were unpredictable (Cochrane & Pratt, 2020). By varying whether participants had to find the oddball color or shape across trials of visual search, Fecteau (2007) demonstrated that the PoP effect was only present when the basis of the task also repeated; however, this result should be interpreted cautiously given the experimental procedure entailed that independent event file updating processes were working in opposition to PoP (see Hommel, 2004; Memelink & Hommel, 2013). An important takeaway from these studies is that, while they demonstrate that the PoP effect can be influenced by top-down interventions, barring putting independent processes in opposition with it (e.g., Fecteau, 2007), the PoP effect cannot be completely eliminated – suggesting that the processes underlying PoP, to some extent, operate in a passive and automatic manner.
As alluded to above, a pattern opposite to the PoP effect can be observed when color imagery is put in opposition. Generally, when color imagery is congruent with the color of an upcoming target, it can aid search performance (Cochrane et al., 2019; Cochrane, Townsend, Lee, et al., 2021; Cochrane, Wang, Pratt, et al., 2021; Moriya, 2018; Reinhart et al., 2015; cf. Clarke et al., 2016). Further, the potency of these imagery effects can be reflected by the reported vividness of the observers’ color imagery (Cochrane, Ng, Khosla, & Milliken, 2021; Cochrane, Ng, & Milliken, 2021; Cochrane, Nwabuike, et al., 2018). Specifically, we have assessed the influence of color imagery on the PoP effect by having participants generate color imagery that was opposite to the previous target color during the temporal interval between trials of a color singleton search task (Cochrane, Nwabuike, et al., 2018; Cochrane, Zhu, & Milliken, 2018). For example, if the target was the red object amongst green distractor objects in the first search display, participants were to imagine green prior to the second. While a typical PoP effect was present for a group of participants who did not implement this imagery instruction, it was reversed for a group of participants who did – responses were faster when the target color switched (i.e., imagery congruent) than repeated (i.e., imagery incongruent) across trials. Further, the PoP effect was absent in two out of three experiments when color imagery was reported as highly vivid, offering preliminary evidence that color imagery shared at least some of the processes responsible for PoP (Cochrane, Ng, & Milliken, 2021).
While color imagery, like other top-down processes, is certainly effortful and deliberate, it is an open issue whether these strategies can become automatic. That is, it may be that once a strategy is established, it is automatically implemented despite the will of the observer, akin to the typical effects produced by the selection history system. In support of this notion, Leber and Egeth (2006; see also Leber et al., 2009) had participants perform either a singleton search task (i.e., find the oddball shape amongst homogeneous distractors) or a feature search task (i.e., find the circle amongst mostly heterogeneous shape distractors) during the training phase of the experiment. During the test phase of the experiment, these participants had to find a circle amongst homogeneous shapes, which, importantly, could be performed using either singleton or feature search strategies. The prior work of Bacon and Egeth (1994) demonstrated that a color singleton distractor has less of an impact on feature than singleton search, and accordingly, Leber and Egeth evaluated whether those trained with feature search would also be less impacted by a color singleton distractor than those trained with singleton search. Leber and Egeth demonstrated that participants trained in feature search were indeed less impacted by the presence of a color singleton distractor than those trained on singleton search during the test phase, suggesting that top-down strategies can have a lingering influence on visual search.
As detailed above, it has been established that the PoP effect reflects a passive and automatic influence on search, and that effortful and deliberate top-down instructions (especially those using color imagery) can influence search when implemented. While other studies (e.g., Leber et al., 2009; Leber & Egeth, 2006) have demonstrated that strategies imposed by the basis of the search task can automatically influence future search, the present study evaluates whether an instruction that is independent of the search task can do so when no longer actively implemented. That is, we instructed participants to generate color imagery that was opposite in color to the previous target in the interval between trials of a color singleton search task. Importantly, on some trials participants were instructed to stop implementing the imagery instruction. If the top-down imagery instruction does not have an automatic influence on visual search, the PoP effect ought to emerge once the imagery instruction is not actively implemented. If the top-down imagery instruction does have an automatic influence on visual search, the PoP effect ought to be absent when the top-down instruction is no longer actively implemented since imagery should continue to have an oppositional influence. The present study will also investigate the robustness of the automaticity of top-down instructions and whether other top-down influences can disrupt them.
Experiment 1
In Experiment 1 search displays were presented in pairs, and prior to a pair of trials participants were cued to either imagine the opposite color of the first singleton search target during the temporal interval prior to the second search display (i.e., the imagine cue) or to not implement this imagery instruction (i.e., the respond cue). Accordingly, if the PoP effect was to emerge here, it would be consistent with the view that it reflects a passive and automatic process, and that the imagery instruction can be flexibly disengaged without a lingering influence.
Method
Participants
Thirty-two McMaster University undergraduates (25 female, seven male, aged 18–25 years, M = 19.5 years) took part in exchange for either course credit or $10 CAD monetary compensation. All participants reported normal or corrected-to-normal visual acuity and normal color vision. A sample size of 32 participants was selected to be identical to Cochrane, Ng et al. (2021; Experiment 1), which had sufficient power to detect a difference in inter-trial repetition effects across their vividness rating procedure (ηp2 = .36; alpha = .05; power = .95).
