
Abstract
Familiar self-associated stimuli such as one’s own name and face are more efficient in guiding attention than other-associated stimuli. Remarkably, a short association of geometric shapes to the self versus others is sufficient to induce prioritization of the self- (vs. other-) associated shape in a matching task. Replications with other tasks measuring different stages of information processing, however, produced mixed results. It thus remains unclear whether the effect can be attributed to the newly associated stimulus alone. Therefore, in Study 1 (N = 28), we implemented the matching task and additionally compared the effectiveness of familiar versus newly self-associated stimuli with that of stranger-related stimuli to hold attention as cues in a dot-probe task. The self and the stranger were either represented by familiar labels (“I” vs. “stranger”), newly associated shapes, or shape–label pairs. In Study 2 (N = 31), participants associated nonwords to themselves and a stranger to compare the attentional impact of familiar and new self-associated letter combinations. Thus, we addressed the potential limitation due to modality present in former studies—which used mostly pictorial stimuli as new representations and letter combinations as familiar representations. Across both studies, in the dot-probe task, responses were faster towards targets following the self-associated stimuli compared with stranger-associated stimuli but only when familiar representations were used. Responses in the matching task were faster when confirming the correct self-associated pair. The results suggest that, under conditions of attentional competition, the prioritization of self-associated compared with other-associated cues does not generalize to newly associated stimuli.
Similar content being viewed by others


Introduction
At any given moment, our body is stimulated by huge amounts of input. Take, for example, a person sitting on a train who sees the scenery changing through the window, smells the perfume which fellow passengers have excessively sprayed on themselves, and senses the vibrations of the train as it runs along the tracks while listening to the train announcements. In order to manage so much information, our cognitive system relies on various mechanisms that allow it to filter, process, and respond to such stimulation. Self-relevance is one important characteristic that influences information processing (Moray, 1959; Wójcik et al., 2018; Yang et al., 2013), even when such relevance has only recently been established (Sui et al., 2012). Indeed, a growing body of research suggests that, recently, self-associated stimuli may benefit from prioritized cognitive processing (Janczyk et al., 2019; Mattan et al., 2015; Schäfer et al., 2016; Truong et al., 2017). However, there are limitations to such effects, and the evidence sustaining them is mixed (Caughey et al., 2021; Falbén et al., 2019; Stein et al., 2016); in particular, the impact of newly self-associated stimuli on attention is still unclear (Dalmaso et al., 2019; Siebold et al., 2015; Sui et al., 2009; Wade & Vickery, 2018; Zhao et al., 2015). Hence, the purpose of this research is to (1) demonstrate how self-association specifically impacts attention holding under conditions of competition between stimuli, and (2) compare attention holding of familiar versus newly self-associated stimuli separately. In addition, it aims at (3) comparing familiar and newly self-associated stimuli by using the same perception modus for both, thereby ruling out the potential limitations in its interpretation due to the modality of stimuli used in the classical setup of prior studies (Study 2).
Attentional prioritization of self-associated versus other-associated stimuli
Familiar self-associated stimuli have consistently been found to impact the distribution of attention (see Sui & Rotshtein, 2019). For example, one’s own name or a picture of one’s own face can capture attention more easily than names or faces of others (Alexopoulos et al., 2012; Arnell et al., 1999; Moray, 1959; Yang et al., 2013). Similarly, one’s name and face are harder to ignore than those of others (Wójcik et al., 2018; Wood & Cowan, 1995). Moreover, targets that follow self-associated stimuli (i.e., occurring at a location that had previously been occupied by a self-associated stimulus) elicit faster responses than targets that follow other-associated stimuli (Alexopoulos et al., 2012). This effect indicates that the attentional focus remains at the location of a self-associated stimulus even after the stimulus has disappeared (Wójcik et al., 2018), reflecting attention holding. With the use of directional stimuli such as faces oriented towards a specific location, responses are faster for targets presented at a location towards which one’s own face (but not another face) is oriented in comparison to targets presented at a distractor location (Liu et al., 2016), indicating that directional self-associated cues orient attention more efficiently than other-associated cues. In summary, familiar self-associated stimuli can efficiently guide attention as well as elicit attentional capture and attention holding with greater ease than other-associated stimuli.
Notably, a growing body of literature has also observed prioritization effects for stimuli that have only recently become self-associated (e.g., Schäfer et al., 2016; Sui et al., 2012; Truong et al., 2017). This means that effects of self-related stimuli like one’s name or face are not solely attributable to familiarity. The established method used to experimentally induce self-association consists of associating neutral stimuli—such as geometric shapes—to the self, a close other (e.g., mother or a friend), and a stranger or neutral object (e.g., a chair). Participants are then asked to complete a response time (RT) matching task comprising random pairs of a geometric shape and a word label (i.e., “I,” “mother,” “stranger”). They are instructed to indicate whether the presented combinations are correct or incorrect according to the initial association. Remarkably, responses are typically fastest when confirming the correct self-associated shape–label pair. The advantage in verifying the self-associated shape–label pair in comparison to any other-associated shape–label combination is called the self-prioritization effect (SPE; Sui et al., 2012). The SPE demonstrates that even newly self-associated stimuli induce prioritization in information processing, but the evidence leaves at least two points unresolved: First, it remains unclear which stage of information processing is affected, and second, the evidence does not allow one to fully conclude whether the newly associated stimulus alone induces prioritization.
Differentiating stages of information processing
The SPE has been interpreted to reflect (among other mechanisms) an attentional prioritization of the self-associated compared with other-associated shape–label pairs, taking into consideration that self-association integrates multiple cognitive processes (Humphreys & Sui, 2015; Sui et al., 2009; Sui & Rotshtein, 2019). However, some evidence also suggests that there may not be a shared mechanism of self-prioritization across different cognitive domains (Nijhof et al., 2020). This would mean that the SPE observed in the matching task may not generalize to other cognitive tasks. Thus, it is important to correlate results from the matching task with other established cognitive measures to compare whether newly self-associated stimuli impact information processing in the same way as familiar self-associated stimuli. Research combining the matching task with other established cognitive tasks has provided mixed evidence suggesting that the SPE may impact perception (Macrae et al., 2017; Sui et al., 2012), attention (Dalmaso et al., 2019; Humphreys & Sui, 2015; Macrae et al., 2018), and memory (Janczyk et al., 2019). While an impact on memory suggests that newly established self-associations consist of internal representations temporarily held in working memory, an effect occurring at an earlier stage of information processing (such as attention or perception) would suggest that a newly self-associated stimulus is immediately rendered more salient without conscious cognitive effort—as also observed with familiar self-associated stimuli (Bargh, 1982; Blume et al., 2017; Bola et al., 2020; Wood & Cowan, 1995). However, the matching task has not yet been combined with established paradigms that can be interpreted as attention holding. Hence, we combine the matching task with an established cuing paradigm—namely, the dot-probe task—to measure attention holding of self-associated stimuli.
Evidence from cuing paradigms: Sequential versus simultaneous presentation
In cognitive psychology, cuing paradigms represent a highly established tool to test the potential of specific stimuli to impact attention holding (Frischen et al., 2007; Hawkins et al., 1990; Posner, 1980; Verghese, 2001). Cuing tasks consist of the presentation of cues varying in salience, followed by a target that requires a response. The targets are presented either at the location that had previously been occupied or indicated by the cue (valid trials) or at a different location (invalid trials). Results typically show faster and more accurate responses on valid trials compared with invalid trials and, more importantly, the effect is enhanced by the salience of the cue (Ehrman et al., 2002; MacMahon et al., 2006). If the difference between valid and invalid trials is enhanced, the stimuli are interpreted to be salient for the respondents.
