
Abstract
Deterioration of diaphragm function is one of the prominent factors that contributes to the susceptibility of serious respiratory infections and development of respiratory failure in patients with Duchenne Muscular Dystrophy (DMD). The NF-κB signaling pathway has been implicated as a contributing factor of dystrophic pathology, making it a potential therapeutic target. Previously, we demonstrated that pharmacological inhibition of NF-κB via a small NEMO Binding Domain (NBD) peptide was beneficial for reducing pathological features of mdx mice. Now, we stringently test the effectiveness and clinical potential of NBD by treating mdx mice with various formulations of NBD and use diaphragm function as our primary outcome criteria. We found that administering DMSO-soluble NBD rescued 78% of the contractile deficit between mdx and wild-type (WT) diaphragm. Interestingly, synthesis of a GLP NBD peptide as an acetate salt permitted its solubility in water, but as a negative consequence, also greatly attenuated functional efficacy. However, replacing the acetic acid counterion of the NBD peptide with trifluoroacetic acid retained the peptide’s water solubility and significantly restored mdx diaphragm contractile function and improved histopathological indices of disease in both diaphragm and limb muscle. Together, these results support the feasibility of using a mass-produced, water-soluble NBD peptide for clinical use.
Similar content being viewed by others


Introduction
Duchenne muscular dystrophy (DMD) is a deadly genetic disease characterized mainly by progressive weakness of the skeletal musculature. Loss of ambulation due to skeletal muscle degeneration is the main recognizable phenotypical feature of patients with DMD. However, more life-threatening complications develop as the disease progresses, with eventual cause of death in nearly all patients as a result of diaphragm and/or cardiac muscle failure (1).
Treatment options for DMD patients are limited with virtually none currently effective for ameliorating contractile dysfunction, including that of the diaphragm. However, we demonstrated recently that NF-κB inhibition improved contractility of diaphragm muscle in mdx mice, a widely used mouse model of DMD (2). Those data warranted further investigation into the possibility of therapeutically targeting the NF-κB signaling pathway as a means of treating dystrophic pathology. Chronic activation of the NF-κB signaling pathway has been described in DMD patients (2,3) and this activity occurs very early in the timescale of the pathology (4). NF-κB was further demonstrated to promote disease progression in mdx mice (2,5,6). Although NF-κB inhibition has yet to be tested in DMD patients (7), the current standard care of using glucocorticoids exert their beneficial effects partially through the inhibition of NF-κB (8,9). Thus, NF-κB therapy may be considered beneficial for the treatment of DMD.
Classical inflammatory NF-κB signaling occurs in response to stimuli such as TNFα and is mediated through the upstream IκB kinase (IKK) complex (10). IKK consists of two catalytic kinase sub-units, IKKα and IKKβ, and a regulatory subunit named IKKγ (or NEMO for NF-κB Essential Modulator) (11). Inactive NF-κB heterodimers composed of p65-p50 are maintained predominately in the cytoplasm by inhibitor IκB proteins. Phosphorylation of IκB by IKK signals ubiquitination and subsequent degradation of IκB by the 26S proteasome, thus allowing p65-p50 to localize to the nucleus, bind DNA, and cooperate with basal transcription factors and coactivators to induce transcription of target genes (12).
An IKK complex containing NEMO is critical for inflammatory signaling through the classical NF-κB pathway (13,14). When the association between IKKα and IKKβ and NEMO is disrupted, inflammatory signaling through IKK is impeded. In our previous investigation, we took advantage of this critical association to inhibit classical NF-κB signaling. We demonstrated that a cell-permeable 11 amino acid peptide that binds to the C-terminal region within IKKα and IKKβ and prevents association with NEMO, called the NEMO Binding Domain (NBD) peptide, blunts the chronic NF-κB signaling in mdx mice (2). This peptide effectively ameliorated pathology in the muscles of mdx mice by reducing inflammation and enhancing regeneration, and improved functional deficits in the diaphragm significantly when compared with administration with a mutant version of NBD.