Apparatus and stimuli
Stimuli were presented using Psychopy v1.82 and displayed on a BenQ 24-in. LED monitor connected to a Mac Mini computer. Each search display contained one target square and four distractor squares, each of which subtended vertical and horizontal visual angles of 2°. Targets and distractors were presented in red and green. The five squares were randomly assigned to one of eight locations that were equidistant from the center of the screen. The distance from the center of the screen to each of these locations subtended a visual angle of 5°. All squares contained a gap in either the left or right side that subtended a visual angle of 0.5°. The white fixation cross subtended a horizontal and vertical visual angle of 0.3°.
Procedure
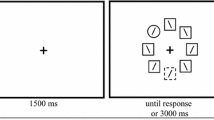
Participants were seated approximately 60 cm from the computer screen. Each trial sequence began with a cue stating “IMAGINE” or “RESPOND.” These cues were randomized on a trial-by-trial basis such that they were equally probable. Once participants read the cue, they pressed the spacebar to initiate the trial sequence. A black screen with a central white fixation cross was then displayed on-screen for 500 ms, followed by a color singleton search task. Here, participants had to find the odd-colored target square amongst four homogeneously colored distractor squares and indicate whether it contained a gap in its left or right side. The side of the gap was randomized on a trial-by-trial basis. Participants indicated that the gap was on the left by pressing the “Z” key with their left index finger and on the right by pressing the “M” key with their right index finger using a standard QWERTY keyboard. The search display remained on-screen until a response was made. A black screen with a central white fixation cross was then displayed for 2,000 ms, which was followed by another color singleton search trial. The target color switched across trials 80% of the time and repeated 20% of the time, which was to incentivize the use of the imagery strategy and does not eliminate the PoP effect (Cochrane, Nwabuike, et al., 2018). Following a response to the search task, participants were to rate the vividness of their imagery by pressing the “1” key for “no imagery,” the “2” key for “low vividness,” the “3” key for “moderate vividness,” and the “4” key for “high vividness.” An example of the trial sequence is depicted in Fig. 1.

An example of the trial sequence when the imagine cue was presented
Prior to the experimental session, participants performed 20 practice trials. For the first five practice trials, participants performed the trial sequence without the cue or imagery vividness ratings. In the next five practice trials, the imagine cue was presented on-screen at the beginning of each trial sequence and participants were instructed to imagine a square opposite in color to the target in the first search display during the temporal interval prior to the second. For example, if the target was red in the first display, participants were to imagine green, and vice versa. For the next five practice trials, the respond cue was presented at the beginning of each trial sequence, and participants were instructed to not use the imagery instruction, but to simply perform the search task. For the final five practice trials, the imagery and respond cues were randomly intermixed and participants were instructed on how to provide the imagery vividness ratings. Specifically, participants were instructed that the “no imagery” rating reflected the situation when “they did not generate any imagery,” the “low vividness” rating reflected the situation when their imagery was “vague and dim,” the “moderate vividness” rating reflected the situation when their imagery was “reasonably clear and vivid,” and the “high vividness” rating reflected the situation when their “imagery was clear and vivid like that of normal vision.” Participants were also informed that their ratings should be implemented in a relativistic manner to reflect their individual capability. Participants then performed 150 trial sequences. Following the experimental session, participants provided percentage estimates of how often they successfully generated imagery for the imagine cue and disengaged from it for the respond cue.
Results
Response times (RTs) and error percentages for the second trial of singleton search in the trial sequence were the primary dependent variables. RTs greater than 2,000 ms or less than 200 ms were excluded from analyses, which removed 2.15% of observations. Correct RTs were further excluded from analyses if they were identified as outliers by the non-recursive moving outlier elimination procedure of Van Selst and Jolicoeur (1994), which removed an additional 2.59% of observations. Mean correct RTs were computed from the remaining observations. An alpha criterion of .05 was used to determine statistical significance in all analyses. The mean post-experiment estimates of imagery use for the imagine cue was 73.08% and imagery instruction disengagement for the respond cue was 75.94%.
Overall results
The mean correct RTs and corresponding error percentages were submitted to within-subject ANOVAs that treated cue (imagine/respond) and inter-trial repetition (switch/repeat) as factors. The overall mean RTs and error percentages are depicted in Fig. 2.

The analysis of RTs revealed a significant two-way interaction of cue and inter-trial repetition, F(1,31) = 63.06, p < .001, η2p = .67. There was a significant main effect of inter-trial repetition, F(1,31) = 28.24, p < .001, η2p = .47, reflecting overall faster responses when the target color switched than repeated across trials. There was a significant main effect of cue, F(1,31) = 13.66, p < .001, η2p = .30, reflecting overall faster responses for the respond than imagine cue. The interaction was explored further by conducting planned paired t-tests that treated inter-trial repetition as a factor for the imagine and respond cues separately. The analysis of the imagine cue revealed a significant effect of inter-trial repetition, t(31) = 7.25, p < .001, d = 0.87, reflecting faster responses when the target color switched (M = 689 ms, SD = 242 ms) than repeated (M = 914 ms, SD = 274 ms) across trials. The analysis of the respond cue revealed no effect of inter-trial repetition, t(31) = 0.57, p = . 57, d = 0.05, reflecting similar RTs when the target color switched (M = 726 ms, SD = 190 ms) and repeated (M = 717 ms, SD = 236 ms) across trials.
For the analysis of error percentages, the interaction of cue and inter-trial repetition, F(1,31) = 0.70, p = .41, η2p = .02, the main effect of inter-trial repetition, F(1,31) = 0.56, p = .46, η2p = .02, and the main effect of cue, F(1,31) = 0.22, p = .64, η2p = .007, were not significant.