There is evidence from cuing paradigms that indicates the impact of self-association on attention holding, but most of the studies do not compare such an impact when newly established self-associated and other-associated stimuli are presented simultaneously. Studies that have combined the matching task with cuing paradigms found that newly established self-associated stimuli hold attention (Dalmaso et al., 2019; Siebold et al., 2015; Sui et al., 2009), but these studies have mostly used setups in which the self-associated and other-associated stimuli are presented sequentially. In visual search, the self-associated shape was detected faster than the other-associated shape (Wade & Vickery, 2018), but this effect was observed only when the self-associated or other-associated shape were presented amongst a set of unfamiliar stimuli. In contrast, latencies of visual search saccades towards self-associated versus stranger-associated geometric shapes did not differ significantly when the self- and stranger-associated shapes were presented simultaneously (Siebold et al., 2015). Notably, in the matching task, the SPE is typically observed in matching trials that present either self-associated pairs, or other-associated shape–label pairs sequentially, but self-associated versus other-associated stimuli never compete for attention. The SPE is usually not observed in nonmatching trials that present both self-associated and other-associated stimuli (i.e., the shape is associated to one, whereas the label is associated to the other) simultaneously (Sui et al., 2012). Since this setup strays from the reality of our environment in which various stimuli simultaneously compete for our attention, we tested whether the established finding that newly established self-relevance causes attentional prioritization actually holds in a context in which two socially salient stimuli—one self-relevant, one other-relevant—are presented simultaneously. The dot-probe task represents a classical cuing paradigm meeting this requirement because self-related and other-related information can be presented simultaneously. Hence, the current research investigated whether new versus familiar self-associations capture and hold attention under conditions of competition for attentional resources.
Disentangling the impact of new versus familiar self-associated stimuli on attention
As reported above, the matching task is interpreted as evidence that even newly self-associated stimuli guide attention. One might, however, criticize that the newly associated stimuli are never presented alone, but rather paired with a familiar label (e.g., “I” or “stranger”; Janczyk et al., 2019; Macrae et al., 2017; Reuther & Chakravarthi, 2017; Sui et al., 2012; Sui & Humphreys, 2015), so that the effect cannot clearly be attributed to the newly associated shape by itself.
To the best of our knowledge, there is one study which directly compared the impact of familiar and new self-associations. Woźniak and Hohwy (2020) used a categorization task in which they asked participants to associate their own name to a stranger’s face, their own face with a stranger’s name, and an additional (stranger’s) name to a (stranger’s) face. Participants were significantly faster at responding in trials which presented their own face paired with a stranger’s name but were slowest in trials which presented their own name paired with a stranger’s face, reflecting a bottom-up effect of self-association. However, this methodology still presents trials that include the combination of familiar and new self-representations.
Only few published studies have investigated whether the self-associated shape alone can elicit prioritization effects in attention. Woźniak and Knoblich (2019) had participants associate arbitrary faces and abstract symbols to the self and others. Afterwards, participants completed the matching task presenting random combinations of the faces and symbols. The typical SPE pattern was observed in participants’ performance in the matching task, providing preliminary evidence that the SPE is indeed found without the use of familiar labels. Nevertheless, the perception of face stimuli entails particularities in information processing that do not apply to other visual stimuli (Kanwisher et al., 1997; Kanwisher & Yovel, 2006; Perrett et al., 1992; Tsao et al., 2006; Yovel & Kanwisher, 2004). Thus, the generalization of this effect for other self-associated symbols awaits further research.
Cuing paradigms in particular have only been scarcely used with recently established self-associated stimuli. Sui et al. (2009) asked participants to associate themselves and a friend with distinctly coloured arrows (i.e., red vs. green). These arrows were used as cues that either pointed towards (valid) or away (invalid) from the location where the target would subsequently occur. The size of the cuing effect was larger for self-associated cues than for friend-associated cues, which was interpreted as a reflection of self-associated cues causing a faster shift of attention towards cued locations than friend-associated cues. As a limitation of this effect, when using auditory targets, the effect of self-associated versus other-associated arrows was only observed with voice targets but not tone targets. The same was true when instead of arrows, generic faces gazing to the right versus to the left were used as cues (Zhao et al., 2015). In regard to attention holding in particular, in an oculomotor task, participants initiated saccades away from self-associated geometric shapes more slowly than saccades away from stranger-associated geometric shapes—an observation interpreted as an increased difficulty to steer attention away from self-associated as opposed to stranger-associated shapes (Dalmaso et al., 2019). This suggests that—like highly familiar self-associated stimuli (Wójcik et al., 2018)—recently established self-associated stimuli may hold attention. However, Dalmaso et al. (2019) observed the effect only in one of two experiments (namely, only when the self/other distinction was task relevant). Furthermore, in a dot-probe location task which presented self-associated and stranger-associated familiar labels and new shapes, participants responded faster to targets cued by self-associated stimuli than targets cued by stranger-associated stimuli only when familiar labels were used—suggesting attentional prioritization of familiar self-associated stimuli but not of new self-associated stimuli (Study 1 in Orellana-Corrales et al., 2020). Yet the use of the dot-probe location task presents limitations in regard to the attribution of its results (see Imhoff et al., 2019). Taken together, it remains unclear whether recently established self-associated stimuli alone elicit attentional prioritization as effectively as highly familiar self-associated cues. Hence, the current research specifically compares the potential of recently established versus familiar self-associated stimuli versus a pair of them to capture and hold attention.
Hypotheses
Based on the evidence indicating an advantage for the information processing of self-associated labels compared with other-associated labels (Alexopoulos et al., 2012; Bargh, 1982; Yang et al., 2013) and the mixed evidence for the advantage of recently self-associated shapes (Dalmaso et al., 2019; Siebold et al., 2015; Sui et al., 2009; Wade & Vickery, 2018; Zhao et al., 2015), we hypothesized that the attentional benefit of the self-associated compared with the stranger-associated cues in the dot-probe task will be more pronounced when familiar stimuli are present (i.e., labels and pairs) compared with when the self and the stranger are represented by the new stimuli alone (i.e., shapes). Interestingly, in the matching task, the size of the SPE increases when two self-associated shapes are presented on matching self-associated shape–label trials compared with when one self-associated shape is presented (Sui & Humphreys, 2015). Similarly, the simultaneous presentation of both new and familiar self-associated stimuli (i.e., pairs) in the dot-probe task might lead to a greater processing advantage compared with the presentation of only the self-associated shape or only the self-associated label (see Lockhead, 1966, for the general concept of redundancy gains). We therefore expected a compound effect of self-association where two self-associated stimuli (regardless of whether these are familiar or newly associated) will impact attention more than one stimulus alone; that is, we expected prioritized responding towards self-associated stimuli to be enhanced when using paired representations, due to the increased number of self-associated elements (i.e., label and shape).
Study 1
Method
Participants
A minimum sample size was established through a priori power calculations using G*Power (Faul et al., 2007). Prior studies have reported the SPE as a medium to large effect in size (dz > 0.81 in Sui et al., 2012, and dz ≥ 0.58 in Schäfer et al., 2016), and a study using self-face and other-face as stimuli in a dot-probe task to measure attentional capture reported a large effect size for congruency between target location and self-association of stimuli (\( {\upeta}_{\mathrm{p}}^2=.19 \) in Wójcik et al., 2018). On this basis, we expected the effect of self-prioritization in the dot-probe task to be medium in size (f = .25; Cohen, 1988). For a repeated-measures multivariate analysis of variance (MANOVA; see O’Brien and Kaiser, 1985, for the use of MANOVA to analyze repeated-measure designs) of mean RTs with one group, six measurements (2 [target position: self vs. stranger] × 3 [type of representation: familiar vs. new vs. pair]), α = .05, correlation among the measures = .50, a minimum sample size of N = 28 is needed to detect an effect with a power effect of 1 − β = .90.
In order to allow for dropouts and exclusion of outlier responses, a total of 34 participants (25 female, Mage = 23 years, SDage = 3.5) completed the study. All participants had normal or corrected-to-normal vision and were able to complete the study in German. Before performing any data analyses, the distribution of RT and error data for the dot-probe task and matching task were individually observed in order to detect outliers. Data from six participants were excluded due to the average of their RTs and error rate falling three interquartile ranges above the third quartile of the sample distribution of all participants in either one of the tasks (Tukey, 1977). The study was carried out according to the principles of the Declaration of Helsinki, on the basis of informed consent.