In addition to NBD, other anti-NF-ºB pharmacological and biological agents (5,15–21) have been shown to be efficacious in rescuing mdx pathology to varying degrees. However, unknown at this point is whether any of these agents are suitable for development into clinically useable drugs. We reasoned that such a compound would need to exhibit significant functional improvement to the diaphragm since this is the muscle in mdx mice that most closely recapitulates the dystrophic process in DMD patients (22). We are mindful that the amount of improvement needed in a rodent model to increase muscle strength or quality of life in a DMD patient is difficult to predict because of in-traspecies differences. Nevertheless, in an effort to move toward our goal of determining whether NBD can be developed as a therapeutic for the treatment of DMD, we performed this study to assess doses and formulations of NBD that are efficacious for improving diaphragm and limb muscle function in the mdx mouse.
Materials and Methods
Mice
Mice (mdx [C57BL/10 ScSn DMDmdx]) purchased from The Jackson Laboratory (Bar Harbor, ME, USA) were housed and bred in the animal facilities of The Ohio State University College of Medicine, under conventional conditions with constant temperature and humidity and fed a standard diet. Both male and female mdx mice were used for these studies. Animal experimentation was approved by The Ohio State University Institutional Animal Care and Use Committee.
Peptide Synthesis
The original NBD peptide (TALDW SWLQTE), fused to an Antennapedia protein transduction domain (23), was generated by using an ABI 430A solid-phase peptide synthesizer (Applied Biosystems, Foster City, CA, USA) with standard tBOC (tert-butyloxycarbonyl)-chemistry (Yale University, New Haven, CT, USA). The peptide was cleaved from the resin and deprotected by using hydrofluoric acid. Resulting crude materials were purified by HPLC on a Vydac (Vydac Separations, Hesperia, CA, USA) C18 preparative column by using gradients of acetonitrile in 0.10% trifluoroacetic acid (TFA). Following lyophilization of the purified fractions, the expected molecular weight of the peptide was confirmed by using matrix-assisted laser desorption ionization mass spectrometry. The resultant peptide was soluble in DMSO, the same as used by our group previously (2).
GLP grade NBD peptide was produced by American Peptides (Sunnyvale, CA, USA) by using similar procedures to the original NBD peptide except with an additional purification step in which gradients of acetate rather than TFA were used to elute peptide from the column. This form is called GLP-NBD.
For conversion of NBD from an acetate salt to TFA, peptide was dissolved in a minimum amount of 1:1 acetonitrile/water. For every 10 mg of peptide used, 18 εL of TFA was added and mixed. The solution then was frozen by using a dry ice and isopropanol mixture and subsequently lyophilized. The peptide was re-suspended with an acetronitrile/water mixture and TFA, and lyophilization steps were repeated an additional two times. This procedure maintained NBD as a water-soluble peptide, which we refer to as TFA-NBD.
Treatment Regimen
Mdx mice between 3.5 to 4 wks of age were treated with peptides at a dose of 40 or 200 εg/mouse 3x weekly by IP injections for a period of 4 wks. Although exact delivery varied slightly, these doses provided an average of 2 or 10 mg/kg, respectively, over the course of the 4-wk treatment period. Vehicle control mdx mice were dosed in a similar manner except, rather than peptide, only the vehicle (10% DMSO/90% saline) was injected. Aged matched wild-type (WT) C57/BL10 mice were included as controls. For GLP synthesized peptides (GLP-NBD and TFA-NBD), water was used as the vehicle.
