
Abstract
The temporal loci of distractor processing were assessed in a flanker task with mutating distractors. We introduce the mutations paradigm, which allows for behavioral assessments of the critical time window during which distractors are processed. A central target was flanked by two identical distractors. While the target remained unchanged throughout the trial, the distractors’ identities mutated once per trial, at a random time during the initial 200 ms following onset. There were three types of trials: incongruent (i.e., disruptive) distractors that mutated to neutral distractors, neutral distractors that mutated to incongruent ones, or neutral distractors that mutated to different neutral distractors (control). The results revealed that presentations of incongruent distractors for a mere 17 ms were sufficient to significantly delay responses. After 50 ms, perceptual information ceased to be accumulated from distractors locations but was still being collected from the target location. We suggest that (a) extensive information about the target and distractors was gathered as early as 17 ms after onset; (b) attentional modulations of processing consummated later, between 34 and 51 ms; and (c) once attentional mechanisms had stepped in (~50 ms), selection achieved full and sustained efficiency. These findings seem to challenge basic assumptions held by early-selection, late-selection, and load theories.
Similar content being viewed by others


Research on attention has mostly focused on the conditions under which the perceptual system fails to filter out irrelevant stimuli during the course of processing targeted stimuli (e.g., Stroop effects: MacLeod, 1991; Stroop, 1935; flanker effects: Eriksen, 1995; Eriksen & Eriksen, 1974; Miller, 1991; global precedence effects: Kimchi, 1992; Navon, 1977; or Simon effects: Simon, 1990). We suggest that in order to explain undesired processing of task-irrelevant distractors, theoretical models have relied on specific assumptions regarding the temporal window during which distractors are processed. Inescapably, the adoption of different assumptions about the time course of attentional modulations of processing has led to fundamentally disparate interpretations of how attention operates. The main focus of the present study was to provide behavioral assessments of the critical time window during which information about distractors is accumulated.
Explicit and hidden assumptions about the temporal locus during which disruptive information about distractors infiltrates attentional selection have directly derived from theoretical propositions regarding the temporal locus of attentional operation. For instance, late-selection theories have suggested that initial perceptual processing is unrestricted and that targets are discerned from nontargets during later, postperceptual stages (Deutsch & Deutsch, 1963; Duncan, 1980; Schneider & Shiffrin, 1977). Thus, according to this view, information about distractors is continually accumulated for as long as the distractors are presented.
Early-selection theories, on the other hand, propose that initial processing proceeds automatically and in parallel, but can only retrieve coarse information. When necessary, resolution is improved by the activation of attentional mechanisms that filter the very inflow of perceptual information (Broadbent, 1958, 1970, 1971, 1981, 1982; Johnston & Dark, 1982; Lachter, Forster, & Ruthruff, 2004). Alternatively, perceptual-load theory (Lavie, 1995, 2005, 2010; Lavie & Tsal, 1994) proposes that if target selection demands less than all available resources (low load), then unused surplus resources end up processing neighboring nontargets. If so, distractor processing can only begin after targets have been selected.
Numerous studies have manipulated and assessed temporal variables in selection tasks. However, the majority of these studies have investigated the time courses of attentional capture (see Theeuwes, 2010, for a review), attentional shifting, or attentional orienting (Eriksen & Webb, 1989; Eriksen & Yeh, 1985; Jonides, 1981; Müller & Rabbitt, 1989; Posner, 1980; Remington, 1980; Taylor, 1977; Tsal, 1983; see Egeth & Yantis, 1997, for a review). These studies have assessed the time course of processing by manipulating presentation asynchronies between targets and distractors or by means of capturing cues or features. In the present experiments, on the other hand, we presented simultaneous stimuli without precues and specifically assessed the critical time window during which a known-to-disrupt distractor must be presented in order to impair responses to a target.
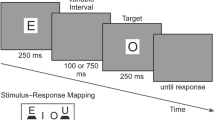
We introduce here the mutations paradigm, which allows us to measure the level of disruption inflicted by a distractor at different stages during the course of a trial. The specific time during which the system gathers information about distractors is inferred from the time window within which a disruptive distractor must appear in order to delay responses. The task comprises a flanker display (Eriksen & Eriksen, 1974), with a central target flanked by two identical distractors, which can be either incongruent (i.e., disruptive) or neutral. Yet, in the mutations paradigm, the distractors change (or mutate) once per trial, while the target remains unchanged throughout the entire trial. Three types of trials are randomly intermixed within each experimental block: trials that begin with incongruent distractors that mutate to neutral ones (incongruent-to-neutral), trials that begin with neutral distractors that mutate to incongruent ones (neutral-to-incongruent), and trials that begin with neutral distractors that mutate to different neutral distractors (neutral-to-neutral), which serve as the control trials. On each trial, the mutation occurs at a random time within the initial 200 ms after stimulus onset (Fig. 1).

Mutations paradigm. On every trial, the distractors mutate once, whereas the target remains unchanged. Both distractors mutate together at a random time during the initial 200 ms after stimulus onset. Both examples above depict an incongruent-to-neutral trial mutated at 85 ms. (a) Experiment 1. (b) Experiment 2, in which three hash marks appeared for 17 ms between the pre- and postmutation stimuli. (c) Six sets of stimuli (see the text)
These manipulations yield straightforward predictions: (a) If distractors are processed only during early stages of a presentation, incongruent-to-neutral trials should render slower response times (RT) than neutral-to-neutral trials. (b) If distractors are continually processed throughout the entire presentation, we should observe a positive correlation between RT and the overall presentation duration of incongruent distractors (irrespective of their onset and offset times). (c) If distractors are processed at later stages, we should expect slower RTs for neutral-to-incongruent than for neutral-to-neutral trials.
