
Abstract
The aim of the study was to assess the effect of supplementation of feed rations with increased calcium and phosphorus doses on the haematological parameters and plasma zinc (Zn), phosphorus (P), magnesium (Mg), copper (Cu), calcium (Ca), and iron (Fe) content as well as the body weight and the growth and development of the first antler in farmed fallow deer (Dama dama Linnaeus, 1758). The mean level of erythrocytes (RBC), haemoglobin (HGB), and haematocrit (HCT) was increased in the Ca- and P-supplemented group after the treatment period. The change was statistically significant (p < 0.05) in the case of RBC and HCT. The other haematological parameters (mean corpuscular volume (MCV), mean corpuscular haemoglobin (MCH), mean corpuscular haemoglobin concentration (MCHC), and platelet count (PLT)) were reduced. An increase in the Zn content was observed in the plasma of slaughtered animals. The concentration of other minerals (P, Mg, and Cu only in group II receiving a higher level of Ca and P in the feeding dose; Ca and Fe only in group I supplemented with a lower content of Ca and P in a nutritional dose) in blood plasma decreased slightly after the supplementation period and declined further after the slaughter. Noteworthy, there was a significant increase in the plasma Cu and Fe levels in group I in group II, respectively, in the post-supplementation period. No significant differences were observed in the body weight between the groups, but there was a beneficial effect of the higher Ca and P dose in the feed ration for the farmed fallow deer on the length of the first antler (p < 0.05). The antlers of animals in group II were on average 2.3 cm longer than in group I.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cervids undergo spectacular physiological changes during the year. The antler cycle begins in spring with the cast of the old set. Afterwards, a new antler grows and is cleaned from velvet at the end of summer (Suttie et al. 1991). However, it has been evidenced that the first stage of cervid nutrition can exert an effect on antler size and weight. The correlation between the skeleton size and the antler weight is one of the longest known allometric dependencies occurring in animals (Zannèse et al. 2006). The growth rate and size of the first antlers developed by young males in the second year of life depend on the quality of food provided by their mothers and by the habitat and ecological conditions existing during the winter period (García et al. 1999; Gómez et al. 2008; Janiszewski et al. 2008). Bartoš (1980) as well as Bartoš and Bubenik (2011) have shown that behavioural aspects, e.g. dominance in group and social structures or other stress factors, can also be a cause of formation of larger antlers in deer.
The length of the first antler in deer is an index trait of the ontogenic quality and a phenotypic trait that reflects the quality of nutrition (Gaspar-López et al. 2008). The first antler grows with an average rate of 1.95 ± 0.05 cm per week, which is the highest at week 14 (Gaspar-López et al. 2008). The conditions provided to a young male in the early months of life exert important effects on the final size and fitness of the adult male. Intensive antler growth (antlerogenesis) requires a supply of a large amount of calcium and phosphorus in a very short time. This demand for the red deer is ca. 100 g/day, while only ca. 34 g/day is sufficient for efficient growth of skeletal bones (Cervus elaphus Linnaeus, 1758). These animals are not able to acquire such an amount of mineral ingredients with natural nutrition (Chen et al. 2008).
The average percent mineral content in the first antler of the Iberian deer (Cervus elaphus hispanicus Hilzheimer, 1909) was shown to be 34% Ca, 31% P, and 44% K. Even the best post-weaning nutrition cannot compensate for the nutrition obtained during lactation (Landete-Castillejos et al. 2007a). It has been found that a 3-kg antler contains 536 g of calcium, 348 g of which is absorbed in the final period of antler mineralisation (Gómez et al. 2012). Landete-Castillejos et al. (2007b) proved that total antler formation requires displacement of approx. 400 g of Ca and 200 g of P from the skeleton, which means transport of even 8.4 g of Ca and 2.4 g of P on each day of the most intensive growth. This demonstrates the scale of the demand for these minerals (Landete-Castillejos et al. 2007b; Kuba 2014). Calcium is the basic component of bones and is present in all animal tissues and body fluids. In terms of the content, phosphorus is the second bone component after calcium; its deficiency causes inhibition of growth and reduction of animal productivity and fertility. These two components are highly important in Cervidae, since these animals produce new antlers every year. The period of antler growth is a time of a rich blood supply, which is a carrier of nutrients, e.g. minerals required for development of antlers (Landete-Castillejos et al. 2007b)
In winter, farm fawns are usually fed with farm feed, which can be supplemented with mineral additives. In turn, they spend summer in pastures. As in domesticated ruminants, this change in feeding can be related with a period of adaptation of rumen microflora and microfauna. This period is frequently associated with diarrhoea, which may cause changes in the haematological profile (Kováč et al. 1997). Moreover blood is a very sensitive indicator of metabolic changes in both the physiological and pathological status of animals (Weiss and Wardrop 2010). In such situations, the haematological indices facilitate immediate detection of emerging potential anomalies or changes in the organism (Neumefister et al. 2001). The influence of many intra- and extrasystemic factors may lead to disruption of the homeostatic balance in the organism, which can be reflected in distinct changes in the values of individual blood components. The aim of the study was to determine the effect of supplementation with increased doses of Ca (calcium) and P (phosphorus) in the diet on the haematological parameters and plasma content of Zn (zinc), P (phosphorus), Mg (magnesium), Cu (copper), Ca (calcium), and Fe (iron). The impact of the supplementation on the body weight as well as growth and development of the first antler in farmed fallow deer was examined in the study.