Design
The data from the dot-probe task consisted of a 2 (target location: self vs. stranger) × 3 (type of representation: familiar vs. new vs. pair) within-participants design. The assignment of shapes as new representations of the self and stranger was randomized and counterbalanced throughout participants, and the target position was randomized and counterbalanced throughout trials. The data from the matching task consisted of a 2 (shape: self-associated vs. stranger-associated) × 2 (trial: matching vs. nonmatching) within-participants design.
Apparatus and materials
The experiment was run on E-Prime 2.0 (Schneider et al., 2002), using Acer Aspire E15 35-573G-54SK 15.6-in. laptops and standard computer mice.
All stimuli were presented at a viewing distance of 50 cm, in white colour and against a black background. Verbal stimuli were presented in Courier New font, 18-pt size type. The geometric shapes were presented at a visual angle of approximately 5° × 5°, and verbal stimuli were presented at a visual angle of about 0.7°.
Procedure
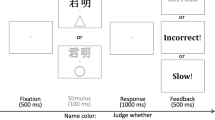
All specific instructions were presented on the computer screen in German language. First, the participants were instructed to associate geometric shapes (square and triangle) to the labels “I” and “stranger” (“Ich” and “Fremder” in German) with the instruction: “You are a [Shape 1] and a stranger will be represented by a [Shape 2].” During this association phase, no images of the shapes were presented. After familiarizing themselves with the instructions, the participants pressed any key to continue with the experiment. Next, the participants completed the dot-probe task (see Fig. 1). They first completed 24 practice trials in which they received feedback about whether their response was incorrect or exceeded 1,500 ms (“incorrect”’ “please respond faster”). Afterwards, they completed 240 experimental trials in which they did not receive feedback. Each trial began with a fixation cross in the centre of the screen (500 ms), followed by the stimuli representing the self and a stranger on opposite sides of the screen (left and right, located on 25% and 75% of the horizontal line of the screen and on 50% of the vertical line of the screen, 200 ms). Representations were a label, a geometric shape, or a matching shape–label pair, with the order of presentation being randomized. Then, a target consisting of either a “q” or “p” was presented on either the left or right side of the screen (on 25% or 75% of the horizontal line of the screen, and on 50% of the vertical line of the screen) until participants provided a discrimination response by pressing “q” or “p” to indicate which target was presented. The target (q or p) and the location of the target (left or right) were randomized between trials. A 1,000 ms pause which consisted of a black screen preceded the next trial.

Schematic depiction of one trial of the dot-probe task (a) and example displays demonstrating how the display differed as a function of the type of representation in Study 1 (familiar vs. new vs. pair; b) and Study 2 (familiar vs. new; c)
As an additional exploratory measure, the participants also completed the dot-probe task following the same procedure but requiring a detection response rather than a discrimination response. The order of the tasks (discrimination and detection) was randomized and counterbalanced across participants. The detection task was included purely for exploratory purposes for future studies, and its trials were not included in the current analysis.Footnote 1
Finally, the matching task was presented in order to replicate the basic SPE finding and compare its results to those of the dot-probe task. The matching task was always presented last for all participants. Each trial began with a black screen (500 ms) followed by a fixation cross (500 ms). A paired representation consisting of a shape and a label underneath was then presented and remained on screen until the participant responded or for a maximum of 1,500 ms. There were two possible responses: The participants had to press “d” to indicate that the shape–label pair matched the association learned during the association phase and “k” to indicate that it did not match the learned combination. They received feedback if their response was incorrect or exceeded 1,500 ms (“incorrect”; “please respond faster”). Initially, four trials were administered as a practice phase, followed by 128 trials of the matching task.
The participants were then thanked, debriefed, and compensated with 6 euros. Students of the Department of Psychology at the University of Tübingen could opt to receive class credit.
Results
The collected data and analyses scripts are available on the Open Science Framework (OSF; https://osf.io/g7wrc/). For all statistical analyses, a significance level of α = .05 was specified. Bonferroni corrections were applied to adjust for multiple comparisons. For RT analyses, only correct responses with RTs above 100 ms and below three interquartile ranges above the third quartile of the overall individual RT distribution were used (Tukey, 1977). Exclusions of trials were performed separately for the dot-probe task and the matching task.
Dot-probe task
Average RTs
Average RTs in the dot-probe task (see Fig. 2) were subjected to a 2 (target location: self vs. stranger) × 3 (type of representation: familiar vs. new vs. pair) within-participants MANOVA. Responses were generally faster when the target was presented at the location previously occupied by the self-representation (M = 505 ms, SD = 35 ms) compared with the stranger-representation (M = 522 ms, SD = 35 ms), F(1, 27) = 89.10, p < .001, ηp2 = .77. Furthermore, a significant main effect of type of representation was observed, F(2, 54) = 61.31, p < .001, ηp2 =.69. Follow-up analyses revealed that, irrespective of the target location, mean RTs were significantly slower for targets following familiar representations (labels; M = 532 ms, SD = 36 ms) than for targets following new representations (shapes; M = 511 ms, SD = 37 ms), t(27) = 5.80, p <.001, dz = 1.10; further, mean RTs were significantly slower for targets following new representations than targets following paired representations (shape–label pairs; M = 499 ms, SD = 35 ms), t(27) = 5.34, p <.001, dz = 1.00. The expected interaction of Target Location × Type of Representation was also significant, F(2, 54) = 12.55, p < .001, ηp2 = .32. Follow-up analyses revealed that responses were significantly faster for targets following the self-representation than stranger-representation, reflecting a significant cuing effect both when the instances were represented by labels (M = 514 ms, SD = 34 ms for self; M = 549 ms, SD = 42 ms for stranger), t(27) = 7.58, p < .001, dz = 1.43, and by pairs (M = 492 ms, SD = 40 ms for self; M = 507 ms, SD = 33 ms for stranger), t(27) = 3.51, p = .002, dz = 0.66. However, the size of the cuing effect yielded by labels was significantly larger than the size of the cuing effect yielded by pairs, t(27) = 2.85, p = .039, dz = 0.54; that is, the difference in response time favouring responses to targets that followed self-representations over stranger-representations was greater when the self and stranger were represented by labels than when they were represented by pairs. Most importantly, however, no cuing effect was observed for the new representation condition, t(27) = 0.81, p = .423, dz = 0.15, BF01= 4.6; that is, responses for targets following the self-representation (M = 510 ms, SD = 38 ms) and responses for targets following the stranger-representation (M = 513 ms, SD = 37 ms) did not differ significantly when the self and stranger were represented by newly associated shapes alone. A Bayesian analysis using JASP (JASP Team, 2020) yielded a moderate level of evidence in support of the null hypothesis that responses did not differ between targets that followed self- and stranger-representations represented by shapes alone.

Mean response times in (a) the dot-probe task as a function of target location (self vs. stranger) and type of representation (familiar vs. new vs. pair) and (b) the matching task as a function of shape (self-associated vs. stranger-associated) and trial type (matching vs. nonmatching) in Study 1. Error bars represent standard errors. ***p < .001
Error rates
Mean error rates (see Table 1) were submitted to a 2 (target location: self vs. stranger) × 3 (type of representation: familiar vs. new vs. pair) MANOVA. Responses to the different types of representations differed significantly in accuracy (labels: M = 3% error rate, SD = 2, shapes: M = 2% error rate, SD = 1, pairs: M = 2% error rate, SD = 2), F(2, 54) = 5.94, p = .005, ηp2 = .18. The main effect of target location was not significant, F(1, 27) = 0.03, p =.875, ηp2 = .001. The interaction of Target Location × Type of Representation, F(2, 54) = 1.66, p = .201, ηp2 = .06, was also nonsignificant.
Matching task
In order to measure the SPE and compare it to the results in the dot-probe task, we analyzed performance in the matching task.