Experiment 1
Method
Participants
Twenty undergraduate students from Tel Aviv University participated in Experiments 1 and 2 for course credit.
Stimuli and procedure
Participants were instructed to respond to the target as quickly as possible by pressing either the z key or the m key on a standard keyboard. A central target (the letter X or Z) was flanked by two identical distractors (either Xs, Zs, or Ps). The distractors were either neutral (P), congruent (identical to the target), or incongruent (i.e., incongruent to the trial’s target; distractors Z for target X, or distractors X for target Z) (Fig. 1c).
On every trial, both distractors mutated once, while the target remained unchanged. Distractors randomly mutated to one of the two other alternatives (e.g., Zs could mutate to either Xs or Ps). The mutation randomly occurred at one out of 11 alternative times, between 17 and 187 ms after onset, in 17-ms time steps.
The stimuli were controlled by a standalone applicationFootnote 1 created with the MATLAB Psychophysics Toolbox (Brainard, 1997), presented on a 19-in. CRT monitor (60 Hz). All stimuli were black on a white background, subtended 1° in height and 0.7° in width, and were separated by 1.2° (center to center).
Each observer participated in one session comprising two training blocks of 30 trials, followed by eight experimental blocks of 200 trials each (four blocks of Exp. 1 and four blocks of Exp. 2, randomly ordered).Footnote 2 On each trial, a fixation cross was presented (400 ms), followed by a blank screen (300 ms) and the stimuli, which remained on screen until response. A blank screen appeared immediately after a response was input and remained on screen for 100 ms, after which a new trial began.
Results and discussion
As is often observed in flanker tasks, neutral displays (PXP and PZP) and congruent displays (XXX and ZZZ) produced nearly identical responses for all trial types (see Fig. 2 and Table S8 and S9)Footnote 3 (see also Lavie, 1995). Accordingly, all patterns observed remained unchanged, both when congruent trials were excluded from analyses and when neutral trials were excluded. Thus, neutral and congruent distractors were combined in all further analyses, and both were labeled as neutral (i.e., nondisruptive).

Experiment 1: Mean reactions times (RTs) by mutation times, showing comparisons between congruent and neutral distractors within the three trial types. (a) Neutral-to-neutral trials. (b) Incongruent-to-neutral trials. (c) Neutral-to-incongruent trials. Error bars represent one standard error. At the bottom of each panel, bars and asterisks denote effect size and p < .05, respectively. The dashed line illustrates η 2 = .5
Errors and responses deviating by two SDs from the mean RT for each participant were discarded (a total of 1,376 errors and 2,756 outliers). Error rates (M = 4.3 %, SD = 2) generally mirrored the RT patterns for the three mutation types across all mutation times (Fig. 3a and d, Tables S1 and S2).

Experiments 1 and 2: (a, b, c) Mean reaction times (RTs) by mutation times for the three display types. (d, e, f) Error rates by mutation times. (a, d) Experiment 1. (b, e) Experiment 2. (c, f) Experiments 1 and 2 combined. At the bottom of each panel, bars and asterisks denote effect size and p < .05, respectively. Blue (or cool-colored) bars and asterisks refer to incongruent-to-neutral versus neutral-to-neutral (control) trials. Red (or warm-colored) bars and asterisks refer to neutral-to-incongruent versus neutral-to-neutral (control) trials. Dashed lines illustrate η 2 = .5
For assessing the time window during which incongruent distractors must be presented in order to impair responses, we must first measure the level of congruency interference (CI) for each mutation time. To evaluate incongruent-to-neutral interference, we compared these trials to neutral-to-neutral (control) trials for each mutation time. Incongruent-to-neutral interferences are represented in Fig. 3 by the distances between the incongruent-to-neutral trials (blue function line) and the neutral-to-neutral trials (black function line). Neutral-to-incongruent interferences were computed in the same manner (depicted by the distances between the red and the black functions in Fig. 3a).
Repeated measures analyses of variance (ANOVAs) were conducted on the three mutation types for each mutation time.Footnote 4 Neutral-to-neutral (control) trials resulted in similar RTs, irrespective of mutation times (M = 406 ms, SD = 13), F(10, 19) = 0.73, p = .686, η 2 = .04.
Incongruent-to-neutral trials significantly impaired performance for all mutation times (see the blue asterisks in Fig. 3a; see also Table S2). The leftmost blue dot in Fig. 3a represents all trials on which incongruent distractors were presented during the initial 17 ms, after which they mutated to neutral distractors that remained on screen until response. These trials resulted in responses that were 9 ms slower than trials on which neutral distractors were presented during the initial 17 ms and then mutated to different neutral distractors (leftmost black dot). This means that a mere 17-ms presentation of incongruent distractors was enough to significantly impair performance: Incongruent-to-neutral trials that mutated at 17 ms (M = 416 ms) resulted in slower RTs than did neutral-to-neutral trials mutated at 17 ms (M = 407 ms), F(1, 19) = 7.27, p = .014, η 2 = .28. In sum, all trials on which incongruent distractors were presented at stimulus onset resulted in significant interference, irrespective of how long those distractors remained on screen.
Neutral-to-incongruent trials, on the other hand, generated interference only when incongruent distractors appeared at the very beginning of the trial. Apart from trials on which neutral distractors mutated to incongruent distractors at 170 ms — when interference was marginally significant (ΔRT = 8 ms), F(1, 19) = 4.45, p = .048, η 2 = .19 — all neutral-to-incongruent mutations that occurred later than 50 ms resulted in RTs virtually identical to those on neutral-to-neutral trials (see red and black functions in Fig. 3a). When incongruent distractors appeared at 17 ms (M = 421 ms), responses were 14 ms slower than neutral-to-neutral trials, F(1, 19) = 8.85, p = .008, η 2 = .32. When incongruent distractors appeared at 34 ms, disruption approached significance (ΔRT = 14 ms), F(1, 19) = 4.31, p = .052.