Diaphragm Force Measurements
After anesthetizing the mice (urethane, 200 mg in 0.7 mL i.p.), the thorax was opened, and the diaphragm, attached to part of the ribcage, was removed. 1.2- to 1.9-mm-wide sections (with a length from rib to tendon) of diaphragm were isolated in presence of BDM to minimize cutting damage (24), and attached to a force transducer (KG7, Scientific Instruments, Heidelberg, Germany) and a micrometer screw that allows for the adjustment of muscle length. All contraction experiments were done at 37° C. Although this temperature deviates from previously established guidelines, it better reflects the physiological temperature for in vivo diaphragm contraction, and thus maximizes extrapolation of the data obtained to the physiological in vivo situation. Moreover, this temperature is inline with our own previous studies (2,25,26), and similar/identical experiments have been done by other laboratories (27–29), indicating feasibility of body-temperature experiments. The muscle was then stretched to the length where twitch contractions are optimal, and allowed to rest for 10 min. Thereafter, they were subjected to a protocol consisting of a series of eight tetanic contractions occurring at 2-min intervals, each with duration of 100 ms with paired-pulse trains of stimuli at 20, 50, 80, 120, 150, 180, 200, and 250 Hz respectively. Force measurements were normalized to cross-sectional area. All assessments and data analysis were done with the investigator blind to the treatment of the mice. To further reduce variability of muscle measurements that are inherent to the preparation, time/personnel permitting, two strips per mouse were measured, and the data was averaged per mouse for inclusion as a single measurement point in the statistical analysis. Following removal of the diaphragm strips, the remainder of the diaphragm muscle was prepared for histological and biochemical analysis.
Whole Body Functional Measurements
Mice were subjected to an exercise regimen that included five successive days of treadmill running at a speed of 25 meters/min at a 15% incline for 35 min. Functional measurements were then obtained by performing rotarod strength/endurance tests where the time of latency to fall was recorded at 25 rpm for 180 seconds.
Assessment of Peptide Stability
Aliquots of NBD either were freshly prepared or stored at different temperatures for different times, and subsequently analyzed by mass spectrometry. LC-MS/MS was performed on an Ultimate 3000 system (Dionex, Sunnyvale, CA, USA) coupled with an LTQ Orbitrap XL high-resolution mass spectrometer (Thermo Finnigan, San Jose, CA, USA). Forty nanograms of the sample were injected onto a reversed-phase C18 column (Michrom Bioresources, Auburn, CA, USA, Magic C18AQ, 200 µm × 150 mm, 3 µm, 200 Å) at a flow rate of 2 µL/min. Sample was eluted by using a linear gradient of mobile phase A (0.1% formic acid) and mobile phase B (0.1% formic acid in acetonitrile). The gradient increased from 2% B to 35% B in 28 min, then to 50% in 5 min and 90% in 4 min. Heated capillary temperature and electrospray voltage were 175°C and 2.0 kV, respectively. The mass spectrometer was operated in data-dependent MS/MS mode in which each full MS scan is followed by five MS/MS scans where the three most abundant pep-tide ions were selected for CID by using a normalized collision energy of 35%. Data were then analyzed manually.
EMSA and Reporter Assays
EMSA and reporter analyses were performed as described previously (30,31). For EMSA, nuclear extracts were prepared from either muscle tissues, or from C2C12 myotubes that were left untreated or treated for 30 min with TNFα. Treated cells were incubated for 1 h prior to TNFα treatment with vehicle (water) or increasing concentrations of TFA-NBD. Reporter assays were performed on C2C12 myotubes containing an NF-κB responsive luciferase reporter. Myotubes were left untreated or preincubated for 1 h with either water or TFA-NBD. TNFα then was added to activate NF-κB and after 4 h, cells were harvested and prepared for luciferase assays.