Average RTs
The RT data (see Fig. 2) were subjected to a 2 (shape: self-associated vs. stranger-associated) × 2 (trial type: matching vs. nonmatching) within-participants MANOVA. The main effects of shape, F(1, 27) = 99.33, p < .001, ηp2 = .79, and trial type, F(1, 27) = 37.66, p < .001, ηp2 = .58, were both significant. The interaction of shape and trial type, F(1, 27) = 51.48, p < .001, ηp2 = .66, was also significant. To follow up on this interaction effect, a t test was run to analyze RTs from matching trials. The analysis revealed a significant difference between RTs in responses towards self-associated and stranger-associated shapes in matching pairs, t(27) = 10.88, p < .001, dz = 2.07, indicating a significant SPE in the RT data; that is, responses were faster for matching self-associated shape–label pairs (M = 683 ms, SD = 129 ms) than for matching stranger-associated shape–label pairs (M = 931 ms, SD = 202 ms). The RTs from nonmatching trials were submitted to the same analysis, which also revealed a significant difference in RTs to self-associated and stranger-associated shapes in nonmatching pairs, t(27) = 2.41, p = .023, dz = 0.46. More precisely, responses were faster for trials presenting the self-associated shape with the label “stranger” (M = 855 ms, SD = 135 ms) than trials presenting the stranger-associated shape with the label “self” (M = 898 ms, SD = 156 ms).
Sensitivity measure d′
Signal detection sensitivity indices (d′) for each shape condition were used to analyze mean error rates (see Table 1; Schäfer et al., 2015, 2016 ; Sui et al., 2012). To this end, responses were defined as follows: For matching trials, correct responses were considered hits and incorrect responses were considered misses; for nonmatching trials, correct responses were considered correct rejections, and incorrect responses were considered false alarms. In order to account for cases with 100% hits or 0% false alarms, the loglinear approach was used; that is, before calculating the rates for hits and false alarms, 0.5 was added to the number of hits and the number of false alarms, and 1 was added to the number of signal trials and the number of noise trials (Hautus, 1995; Stanislaw & Todorov, 1999). Resulting sensitivity measures were submitted to a one-factorial (shape: self-associated vs. stranger-associated) MANOVA. A significant main effect of shape was observed, F(1, 27) = 35.37, p < .001, ηp2 = .57, indicating a higher sensitivity for self- than for stranger-associated shapes (i.e., a significant SPE in d′).
Comparing measures of SPE and cuing effect
A correlation analysis was run in order to compare the results observed in the dot-probe task regarding the cuing effect of familiar, new, and pair representations of self and stranger with the SPE. We did not observe a significant correlation between RT measures of the SPE and the cuing effect of self-representations, regardless of whether these were familiar, r = .06, p = .766, BF01= 3.1, new, r = −.16, p = .412, BF01= 3.2, or pair representations, r = − .04, p = .853, BF01= 1.5. A Bayesian analysis yielded a moderate level of evidence in support of the null hypothesis that RT measures of the SPE and the cuing effect for familiar and new representations did not correlate. The analysis yielded only anecdotal evidence in support of the null hypothesis that RT measures of the SPE and the cuing effect for pair representations did not correlate. We did not observe a significant correlation between d′ measures of the SPE and the cuing effect of familiar, r = −.10, p = .622, BF01= 3.8, new, r = −.15, p = .459, BF01= 4.4, or pair representations, r = .13, p = .517, BF01= 4.3. Bayesian analysis yielded a moderate level of evidence in support of the null hypothesis that d′ measures of the SPE and the cuing effect did not correlate.
Discussion
Study 1 investigated how self-relevance impacts the distribution of attention. In detail, we compared (1) self-associated versus stranger-associated cues and (2) familiar versus newly established self-versus stranger-associated cues versus pairs. We assessed the cues’ efficiency in enhancing the identification of a subsequent probe target (attention holding) under conditions of competition between stimuli.
In the matching task, the participants were faster and more accurate when responding to the matching self-associated shape–label combination than when responding to any other combination. These results replicate the SPE as described in the literature (Schäfer et al., 2016; Sui et al., 2012); confirming that our manipulation to induce self-association of a geometric shape was successful.
In the dot-probe task, as expected, the participants were generally faster and more accurate in identifying the probe target when it occurred at the location that had previously been occupied by self-associated compared with the stranger-associated stimuli. This indicates that self-associated stimuli captured attention and that attention remained at the respective location. We interpret this to reflect that self-association can elicit attention holding.
Importantly, the effect of self-association only occurred for the familiar but not for the newly established cues. Specifically, significant cuing effects were observed when the self and stranger were represented by the corresponding labels or by shape–label pairs (both containing familiar representations), but not when represented by the corresponding shapes (new representations) alone, either in RTs or in error rates. This is in line with our hypothesis that familiar self-associated versus other-associated cues are more efficient in eliciting attentional capture and attention holding than newly established cues.
Contrary to what was expected, the shape–label pairs were no more efficient in eliciting cuing effects than labels only. Rather, labels alone elicited a larger cuing effect than shape–label pairs. Hence, adding the newly self-associated shape to the familiar self-associated label did not increase differences between self-and other-related stimuli. This is not in line with the reasoning that redundancy gains (Lockhead, 1966), induced by the presentation of the shape–label pair compared with only the shape or label alone, would enhance the effect of self-relevance in the dot-probe task (see Sui & Humphreys, 2015). Given that we did not observe a cuing effect for the shape alone, it is reasonable that we did not observe redundancy gains when pairing the label with the shape. Still, responses were generally faster for targets following pairs as compared with targets following shapes or labels only, which demonstrates that there is a facilitated processing of information if two cues are presented instead of one, independently of the self- versus other- association.
Taken together, the findings in Study 1 demonstrate that the advantage of self-associated stimuli to capture and hold attention cannot necessarily be induced by associating new stimuli to the self shortly before. One might criticize, however, that word-labels were used as familiar representations and pictorial geometric shapes were used as newly established representations in former studies (e.g., Dalmaso et al., 2019; Janczyk et al., 2019; Schäfer et al., 2016; Sui et al., 2012), as well as in Study 1. Thus, it is not possible to deduce from this data whether the effects are due solely to the differences in familiarity or whether they are somehow influenced by the difference in modalities of representation. Pictorial stimuli and written words are intrinsically different: These two types of stimuli are processed at different speeds (Shor, 1971) and differ in their cognitive impact even when used to represent the same concept (see Jenkins et al., 1967; Sperber et al., 1979). Thus, it can be criticized that the evidence available may only be generalized to a limited extent. Thus far, this limitation has only been approached by using participants’ faces as familiar representations and arbitrary, newly self-associated faces to measure the impact of self-association in the matching task, which yielded limited effects of enhancement in responses towards self-associated stimuli (Woźniak et al., 2018; Woźniak & Hohwy, 2020). However, as previously mentioned, face stimuli are processed in particular ways that differ from that of other visual stimuli (Kanwisher et al., 1997; Kanwisher & Yovel, 2006; Perrett et al., 1992; Tsao et al., 2006; Yovel & Kanwisher, 2004). Word stimuli, in contrast, can easily allow for the manipulation of familiarity of self-representations.
In Study 2, we therefore address the use of different modalities to represent familiar and newly self-associated stimuli by comparing familiar and newly self-associated letter combinations (i.e., words vs. nonwords) with each other. Again, we expect an attentional benefit towards self-associated letter combinations in comparison to stranger-associated letter combinations when such stimuli are familiar (i.e., for words). Based on the findings of Study 1, however, we do not expect to find this self-associated versus other-associated attentional benefit when newly associated stimuli (i.e., for nonwords) are presented alone.
Study 2
We aimed to test whether the effects observed in Study 1 replicate when both familiar and new representations are represented by stimuli of the same modality. To this end, we compared familiar and new self-associated letter combinations (i.e., words vs. nonwords). In line with Study 1, we expected to observe an attentional benefit towards self-associated letter combinations in comparison to stranger-associated letter combinations only when such stimuli are familiar (i.e., for words) but not when stimuli are new (i.e., for nonwords).
Method
Study 2 was carried out according to the principles of the Declaration of Helsinki, on the basis of informed consent and preregistered in the OSF (https://osf.io/s68pm).
Participants
The minimum require sample size was established through a priori power calculations which were performed in G*Power (Faul et al., 2007). In Study 1, we observed a large effect size for the interaction between target location and type of representation (ƒ = .72) and a high correlation among measures (between r = .77 and r = .92). For a repeated-measures MANOVA with one group, 4 measurements (2 [target location: self vs. stranger] × 2 [type of representation: familiar vs. new]), α = .05, correlation among measures = .70, nonsphericity correction ε = 1, and a conservatively chosen medium effect size of ƒ = .25 for the expected interaction effect, a minimum sample size of N = 25 is needed to detect an effect with a power of 1 − β = .95.