Materials and methods
Experimental design
The research was carried out at the Research Station of the Institute of Parasitology, Polish Academy of Sciences, Kosewo Górne (Region of Warmia and Mazury; Poland; 53o48’ N; 21o23’ E) in 2016–2017 (December–August). The study involved fawns born in the same year (the same age). In order to provide dose adjustment for males, the animals had to be placed in different enclosures, thus modifying the social environment. Both research groups had the same social structures and were influenced by the same stress factors. It has been demonstrated that fallow deer live in unisexual groups for most of the year (Braza et al. 1990). All animals were adapted to routine management and maintained in good health and body condition during the experiment. Handling procedures and sampling frequency were designed to reduce stress and health risk for the animals. During the experiment, no negative behaviour and anomalies were observed. No record of individual intake of feed was attempted at any stage. The 24 male fawns in the first year of life were divided into two equal groups (n = 12) receiving different nutrition for 5 months (December–April):
group I - standard nutrition supplemented with a complementary mineral feed mixture for fawns “Cielak B Plus©” from LNB (Cargill, Poland). The supplement constituted 2.5% of the standard farm nutrition - 6.5 g (Table 1),
group II - standard nutrition supplemented with a complementary mineral feed mixture for fawns “Cielak B Plus©” from LNB (Cargill, Poland). The supplement constituted 2.5% of the standard farm nutrition - 6.5 g; it was additionally enriched with monocalcium phosphate (0.9 g) to increase the Ca and P level up to 25% and 13%, respectively (Table 1).
Table 1 Composition of the dietary supplement provided to farmed fallow deer fawns
The demand for Ca and P in cervids (animals exceeding 60 kg of body weight) is 2.7 g per day and 2.2 g per day, respectively (NRC 2007). These recommendations are indispensable for animal survival. The antler mineralisation process is such a big effort for the organism (Landete-Castillejos et al. 2012) that the typical diet may be insufficient to cover the large demand for Ca and P at this time. At the beginning of the experiment, the animals from both groups were at the age of 6 months and weighed on average by half less (29.3 kg); therefore, the supplementation was adjusted. The first experimental group (I) received 1.2 g of Ca per day and 0.47 g of P per day, while the second experimental group (II) received 1.8 g of Ca per day and 0.9 g of P per day as well as standard farm nutrition. To increase the concentration of Ca and P in the nutritional dose, the animals were administered “Cielak B Plus” from LNB (Cargill, Poland) and monocalcium phosphate.
Measurements
The animals spent the winter (December–April) in a shelter situated in the wintering ground divided into two identical rooms, where the amount of feed taken was controlled. The walls of the shelter were made of horizontal wooden boards and provided protection against the adverse impact of weather conditions. In the shelter, there was a feeding rack for haylage, water containers, and feeding troughs for concentrated feed. The concentrate was provided once daily. The animals had free access to water over the entire wintering period. After the winter period, the animals stayed on the available pasture without supplementation (May–August). In European countries (e.g. the Czech Republic, Denmark, France, and Great Britain), it is common practice that weaned red or fallow deer calves are wintered indoors from September to the following spring in order to protect them from aversive weather conditions, reduce expenses for winter feeding, and encourage taming of the animals (Bartoš and Šiler 1993). The animals were provided with adequate space during the winter and grazing periods as recommended in DEFRA (2018), FEDFA (2018), and Mattiello (2009). Body weight was measured with the use of a set of MP 800 sensors coupled with a Tru-test DR 3000 weight reader. As declared by the manufacturer, the accuracy of this set is ± 1% and the minimum resolution is 100 g. To obtain additional information about the ontogeny of the young fallow deer males, the growing antlers with the pedicle (at the skull base) were measured when the development was completed. The measurements were carried out using a tape measure at the age of 14 months (17 August 2017) and the mean values of the measurement of both beams were noted.
Sampling
Blood was sampled from vena jugularis externa always at the same time (from 1 to 3 h after dawn) to avoid variations associated with circadian rhythms before the supplementation period (28 December 2016), after the supplementation period (20 April 2017), and during slaughter (17 September 2017). There was no need for sedation and it was carried out as in research Langridge (1992), García et al. (2002), and Gaspar-López et al. (2011). For haematological analyses, 5-ml volumes of blood were collected into vacuum tubes containing an anticoagulant agent (EDTA). The samples were chilled (4–8 °C) for 15 min after collection and analysed 2 or 3 h later. The analyses were carried out with the use of an automatic haematological analyser Mythic 18. The device was calibrated each time before the analysis of the samples. The blood was analysed to determine the levels of leukocytes (WBC), erythrocytes (RBC), haemoglobin (HGB), haematocrit (HCT), mean corpuscular volume (MCV), mean corpuscular haemoglobin (MCH), mean corpuscular haemoglobin concentration (MCHC), and platelets (PLT).