Histology
Serial frozen muscle sections (10 µm) were cut from the proximal, midbelly, and distal portions of the TA. For diaphragm, serial sections from muscle strips not used for functional analysis were cut in three separate places along the muscle in an attempt to obtain analysis representative of the entire muscle. Sections from each portion of the muscles were labeled for immunofluorescence. Primary antibodies were E-MyHC (1:50; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA, USA), Collagen IV, (1:500; Millipore, Billerica, MA, USA), and F4/80 (1:200; Serotec, Kidlington, UK). Alexa Fluor 488- and Alexa Fluor 568-conjugated secondary antibodies (1:250; Molecular Probes; Invitrogen, Carlsbad, CA, USA) were used for indirect immunofluorescence, and DAPI was used for nuclear detection. Staining was quantitated on an Olympus BX51 microscope with Microsuite Five software (Olympus Soft Imaging Solutions GmbH, Center Valley, PA, USA). Three entire sections (one from each portion of the muscle cut) were analyzed. Sections were quantitated by calculating the percentage of a section positive for IgG, eMyHC or F4/80 staining in proportion to the total area of the section (positive area/total area × 100%).
Statistics
Contractile forces were analyzed by using unpaired t tests and analysis of variance (ANOVA), followed by post hoc t tests where applicable. A maximum of two muscles per mouse were included in the study, and these were averaged per mouse for statistical analyses. Histology was analyzed by using an unpaired Student t test. A two-tailed P value of < 0.05 was considered significant. Data are given as mean ± SEM.
Results
We initiated our study by using the same non-GLP version of the NBD peptide that we and many other laboratories have used to demonstrate NF-κB involvement in a specific model of disease (13,23,32–37). This peptide is formulated in DMSO and was administered as a 10% solution in saline. The final dose of the compound was 200 µg per mouse, which, at the weights of the animals that we were using, converts approximately to a dose of 10 mg/kg. We found that the formulation of NBD as used in our studies, soluble in DMSO, exhibited considerable stability as assessed by mass spectrometry analysis. When compared with freshly prepared peptide, NBD remained stable after 1-day storage at room temperature and 2 wks at 37° C (Figure 1A).

NBD peptide is stable and improves mdx whole body function. (A) Mass spectrometry analysis of a NBD peptide depicts the strong stability of such a peptide after 1 d at room temperature and 2 wks at 37° C. (B) Rotarod strength/endurance measurements recorded from male and female WT or mdx mice treated with vehicle or NBD. Duration of treatment was 4 wks starting at 4 wks of age. Rotarod latency time was statistically different between NBD and vehicle mdx treated groups, but not between genders. *P< 0.05.
To determine how effective NBD was in restoring whole body function, we administered this peptide over a 4-wk period to male and female mdx mice, and rotarod strength/endurance tests subsequently were performed at 8 wks of age. Results from a treadmill exercise regimen showed that after 4 d of exercise, vehicle-treated (10% DMSO, 90% saline) mdx mice had a 20% to 25% reduction in whole body muscle strength/endurance, while animals treated with NBD exhibited a 16% (ANOVA, P < 0.05) improvement over this level (Figure 1B). This equated to a functional rescue of between 64% to 80% toward that of WT mice (deficit between WT and vehicle-treated mdx taken as 100% deficit). Similar results were obtained when performing an endurance test after 5 d of treadmill running (data not shown), and no differences between male and female mice were observed in any of the groups. Therefore, in the subsequent studies, both genders were used for analysis.
Next, specific diaphragm contractile function was assessed. Vehicle-treated WT (n = 16 from 10 mice) and vehicle-treated mdx (n = 37 from 22 mice) muscles were measured longitudinally to mdx mice subjected to various treatment regimens over the duration of the entire project, always with the use of an identical protocol. The specific force generated by 150 Hz tetani in WT mice was on average 167 mN/mm2. Owing to the paired-pulse protocol used in our studies, at lower frequencies we already obtained maximal tetanic force, an average of 181 mN/mm2 in WT mice. These levels of specific force are nearly identical with similar studies performed at 37° C; 180 mN/mm2 by Rucker et al. (29), 172 mN/mm2 by Prezant et al. (28), and 190 mN/mm2 by Diaz et al. (27). In WT mice that received our protocols, the isometric single-pulse twitch-to-isometric tetanus ratio was 3.6 ± 0.6, a value in-line with all above-mentioned previous studies, and was not different in any of the treatment groups. We found that at an initial frequency of 20 Hz, WT diaphragm muscles generated 62% (P < 0.005) more force than muscles from mdx vehicle-treated mice. At this same stimulation, NBD treatment (25 muscles from 14 mice) improved mdx diaphragm function by 43%, representing an average rescue toward WT of 78%. Significantly, this difference in force by NBD was maintained at the tetanic frequency of 150 Hz (Figure 2A).