The study was completed by 34 participants (23 female; Mage = 21 years, SDage = 2.8). Before analyzing the data, each participant’s mean RT and error rate were compared with the sample distribution, separately for the dot-probe task and the matching task, in order to detect outliers in the data. The average RT of two participants were observed within three interquartile ranges above the third quartile of the sample distribution of all participants (Tukey, 1977); their data were therefore excluded from all analyses.
Apparatus and materials
The experiment was conducted using the same apparatus and materials described in Study 1, with the exception of the use of nonword stimuli as new self-associated and stranger-associated stimuli (instead of geometric shapes). The nonwords used were letter combinations that phonologically resemble real German words, but have no semantic meaning. Based on materials used by Landkammer et al. (2019), they consisted of the following: sfartku, ambelde, teirnen, kes, muf, lor. Considering the difference in character length of the German words that were used (three characters for “Ich” and seven characters for “Fremder”), we also used nonwords consisting of three and seven characters. The assignment of nonwords to the self and stranger was randomized and counterbalanced across participants. As the character length of the nonwords might affect reaction times, the length of the nonwords that were associated to the self and stranger was also randomized and counterbalanced across participants: One half of the participants were assigned either a short nonword to the self and a long nonword to the stranger (thus mirroring the lengths of the familiar words in German), whereas the second half of the sample received long nonwords related to the self and short nonwords related to the stranger.
Procedure
At the beginning of the experiment, the participants were asked to associate one nonword to the concept “I” and one nonword to the concept “stranger.” To this end, the randomly assigned nonword was presented above the word to which it should be associated (i.e., “self” or “stranger”). Each combination was presented four times for a duration of 3,000 ms, alternating between the self-associated and stranger-associated pair. In order to mimic the way in which the association instructions are often worded in the SPE literature (e.g., Schäfer et al., 2015; Sui et al., 2012), the self-associated pair was always presented first.
The experiment then proceeded with a discrimination dot-probe task as described in Study 1, followed by the SPE matching task as also described in Study 1. In both tasks, nonwords were used instead of geometric shapes.
Design
Data regarding attentional prioritization in the dot-probe task consisted of a 2 (target location: self vs. stranger) × 2 (type of representation: familiar vs. new) within-participants design. The target location was randomized and counter-balanced throughout trials. Data regarding the SPE matching task consisted of a 2 (association: self vs. stranger) × 2 (trial type: matching vs. nonmatching) within-participants design.
Results
The collected data and analyses scripts are available on the OSF (https://osf.io/4cwrv/).
Dot-probe task
Average RTs
Average RTs in the dot-probe task (Fig. 3) were submitted to a 2 (target location: self vs. stranger) × 2 (type of representation: familiar vs. new) within-participants MANOVA. Responses were faster when the target was presented at the location previously occupied by a self-associated stimulus (M = 542 ms, SD = 63 ms) than when the target was presented at the location previously occupied by a stranger-associated stimulus (M = 557 ms, SD = 65 ms), F(1, 30) = 17.49, p < .001, ηp2 = .37. The effect of type of representation was not significant, F(1, 30) = 0.12, p = .731, ηp2 = .004. However, a significant interaction between target location and type of representation was observed, F(1, 30) = 8.75, p = .006, ηp2 = .22. Follow-up t tests revealed that, when the self and stranger were represented by familiar representations (words), responses were significantly faster for targets following the self-associated letter-strings (M = 537 ms, SD = 63 ms) than for targets following the stranger-associated letter-strings (M = 563 ms, SD = 67 ms), t(30) = 6.23, p < .001, dz = 1.12. As expected, no such cuing effect was observed for the new representation (nonword) condition, t(30) = 0.36, p = .720, dz = 0.06, BF01= 5.7. The Bayesian analysis yielded a moderate level of evidence supporting the null hypothesis that responses did not differ between targets that followed self- and stranger-representations represented by shapes alone.

Mean response times in (a) the dot-probe task as a function of target location (self vs. stranger) and type of representation (familiar vs. new) and (b) the matching task as a function of shape (self-associated vs. stranger-associated) and trial type (matching vs. nonmatching) in Study 2. Error bars represent standard errors. ***p < .001
Error rates
Mean error rates (see Table 1) were submitted to a 2 (target location: self vs. stranger) × 2 (type of representation: new vs. familiar) MANOVA. Neither the effects of target location, F(1, 30) = 1.01, p = .324, ηp2 = .03, nor type of representation, F(1, 30) = 0.23, p = .634, ηp2 = .01, nor the interaction effect, F(1, 30) = 0.32, p = .579, ηp2 = .01, were significant.
Matching task
Average RTs
Mean RTs (see Fig. 3) were subjected to a 2 (association: self-associated vs. stranger-associated) × 2 (trial type: matching vs. nonmatching) within-participants MANOVA. A main effect of association was observed, F(1, 30) = 26.65, p < .001, ηp2 = .47, while the effect of trial type was nonsignificant, F(1, 30) = 0.53, p = .473, ηp2 = .02. However, a significant interaction between association and trial type was observed, F(1, 30) = 15.42, p < .001, ηp2 = .34. The follow-up t test on matching trials revealed a significant SPE, t(30) = 5.81, p < .001, dz = 1.05; that is, the responses were faster for matching self-associated combinations (M = 615 ms, SD = 101 ms) than for matching stranger-associated combinations (M = 701 ms, SD = 149 ms). Submitting RTs from nonmatching trials to the same analysis did not reveal an effect, t(30) = 1.05, p = .301, dz = .19; that is, the responses to the combination of the self-associated nonword and the label “stranger” (M = 658 ms, SD = 117 ms) and the responses to the combination of the stranger-associated nonword and the word “self” (M = 670 ms, SD = 129 ms) did not differ significantly.
Sensitivity measure d′
Mean error rates are presented in Table 2. Measures of d′ were calculated as described in Study 1 and used to analyze error rates. The sensitivity measures were submitted to a one-factorial (association: self-associated vs. stranger-associated) MANOVA. A significant main effect of association was observed, F(1, 30) = 20.27, p < .001, ηp2 = .40, indicating a significant SPE in d′.
Comparing measures of SPE and cuing effect
Correlation analyses indicated no significant correlation between the SPE in RTs and the cuing effect for familiar, r = −.16, p = .391, BF01= 3.1, or new representations, r = .04, p = .840, BF01= 4.6. A Bayesian analysis yielded a moderate level of evidence in support of the null hypothesis that RT measures of the SPE and the cuing effect did not correlate. Measures of d′ for the SPE did not correlate with measures of the cuing effect for familiar, r = .25, p = .185, BF01= 2.7, or new representations, r = .06, p = .749, BF01= 4.0, either. A Bayesian analysis yielded a moderate level of evidence in support of the null hypothesis that d′ measures of the SPE and the cuing effect for new representations did not correlate but only anecdotal evidence regarding the lack of correlation between d′ measures of the SPE and familiar representations.
Discussion
Study 2 aimed to replicate the findings from Study 1 but used letter combinations for both familiar and new self-associated stimuli in order to rule out the potential influence of modality in the material.
As in Study 1, in the matching task, the participants were faster and more accurate at confirming the matching self-associated combination in comparison to any other combination. This reflects a significant SPE and demonstrates that nonwords can become associated to the self.
As expected, in the dot-probe task, we observed that the participants were faster at identifying the target when it was presented on the location previously occupied by the self-associated compared with the stranger-associated word. This indicates that, under conditions of attentional competition (i.e., when stimuli are presented simultaneously), self-associated versus stranger-associated letter combinations are prioritized in attention. However, no such difference was observed when nonwords were used: Self-associated letter combinations did not produce an advantage in identifying targets in comparison to stranger-associated letter combinations when such letter combinations had been newly associated to the self and stranger.
Taken together, Study 2 replicates the pattern of Study 1 and confirms that if newly self-associated stimuli stand alone, they do not have any advantage in capturing or holding attention when several stimuli compete for one’s attention.