The blood samples (5 ml) were mixed with heparin as an anticoagulant. Plasma for analysis of the biochemical parameters was obtained by centrifugation of whole blood at 3000 rpm for 10 min in a laboratory centrifuge (MPW-350R, MPW Medical Instruments, Warsaw, Poland) at a temperature of 4 °C. After centrifugation, plasma Zn, P, Mg, Cu, Ca, Fe levels were determined using reagent kits (BioMaxima, Lublin, Poland) according to the manufacturer’s protocols and a random access biochemical analyser Metrolab 2300 GL (Metrolab SA, Buenos Aires, Argentina). Haematological parameters were determined in the first two terms (before and after the supplementation, i.e. at the beginning of antler growth in April), since slaughter exerts adverse effects and the results may not be comparable to those obtained in the other periods (Marco and Lavín 1999; Kocan et al. 1981). The plasma mineral content was determined in the three terms (before the supplementation, after the supplementation at the beginning of antler growth in April, and post slaughter - after ossification of antlers). The article is part of the research of a large scientific project consisting of several studies.
Statistical analysis
The results were subjected to statistical analysis. The analysed values were presented as a mean value and standard deviation in the case of measurable parameters and as cardinality and percentage in the case of non-measurable variables.
The normality of the distribution of variables in the analysed groups was verified with the Shapiro-Wilk test. The differences between the two groups were assessed with the Student’s t test or with the Mann-Whitney test when the former could not be applied. The comparison of the haematological results and plasma mineral concentrations before and after the supplementation as well as before the supplementation and post slaughter was performed with a Student’s t test for dependent variables or a paired-samples Wilcoxon rank test when the conditions for application of the former test were not met. The comparison of the weight results obtained in the subsequent measurements was carried out with a Friedman ANOVA analysis, and pairwise comparisons were performed with a paired-samples Wilcoxon rank test with Bonferroni correction. A significance level of p < 0.05 was assumed, which indicated the presence of statistically significant differences or correlations. The database was compiled and statistical analyses were carried out in the Statistica 9.1 software (StatSoft, Polska).
Results
In the study, many interesting relationships were observed between the haematological parameters and the content of selected minerals supplemented in the nutritional dose. There was a significant increase in RBC in the study groups in the post-supplementation period: from 11.035 × 106 μl−1 to 12.954 × 106 μl−1 in group I and from 10.355 × 106 μl−1 to 12.942 × 106 μl−1 in group II receiving the higher Ca and P dose in the diet. Similarly, the increase in HCT was higher in group II, i.e. from 37.983% to 47.200%, than in group I (from 40.481% to 48.127%). In turn, the MCHC value was reduced from 40.509 g dl−1 to 38.718 g dl−1 in group I and from 42.825 g dl−1 to 39.725 g dl−1 in group II. There were significant differences in the blood haematological parameters, i.e. in the level of RBC, HCT, and MCHC (p < 0.05), within both groups and between the groups before the winter supplementation. There were no significant differences at the end of this period (Table 2). In contrast, the content of RBC and HCT changed significantly (p < 0.05) in the post-supplementation period (Table 3). Furthermore, significant differences in the level of RBC, HGB, HCT, and MCHC were noted between both study groups before and after the supplementation. Additionally, there was a significant decline in PLT after the supplementation in group I (from 457.182 × 103 μl−1 to 378.545 × 103 μl−1, p = 0.033). In group II, there was a significant change in the WBC value (from 4.808 × 103 μl−1 to 4.214 × 10 3 μl−1, p < 0.001) and MCH (from 15.875 pg to 14.458 pg, p = 0.014) (Table 4).
The plasma mineral content differed significantly between the study groups in the pre-supplementation period in the case of Zn (35.821 μmol L−1 in group I and 45.600 μmol L−1 in group II, p = 0.001), Mg (0.976 mmol L−1 in group I and 0.825 mmol L−1 in group II, p < 0.001), Cu (14.694 μmol L−1 in group I and 13.964 μmol L−1 in group II, p = 0.018), and Fe (41.695 μmol L−1 in group I and 32.005 μmol L−1 in group II, p = 0.003). After the supplementation period, significant differences were observed only in the case of Cu (20.780 umol L−1 in group I and 11.953 umol L−1 in group II, p = 0.035). In turn, there were differences in the levels of Zn (57.783 μmol L−1 in group I and 49.650 μmol L−1 in group II, p = 0.001) and P (1.640 mmol L−1 in group I and 1.838 mmol L−1 in group II, p = 0.038) in the post-slaughter period following the summer grazing period. Additionally, the plasma Zn content was increased in the post-slaughter period (by 21.9–22.1 μmol L−1 in group I and by only 4.05–14.5 μmol L−1 in group II). The content of other minerals (P, Mg, and Cu only in group II; Ca and Fe only in group I) in the blood plasma in the post-supplementation period decreased slightly and was even lower after the slaughter. Noteworthy, there was a significant increase in the plasma levels of Cu (from 14.694 μmol L−1 to 20.780 μmol L−1) in group I and Fe (from 32.005 μmol L−1 to 34.475 μmol L−1) in group II after the supplementation period (Table 5). Both groups showed significant changes in the P and Cu content in the post-supplementation period and in the Zn, Mg, and Ca levels after the slaughter (Table 6).