These results suggest that disruptive information about distractors was accumulated during the initial 50 ms, after which the distractors’ identities became inconsequential. These conclusions are further supported by four additional patterns. First, incongruent-to-neutral trials resulted in significantly slower RTs than neutral-to-incongruent trials for all mutation times later than 34 ms (Table S2). Second, ANOVA comparisons of interferences between the conditions (Table 1) revealed that incongruent-to-neutral interference was drastically larger than neutral-to-incongruent interference for all mutations that occurred later than 34 ms.
Third, prolonged presentations of incongruent distractors on incongruent-to-neutral trials increased interference only within the initial 50 ms, after which interference plateaued: Incongruent-to-neutral trials mutated between 51 and 187 ms resulted in similar interference (M = 18 ms), F(8, 19) = 0.84, p = .567, η 2 = .04. Incongruent-to-neutral trials mutated at 17 ms (mean interference = 9 ms) resulted in significantly less interference than all trials mutated later than 34 ms (M = 18 ms), F(1, 19) = 4.93, p = .038, η 2 = .21, as did trials mutated at 34 ms (M = 12 ms), F(1, 19) = 1.98, p = .017, η 2 = .09. Together, incongruent-to-neutral mutations that occurred earlier than 50 ms (M = 10 ms) resulted in significantly less interference than did mutations later than 50 ms (M = 18 ms), F(1, 19) = 5.07, p = .036, η 2 = .21.
Fourth, neutral-to-incongruent trials on which distractors mutated before 50 ms (M = 14 ms) resulted in significantly greater interference than trials on which the mutation occurred after 50 ms: difference in congruency interference (ΔCI) = 11 ms, F(1, 19) = 4.87, p = .039, η 2 = .20. After 50 ms, neutral-to-incongruent mutations generated similar interference (M = 3 ms), F(8, 19) = 0.55, p = .817, η 2 = .02.
On the other hand, two patterns reveal a small, yet statistically significant, amount of distractor processing that persisted beyond 50 ms. First, although the ANOVA for incongruent-to-neutral interference after 50 ms revealed no main effect (M = 18 ms), F(8, 19) = 0.84, p = .567, η 2 = .04, Pearson’s linear correlation coefficients indicated a possible gradual increase of interference for incongruent-to-neutral mutations later than 50 ms (see Table 2). When interference was computed as ΔRT, it was significantly correlated with mutation times (r = .69, p = .038). Yet, when interference was computed as a percentage of the total RT (see the parentheses in Table 2), this correlation only approached significance (r = .66, p = .052).
Second, the differences in error rates between neutral-to-incongruent and neutral-to-neutral trials on which distractors mutated after 50 ms (represented by the distances between the red and black functions in Fig. 3d) were negatively correlated with mutation times (r = −.76, p = .017).Footnote 5 In sum, it seems that although the bulk of disruption by incongruent distractors took place within the initial 50 ms, prolonged presentations of incongruent distractors may have continued to (marginally) increase disruption.
It should be noted that the conclusion that most disruptive information about distractors was accumulated during the initial 50 ms does not mean that the extracted information was immediately processed, nor that the distractors’ identities were recognized at 50 ms. It is quite possible that at this stage the system just collected perceptual information, whereas the actual processing, identification, and selection occurred later, during higher-level processing.
In summary, these results indicate that incongruent distractors impaired performance only when presented within the initial 50 ms. The onset time and presentation duration of incongruent distractors presented later than 50 ms were virtually inconsequential. The gist of these patterns is the drastic drop in RTs for neutral-to-incongruent trials on which the mutation occurred later than 50 ms—depicted in Fig. 3a, where the neutral-to-incongruent function suddenly plunges and stabilizes within the same RT values as the neutral-to-neutral (control) trials function.
Experiment 2
The chief purpose of Experiment 2 was to replicate the main patterns observed in Experiment 1 and to reassess the conclusion that the accumulation of information about distractors practically terminated around 50 ms after onset. In Experiment 2 we followed the exact same procedures as in Experiment 1, with the addition of a 17-ms interruption of stimulus presentation at different times after onset. If the target and the distractors are processed at different times, the interruption should affect responses differently depending on when it occurred.
The interruption was effectuated by three hash marks (#) presented for 17 ms between the premutation and postmutation stimuli (see Fig. 1b). In this manner, Experiment 2 was identical to Experiment 1, except for the 17-ms interruption entailed in each mutation. This allowed us not only to assess interactions between the interrupted mutations and times of mutation, but also to directly compare corresponding trials in both experiments (i.e., trials of the same type and same mutation time).Footnote 6 Phenomenologically, participants could not discriminate between the equivalent trials of the two experiments.
The addition of an interruption at different times after onset yielded the following predictions: (a) If not only the distractors, but also the target ceases to be perceptually processed at 50 ms, then all interruptions later than 50 ms should be inconsequential, and Experiment 2 mutations later than 50 ms should result in the same RTs as their equivalents in Experiment 1. If, on the other hand, targets continue to be processed after 50 ms, the Experiment 2 mutations later than 50 ms should result in slower responses than their counterparts of Experiment 1. This delay should be observed for as long as target processing persists. (b) If interruptions occurring before 50 ms disrupt the processing of all three stimuli, whereas interruptions after 50 ms disrupt only target processing, then neutral-to-neutral interrupted mutations occurring before 50 ms should render slower RTs than neutral-to-neutral interrupted mutations occurring later than 50 ms.