General discussion
The present research set out to test whether new and familiar self-associated stimuli benefit from advantages in attention holding compared with other-associated stimuli under conditions of competition for attentional resources. Study 1 used familiar labels and newly self-associated shapes, whereas Study 2 used familiar versus newly self-associated letter combinations. Other than in the matching task (Sui et al., 2012), familiar and newly self-associated stimuli were also deliberately presented alone in order to disentangle the effects. In both studies, it was found that the familiar self-related stimuli (i.e., labels and, in Study 1, pairs) guide attention on a dot-probe task when competing for attention with other-related stimuli, whereas newly associated stimuli to the self (i.e., shapes in Study 1, letter combinations in Study 2) do not produce such an advantage in attention holding.
The finding that familiar self-representations guide attention is in line with former research: Responses towards one’s own name or face are faster than responses towards stimuli strongly linked to other persons, such as other’s names or faces (Alexopoulos et al., 2012; Arnell et al., 1999; Bargh, 1982; Brédart et al., 2006; Moray, 1959; Wójcik et al., 2018; Yang et al., 2013). The current results, however, question the conclusion that newly established self-representations produce a similar advantage in attention holding. Based on evidence obtained in the matching task that was introduced by Sui et al. (2012), it was interpreted that self-association can guide attention even when it has only recently been established. The matching task can, however, be criticized for presenting pairs of familiar and newly self-associated stimuli. Strictly speaking, the matching task does not test the impact of newly self-associated stimuli. The present study contributes to a better understanding of the impact of newly self-associated stimuli alone on attention by combining the classical matching task with a dot-probe task.
Former research that has used newly self-associated stimuli alone to test their impact on information processing has produced mixed evidence. Sui et al. (2009) demonstrated that in a cuing task, newly self-associated arrows can serve to orient attention—but this effect seems to hold only under very specific conditions (see Zhao et al., 2015). As to attention holding, Dalmaso et al. (2019) observed that saccades away from self-associated geometric shapes are initiated more slowly than saccades away from stranger-associated shapes when the self/other distinction was task-relevant. Similarly, Wade and Vickery (2018) observed faster detection of self-associated versus stranger-associated geometric shapes in complex visual search tasks. However, Siebold et al. (2015) did not observe faster detection of self-associated compared with stranger-associated shapes. The present data, likewise, did not confirm any attentional advantage of newly self-associated stimuli compared with other-associated stimuli.
Considering this mixed evidence, why do we sometimes find evidence for the prioritization of newly self-associated stimuli and other times not? Interestingly, the cuing tasks used by Sui et al. (2009) and the oculomotor task used by Dalmaso et al. (2019) present the self-associated and other-associated cue sequentially in different trials. Likewise, Wade and Vickery (2018) displayed stimuli including neutral and either self-associated or stranger-associated shapes. In contrast, in the study of Siebold et al. (2015), self-associated and stranger-associated cues are simultaneously presented in the dot-probe task on the same visual search display; that is, the latter study and our own studies, more specifically, the studies that did not support any attentional prioritization of newly self-associated stimuli over other-associated stimuli, test the potential of self-relevance to guide attention in contexts in which self-associated and stranger-associated stimuli have to be processed simultaneously and compete for attentional resources. The evidence under this condition suggest that attentional self-prioritization of newly associated stimuli may not transfer to an attention competition task. All in all, these findings suggest that the attentional impact of newly associated self-associated versus stranger-associated stimuli may depend on the visual context in which it is presented. Future research will have to systematically test the impact of different visual presentations as a moderating factor of the effect.
It could be criticized that the task order (i.e., presenting the matching task always at the end) may have affected the participants’ performance in the dot-probe task. More specifically, some studies present the matching task before any other cognitive task through which the associations between the shapes and the self and other(s) can (arguably) be strengthened (Dalmaso et al., 2019; Siebold et al., 2015; Zhao et al., 2015). In this case, it could be argued that the reason we did not observe a benefit for new self-representations in comparison to stranger-representations in the dot-probe task is due to the dot-probe task being presented before the matching-task, whereas an initial presentation of the matching task might have reinforced the self-association. However, it is generally considered that the SPE is an effect which is produced by the initial instructions that we used as a manipulation (Schäfer et al., 2015; Sui et al., 2012; Sui & Rotshtein, 2019), and measured with the matching task. Furthermore, Orellana-Corrales et al. (2020) tested the impact of self-associated and other-association of geometric shapes in a dot-probe location both when presenting the matching task before and after the dot-probe task and observed no impact of task order on the results.
The present findings contribute to the body of evidence that replicates the SPE in the matching task: Pairs of new and familiar self-stimuli are prioritized in comparison with pairs of new and familiar other-stimuli. The matching task has, however, been criticized for not allowing for conclusions about the stages of information processing that is affected by self-relevance (e.g., Janczyk et al., 2019; Schäfer et al., 2015). The present data also adds to literature suggesting that the SPE does not correlate with measures of self-prioritization in attention (Nijhof et al., 2020). The present data is the first to combine the matching task with a dot-probe discrimination task, thus using a method that allows for a more specific understanding of what stage of information processing is affected by self-association. Our study extends the body of literature showing that self-relevance elicits attentional capture (Arnell et al., 1999; Wade & Vickery, 2018) and serves to orient attention (Sui et al., 2009; Zhao et al., 2015) by demonstrating further support for the assumption that it also enhances attention holding (see Alexopoulos et al., 2012; Dalmaso et al., 2019; Wójcik et al., 2018). Furthermore, it shows that such attentional effects are limited by the familiarity of self-representations and are not elicited by newly self-associated stimuli. Future research should address these different aspects of attention with material that tests the impact of familiar and newly self-associated stimuli separately (as in the present studies), to gain a better understanding of the stages of information processing that are affected by self-relevance. Direct comparisons of these effects should also be compared with further understand how such effects of self-relevance relate to each other.
The present findings furthermore demonstrate that the lack of evidence for newly self-associated stimuli cannot be attributed to the modality of the stimuli: We still did not observe an impact of newly self-associated stimuli on attention when using the same stimulus modality as both familiar and new representations of the self and stranger in Study 2. Thus, we addressed a potential limitation in the body of evidence: The majority of the studies implemented words as familiar stimuli while using pictorial stimuli as newly associated stimuli. To the best of our knowledge, the only evidence so far that compared stimuli of the same modality has used two newly self-associated stimuli but not familiar stimuli. Woźniak and Knoblich (2019) had participants associate themselves and others with two new stimuli: generic faces and shapes. In their data, the classic SPE pattern was found in the matching task that combined a face and a shape. Even though faces and shapes are both pictorial material, this finding may be influenced by the particularities of the perceptual processing of faces (Kanwisher & Yovel, 2006; Tsao et al., 2006). At the very least, as previously mentioned, the use of face stimuli poses a limitation in regard to the generalization of the observed effects. When the stimuli are more comparable, as in Study 2, the effect might no longer hold. Future research will have to test a greater variety of modalities to understand whether the kind of modality has an impact on the overall self-prioritization effect of newly associated stimuli. In doing so, future research should also aim at varying the factors familiarity (familiar vs. newly associated) and type of modality (e.g., pictorial vs. letter-based) in one design and thus test the impact of these factors on self-prioritization systematically in the same design.
One might criticize that the length of the letter combination may influence the results: For familiar letter combinations, the word was always shorter for the self than for the stranger because the familiar words are given by natural language. For one half of the sample, the nonwords were also shorter for the self than for the stranger. As the familiar stimuli (i.e., the natural words “Ich” and “Fremder”) cannot randomly be allocated to long versus short letter combinations (i.e., we cannot use long familiar self-associated and short stranger-associated labels), the factor of word length cannot be analyzed as an independent factor in the analyses. However, if short words produce fast RTs and word length does not vary independently, then word length might be an alternative explanation for the present result pattern. In fact, most former evidence on the SPE that used either German or English labels might be questioned as influenced by word length, too, as in both languages the self-related familiar word is shorter than the other-related word. However, there is evidence that the SPE result pattern can be observed even when using the longer label “yourself” in comparison to “friend” and “stranger” to control for word length (Study 3C in Sui et al., 2012). Furthermore, research that used the participants’ names (thus having any lengths of letter combinations for the self-associated familiar stimulus) has consistently observed an attentional impact of self-association regardless of word length in attentional tasks (e.g., Alexopoulos et al., 2012; Nijhof et al., 2020; Tacikowski & Nowicka, 2010; Yang et al., 2013) and, most recently, also in the matching task (producing the usual SPE pattern; Nijhof et al., 2020). Thus, although in the current data an effect of word length cannot be ruled out as contributing to the effects, in light of the former evidence, it is likely that word length does not fully account for the findings. Still, future research should vary and test the influence of word length in the observed effects independently by using, for instance, names instead of the terms “I” and “stranger” or by comparing different languages.