Disengagement results
Additional analyses were conducted to evaluate whether the PoP effect returned when participants reported no imagery. To do so, RT and error percentage analyses were only conducted on observations where “no imagery” was indicated for the respond cue and imagery of any vividness was indicated for the imagine cue. Participants reported “no imagery” on 65.70% of respond cue trials, and imagery of any vividness on 93.26% of imagine cue trials. The corresponding mean RTs and error percentages were submitted to a within-subject ANOVA that treated inter-trial repetition (repeat/switch) and cue (imagine/respond) as factors. The mean RTs and error percentages of the disengagement results are depicted in Fig. 3.

The analysis of RTs revealed a significant interaction of inter-trial repetition and cue, F(1,31) = 71.12, p < .001, η2p = .71. There was a significant main effect of inter-trial repetition, F(1,31) = 24.34, p < .001, η2p = .44, reflecting overall faster responses when the target color switched than repeated. There was a significant main effect of cue, F(1,31) = 14.66, p < .001, η2p = .32, reflecting overall faster responses for the respond than imagine cue. The interaction was explored further by conducting planned paired t-tests that treated inter-trial repetition as a factor for the imagine and respond cues separately. The analysis of the imagine cue revealed a significant effect of inter-trial repetition, t(31) = 7.09, p < .001, d = 0.91, reflecting faster responses when the target color switched (M = 687 ms, SD = 244 ms) than repeated (M = 927 ms, SD = 280 ms) across trials. The analysis of the respond cue revealed no effect of inter-trial repetition, t(31) = 0.93, p = .36, d = 0.09, indicating there were similar RTs when the target color switched (M = 713 ms, SD = 188 ms) and repeated (M = 693 ms, SD = 229 ms) across trials.
For the analysis of error percentages, there was no interaction of cue and inter-trial repetition, F(1,31) = 1.06, p = .31, η2p = .03, main effect of inter-trial repetition, F(1,31) = 1.35, p = .25, η2p = .02, or main effect of cue, F(1,31) = 0.13, p = .72, η2p = .004.
Discussion
First, the present experiment revealed that when the imagery instruction was implemented, responses were faster when the target color switched (i.e., imagery congruent) than repeated (i.e., imagery incongruent), demonstrating that imagery can produce an effect opposite to PoP. Second, when participants were cued to not implement the imagery instruction, neither the PoP nor the imagery congruency effect was observed. It is important to highlight that this study used an experimental procedure that was very similar to Cochrane, Nwabuike et al. (2018; Experiment 1a), which demonstrated a robust PoP effect for participants who never generated imagery (77 ms; d = 1.13). Third, we assessed whether the imagery instruction was better disengaged from when participants reported that they did not generate imagery following the trial sequence. Here the PoP effect was still absent, suggesting that imagery instruction disengagement was consciously inaccessible to participants (although see Cochrane, Ng, Khosla, & Milliken, 2021). However, the mean post-experiment estimate of imagery instruction disengagement was 75.94%, indicating that participants indicated that the instruction was not readily abandoned. Overall, the present findings suggest that prior instructions continue to influence search even when not actively implemented.
Experiment 2
A possible reason for participants being unable to fully disengage from the imagery instruction is that the imagine and respond cues were randomly implemented on a trial-by-trial basis. That is, it could be that when the imagery instruction was recently used, it interferes with the participants’ ability to disengage from it. Accordingly, Experiment 2 assessed whether the imagery instruction continued to influence performance long after it was last cued. If the imagery instruction has a long-lasting influence on search, participants instructed to disengage from imagery should continue to be influenced by the imagery instruction even when it was last cued as many as 150 trial sequences earlier.
Method
Participants
Thirty-two McMaster University undergraduates (28 female, four male, aged 18–23 years, M = 18.75 years) took part in exchange for either course credit or $10 CAD monetary compensation. All participants reported normal or corrected-to-normal visual acuity and normal color vision. While the sample size was selected to be identical to the previous experiment, it is worth noting that the power analysis reported in Experiment 1 is not directly applicable given the additional block order factor.
Apparatus and stimuli
The apparatus and stimuli were identical to the previous experiment.
Procedure
The procedure was identical to the previous experiment with the exception that participants now performed two blocks of 150 trial sequences each. For one block, only the imagine cue was presented, and for the other, only the respond cue was presented. Block order was counter-balanced across participants. Across both blocks, participants were required to rate the vividness of their color imagery following each trial sequence, even though the participants who performed the respond cue block first were not briefed on the imagery instruction. Accordingly, these participants should have been unable to adequately rate the vividness of their visual imagery as it pertains to the “imagine the opposite color” instruction. Participants were briefed on the imagery and respond cue instructions at the start of the corresponding block, at which point they performed ten practice trials.
Results
RTs and error percentages for the second search task of the trial sequence were the primary dependent variables. RTs greater than 2,000 ms or less than 200 ms were excluded from analyses, which removed 1.51% of observations. The non-recursive moving outlier elimination procedure of Van Selst and Jolicoeur (1994) removed an additional 3.18% of observations. Mean correct RTs were computed from the remaining observations. An alpha criterion of .05 was used to determine statistical significance in all analyses. The mean post-experiment estimates of imagery use during the imagine block was 74.39%.