Significant changes in the plasma P and Mg content were demonstrated in both groups in the pre- and post-supplementation periods and in the post-slaughter period. Additionally, there was a significant change in the plasma Zn and Cu levels in group I between the post-supplementation and post-slaughter periods. The content of Ca and Fe in the plasma changed significantly in group I in the pre- and post-supplementation periods as well as the post-supplementation and post-slaughter periods. The group receiving the higher dose of Ca and P in the feed ration (II) exhibited a significant change in the plasma Zn level after the supplementation (Table 7).
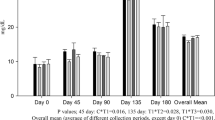
The determination of the effects of the supplementation on blood parameters was accompanied by an analysis of its impact on the body weight and the length of the first antler in the farmed fallow deer. There were no significant differences in the body weight of the animals between the groups, whereas the increased Ca and P content in the feed ration was shown to exert a significant positive effect on the first antler length (p < 0.05). The animals from group II developed on average 2.3 cm longer antlers than the individuals in group I (Online Resource: Table A1). Additionally, there were significant correlations in the case of the body weight in both groups of the farmed fallow deer in almost all the terms of weighing, except for term 2 and 3 (Online Resource: Table A2, A3).
Discussion
There are few studies on the effect of supplementation on haematological parameters and mineral concentration in the plasma of farmed fallow deer. This type of research is important for breeders since this species is the most commonly bred Cervidae in Poland (FEDFA 2018). Commercial deer breeding in Poland began in 2002. As demonstrated by data presented in 2014 by the Polish Union of Deer Breeders at the FEDEFA congress, it can be assumed that there are 6000–9000 red deer and 25000–30000 fallow deer in Poland (Janiszewski et al. 2014).
In the present experiments, a decline in the WBC count was observed in both groups in the spring (after the supplementation period). In group II supplemented with the increased amounts of Ca and P, this parameter had higher values, i.e. 0.6 × 103 μl−1, than in group I (0.3 × 103 μl−1). This may indicate a higher rate of antler growth in the fallow deer, which is in agreement with the findings reported by Gaspar-López et al. (2011), who showed that the size of antlers was negatively correlated with the WBC count in cervids’ blood. As shown by their results, larger antler sizes were found in animals that had lower values of the WBC count, MCHM, and MPV (Gaspar-López et al. 2011; Weiss and Wardrop 2010). The WBC and antler score were negatively correlated. This negative relationship between the leukocyte count and antler size may be explained by the theory proposed by Coop and Kyriazakis (1999) that the requirements of the immune system are only covered after the needs of maintenance of body protein or reproduction. Another possible explanation is the weaker condition of the first defence line in animals with smaller antlers, as suggested by Landete-Castillejos et al. (2002). This would be associated with a higher expense in the immune system, reducing the amount of resources available for antler growth (Landete-Castillejos et al. 2007b).
The mean leukocyte count was slightly higher than that reported by Mohri et al. (2000) in Persian fallow deer (Dama mesopotamica Brooke, 1875) and Chapman and Chapman (1982) in fallow deer (D. dama), but it was the same as the result presented by English and Lepherd (1981), Zomborsky et al. (1997), and Kováč et al. (1997). The differences may have been related to the method applied for immobilisation of the animals during blood collection. The use of physical methods, as in the case of farm fallow deer, may result in elevated leukocyte counts, in comparison with chemical methods. As shown by Tomkins et al. (1991), the diet may be another cause of reduced WBC and elevated RBC counts. In the present study, there was an increase in the mean number of erythrocytes in the post-supplementation period; a higher increase, i.e. by 2.6 × 106 μl−1, was observed in group II. Increased blood platelet counts are observed in cervids in spring (Gaspar-López et al. 2011); however, the value of this parameter measured in the experimental animals in April was reduced by 78.6 × 103 μl−1 in group I and by 69.2 × 103 μl−1 in group II. The increase in the haematological parameters as well as haemoglobin and haematocrit may support the positive effect of supplementation with an increased amount of Ca and P in the diet.
The mean number of erythrocytes in the fallow deer blood was similar to the value reported by Mohri et al. (2000) as well as Chapman and Chapman (1982). Similarly, the level of haemoglobin was similar to that shown by Mohri et al. (2000), Barić Rafaj et al. (2011), Chapple et al. (1991), and Cross et al. (1994) and slightly higher than that reported by Rosef et al. (2004). The value of haematocrit was higher in the farmed fallow deer in the post-supplementation period and similar in the pre-supplementation period to the values reported by Mohri et al. (2000), Barić Rafaj et al. (2011), Chapple et al. (1991), and Cross et al. (1994). The MCV parameter was slightly lower in the farmed fallow deer than that shown by Cross et al. (1994) and Rosef et al. (2004) and higher than the value demonstrated by Chapple et al. (1991). The MCH value was lower than in Persian fallow deer (Mohri et al. 2000), similar to that reported by Barić Rafaj et al. (2011) and Cross et al. (1994), and higher than in investigations conducted by Chapple et al. (1991). The MCHC parameter exhibited higher values than those presented by Mohri et al. (2000) or those determined in other cervid species by Audigé (1992), Barić Rafaj et al. (2011), Chapple et al. (1991), and Cross et al. (1994). The mean number of platelets was higher than the value reported from Persian fallow deer (Mohri et al. 2000) or shown by Barić Rafaj et al. (2011); yet, it was in the range reported by Cross et al. (1994). These seasonal changes in the haematological parameters may have been associated with the changes in the diet, as demonstrated by DelGuidice et al. (1992). Nevertheless, it should be borne in mind that haematological parameters usually have higher values in young animals up to 18 months of age (Weiss and Wardrop 2010).