Results and discussion
Errors and responses deviating by two SDs from the mean RT of each participant were discarded (a total of 1,408 errors and 2,754 outliers). Error rates (M = 4.4 %, SD = 2.2) mirrored the RT patterns for the three mutation types across all mutation times (Fig. 3b and e). Repeated measures ANOVAs conducted across the three mutation types for each mutation time (Tables S3 and S4) revealed patterns similar to those observed in Experiment 1. They converged toward the conclusion that the accumulation of information about distractors was restricted to the initial 50 ms after onset.
Regarding the interruption manipulation, had the interrupted mutations, instead of temporarily interrupting accumulation, ended up masking the premutation stimuli or resetting processing anew, then we would observe a strong bias toward the postmutation distractors for all mutation types. This was clearly not the case.
In Fig. 3b, we can see that incongruent-to-neutral trials (blue function line) consistently rendered longer RTs than neutral-to-incongruent trials (red function line) for all mutations later than 34 ms, and resulted in significant interference for most mutation times. Neutral-to-incongruent mutations, on the other hand, only caused interference when the incongruent distractors were presented at 17 ms following onset.
Foremost, ANOVA comparisons between interference patterns (Table 1) revealed that incongruent-to-neutral interference was drastically greater than neutral-to-incongruent interference for all mutations that occurred later than 34 ms, apart from mutations at 136 ms.Footnote 7
Beyond the Experiment 1 replications described above, the main focus of Experiment 2 was to assess the effects of the 17-ms interruption entailed in each mutation. Interruptions (i.e., mutations) within the initial 50 ms were expected to disrupt the perceptual processing of all three stimuli, whereas interruptions later than 50 ms were expected to disrupt only target processing. Two convergent patterns substantiate this hypothesis.
First, contrary to Experiment 1, in which neutral-to-neutral trials resulted in similar RTs for all mutation times, in Experiment 2, neutral-to-neutral mutations that occurred before 50 ms resulted in much slower RTs than mutations later than 50 ms (black function in Fig. 3b). Whereas we observed no RT differences between neutral-to-neutral mutations that occurred later than 50 ms (M = 419 ms), F(8, 19) = 1.04, p = .408, η 2 = .05, mutations that occurred at 17 ms (M = 440 ms) caused RTs to be 21 ms slower than all mutations later than 50 ms, F(1, 19) = 20.40, p < .001, η 2 = .52. Mutations at 34 ms (M = 433 ms) led to RTs 14 ms slower than all mutations later than 50 ms, F(1, 19) = 7.05, p = .015, η 2 = .27.
Second, Fig. 4 shows that the interrupted mutations of Experiment 2 indeed interrupted processing and significantly delayed responses in comparison to the equivalent trials of Experiment 1 for most mutation times and types (see also Table S7). This RT cost was observed as late as 187 ms after onset (Fig. 4c). More importantly, in accord with our predictions, interruptions that occurred within the initial 50 ms disrupted responses twice as much as interruptions that took place later than 50 ms. Experiment 2 trials on which the mutation occurred before 50 ms led to significantly slower responses than their equivalents in Experiment 1, by an average of 26 ms, F(1, 19) = 30.93, p < .001, η 2 = .62. Experiment 2 trials on which the mutation occurred later than 50 ms delayed responses by an average of only 13 ms, F(1, 19) = 14.79, p = .001, η 2 = .43. The mean response delay observed before 50 ms (ΔRT = 26 ms) was significantly greater than the delay observed later than 50 ms (ΔRT = 13 ms), F(1, 19) = 29.01, p < .001, η 2 = .60.

Mean reaction times (RTs) by mutation times, and pairwise comparisons between the equivalent trials of Experiments 1 and 2. (a) Neutral-to-neutral trials. (b) Incongruent-to-neutral trials. (c) Neutral-to-incongruent trials. Error bars represent one standard error of the mean. At the bottom of each panel, bars and asterisks denote effect size and p < .05, respectively. The dashed line illustrates η 2 = .5
We have attributed the slow responses in interrupted mutations that occurred later than 50 ms to disruptions of perceptual processing of the targets. Yet the fact that target processing could have been disrupted as late as 187 ms does not mean that the information accumulated at that time was still insufficient for identifying the target. It is possible that the system had gathered sufficient information for a response much earlier, maybe as early as 30 ms, but then still continued to monitor the target for another 200 ms or more. This could reflect confidence thresholds for a sure decision rather than an ongoing accumulation of perceptual information necessary for reaching a decision about the target’s identity.
To our understanding, the main conclusion to be drawn from the comparisons between Experiments 1 and 2 is that the events observed around 50 ms reflect top-down attentional modulation of distractor processing, and not a spontaneous dwindling of perceptual activity in general. These comparisons show that the changes in processing patterns observed before and after 50 ms were different for targets and nontargets. Even if we assume that the system ceased to actively amass critical information after 50 ms, it is still clear that the system continued to monitor eventual changes onscreen until (at least) 187 ms, but this monitoring was confined exclusively to the targeted object.
General discussion
The results of both experiments (Fig. 3c and f, Tables S5 and S6) revealed two main patterns: First, a mere 17-ms presentation of incongruent distractors sufficed to significantly delay responses. Second, incongruent (i.e., disruptive) distractors impaired performance only if they were presented within the initial ~50 ms.Footnote 8
These patterns suggest that the system gathered extensive information about all presented stimuli during the initial 50 ms following onset. At some point between 34 and 51 ms, the accumulation of information from the distractors’ locations abruptly halted, whereas the collection of information from the target’s location persisted (at least to some extent). We concluded that the changes observed around 50 ms reflect attentional modulations of perceptual processing that deliberately and effectively discriminated the target from nontargets (see the “Results and discussion” section of Exp. 2).