Overall results
The mean correct RTs and the corresponding error percentages were submitted to mixed factor ANOVAs that treated cue block (imagine/respond) and inter-trial repetition (switch/repeat) as within-subject factors, and block order (imagine-first/imagine-second) as a between-subjects factor. The mean RTs and error percentages are depicted in Fig. 4.

The analysis of RTs revealed no three-way interaction of cue block, inter-trial repetition, and block order, F(1,30) = 0.04, p = .85, η2p = .001. There was no two-way interaction of inter-trial repetition and block order, F(1,30) = 0.88, p = .36, η2p = .03, or cue and block order, F(1,30) = 0.15, p = .70, η2p = .04. There was a significant two-way interaction of inter-trial repetition and cue block, F(1,30) = 39.08, p < .001, η2p = .57. There was no main effect of block order, F(1,30) = 1.11, p = .30, η2p = .04. There was a significant main effect of inter-trial repetition, F(1,30) = 18.05, p < .001, η2p = .37, reflecting overall faster responses for switch than repeat trials. There was a significant main effect of cue block, F(1,30) = 25.72, p < .001, η2p = .46, reflecting overall faster responses for the respond than imagine cue block. Given a priori interests, mixed factor ANOVAs that treated block order and inter-trial repetition as factors were conducted for the imagine and respond cue blocks separately.
The analysis of the imagine cue block revealed a significant main effect of inter-trial repetition, F(1,30) = 28.40, p < .001, η2p = .49, reflecting faster responses when the target color switched (first-block: M = 732 ms, SD = 147 ms; second-block: M = 669 ms, SD = 77 ms) than repeated (first-block: M = 925 ms, SD = 220 ms; second-block: M = 907 ms, SD = 291 ms) across trials. The main effect of block order, F(1,30) = 0.48, p = .49, η2p = .02, and interaction of inter-trial repetition and block order, F(1,30) = 0.32, p = .58, η2p = .01, were not significant.
The analysis of the respond cue block revealed a significant main effect of inter-trial repetition, F(1,30) = 23.50, p < .001, η2p = .43, reflecting overall faster responses when the target color repeated than switched across trials. There was no effect of block order, F(1,30) = 0.19, p = .66, η2p = .006. There was an interaction of inter-trial repetition and block order that approached significance, F(1,30) = 3.72, p = .063, η2p = .11. This finding was explored further by conducting paired t-tests that treated inter-trial repetition as a factor for each level of block order separately. When the respond cue block was performed first, there was a significant effect of inter-trial repetition, t(15) = 5.26, p < .001, d = 0.76, reflecting faster responses when the target color repeated (M = 662 ms, SD = 64 ms) than switched (M = 715 ms, SD = 74 ms) across trials. When the respond cue block was performed second, the effect of inter-trial repetition only approached significance, t(15) = 1.91, p = .076, d = 0.27, reflecting statistically similar RTs when the target color repeated (M = 666 ms, SD = 89 ms) and switched (M = 688 ms, SD = 81 ms) across trials.
The analysis of error percentages revealed a significant main effect of block, F(1,30) = 9.99, p = .004, η2p = .24, reflecting higher error percentages for the imagine than respond cue block, and a main effect of inter-trial repetition, F(1,30) = 15.8, p < .001, η2p = .34, reflecting higher error percentages when the target color switched than repeated. No other effects in the analysis of error percentages were significant (all F ≤ 3.13, p ≥ .087).
Disengagement results
Like in Experiment 1, additional analyses were conducted only on observations where “no imagery” was indicated for the respond cue and some form of imagery was indicated for the imagine cue. Participants reported “no imagery” on 91.89% (response block first: 93.89%; response block second: 89.86%) of respond cue trials, and imagery of some vividness on 91.64% (imagery block first: 92.82%; imagery block second: 90.47%) of imagine cue trials. The corresponding mean RTs were submitted to a mixed factor ANOVA that treated inter-trial repetition (repeat/switch) and cue block (imagine/respond) as within-subject factors and block order (imagery-first/imagery-second) as a between-subjects factor. The mean RTs and error percentages of the disengagement results are depicted in Fig. 5.

This analysis revealed that the three-way interaction of inter-trial repetition, cue block, and block order was not significant, F(1,30) = 0.10, p = .75, η2p = .003. There was no two-way interaction of cue and cue block, F(1,30) = 0.04, p = .83, η2p = .002, or inter-trial repetition and cue block, F(1,30) = 0.88, p = .35, η2p = .03. There was a significant two-way interaction of inter-trial repetition and cue, F(1,30) = 38.45, p < .001, η2p = .56. There was no main effect of cue block, F(1,30) = 1.16, p = .29, η2p = .04. There was a significant main effect of inter-trial repetition, F(1,30) = 19.56, p < .001, η2p = .39, reflecting overall faster responses when the target color switched than repeated across trials. There was a significant main effect of cue block, F(1,30) = 22.35, p < .001, η2p = .56, reflecting overall faster responses for the respond than imagine cue block. Given a priori interests, mixed factor ANOVAs that treated block order and inter-trial repetition as factors were conducted for the imagine and respond cue blocks separately.
The analysis of the imagine cue block revealed a significant main effect of inter-trial repetition, F(1,30) = 29.09, p < .001, η2p = .49, reflecting faster responses when the target color switched (first-block: M = 726 ms, SD = 144 ms; second-block: M = 661 ms, SD = 73 ms) than repeated (first-block: M = 932 ms, SD = 227 ms; second-block: M = 923 ms, SD = 320 ms) across trials. There was no main effect of block order, F(1,30) = 0.37, p = .55, η2p = .01, or interaction of inter-trial repetition and block order, F(1,30) = 0.41, p = .53, η2p = .01.