Besides haematological parameters, the content of some minerals in the blood plasma was analysed. The highest concentration of most minerals (P, Mg, Cu only in group II, Ca and Fe only in group I) was detected in the first period, i.e. before the supplementation was included in the youngest animals. This can be explained by the high activity of osteoblasts in calves in bone tissue remodelling (Rosef et al. 2004; Kučer et al. 2013). Additionally, the slight decrease in the plasma mineral content in the post-supplementation period may have been caused by the change in the diet and the onset of the antler development period. However, in contrast to the present research, Graham et al. (1962) as well as Morris and Bubenik (1983) showed that the concentration of minerals in the plasma remained stable during antler growth.
The increase in the Zn concentration in the plasma of the farmed fallow deer observed in the post-slaughter period, i.e. after cleaning off the velvet and ossification of antlers, may have been related to the approaching mating season. Zn is responsible for normal testosterone concentrations and proper function of the immune system (Weiss and Wardrop 2010; Bartoskewitz et al. 2007). After antler growth and mating periods, the immune system of cervids can return to homeostasis, hence the increased uptake of this mineral in the blood (Kun et al. 2015). This may also result from the supplementation, as in the investigations conducted by Suresh et al. (2013).
Scharfe et al. (1998) and Chapple et al. (1991) found a P concentration of 2.40 mmol L−1 in neonatal fallow deer. A similar concentration of this mineral was determined in the plasma of the farmed fawns in the pre-supplementation period. The value declined in the subsequent terms with the age of animals. There were no significant differences between the study groups despite the different P content in the diet. In other studies, the mean concentration of P at unrestricted access was 3.20 mmol L−1 in a population of young red deer and 1.48 mmol L−1 in adult animals. The decline with age is understandable, as the P demand is higher during early growth (Kučer et al. 2013).
It is known that the Mg concentration decreases with age due to stress susceptibility, defective membrane functions, and disruption of intracellular calcium metabolism, inflammation, cardiovascular diseases, including atherosclerosis and ischemic injury, diabetes, fibrosis, immune dysfunction, and other diseases associated with aging (Rayssiguier et al. 1993). The Mg concentration in the farmed fallow deer was similar to that reported by Kučer et al. (2013), and a significant decline in this parameter was noted after the grazing period without supplementation.
Cu deficiency is common in farmed red deer livestock. Padilla et al. (2000) and Bao et al. (2010), however, detected low serum Cu (9.86 μmol L−1) values in young red deer kept on pasture. Values lower than 8 μmol L−1 are below the critical level for Cu deficiency (Mackintosh et al. 1987). Probably, the distinct decline in the plasma Cu content noted in the post-slaughter period in the farmed fallow deer was caused by the preceding pasture period, which is sometimes characterised by deficiencies of this mineral in pasture-kept animals. In turn, the study conducted by Rosef et al. (2004) showed a mean Cu value of 13.0 μmol L−1. A similar level of this element was noted in the farmed fallow deer in the pre-supplementation period before the winter.
The mean values of Ca, Mg, and P in the fallow deer blood were higher than the concentrations described by Marco and Lavín (1999) and Rosef et al. (2004). This confirms the positive effect of the supplementation applied during the winter period. However, the Ca to Mg ratio is important in the absorption, utilisation, and excretion of these minerals (Somer 1995). Magnesium and calcium act in conjunction; for instance, they are involved in the regulation of nerve and muscle tone. A high Ca2+/Mg2+ ratio also predisposes to arterial spasms and increases catecholamine release (Sheehan and Seelig 1984). Probably, the changes in the Fe concentration in the blood of the group I fallow deer were induced by an unfavourable Ca/Mg ratio in the diet. Evidently, the group II animals received a better-balanced and more beneficial diet containing an increased content of Ca and P in the feed ration. At a proper and constant level of nutrition, the concentration of Ca and P in the blood does not change despite the period of antler growth (NRC 2007; Gaspar-López et al. 2011).