NBD treatment improves diaphragm contractility and is dose dependent. Measurements of diaphragm tetanic force were assessed at 37° C at specific forces of 150 Hz. Values were averaged from WT (n = 16 muscles/10 mice), vehicle-treated mdxmice (Vehicle, n = 37 muscles/22 mice) and NBD (40 µg, n = 22 muscles/12 mice, and 200 µg, n = 25 muscles/14 mice) treated mdxmice. Duration of treatment was 4 wks starting at 4 wks of age. The lower dose was much less effective than a higher dose at improving diaphragm contractile force. *P < 0.05.
We considered whether a similar efficacy in mdx mice could be achieved by using a lower dose of NBD. We reasoned that obtaining a similar improvement would be beneficial from a cost perspective as well as from a potential safety profile. Therefore, treatments were repeated with the identical protocol except that five times less peptide was used. However, as results show (see Figure 2A), administrating NBD at a dose of 40 µg/mouse exhibited only a marginal improvement in diaphragm function (n = 22 muscles from 12 mice) over that of mdx vehicle-treated mice compared with WT or mdx mice treated with the original higher dose. These results suggest that under the current experimental approach, NBD improvement of mdx muscle function requires a dose above 40 µg/mouse.
In our efforts to develop NBD as a compound suitable for the clinic, a version of the peptide was synthesized under GLP conditions. The amino acid sequence including the Antennapedia protein transduction domain was identical to the parent non-GLP compound. However, the new version was soluble in water, which we considered beneficial for patient safety. Unfortunately, in sharp contrast to the parent compound (NBD), GLP-NBD showed only marginal and statistically nonsignificant improvement in diaphragm function of mdx mice (n = 15 muscles from nine mice, Figure 3). This less than efficacious response was maintained over the entire range of frequency stimulations, leading us to conclude that the GLP-NBD peptide did not provide a significant functional benefit to dystrophin-deficient respiratory muscles.

GLP-NBD does not improve mdx diaphragm function significantly. Composite representation of the force generated from diaphragm muscles at 150 Hz tetanus frequency from WT (n = 16 muscles/ 10 mice), vehicle-treated mdx (n = 37 muscles/22 mice), and mdx treated with GLP-NBD (n = 15). Duration of treatment was 4 wks starting at 4 wks of age. *P < 0.05 between indicated groups.
Given this result, it was pertinent to dissect the differential responses between the NBD and GLP-NBD peptides. Because the sequences of these compounds were identical, we reasoned that one explanation for the discrepancy in our results could be due to the manner in which each peptide was purified. Production of NBD utilized TFA as a counterion in the standard reverse phase HPLC purification phase, while GLP-NBD was generated with an extra purification step using acetate as a counterion. We thus speculated that the counterion that remained along with NBD during the lyophilization step could account for the solubility differences and thus potentially also explain differences in the functional outcome we observed. To test this possibility, we changed the purification counterion in GLP-NBD from acetate to TFA in an attempt to mimic the initially produced version of NBD. This formulation of the peptide was called TFA-NBD. Importantly, TFA-NBD was still soluble in water. However this advantage in solubility was in exchange for a marginal loss of peptide stability, as detected by mass spectrometry (data not shown).