The analysis of the respond cue block revealed a significant main effect of inter-trial repetition, F(1,30) = 24.3, p < .001, η2p = .44, reflecting overall faster responses when the target color repeated than switched across trials. There was no main effect of block order, F(1,30) = 0.65, p = .43, η2p = .02, and interaction of inter-trial repetition and block order, F(1,30) = 3.06, p = .090, η2p = .09. This finding was explored further by conducting paired t-tests that treated inter-trial repetition as a factor for each level of block order separately. When the respond cue block was performed first, there was a significant effect of inter-trial repetition, t(15) = 4.73, p < .001, d = 0.72, reflecting faster responses when the target color repeated (M = 666 ms, SD = 70 ms) than switched (M = 719 ms, SD = 74 ms) across trials. When the respond cue block was performed second, there was a significant effect of inter-trial repetition, t(15) = 2.25, p = .040, d = 0.30, reflecting faster responses when the target color repeated (M = 660 ms, SD = 84 ms) than switched (M = 684 ms, SD = 81 ms) across trials.
The analysis of error percentages revealed a significant interaction of inter-trial repetition and block order, F(1,30) = 4.81, p = .036, η2p = .14. There was a significant main effect of inter-trial repetition, F(1,30) = 9.51, p = .004, η2p = .24, reflecting higher error percentages when the target color switched than repeated. No other effects in the analysis of error percentages were significant (all F ≤ 3.61, p ≥ .067).
Discussion
First, the imagery congruency effect was robust when participants implemented the imagery instruction, regardless of whether the imagery cue block occurred first or second. Second, when the respond cue occurred first, and, therefore, participants were unaware of the imagery instruction, a robust PoP effect was present. Third, while the PoP effect approached significance when the respond cue block was performed after the imagine cue block, its magnitude was greatly reduced relative to when the respond cue block was performed without the knowledge of the imagery instruction. All told, the present results suggest that top-down instructions can be persistent and not so easily extinguished – they continue to influence the PoP effect long after the task requirements had led them to be abandoned.
Experiment 3
At this point, it has been revealed that top-down instructions have a lingering influence on visual search, while at the same time, the basis of this phenomenon is unclear. What we do know is this top-down influence is operating outside of conscious awareness, as it cannot be willfully abandoned. What we don’t know is how this phenomenon is affected by other top-down tasks. That is, it may be that when an instruction is implemented, it will remain intact until supplanted, and this posed a particular problem for the PoP effect since the imagery instruction was in direct opposition with it. If a top-down task can be disrupted by another, the PoP effect should return when the supplanting task does not affect it. To evaluate this possibility, we replicated the experimental procedure of Experiment 1, except that instead of being instructed to not implement the imagery instruction, participants were to repeat a random word aloud during the interval between search displays of the trial sequence.
Method
Participants
Thirty-two McMaster University undergraduates (21 female, 11 male, aged 18–33 years, M = 19.75 years) took part in exchange for either course credit or $10 CAD monetary compensation. All participants had normal or corrected-to-normal visual acuity and normal color vision. The sample size was selected based on the previous experiments.
Apparatus and stimuli
The apparatus and stimuli were identical to the previous experiments.
Procedure
The procedure was identical to Experiment 1 except that the respond cue was replaced by the word cue. For the word cue, a random five-letter word of medium frequency (e.g., “PAINT,” “CLOUD,” etc.) was displayed at the beginning of the trial pair sequence and participants were to repeatedly say this word aloud during the temporal interval between the pair of search displays. The word and imagine cues were randomly intermixed across the experimental session. The imagine cue now contained three asterisks on each side (i.e., “***IMAGINE***”) to distinguish it from the word cue. For the word cue trial sequences, the imagery vividness probe was replaced by ratings that evaluated how well participants implemented the word cue instruction. Here, a keypress of “1” indicated “not at all,” a keypress of “2” indicated “a little bit,” a keypress of “3” indicated “moderately well,” and a keypress of “4” indicated “to the best of my ability.”
Results
RTs and error percentages for the second trial of singleton search of the trial sequence were the primary dependent variables. RTs greater than 2,000 ms or less than 200 ms were excluded from analyses, which removed 2.21% of observations. The non-recursive moving outlier elimination procedure of Van Selst and Jolicoeur (1994) removed an additional 2.57% of observations. Mean correct RTs were computed from the remaining observations. An alpha criterion of .05 was used to determine statistical significance in all analyses. The mean post-experiment estimates of imagery use for the imagine cue was 73.11% and implementation of the word cue instruction for the word cue was 88.94%. Given that participants rarely indicated failing to perform the word task instructions (i.e., 0.01% of observations), the Disengagement Results section was not included.
Overall results
The mean correct RTs and corresponding error percentages were submitted to within-subject ANOVAs that treated cue (imagine/word) and inter-trial repetition (switch/repeat) as factors. The overall mean RTs and error percentages are depicted in Fig. 6.