Proper nutrition is especially important in Cervidae calves, whose survival of the first winter is determined by achievement of normal body weight (Fennessy et al. 1991). A comparison of the effect of supplementation of feed for farmed Iberian red deer (C. elaphus hispanicus) demonstrated a significant difference in biometric parameters between the control group and the group of animals receiving enriched diets (Olguin et al. 2013). The present study showed no effect of the supplementation on the body weight in the analysed fallow deer. In other studies of farmed fallow deer, fawns that spent winter with does were characterised by an average 5.43% decline in body weight, in comparison with fawns wintered under a shelter - 10.24% (Janiszewski et al. 2008). Appropriate conditions provided to animals during the first winter are essential, as confirmed by the above-mentioned experiments and the present study, which demonstrated a positive effect of the supplementation on the body weight and antler length. Moreover, the final antler size was negatively related to the leukocyte count and the body condition had a more important effect on the final antler size, which was bigger than that exerted by the body weight, as also confirmed by Gaspar-López et al. (2011). However, one cannot forget that the size of antlers can also be influenced by animal behaviour and their social structure (Bartoš 1980; Bartoš and Bubenik 2011).
The increased Ca and P supply contributed to elevation of the plasma Zn content and improved some haematological indices, e.g. RBC, HGB, and HCT. Moreover, the increased doses of Ca and P in the diet exerted a positive effect on the concentration of minerals in the blood of cervids. Both animal groups exhibited a significant increase in the plasma Cu and Fe levels in group I in the post-supplementation period. Thus, the supplementation of the fawn nutrition had a positive effect on animal fitness and antler growth.
References
Audigé L (1992) Haematological values of rusa deer (Cervus timorensis russa) in New Caledonia. Aust Vet J 69(11):265–268. https://doi.org/10.1111/j.1751-0813.1992.tb09886.x
Bao K, Li GY, Cui XZ (2010) Effects of different copper sources on serum biochemical parameters and nutrient digestibilities in growing male sika deer (Cervus nippon). Chinese J Anim Nutr 22:717–722
Barić Rafaj R, Tončić J, Vicković I, Šoštaric B (2011) Haematological and biochemical values of farmed red deer (Cervus elaphus). Vet Arhiv 81(4):513–523
Bartoš L (1980) The date of antler casting, age and social hierarchy relationships in the red deer stag. Behav Proc 5:293–301. https://doi.org/10.1016/0376-6357(80)90013-3
Bartoš L, Bubenik GA (2011) Relationships between rank-related behaviour, antler cycle timing and antler growth in deer: behavioural aspects. Anim Prod Sci 51:303–310
Bartoš L, Šiler J (1993) Survey of game farming in Europe. FAO Ed., Roma, Italy
Bartoskewitz ML, Hewitt DG, Laurenz JC (2007) Effect of dietary copper and zinc concentrations on white-tailed deer antler growth, body size, and immune system function. Small Rumin Res 73:87–94. https://doi.org/10.1016/j.smallrumres.2006.11.005
Braza F, San Jose C, Blom A, Cases V, García JE (1990) Population parameters of fallow deer at Donana National Park (SW Spain). Acta Theriol 35(3–4):277–288
Chapman DI, Chapman NG (1982) Some haematological data for fallow deer (Dama dama) in England. Res Vet Sci 33:205–207
Chapple RS, English AW, Mulley RC, Lepherd EE (1991) Haematology and serum biochemistry of captive unsedated chital deer (Axis axis) in Australia. J Wildl Dis 27(3):396–406. https://doi.org/10.7589/0090-3558-27.3.396
Chen PY, Lin AY, M, Lin YS, Seki Y, Stokes AG, Peyras J, Olevsky EA, Meyers MA, McKittrick J (2008) Structure and mechanical properties of selected biological materials. J Mech Behav Biomed Mater 1(3):208–226. doi:https://doi.org/10.1016/j.jmbbm.2008.02.003
Coop RL, Kyriazakis I (1999) Nutrition-parasite interaction. Vet Parasitol 84:187–204 https://doi.org/10.1016/S0304-4017(99)00070-9
Cross JP, Mackintosh CG, Griffin JFT (1994) Haematological reference values for farmed red deer (Cervus elaphus) in New Zealand. Comp Haematol Int 4:76-85
DEFRA (2018) Code of recommendations for the welfare of farmed deer. (http://www.defra.gov.uk/animalh/welfare/farmed/othersps/ deer/pb0055/deercode.htm). Accessed 24 April 2018
DelGuidice GD, Mech LD, Kunkel KE, Gese EM, Seal US (1992) Seasonal patterns of weight, hematology and serum characteristics of free-ranging female white-tailed deer in Minnesota. Can J Zool 70:974–983. https://doi.org/10.1139/z92-139
English AW, Lepherd EE (1981) The hematology and serum biochemistry of wild fallow deer (Dama dama) in New South Wales. J Wildl Dis 17:289–295
FEDFA (2018) The Federation of European Deer Farmers Associations (https://www.fedfa.com/en/fedfa-members/#1364). Accessed 10 November 2018