If indeed the events observed at ~50 ms reflect attentional activity, this means that flanker interference stemmed from perceptual processing that occurred before attentional operation, and not as a consequence of an incapacity of attentional mechanisms. In fact, once attentional modulations consummated (at ~50 ms), selection became fully and steadily efficient. This order of events seems to challenge early-selection claims that attentional operation precedes distractor interference, and that the latter stems from inherently flawed selection mechanisms. Late selection, on the other hand, is based on the assumption that attentional activity is strictly postperceptual, whereas here we observed strong attentional modulations of perception within the initial 50 ms of processing.
The early-selection approach (Broadbent, 1958, 1970, 1971, 1981, 1982; Treisman, 1969) assumes a critically limited capacity for initial perceptual processing, which begins in an automatic and parallel mode but remains restricted to rudimentary resolution, insufficient to attain stimulus identification. If the system becomes overloaded or fails to reach the demanded levels of resolution, attentional mechanisms step in and allow only selected stimuli to reach further processing. Attentional selection can be accomplished by different strategies, such as filtering, pigeon-holing, or categorizing (Broadbent, 1971), which are determined by the current task demands and differ between individuals (Broadbent, 1981). Identifications can only be accomplished at later stages that include access to memory buffers (for reviews, see Broadbent, 1982; Johnston & Dark, 1982; Kahneman & Treisman, 1984; Lachter et al., 2004).
Early selection theories have interpreted distractor interference as ineffective attentional deployment, because “if the person knows what part of the stimulus field to select, the interference disappears. The interference must therefore be before the choice of response, and the selection must be earlier still” (Broadbent, 1982, p. 281). That is, if the system ended up resorting to attentional selection, it must have preceded stimulus identification (see also Driver, 2001; Johnston & Dark, 1986; Paquet, 2001; Yantis & Johnston, 1990). Yet, as we have described above, the present results show that the distractors were identified as early as 17 ms after onset, whereas attentional selection took at least twice that amount of time to consummate (between 34 and 51 ms following onset).
Specifically regarding flanker effects, early-selection theories and their present-day corollaries have described flanker interference as an undesired leakage or slippage of perceptual processing toward flanking distractors (Baylis & Driver, 1992; Broadbent, 1982; Driver, 2001; Lachter et al., 2004; Lavie & Tsal, 1994; Miller, 1991; Yantis & Johnston, 1990). Attentional mechanisms have been considered unable to concentrate resources exclusively within the target, maybe as a consequence of Gestalt-like tendencies to chunk adjacent letters into one object (Baylis & Driver, 1992; Driver & Baylis, 1989; Prinzmetal, 1981), or possibly because the target’s area in a flanker task is usually too small— it has been generally suggested that distractors located less than 1° away from the target are inevitably processed (Broadbent, 1982; Eriksen & Eriksen, 1974; Miller, 1991; Paquet, 2001; Paquet & Craig, 1997).Footnote 9
Accordingly, attentional performance in flanker tasks has often been described as a colander with holes that are too large, through which not only the target, but also distractors pass unhindered (also described as “attentional leakage”; Yantis & Johnston, 1990). This means that even after resources have been concentrated (which should bring the attentional apparatus to its very best), the attentional filter remains structurally unable to filter out distractors. This description is incompatible with our results, because if the filter is structurally flawed, it should remain irrevocably flawed at all times, allowing distractors to be continually processed throughout their presentation. If so, in our experiments, incongruent distractors should have consistently delayed responses at all times, including when they were presented in later portions of a trial (i.e., the red functions in Fig. 3 should have remained above the black functions for all mutation times).
Lachter et al. (2004) presented strong and comprehensive arguments in favor of Broadbent’s core claim that identification cannot be achieved without attentional deployment, and concluded that flanker effects are a result of attentional slippage, but not leakage. The latter is defined as semantic processing of distractors despite attentional allocation of resources upon targets. That is, the attentional allocation itself is in accord with the task, but the deployed resources end up leaking toward distractors. Slippage, on the other hand, is defined as unintentional allocation of resources toward distractors, despite the task instructions.
Although slippage describes flanker interference as a circumstantial inefficiency, instead of an intrinsic deficiency (as leakage does), both slippage and leakage predict the same order of events: After attentional resources have slipped or leaked, distractor processing begins and persists. Here, the opposite was observed: Distractors were processed until attentional deployment consummated (within the initial 50 ms), after which distractors were completely ignored.
Also assuming early limited capacity, the feature integration theory (Treisman & Gelade, 1980; Treisman & Gormican, 1988) proposes that during early, preattentive stages, most physical features are processed automatically and in parallel. The percepts of these features remain floating and unconnected until attentional mechanisms have anchored them onto a map of locations; only then can the features be bound together into specific and discriminable objects. That is, the system amasses extensive perceptual information about all stimuli, but objects can only be assembled after a map of locations has been formed. If so, the targeted object can only be discriminated from irrelevant objects after attentional mechanisms have discerned between the relevant and irrelevant locations.
Assuming that representations of floating features are able to disrupt responses, the description above can accommodate the sequence of events observed in our results. The early interferences indicates that the (floating) features of all objects had already been extracted as early as 17 ms. Floating features remained unbound to geographic coordinates for an additional ~20–30 ms, until the system could discern the targeted object from nontargets and terminate distractor processing accordingly (between 34 and 51 ms).