The analysis of RTs revealed a significant main effect of inter-trial repetition, F(1,31) = 26.50, p < .001, η2p = .24, reflecting overall faster responses when that target color switched than repeated across trials. There was a significant main effect of cue, F(1,31) = 11.99, p = .002, η2p = .28, reflecting overall faster responses when the word than imagine cue was presented. There was a significant interaction of cue and inter-trial repetition, F(1,31) = 26.50, p < .001, η2p = .49. This interaction was explored further by conducting planned paired t-tests that treated inter-trial repetition as a factor for the imagine and word cues separately. The analysis of the imagine cue revealed a significant effect of inter-trial repetition, t(31) = 4.55, p < .001, d = 0.49, reflecting faster responses when the target color switched (M = 808 ms, SD = 238 ms) than repeated (M = 939 ms, SD = 297 ms) across trials. The analysis of the word cue revealed a significant effect of inter-trial repetition, t(31) = 3.19, p = .003, d = 0.22, reflecting faster responses when the target color repeated (M = 761 ms, SD = 171 ms) than switched (M = 800 ms, SD = 183 ms) across trials.
For the analysis of error percentages, there was no interaction of cue and inter-trial repetition, F(1,31) = 0.69, p = .41, η2p = .02, main effect of inter-trial repetition, F(1,31) = 0.52, p = .42, η2p = .01, or main effect of cue, F(1,31) = 1.97, p = .17, η2p = .06.
Discussion
First, and as in previous experiments, when participants were cued to implement the imagery instruction, a robust imagery congruency effect emerged. Second, when participants performed the word task instruction, a robust PoP effect emerged. This was despite participants having the knowledge of and recently implementing the imagery instruction. Accordingly, it appears that the imagery instruction can lead to automatic influences on visual search, but its automaticity can be overridden when the top-down system is occupied by another task.
General discussion
The present study evaluated whether top-down instructions can have an automatic influence on visual search. To assess this issue, participants imagined a color that was opposite to the previous target of a color singleton search task during the temporal interval between search displays. Automaticity was then assessed by evaluating whether the PoP effect emerged when participants were instructed to stop implementing the imagery instruction. It was revealed that when participants implemented the imagery instruction, responses were faster when the target color switched (i.e., imagery congruent) than repeated (i.e., imagery incongruent) across trials. When participants were cued to not use the imagery instruction, the PoP effect was not observed, suggesting that the imagery instruction influenced search. Further, it appeared that participants did not have conscious access to the influence of the imagery instruction, since the PoP effect did not emerge even when participants reported successfully not implementing it. Even further, the imagery instruction continued to influence search long after it was last cued, as the magnitude of the PoP effect was greatly reduced compared to participants who had no knowledge of the imagery instruction when they performed the search task. It was only when the imagery disengagement instruction was replaced by an extraneous top-down task that a robust PoP effect emerged when participants had recently implemented the imagery instruction. Overall, the present findings suggest that top-down instructions can lead to automatic influences on search unless supplanted by another top-down task.
A consistent finding across all experiments of the present study was that responses were especially slow when the target color was incongruent with color imagery. Accordingly, it seems that the present findings were primarily due to an incongruency cost rather than a faciliatory benefit. However, we have demonstrated elsewhere that color imagery can produce a faciliatory benefit when it is congruent with an upcoming target, and a cost when it is congruent with a distractor (Cochrane, Wang, et al., 2021). We believe that a consistent faciliatory benefit was not observed in the present study because the processes underlying imagery congruency and PoP tap into the same system integral to attentional guidance. Indeed, we have demonstrated elsewhere that when imagery is reported as being highly vivid, in two out of three experiments, the magnitude of imagery congruency effects were statistically identical when the previous target color was repeated and switched (Cochrane, Ng, & Milliken, 2021). In other words, the lack of an additive contribution from PoP suggests that imagery is overriding its influence, and, thus, there is overlap of the underlying processes. While that explains the absence of the faciliatory benefit, why were there large incongruency costs for imagery? We suspect this has to do with imagery producing a stickier form of attention capture, so to speak. That is, while color imagery and target repetitions both direct spatial attention to the location of congruent colors, they are not easily disengaged from when the templates are constructed with imagery. This is not problematic when imagery and the target color are congruent since disengagement is not required. However, when imagery is incongruent with the target (i.e., congruent with the distractors), participants are required to disengage from the distractor prior to orienting and identifying the target. While the reason for this phenomenon is not yet clear, we suspect that it is due to the active basis of imagery – that active maintenance of a representation in memory strengthens attentional engagement to congruent perceptual representations.
There are at least two plausible ways that the top-down color imagery instruction influenced visual search when participants were no longer actively implementing it. The first is the interference hypothesis – that once participants sufficiently implemented the imagery instruction, the corresponding color representation was incorporated into the attentional control setting. This then interfered with the color representations incorporated into the attentional control setting produced by the previous target color, such that attention could not be guided sufficiently to either representation. In other words, the capacity to which the previous target color could guide attention was functionally nullified by the competing representation of the imagery instruction. The second is the emergence hypothesis – that once participants sufficiently implemented the imagery instruction, this instruction could intrude when participants were instructed to disengage from it. This hypothesis is akin to the “white bear phenomenon” – that an instruction to “not think of a white bear” can lead participants to think of it (Wegner, 1994; Wegner et al., 1987). Presently, it is not clear which of these two hypotheses best explain the present results. In favor of the interference hypothesis, the vividness rating results indicated that participants were unaware of the influence of the imagery instruction. However, it may be that the influence of the imagery instruction did not reach conscious awareness, or that the vividness rating did not adequately measure conscious disengagement from the imagery instruction. Indeed, we are also hesitant to come to strong conclusions regarding the vividness rating findings, given we have found somewhat contradictory results elsewhere (Cochrane, Ng, Khosla, & Milliken, 2021). In favor of the emergence hypothesis, when participants performed the word task of Experiment 3, a robust PoP effect emerged – that performing the word task sufficiently occupied cognitive resources, which meant they could not be allocated to performance of the imagery instruction. In all, it is our position that the emergence hypothesis is best supported by the present results, but further investigation of this issue is needed.