Fennessy PF, Thomson JM, Suttie JM (1991) Season and growth strategy in red deer. Evolutionary implications and nutritional management. In: Wildlife production: conservation and sustainable development. University of Alaska, Fairbanks, pp 495–501
García A, Landete-Castillejos T, Molina A, Albiñana B, Fernández C, Garde J, Gallego L (1999) Lactation curves in captive Iberian red deer (Cervus elaphus hispanicus). J Anim Sci 77:3150–3155. https://doi.org/10.2527/1999.77123150x
García AJ, Landete-Castillejos T, Garde JJ, Gallego L (2002) Reproductive seasonality in female Iberian red deer (Cervus elaphus hispanicus). Theriogenology 8656:1–10
Gaspar-López E, García AJ, Landete-Castillejos T, Carrion D, Estévez JA, Gallego L (2008) Growth of the first antler in Iberian red deer (Cervus elaphus hispanicus). Eur J Wildl Res 54:1–5. https://doi.org/10.1007/s10344-007-0096-0
Gaspar-López E, Landete-Castillejos T, Estevez JA, Ceacero F, Gallego L, García AJ (2011) Seasonal variations in red deer (Cervus elaphus) hematology related to antler growth and biometric measurments. J Exp Zool 315:242–249. https://doi.org/10.1002/jez.670
Gómez JA, Landete-Castillejos T, García AJ, Gaspar-López E, Estévez JA, Gallego L (2008) Lactation growth influences mineral composition of first antler in Iberian red deer (Cervus elaphus hispanicus). Wildl Biol 14:331–338. https://doi.org/10.2981/0909-6396(2008)14[331:LGIMCO]2.0.CO;2
Gómez JA, Ceacero F, Landete-Castillejos T, Gaspar-López E, García AJ, Gallego L (2012) Factors affecting antler investment in Iberian red deer. Anim Prod Sci 52:867–873. https://doi.org/10.1071/AN11316
Graham EA, Rainey R, Kuhlman RE, Houghton EH, Moyer CA (1962) Biochemical investigations of deer antler growth: part I. alterations of deer blood chemistry resulting from Antlerogenesis. J Bone Joint Surg 44(3):482–488
Janiszewski P, Dmuchowski B, Gugołek A, Żełobowski R (2008) Body weight characteristics of farm-raised fallow deer (Dama dama L.) over the winter period. JCEA 9(2):337–342
Janiszewski P, Bogdaszewska Z, Bogdaszewski M, Bogdaszewski P, Ciululko-Dołęga J, Nasiadka P, Steiner Ż (2014) Rearing and breeding of cervids. Publishing house UWM, Olsztyn. [in Polish]
Kocan AA, Glenn BL, Thedford TR, Doyle R, Waldrup K, Kubat G, Shaw MG (1981) Effects of chemical immobilization on hematologic and serum chemical values in captive white-tailed deer. J Am Vet Med Assoc 179:1153–1156
Kováč G, Ciberej J, Paulikova I (1997) Haematological indices in fallow deer. Acta Vet Brno 66:203–211
Kuba J (2014) Analysis of changes in the concentration of ca, P and mg in blood serum of red deer (Cervus elaphus) immature males in farm breeding. Acta Sci Pol Zootech 13(2):31–40
Kučer N, Kuleš J, Barić Rafaj R, Tončić J, Vicković I, Štokovic D, Potočnjak D, Šoštaric B (2013) Mineral concentrations in plasma of young and adult red deer. Vet Arhiv 83(4):425–434
Kun B, Weili S, Chunyi L, Kaiying W, Zhipeng L, Shidan B, Guangyu L (2015) Effects of dietary zinc supplementation on nutrient digestibility, haematological biochemical parameters and production performance in male sika deer (Cervus nippon). Anim Prod Sci 56(6):997–1001. https://doi.org/10.1071/AN15039
Landete-Castillejos T, García AJ, Gómez JA, Laborda J, Gallego L (2002) Effects of nutritional stress during lactation on immunity costs and indices of future reproduction in Iberian red deer (Cervus elaphus hispanicus). Biol Reprod 67:1613–1620
Landete-Castillejos T, García A, Gallego L (2007a) Body weight, early growth and antler size influence antler bone mineral composition of Iberian red deer (Cervus elaphus hispanicus). Bone 40(1):230–235. https://doi.org/10.1016/j.bone.2006.07.009
Landete-Castillejos T, Estévez JA, Martínez A, Ceacero F, García AJ, Gallego L (2007b) Does chemical composition of antler bone reflect the physiological effort made to grow it? Bone 40:1095–1102. https://doi.org/10.1016/j.bone.2006.11.022
Landete-Castillejos T, Estevez AJ, Ceacaro F, García AJ, Gallego L (2012) A review of factors affecting antler composition and mechanics. Front Biosci 4:2328–2339. https://doi.org/10.2741/E545
Langridge M (1992) Establishing a fallow deer farm: basic principles. In: Asher GW (ed) Proc 1st world forum on fallow deer farming, Mudgee, pp 79–94
Mackintosh CG, Wilson PR, Beatson NS, Turner K, Johnstone P (1987) Preliminary report of the liver/serum copper relationship in red deer. In: Proceedings of a deer course for veterinarians. Rotura 1986. New Zealand veterinary association, Wellington, pp 156–157
Marco I, Lavín S (1999) Effect of the method of capture on the haematology and blood chemistry of red deer (Cervus elaphus). Res Vet Sci 66:81–84. https://doi.org/10.1053/rvsc.1998.0248
Mattiello S (2009) Welfare issues of modern deer farming. Ital J Anim Sci 8:205–217. https://doi.org/10.4081/ijas.2009.s1.205