The zoom-lens metaphor (Eriksen & St. James, 1986; Eriksen & Yeh, 1985) is rooted in the early-selection claim that initial dispersion of resources can only attain coarse resolution, which can be improved by attentional reallocation of resources upon the targeted area (Broadbent, 1982; Eriksen & Yeh, 1985; Murphy & Eriksen, 1987; Tsal, Meiran, & Lamy, 1995). This model ascribes the mechanic attributes of zoom lenses to the attentional modulation of processing. If we assume that, in our experiments, (a) every trial began with a wide-open window, (b) that it took ~50 ms to gradually zoom in from a wide window to a narrow one that encompassed only the target, and (c) that during the zooming process there was sufficient perceptual resolution, then at least some information about distractors was accumulated during the initial ~50 ms. Once the window closed in on the target, distractors remained outside the attentional window until response. That could explain why incongruent distractors delayed responses only when they were presented within the initial 50 ms, and were completely inconsequential afterward.Footnote 10
Nevertheless, even if the zoom lens metaphor is in line with the present patterns, it should not be taken as a full model of attention, because it can only describe selection within a contiguous visual field region (as in our experiments) but cannot account for attentional deployment to simultaneous stimuli selected through a shared feature, such as movement or color (Baylis & Driver, 1992, 1993; Driver & Baylis, 1989; Prinzmetal, 1981).
While early-selection theories have attributed flanker interference to inherent failures of the attentional apparatus, the late-selection approach has interpreted it as evidence reflecting unrestricted parallel processing and full identification of unattended stimuli. The late-selection approach (Deutsch & Deutsch, 1963; Duncan, 1980; Schneider & Shiffrin, 1977) has claimed that perceptual capacity is not critically limited, such that extensive information is indiscriminately extracted from all stimuli. The attentional mechanisms responsible for selecting the target operate only later, upon postperceptual representations of the already analyzed and identified stimuli. Our results indicate that extensive information about distractors (sufficient for impairing responses) was accumulated within the initial 17 ms, approximately 30 ms before attentional mechanisms succeeded in filtering the distractors out.
On the one hand, these patterns support the claim that extensive information is extracted from all stimuli before attentional activity. On the other hand, the late-selection approach is based on the assumption that there is no perceptual bottleneck and that the discrimination of targets from distractors takes place after perceptual processing, when attentional mechanisms work on postperceptual representations. Yet the clear-cut termination of distractor processing at ~50 ms suggests that attention does indeed modulate perceptual processing and is not strictly extraperceptual (Johnston & Dark, 1982; see also Broadbent, 1982, for a review of late-selection claims and shortcomings).
Perceptual load theory (Lavie, 1995, 2005, 2010; Lavie & Tsal, 1994) has proposed a resolution for the early versus late selection debate. When a task imposes low perceptual load, target selection is easy, surplus resources remain unused, and neighboring distractors end up being processed (late selection). Alternatively, high-load tasks exhaust all available resources, and distractors remain unprocessed (early selection). According to this description, distractors are processed by surplus resources, which can only happen after targets have already been selected. This order of events is the very opposite from the one observed here. Nevertheless, assuming that our two experiments imposed a low perceptual load, the interferences observed at 17 ms corroborate the prediction that low load leads to late selection.Footnote 11
Finally, the patterns observed here may stem from unlimited perceptual processing, in which all pertinent information about all presented stimuli was gathered within the first 50 ms, followed by active inhibition of the nontarget representations and suppression of additional processing of recognized distractors (Tipper, 1985). Dual-process models of attention (Anderson & Spellman, 1995; Desimone & Duncan, 1995; Driver & Tipper, 1989; Houghton & Tipper, 1994, 1996) propose that target selection is accomplished by enhancements of target processing, accompanied by inhibition of distractor processing.Footnote 12 That is, extensive information about distractors is accumulated from stimulus onset until attentional modulations inhibit nontargets. Our manipulations could not reveal when or whether perceptual processing of targets was amplified, but the suppressions observed at ~50 ms were enforced exclusively on distractors, which means that they were first recognized as to-be-ignored objects—or at least that their locations were tagged as such, which boils down to the same idea.
In conclusion, our results indicate that distractors affected performance exclusively within the initial ~50 ms following onset, after which attentional mechanisms modulated perceptual processing by suppressing the inflow of information from distractors’ locations and sustaining processing within an overall window narrower than 1.7° of visual field. We suggest that flanker interference stems from the collection of perceptual information that precedes attentional modulations, and not from inherently flawed attentional mechanisms.
Notes
The standalone application is available upon request from the authors.
Both experiments were run on the same participants within the same experimental session.
Alpha adjustments for multiple comparisons were unnecessary here, because the comparisons within each mutation time comprised different and independent sets of data. In addition, our inferences were based on patterns of significant differences across mutation times. Alpha adjustments would be necessary if, for instance, an inference were based on the presence of one significant difference in any of the 11 mutation times (because, theoretically, for every 20 nonsignificant comparisons, we could expect one false-positive result).
At 187 ms, neutral-to-neutral trials resulted in 4.4 % errors, which, compared to the 2.2 % error rate in neutral-to-incongruent trials, seems to be a conceptually illogical outlier, rather than a genuine facilitation for neutral-to-incongruent mutations. Its removal rendered the correlation less than significant (r = −.64, p = .089).
Both experiments were run on the same participants within the same experimental session.
The lack of statistical significance at 136 ms does not suggest any kind of distractor processing at this stage, since we can see in Fig. 3b that it stems from an RT drop on incongruent-to-neutral trials and not from increased interference on neutral-to-incongruent trials.
Yet see the results of Experiment 1, in which ancillary analyses suggest that prolonged presentations of incongruent distractors beyond 50 ms may have (marginally) increased disruption.
But see Humphreys (1981) for efficient filtering out of distractors located 0.5° from the target when the target was presented immediately following the fixation sign, without a blank interval between them.
We thank Dominique Lamy for this insight.
But see Max and Tsal (2014), who manipulated perceptual load in a series of mutation experiments that resulted in convergent reversals of perceptual load predictions: That is, high-load tasks (slow RTs) consistently rendered greater interference than low-load tasks.