If the emergence hypothesis is true, it may be that the imagery vividness rating procedure aided the resiliency of the imagery instruction. In other words, while it was revealed that the imagery vividness ratings did not reflect the strength of imagery disengagement, it may be that the imagery vividness ratings reminded the participants of the imagery instruction, and thus, impaired imagery instruction disengagement. That is, it could be that the reason the PoP effect only approached significance in Experiment 2 for participants who performed the respond cue block after the imagery cue block was that the imagery vividness rating procedure was implemented during the respond cue block. While we suspect that the imagery vividness rating procedure does not fully explain the absence of the PoP effect during imagery instruction disengagement, the role of implicit reminders in instruction abandonment is a fruitful avenue for future research.
Another possibility is that instructing participants to not implement the imagery instruction reduced the PoP effect in and of itself. That is, the present work exploited the automaticity of the PoP effect to evaluate successful disengagement from the top-down imagery instruction. We then have interpreted the absence of the PoP effect as participants being unable to stop implementing the imagery instruction when instructed to do so. However, it is possible that participants were able to stop implementing the imagery instruction when instructed to do so, but that act of disengaging from the imagery instruction affected the processes driving the PoP effect. Hypothetically, what if to disengage from visual imagery a participant imagined a neutral color to that of imagery and the previous target, which prevented attentional guidance to both representations? While this interpretation is possible, such a finding would be remarkable considering research noting the resilience of the PoP effect (e.g., Cochrane & Pratt, 2020; Pascucci et al., 2012; Shurygina et al., 2019; Thomson et al., 2013). For example, we consistently observed robust PoP effects when encouraging participants to expect the opposite color and giving them a reason to do so by making target color switches more likely than repeats (Cochrane, Nwabuike, et al., 2018). In all, while it is possible that the disengagement instruction in and of itself eliminated the PoP effect, we suspect this was not the reason for the present findings.
The present findings further our understanding of the interactions of the top-down and selection history systems. Theeuwes (2019) defines selection history effects as “previous attentional deployments [that] elicit lingering and enduring selection biases that are unrelated to current goals nor related to the stimulus-driven salience of objects,” later indicating that “whenever attentional selection is driven by experiences with previous selection episodes one speaks about ‘history-driven selection’.” This description of history-driven selection fits well with the instruction-based effects here – that previously implemented instructions produce a lingering and enduring influence even when no longer actively implemented. That is, the present findings indicate an important interaction across the top-down and selection history systems. It appears that the top-down system can be used to establish an attentional control setting that can influence visual search while actively engaged, and once this attentional control setting is established, it can have an automatic influence on visual search. The benefit here is that the top-down system can work to deliberately intervene to aid performance, and once the behavior is established, the selection history system can take over to free up cognitive resources such that these associated behaviors may be efficiently executed.
Conclusion
The present study evaluated whether top-down instructions could have a lingering influence on visual search when they were to not be implemented. The results demonstrated that, by means of the PoP effect, participants were unable to disengage from the imagery instruction when instructed to do so. It was only when participants replaced the disengagement instruction with an extraneous top-down task that successful disengagement was possible, suggesting that automatic retrieval of prior implemented instructions can occur, but it is affected by other top-down tasks.
References
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16(8), 437–443.
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496.
Chun, M. M., & Jiang, Y. (1998). Contextual cueing: Implicit learning and memory of visual context guides spatial attention. Cognitive Psychology, 36(1), 28–71.
Clarke, A. D. F., Barr, C., & Hunt, A. R. (2016). The effect of visualization on visual search performance. Attention, Perception, & Psychophysics, 78, 2357–2362.
Cochrane, B. A., & Pratt, J. (2020). Re-examining Maljkovic and Nakayama (1994): Conscious expectancy does affect the Priming of Pop-out effect. Attention, Perception, & Psychophysics, 82(5), 2693–2702.
Cochrane, B. A., Nwabuike, A. A., Thomson, D. R., & Milliken, B. (2018). An imagery-induced reversal of intertrial priming in visual search. Journal of Experimental Psychology: Learning, Memory, and Cognition, 44(4), 572–587.
Cochrane, B. A., Zhu, H., & Milliken, B. (2018). Strategic visual imagery and automatic priming effects in pop-out visual search. Consciousness & Cognition, 65, 59–70.
Cochrane, B. A., Siddhpuria, S., & Milliken, B. (2019). Cueing color imagery: A critical analysis of imagery-perception congruency effects. Journal of Experimental Psychology: Learning, Memory, and Cognition, 45(8), 1410–1421.
Cochrane, B. A., Ng, W., & Milliken, B. (2021). Top-down imagery overrides the influence of selection history effects. Consciousness and Cognition, 93, 103153.
Cochrane, B. A., Ng, W., Khosla, A., & Milliken, B. (2021). Looking into the mind’s eye: Directed and evaluated imagery vividness modulates imagery-perception congruency effects. Psychonomic, Bulletin, & Review, 28(3), 862–869.