Mohri M, Aslani MR, Shahbazian N (2000) Haematology of persian fallow deer (Dama mesopotamica). Comp Haematol Int 10:183–186. https://doi.org/10.1007/s005800170001
Morris JM, Bubenik GA (1983) Seasonal levels of minerals, enzymes,nutriens and metabolic products in plasma of intact and castrated adult male white-tailed deer (Odocoileus virginianus). Comp Biochem Physiol A Comp Physiol 74A(1):21–28
Neumefister B, Besenthal I, Liebich H (eds) (2001) Laboratory diagnostics - clinical guide. Elsevier Urban & Partner, Warszawa. [in Polish]
NRC (2007) National Research Council Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids. The National Academies Press, Washington, DC
Olguin CA, Landete-Castillejos T, Ceacero F, García AJ, Gallego L (2013) Effects of feed supplementation on mineral composition, mechanical properties and structure in femurs of Iberian red deer hinds (Cervus elaphus hispanicus). PLoS One 8(6):e65461. https://doi.org/10.1371/journal.pone.0065461
Padilla S, Bouda J, Quiroz-Rocha GF, Davalos JL, Sanches A (2000) Biochemical and haematological values in venous blood of captive red deer (Cervus elaphus) at high altitude. Acta Vet Brno 69:327–331. https://doi.org/10.2754/avb200069040327
Rayssiguier Y, Durlach J, Gueux E, Rock E, Mazur A (1993) Magnesium and ageing. I. Experimental data: importance of oxidative damage. Comp Haematol Int 6:369–378
Rosef O, Nystøyl HL, Solenes T, Arnemo JM (2004) Haematological and serum biochemical reference value in free-ranging red deer (Cervus elaphus atlanticus). Rangifer 24(2):79–85. https://doi.org/10.7557/2.24.2.304
Scharfe ST, Wittek K, Elze J, Gruhle W, Hock (1998) Untersuchungen zu hämatologischen sowie klinisch-chemischen Parametern im Blutserum neugeborener Damwildkälber (Dama dama L.). Teil 1: Geburtsgewichte und hämatologische Parameter. Tierärztl Prax 26:97–103
Sheehan JP, Seelig MS (1984) Interactions of magnesium and potassium in the pathogenesis of cardiovascular disease. Magnesium 3:301–314
Somer E (1995) Minerals. In: Somer E (ed) the essential guide to vitamins and minerals. NY Harper Perennial, New York
Suresh C, Das A, Katole S, Saini M, Swarup D (2013) Effect of concentrate supplementation on feed consumption, nutrient utilization and blood metabolite profile in captive spotted deer (Axis axis) fed oat (Avena sativa) and berseem (Trifolium alexandrinum) fodders based diet. Zoobiology 32(2):195–203. https://doi.org/10.1002/zoo.21058
Suttie JM, Fennessy PF, Crosbie SF, Corson ID, Lass FJ, Elgar HJ, Lapwood KR (1991) Temporal changes in LH and testosterone and their relationships with the firs antler in red deer (Cervus elaphus) stags from 3 to 15 months of age. J Endocrinol 131:467–474. https://doi.org/10.1677/joe.0.1310467
Tomkins NW, McMeniman NP, Daniel RCW (1991) Voluntary feed intake and digestibility by red deer (Cervus elaphus) and sheep (Ovis ovis) of pangola grass (Digitaria decumbens) with or without a supplement of leucaena (Leucaena leucocephala). Small Ruminant Res 5(4):337–345. https://doi.org/10.1016/0921-4488(91)90071-W
Weiss DJ, Wardrop KJ (2010) Schalm’s veterinary hematology. Sixth Edn. Blackwell Publishing Ltd, Iowa
Zannèse A, Morellet N, Targhetta C, Coulon A, Fuser S, Hewison AJM, Ramanzin M (2006) Spatial structure of roe deer populations: towards defining management units at a landscape scale. J Appl Ecol 43:1087–1097. https://doi.org/10.1111/j.1365-2664.2006.01229.x
Zomborsky Z, Horn E, Tuboly Gyodi P (1997) Some haematological and immunological parameters of farmed deer in Hungary. Acta Vet Hung 45:75–84
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical statement
This study was carried out in strict accordance with the Polish legislation for the use of animals in research (Act of 15 January 2015 on the protection of animals used for scientific or educational purposes). The protocol was approved by the Local Ethics Committee 0069, Resolution No. 42/2016 at the University of Warmia and Mazury in Olsztyn. Animals were restrained using a cushioned crush specifically designed to restrain deer movements - a handling box (2 m × 2 m × 0.6 m) only for a few minutes. At that time, the fallow deer were weighed, the blood was collected, and the antlers were measured.
Conflict of interest
No potential conflicts of interest were reported by the authors.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 27.1 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Tajchman, K., Steiner-Bogdaszewska, Ż., Kowalczuk-Vasilev, E. et al. Effect of Ca and P supplementation on the haematological parameters and content of selected minerals in the blood of young farmed fallow deer males (Dama dama). Biologia 75, 401–411 (2020). https://doi.org/10.2478/s11756-019-00310-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2478/s11756-019-00310-2