In contrast to single-process models that propose that target selection can be fully explained by amplifications of target processing, without the need to assume inhibitions of distractors (MacLeod, Dodd, Sheard, Wilson, & Bibi, 2003).
References
Anderson, M. C., & Spellman, B. A. (1995). On the status of inhibitory mechanisms in cognition: Memory retrieval as a model case. Psychological Review, 102, 68–100. doi:10.1037/0033-295X.102.1.68
Baylis, G. C., & Driver, J. (1992). Visual parsing and response competition: The effect of grouping factors. Perception & Psychophysics, 51, 145–162. doi:10.3758/BF03212239
Baylis, G. C., & Driver, J. (1993). Visual attention and objects: Evidence for hierarchical coding of location. Journal of Experimental Psychology: Human Perception and Performance, 19, 451–470. doi:10.1037/0096-1523.19.3.451
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10, 433–436. doi:10.1163/156856897X00357
Broadbent, D. E. (1958). Perception and communication. Elmsford: Pergamon Press. doi:10.1037/10037-000
Broadbent, D. E. (1970). Stimulus set and response set: Two kinds of selective attention. In D. I. Mostofsky (Ed.), Attention: Contemporary theory and analysis (pp. 51–60). New York: Appleton-Century-Crofts.
Broadbent, D. E. (1971). Decision and stress. London: Academic Press.
Broadbent, D. E. (1981). Selective and control processes. Cognition, 10, 53–58
Broadbent, D. E. (1982). Task combination and selective intake of information. Acta Psychologica, 50, 253–290.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222. doi:10.1146/annurev.ne.18.030195.001205
Deutsch, J. A., & Deutsch, D. (1963). Attention: Some theoretical considerations. Psychological Review, 70, 80–90. doi:10.1037/h0039515
Driver, J. (2001). A selective review of selective attention research from the past century. British Journal of Psychology, 92, 53–78. doi:10.1348/000712601162103
Driver, J., & Baylis, G. C. (1989). Movement and visual attention: The spotlight metaphor breaks down. Journal of Experimental Psychology: Human Perception and Performance, 15, 448–456. doi:10.1037/0096-1523.15.3.448
Driver, J., & Tipper, S. P. (1989). On the nonselectivity of “selective” seeing: Contrasts between interference and priming in selective attention. Journal of Experimental Psychology: Human Perception and Performance, 15, 304–314. doi:10.1037/0096-1523.15.2.304
Duncan, J. (1980). The locus of interference in the perception of simultaneous stimuli. Psychological Review, 87, 272–300. doi:10.1037/0033-295X.87.3.272
Egeth, H. E., & Yantis, S. (1997). Visual attention: Control, representation, and time course. Annual Review of Psychology, 48, 269–297. doi:10.1146/annurev.psych.48.1.269
Eriksen, C. W. (1995). The flankers task and response competition: A useful tool for investigating a variety of cognitive problems. Visual Cognition, 2, 101–118.
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16, 143–149. doi:10.3758/BF03203267
Eriksen, C. W., & St. James, J. D. (1986). Visual attention within and around the field of focal attention: A zoom lens model. Perception & Psychophysics, 40, 225–240. doi:10.3758/BF03211502
Eriksen, C. W., & Webb, J. M. (1989). Shifting of attentional focus within and about a visual display. Perception & Psychophysics, 45, 175–183. doi:10.3758/BF03208052
Eriksen, C. W., & Yeh, Y.-Y. (1985). Allocation of attention in the visual field. Journal of Experimental Psychology: Human Perception and Performance, 11, 583–597. doi:10.1037/0096-1523.11.5.583
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. H. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 52–112). San Diego: Academic Press.
Houghton, G., & Tipper, S. P. (1996). Inhibitory mechanisms of neural and cognitive control: Applications to selective attention and sequential action. Brain and Cognition, 30, 20–43. doi:10.1006/brcg.1996.0003
Humphreys, G. W. (1981). On varying the span of visual attention: Evidence for two modes of spatial attention. Quarterly Journal of Experimental Psychology, 33A, 17–31. doi:10.1080/14640748108400765
Johnston, W. A., & Dark, V. J. (1982). In defense of intraperceptual theories of attention. Journal of Experimental Psychology. Human Perception and Performance, 8, 407–421. doi:10.1037/0096-1523.8.3.407
Johnston, W. A., & Dark, V. J. (1986). Selective attention. Annual Review of Psychology, 37, 43–75. doi:10.1146/annurev.ps.37.020186.000355
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eye’s movement. In J. Long & A. Baddeley (Eds.), Attention and performance IX (pp. 187–203). Hillsdale: Erlbaum.
Kahneman, D., & Treisman, A. (1984). Changing views of attention and automaticity. In R. Parasuraman & D. R. Davies (Eds.), Varieties of attention (pp. 29–61). Orlando: Academic Press.
Kimchi, R. (1992). Primacy of wholistic processing and global/local paradigm: A critical review. Psychological Bulletin, 112, 24–38. doi:10.1037/0033-2909.112.1.24
Lachter, J., Forster, K. I., & Ruthruff, E. (2004). Forty-five years after Broadbent (1958): Still no identification without attention. Psychological Review, 111, 880–913. doi:10.1037/0033-295X.111.4.880
Lavie, N. (1995). Perceptual load as a necessary condition for selective attention. Journal of Experimental Psychology: Human Perception and Performance, 21, 451–468. doi:10.1037/0096-1523.21.3.451
Lavie, N. (2005). Distracted and confused?: selective attention under load. Trends in Cognitive Sciences, 9, 75–82. doi:10.1016/j.tics.2004.12.004
Lavie, N. (2010). Attention, distraction, and cognitive control under load. Current Directions in Psychological Science, 19, 143–148. doi:10.1177/0963721410370295
Lavie, N., & Tsal, Y. (1994). Perceptual load as a major determinant of the locus of selection in visual attention. Perception & Psychophysics, 56, 183–197. doi:10.3758/BF03213897
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review. Psychological Bulletin, 109, 163–203. doi:10.1037/0033-2909.109.2.163
MacLeod, C. M., Dodd, M. D., Sheard, E. D., Wilson, D. E., & Bibi, U. (2003). In opposition to inhibition. In B. H. Ross (Ed.), The psychology of learning and motivation (Vol. 43, pp. 163–214). San Diego: Academic Press.