In conclusion, our results indicate that familiarity (but not the stimulus modality) determines the attentional impact of self-associated versus stranger-associated stimuli; that is, familiar self-associated stimuli robustly elicit attention holding, whereas recently established self-associations may not be sufficient to induce such an attentional prioritization when the self-associated stimuli need to compete for attentional resources with other-associated stimuli.
Notes
The detection task produces a different pattern than the discrimination task, which might be due to different effects being observed by each task (inhibition of return vs. cuing effect) stimulated by different stimulus onset asynchrony requirements of the tasks (see supplementary material for results and discussion).
References
Alexopoulos, T., Muller, D., Ric, F., & Marendaz, C. (2012). I, me, mine: Automatic attentional capture by self-related stimuli. European Journal of Social Psychology, 42(6), 770–779. https://doi.org/10.1002/ejsp.1882
Arnell, K. M., Shapiro, K. L., & Sorensen, R. E. (1999). Reduced repetition blindness for one’s own name. Visual Cognition, 6(6), 609–635. https://doi.org/10.1080/135062899394876
Bargh, J. A. (1982). Attention and automaticity in the processing of self-relevant information. Journal of Personality and Social Psychology, 43(3), 425–436. https://doi.org/10.1037/0022-3514.43.3.425
Blume, C., del Giudice, R., Lechinger, J., Wislowska, M., Heib, D. P. J., Hoedlmoser, K., & Schabus, M. (2017). Preferential processing of emotionally and self-relevant stimuli persists in unconscious N2 sleep. Brain and Language, 167, 72–82. https://doi.org/10.1016/j.bandl.2016.02.004
Bola, M., Paź, M., Doradzińska, Ł., & Nowicka, A. (2020). The self-face automatically captures attention without consciousness [Preprint]. Neuroscience. https://doi.org/10.1101/2020.01.22.915595
Brédart, S., Delchambre, M., & Laureys, S. (2006). One’s own face is hard to ignore. Quarterly Journal of Experimental Psychology, 59(1), 46–52. https://doi.org/10.1080/17470210500343678
Caughey, S., Falbén, J. K., Tsamadi, D., Persson, L. M., Golubickis, M., & Neil Macrae, C. (2021). Self-prioritization during stimulus processing is not obligatory. Psychological Research, 85(2), 503–508. https://doi.org/10.1007/s00426-019-01283-2
Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). Erlbaum
Dalmaso, M., Castelli, L., & Galfano, G. (2019). Self-related shapes can hold the eyes. Quarterly Journal of Experimental Psychology. https://doi.org/10.1177/1747021819839668
Ehrman, R. N., Robbins, S. J., Bromwell, M. A., Lankford, M. E., Monterosso, J. R., & O’Brien, C. P. (2002). Comparing attentional bias to smoking cues in current smokers, former smokers, and nonsmokers using a dot-probe task. Drug and Alcohol Dependence, 67(2), 185–191. https://doi.org/10.1016/S0376-8716(02)00065-0
Falbén, J. K., Golubickis, M., Balseryte, R., Persson, L. M., Tsamadi, D., Caughey, S., & Neil Macrae, C. (2019). How prioritized is self-prioritization during stimulus processing? Visual Cognition, 27: 1–6. https://doi.org/10.1080/13506285.2019.1583708
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191. https://doi.org/10.3758/BF03193146
Frischen, A., Bayliss, A. P., & Tipper, S. P. (2007). Gaze cueing of attention: Visual attention, social cognition, and individual differences. Psychological Bulletin, 133(4), 694–724. https://doi.org/10.1037/0033-2909.133.4.694
Hautus, M. J. (1995). Corrections for extreme proportions and their biasing effects on estimated values of d′. Behavior Research Methods, Instruments, & Computers, 27(1), 46–51. https://doi.org/10.3758/BF03203619
Hawkins, H. L., Hillyard, S. A., Luck, S. J., Mouloua, M., Downing, C. J., & Woodward, D. P. (1990). Visual attention modulates signal detectability. Journal of Experimental Psychology: Human Perception and Performance, 16(4), 802–811. https://doi.org/10.1037/0096-1523.16.4.802
Humphreys, G. W., & Sui, J. (2015). Attentional control and the self: The self-attention network (SAN). Cognitive Neuroscience, 7(1–4), 5–17. https://doi.org/10.1080/17588928.2015.1044427
Imhoff, R., Lange, J., & Germar, M. (2019). Identification and location tasks rely on different mental processes: A diffusion model account of validity effects in spatial cueing paradigms with emotional stimuli. Cognition and Emotion, 33(2), 231–244. https://doi.org/10.1080/02699931.2018.1443433
Janczyk, M., Humphreys, G. W., & Sui, J. (2019). The central locus of self-prioritisation. Quarterly Journal of Experimental Psychology, 72(5), 1068–1083. https://doi.org/10.1177/1747021818778970
JASP Team. (2020). JASP (Version 0.14.1) [Computer software].
Jenkins, J. R., Neale, D. C., & Deno, S. L. (1967). Differential memory for picture and word stimuli. Journal of Educational Psychology, 58(5), 303–307. https://doi.org/10.1037/h0025025
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The fusiform face area: A module in human extrastriate cortex specialized for face perception. The Journal of Neuroscience, 17(11), 4302–4311. https://doi.org/10.1523/JNEUROSCI.17-11-04302.1997
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: A cortical region specialized for the perception of faces. Philosophical Transactions of the Royal Society, B: Biological Sciences, 361(1476), 2109–2128. https://doi.org/10.1098/rstb.2006.1934
Landkammer, F., Winter, K., Thiel, A., & Sassenberg, K. (2019). Team sports off the field: Competing excludes cooperating for individual but not for team athletes. Frontiers in Psychology, 10: 2470. https://doi.org/10.3389/fpsyg.2019.02470
Liu, M., He, X., Rotsthein, P., & Sui, J. (2016). Dynamically orienting your own face facilitates the automatic attraction of attention. Cognitive Neuroscience, 7(1/4), 37–44. https://doi.org/10.1080/17588928.2015.1044428
Lockhead, G. R. (1966). Effects of dimensional redundancy on visual discrimination. Journal of Experimental Psychology, 72(1), 95–104. https://doi.org/10.1037/h0023319
MacMahon, K. M. A., Broomfield, N. M., & Espie, C. A. (2006). Attention bias for sleep-related stimuli in primary insomnia and delayed sleep phase syndrome using the dot-probe task. Sleep, 29(11), 1420–1427. https://doi.org/10.1093/sleep/29.11.1420
Macrae, C. N., Visokomogilski, A., Golubickis, M., Cunningham, W. A., & Sahraie, A. (2017). Self-relevance prioritizes access to visual awareness. Journal of Experimental Psychology: Human Perception and Performance, 43(3), 438–443. https://doi.org/10.1037/xhp0000361
Macrae, C. N., Visokomogilski, A., Golubickis, M., & Sahraie, A. (2018). Self-relevance enhances the benefits of attention on perception. Visual Cognition, 26(7), 475–481. https://doi.org/10.1080/13506285.2018.1498421
Mattan, B., Quinn, K. A., Apperly, I. A., Sui, J., & Rotshtein, P. (2015). Is it always me first? Effects of self-tagging on third-person perspective-taking. Journal of Experimental Psychology: Learning, Memory, and Cognition, 41(4), 1100–1117. https://doi.org/10.1037/xlm0000078
Moray, N. (1959). Attention in dichotic listening: Affective cues and the influence of instructions. Quarterly Journal of Experimental Psychology, 11(1), 56–60. https://doi.org/10.1080/17470215908416289