Cochrane, B. A., Townsend, B., Lee, R. C., Legere, J. K., Milliken, B., & Shedden, J. M. (2021). Visual imagery influences attentional guidance during visual search: Behavioral and electrophysiological evidence. Attention, Perception, & Psychophysics, 83(1), 58–66.
Cochrane, B. A., Wang, C., Pratt, J., Milliken, B., & Sun, H. J. (2021). Comparing imagery and perception: Using eye movements to dissociate mechanisms in search. Attention, Perception, & Psychophysics, 83(7), 2870–2890.
Cousineau, D. (2005). Confidence intervals in within-subject design: a simpler solution to Loftus and Masson’s method. Tutorials in Quantitative Methods for Psychology, 1(1), 42–45.
Fecteau, J. H. (2007). Priming of pop-out depends upon the current goals of observers. Journal of Vision, 7(6), 1–11.
Found, A., & Müller, H. J. (1996). Searching for unknown feature targets on more than one dimension: Investigating a “dimension-weighting” account. Perception & Psychophysics, 58(1), 88–101.
Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends in Cognitive Science, 8(11), 494–500.
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eye’s movement. In J. B. Long & A. D. Baddeley (Eds.), Attention and Performance IX (pp. 187–203). Erlbaum.
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72(1), 5–18.
Leber, A. B., & Egeth, H. E. (2006). It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review, 13(1), 132–138.
Leber, A. B., Kawahara, J., & Gabari, Y. (2009). Long-term, abstract learning of attentional set. Journal of Experimental Psychology: Human Perception and Performance, 35(5), 1385–1397.
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22(6), 657–672.
Maljkovic, V., & Nakayama, K. (1996). Priming of pop-out II. The role of position. Perception & Psychophysics, 58(7), 977–991.
Maljkovic, V., & Nakayama, K. (2000). Priming of popout: III. A short-term implicit memory system beneficial for rapid target selection. Visual Cognition, 7(5), 571–595.
Memelink, J., & Hommel, B. (2013). Intentional weighting: A basic principle in cognitive control. Psychological Research, 77, 249–259.
Morey, R. D. (2008). Confidence intervals from normalized data: a correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4(2), 61–64.
Moriya, J. (2018). Visual mental imagery influences attentional guidance in a visual-search task. Attention, Perception, & Psychophysics, 80(5), 1127–1142.
Pascucci, D., Mastropasqua, T., & Turatto, M. (2012). Permeability of priming of pop out to expectations. Journal of Vision, 12(10), 1–13.
Reinhart, R. M. G., McClenahan, L. J., & Woodman, G. F. (2015). Visualizing trumps vision in training attention. Psychological Science, 26(7), 114–122.
Shurygina, O., Kristjánsson, Á., Tudge, L., & Chetverikov, A. (2019). Expectations and perceptual priming in a visual search task: from eye movements and behavior. Journal of Experimental Psychology: Human Perception and Performance, 45(4), 489–499.
Theeuwes, J. (2019). Goal-driven, stimulus-driven, and history-driven selection. Current Opinions in Psychology, 29, 97–101.
Thomson, D. R., D’Ascenzo, M., & Milliken, B. (2013). Learning what to expect: context-specific control over intertrial priming effects in singleton search. Memory & Cognition, 41(4), 522–546.
Van der Stigchel, S., Belopolsky, A. V., Peters, J. C., Wijnen, J. G., Meeter, M., & Theeuwes, J. (2009). The limits of top-down control of visual attention. Acta Psychologica, 132(3), 201–212.
Van Selst, M., & Jolicoeur, P. (1994). A solution to the effect of sample size on outlier elimination. The Quarterly Journal of Experimental Psychology, 47A(3), 631–650.
Wang, B., & Theeuwes, J. (2018). Statistical regularities modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 44(1), 13–17.
Wegner, D. M. (1994). Ironic processes of mental control. Psychological Review, 101(1), 34–52.
Wegner, D. M., Schneider, D. J., Carter, S. R., & White, T. L. (1987). Paradoxical effects of thought suppression. Journal of Personality and Social Psychology, 53(1), 5–13.
Wolfe, J. M., Cave, K. R., & Franzel, S. L. (1989). Guided search: An alternative to the feature integration model of visual search. Journal of Experimental Psychology: Human Perception and Performance, 15(3), 419–433.
Wolfe, J. M., Butcher, S. J., Lee, C., & Hyle, M. (2003). Changing your mind: On the contributions of top-down and bottom-up guidance in visual search for feature singletons. Journal of Experimental Psychology: Human Perception and Performance, 29(2), 483–502.
Funding
Financial support for this study was provided in part by a Natural Sciences and Engineering Research Council of Canada Discovery Grant awarded to Bruce Milliken. The funding agreement ensured the authors’ independence in designing the study, interpreting the data, writing, and publishing the report. The authors report no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Open practices statement
The experiments reported in this article were not preregistered. All data have been made publicly available on the Center for Open Science website (osf.io/jh3fb). Requests for materials can be sent via email to the corresponding author.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cochrane, B.A., Pratt, J. & Milliken, B. Top-down then automatic: Instructions can continue to influence visual search when no longer actively implemented. Atten Percept Psychophys 85, 76–87 (2023). https://doi.org/10.3758/s13414-022-02558-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-022-02558-4