Max, R., & Tsal, Y. (2014). The exact timecourse of attention: The mutations paradigm. Journal of Vision, 14, 341. doi:10.1167/14.10.341
Miller, J. (1991). The flanker compatibility effect as a function of visual angle, attentional focus, visual transients, and perceptual load: A search for boundary conditions. Perception & Psychophysics, 49, 270–288. doi:10.3758/BF03214311
Müller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary orienting of visual attention: Time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance, 15, 315–330. doi:10.1037/0096-1523.15.2.315
Murphy, D. T., & Eriksen, C. W. (1987). Temporal changes in the distribution of attention in the visual field in response to precues. Perception & Psychophysics, 42, 576–586.
Navon, D. (1977). Forest before trees: The precedence of global features in visual perception. Cognitive Psychology, 9, 353–383. doi:10.1016/0010-0285(77)90012-3
Paquet, L. (2001). Eliminating flanker effects and negative priming in the flankers task: Evidence for early selection. Psychonomic Bulletin & Review, 8, 301–306. doi:10.3758/BF03196165
Paquet, L., & Craig, G. L. (1997). Evidence for selective target processing with a low perceptual load flankers task. Memory & Cognition, 25, 182–189.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25. doi:10.1080/00335558008248231
Prinzmetal, W. (1981). Principles of feature integration in visual perception. Perception & Psychophysics, 30, 330–340.
Remington, R. W. (1980). Attention and saccadic eye movements. Journal of Experimental Psychology: Human Perception and Performance, 6, 726–744. doi:10.1037/0096-1523.6.4.726
Schneider, W., & Shiffrin, R. M. (1977). Controlled and automatic human information processing: I. Detection, search, and attention. Psychological Review, 84, 1–66. doi:10.1037/0033-295X.84.1.1
Simon, J. R. (1990). The effects of an irrelevant directional CUE on human information processing. Advances in Psychology, 65, 31–86. doi:10.1016/S0166-4115(08)61218-2
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18, 643–662. doi:10.1037/0096-3445.121.1.15
Taylor, D. A. (1977). Time course of context effects. Journal of Experimental Psychology: General, 106, 404–426. doi:10.1037/0096-3445.106.4.404
Theeuwes, J. (2010). Top-down and bottom-up control of visual selection. Acta Psychologica, 135, 77–99. doi:10.1016/j.actpsy.2010.02.006
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology, 37, 571–590. doi:10.1080/14640748508400920
Treisman, A. M. (1969). Strategies and models of selective attention. Psychological Review, 76, 282–299. doi:10.1037/h0027242
Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cognitive Psychology, 12, 97–136. doi:10.1016/0010-0285(80)90005-5
Treisman, A. M., & Gormican, S. (1988). Feature analysis in early vision: Evidence from search asymmetries. Psychological Review, 95, 15–48. doi:10.1037/0033-295X.95.1.15
Tsal, Y. (1983). On interpreting the effects of location preknowledge: A critique of Duncan. Perception & Psychophysics, 34, 297–298. doi:10.3758/BF03202960
Tsal, Y., Meiran, N., & Lamy, D. (1995). Towards a resolution theory of visual attention. Visual Cognition, 2, 313–330. doi:10.1080/13506289508401736
Yantis, S., & Johnston, J. C. (1990). On the locus of visual selection: Evidence from focused attention tasks. Journal of Experimental Psychology: Human Perception and Performance, 16, 135–149. doi:10.1037/0096-1523.16.1.135
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Figure S1
Experiment 2. Mean RT by mutation time. Comparisons between congruent distractors vs neutral distractors within the three trial types (a) Neutral-to-Neutral trials; (b) Incongruent-to-Neutral trials; (c) Neutral-to-Incongruent trials. Error bars represent one standard error and asterisks denote p < 0.05. Dashed line illustrates η 2 = .5 (PDF 70.1 kb)
Figure S2
Experiment 1 and 2. Mean RT by mutation time. Comparisons between congruent distractors vs neutral distractors within the three trial types (a) Neutral-to-Neutral trials; (b) Incongruent-to-Neutral trials; (c) Neutral-to-Incongruent trials. Error bars represent one standard error and asterisks denote p < 0.05. Dashed line illustrates η 2 = .5 (PDF 69.6 kb)
Table S1
(PDF 28 kb)
Table S2
(PDF 26 kb)
Table S3
(PDF 25 kb)
Table S4
(PDF 25 kb)
Table S5
(PDF 25 kb)
Table S6
(PDF 25 kb)
Table S7
(PDF 25 kb)
Table S8
(PDF 20 kb)
Table S9
(PDF 21 kb)
Table 10
(PDF 20 kb)
Table 11
(PDF 21 kb)
Table 12
(PDF 20 kb)
Table 13
(PDF 21 kb)
Rights and permissions
About this article

Cite this article
Max, R., Tsal, Y. The mutations paradigm: Assessing the time course of distractor processing. Atten Percept Psychophys 77, 2344–2355 (2015). https://doi.org/10.3758/s13414-015-0928-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-015-0928-z