Nijhof, A. D., Shapiro, K. L., Catmur, C., & Bird, G. (2020). No evidence for a common self-bias across cognitive domains. Cognition, 197, Article 104186. https://doi.org/10.1016/j.cognition.2020.104186
O'Brien, R. G., Kaiser, M. K. (1985)1 MANOVA method for analyzing repeated measures designs: An extensive primer. Psychological Bulletin, 97(2), 316-333. https://doi.org/10.1037/0033-2909.97.2.316
Orellana-Corrales, G., Matschke, C., & Wesslein, A.-K. (2020). Does self-associating a geometric shape immediately cause attentional prioritization? Comparing familiar versus recently self-associated stimuli in the Dot-Probe Task. Experimental Psychology, 67(6), 335–348. https://doi.org/10.1027/1618-3169/a000502
Perrett, D. I., Hietanen, J. K., Oram, M. W., & Benson, P. J. (1992). Organization and functions of cells responsive to faces in the temporal cortex. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 335(1273), 23–30. https://doi.org/10.1098/rstb.1992.0003
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32(1), 3–25. https://doi.org/10.1080/00335558008248231
Reuther, J., & Chakravarthi, R. (2017). Does self-prioritization affect perceptual processes? Visual Cognition, 25(1/3), 381–398. https://doi.org/10.1080/13506285.2017.1323813
Schäfer, S., Wentura, D., & Frings, C. (2015). Self-prioritization beyond perception. Experimental Psychology, 62(6), 415–425. https://doi.org/10.1027/1618-3169/a000307
Schäfer, S., Wesslein, A.-K., Spence, C., Wentura, D., & Frings, C. (2016). Self-prioritization in vision, audition, and touch. Experimental Brain Research, 234(8), 2141–2150. https://doi.org/10.1007/s00221-016-4616-6
Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-Prime: User’s guide (2.0) [Computer software]. Psychology Software Incorporated
Shor, R. E. (1971). Symbol processing speed differences and symbol interference effects in a variety of concept domains. The Journal of General Psychology, 85(2), 187–205. https://doi.org/10.1080/00221309.1971.9920672
Siebold, A., Weaver, M. D., Donk, M., & van Zoest, W. (2015). Social salience does not transfer to oculomotor visual search. Visual Cognition, 23(8), 989–1019. https://doi.org/10.1080/13506285.2015.1121946
Sperber, R. D., McCauley, C., Ragain, R. D., & Weil, C. M. (1979). Semantic priming effects on picture and word processing. Memory & Cognition, 7(5), 339–345. https://doi.org/10.3758/BF03196937
Stanislaw, H., & Todorov, N. (1999). Calculation of signal detection theory measures. Behavior Research Methods, Instruments, & Computers, 31(1), 137–149. https://doi.org/10.3758/BF03207704
Stein, T., Siebold, A., & van Zoest, W. (2016). Testing the idea of privileged awareness of self-relevant information. Journal of Experimental Psychology: Human Perception and Performance, 42(3), 303–307. https://doi.org/10.1037/xhp0000197
Sui, J., He, X., & Humphreys, G. W. (2012). Perceptual effects of social salience: Evidence from self-prioritization effects on perceptual matching. Journal of Experimental Psychology: Human Perception and Performance, 38(5), 1105–1117. https://doi.org/10.1037/a0029792
Sui, J., & Humphreys, G. W. (2015). More of me! Distinguishing self and reward bias using redundancy gains. Attention, Perception, & Psychophysics, 77(8), 2549–2561. https://doi.org/10.3758/s13414-015-0970-x
Sui, J., Liu, C. H., Wang, L., & Han, S. (2009). Attentional orientation induced by temporarily established self-referential cues. Quarterly Journal of Experimental Psychology, 62(5), 844–849. https://doi.org/10.1080/17470210802559393
Sui, J., & Rotshtein, P. (2019). Self-prioritization and the attentional systems. Current Opinion in Psychology, 29, 148–152. https://doi.org/10.1016/j.copsyc.2019.02.010
Tacikowski, P., & Nowicka, A. (2010). Allocation of attention to self-name and self-face: An ERP study. Biological Psychology, 84(2), 318–324. https://doi.org/10.1016/j.biopsycho.2010.03.009
Truong, G., Roberts, K. H., & Todd, R. M. (2017). I saw mine first: A prior-entry effect for newly acquired ownership. Journal of Experimental Psychology: Human Perception and Performance, 43(1), 192–205. https://doi.org/10.1037/xhp0000295
Tsao, D. Y., Freiwald, W., Tootell, R. B. H., & Livingstone, M. (2006). A cortical region consisting entirely of face-selective cells. Science, 311(5761), 670–674. https://doi.org/10.1126/science.1119983
Tukey, J. W. (1977). Exploratory data analysis. Addison-Wesley
Verghese, P. (2001). Visual search and attention. Neuron, 31(4), 523–535. https://doi.org/10.1016/S0896-6273(01)00392-0
Wade, G. L., & Vickery, T. J. (2018). Target self-relevance speeds visual search responses but does not improve search efficiency. Visual Cognition, 26(8), 563–582. https://doi.org/10.1080/13506285.2018.1520377
Wójcik, M. J., Nowicka, M. M., Kotlewska, I., & Nowicka, A. (2018). Self-face captures, holds, and biases attention. Frontiers in Psychology, 8. https://doi.org/10.3389/fpsyg.2017.02371
Wood, N., & Cowan, N. (1995). The cocktail party phenomenon revisited: How frequent are attention shifts to one’s name in an irrelevant auditory channel? Journal of Experimental Psychology: Learning, Memory, and Cognition, 21(1), 255–260. https://doi.org/10.1037/0278-7393.21.1.255
Woźniak, M., & Hohwy, J. (2020). Stranger to my face: Top-down and bottom-up effects underlying prioritization of images of one’s face. PLoS ONE, 15(7), e0235627. https://doi.org/10.1371/journal.pone.0235627
Woźniak, M., & Knoblich, G. (2019). Self-prioritization of fully unfamiliar stimuli. Quarterly Journal of Experimental Psychology, 72(8), 2110–2120. https://doi.org/10.1177/1747021819832981
Woźniak, M., Kourtis, D., & Knoblich, G. (2018). Prioritization of arbitrary faces associated to self: An EEG study. PLoS ONE, 13(1), e0190679. https://doi.org/10.1371/journal.pone.0190679
Yang, H., Wang, F., Gu, N., Gao, X., & Zhao, G. (2013). The cognitive advantage for one’s own name is not simply familiarity: An eye-tracking study. Psychonomic Bulletin & Review, 20(6), 1176–1180. https://doi.org/10.3758/s13423-013-0426-z
Yovel, G., & Kanwisher, N. (2004). Face perception: Domain specific, not process specific. Neuron, 44(5), 889–898. https://doi.org/10.1016/j.neuron.2004.11.018
Zhao, S., Uono, S., Yoshimura, S., & Toichi, M. (2015). Self make-up: The influence of self-referential processing on attention orienting. Scientific Reports, 5(1), Article 14169. https://doi.org/10.1038/srep14169
Acknowledgements
This research was conducted as part of the Leibniz-WissenschaftsCampus Tübingen “Cognitive Interfaces.”
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Open practices statement
The collected data and analyses scripts for Study 1 are available on the Open Science Framework (OSF) (https://osf.io/g7wrc/)
Study 2 was preregistered in the OSF (https://osf.io/s68pm). The collected data and analyses scripts are available on the OSF (https://osf.io/4cwrv/).
Conflicts of interest
The authors report no conflicts of interest that are relevant to this article.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 69.3 kb)
Rights and permissions
About this article

Cite this article
Orellana-Corrales, G., Matschke, C. & Wesslein, AK. The impact of newly self-associated pictorial and letter-based stimuli in attention holding. Atten Percept Psychophys 83, 2729–2743 (2021). https://doi.org/10.3758/s13414-021-02367-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-021-02367-1