To test the efficacy of the TFA-NBD peptide in vitro, C2C12 myotubes were pretreated with peptide and subsequently stimulated with TNFα for 30 min, at which time cells were harvested and analyzed for NF-κB DNA binding activity by an electrophoretic mobility shift assay (EMSA). Results showed that TFA-NBD, at two separate doses, was effective at inhibiting NF-κB DNA binding (Figure 4A). Similar results were obtained when TFA-NBD was tested for the ability to inhibit NF-κB transcriptional activity by using an NF-κB responsive reporter (Figure 4B). We next tested the ability of TFA-NBD to attenuate NF-κB activation in the muscles of mdx mice. After 4 wks of treatment (starting at 4 wks of age), NF-κB DNA binding was generally reduced in diaphragm muscles of TFA-NBD treated versus vehicle treated mdx mice (Figure 4C). These results support that TFA-NBD was indeed inhibiting NF-κB activity in vivo. Most importantly, when TFA-NBD was tested for efficacy in mdx mice, a 37% (P < 0.05) improvement in diaphragm function (n = 18 muscles from 11 mice) was achieved compared with control (Figure 4D). This represented a 63% rescue of the generated force produced by WT muscles, an improvement that was maintained over the entire range of tetanic frequencies employed. These values of improvement were within the statistical range recorded for the original DMSO NBD peptide, indicating that we were able to achieve diaphragm functional rescue successfully with an NBD peptide formulation viewed more favorable for clinical use.

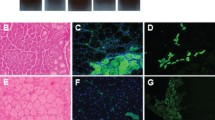
Conversion to TFA-NBD is effective in inhibiting NF-κB and improving diaphragm function and histopathology of mdx mice significantly. (A) EMSA analysis from untreated or TNFα treated C2C12 myotubes exposed to vehicle (water), 50 µmol/L, or 100 µmol/L of TFA-NBD. (B) C2C12 myotubes containing an NF-κB responsive luciferase reporter either were left untreated or were treated with TNFα after being exposed to either water, 100 µmol/L TFA-NBD or 200 µmol/L TFA-NBD. (C) EMSA analysis from vehicle or TFA-NBD treated diaphragm muscles. Duration of treatment was 4 wks starting at 4 wks of age. (D) Force measurements set at 150 Hz tetanus frequency were generated in diaphragm muscles from mdx mice treated with NBD converted to TFA (TFA-NBD, gray bar, n = 18 muscles/11 mice) compared with levels obtained with vehicle (mdx, hatched, n = 37 muscles/22 mice) and WT mice (open, n = 16 muscles/10 mice). (E-G) Percentages of total area within diaphragm and tibialis interior (TA) muscle sections positive for IgG (F), F4/80 (G) and eMyHC (H) between vehicle- and TFA-NBD-treated mice. *P < 0.05 among groups.
In addition to rescuing functional deficits in mdx mice, our previous results demonstrated that the DMSO NBD version of NBD was efficacious both in reducing injury and inflammation and in enhancing regeneration of mdx limb muscles (2). To assess whether similar benefits could be achieved with TFA-NBD, we performed histological analyses with skeletal muscles isolated from treated mdx mice. Results showed that TFA-NBD treatment was successful in reducing myofiber damage in diaphragm muscles by 79%, and to a lesser extent (64%) in limb muscles, as measured by IgG accumulation that reflects the level of muscle damage due to necrosis (Figure 4E). Macrophage accumulation, using F4/80 as a marker, also was reduced in diaphragm and limb muscle of mice treated with TFA-NBD (Figure 4F) by 56% and 72% respectively. Furthermore, eMyHC (embryonic myosin heavy chain) expression was significantly increased by 92% in diaphragm, and by 478% in limb muscles (Figure 4G), signifying that TFA-NBD treatment also produces a robust regenerative response that is characteristic of NF-κB inhibition in the presence of muscle injury. Together, these data demonstrate that in addition to providing similar functional rescue, TFA-NBD is equally effective for alleviating inflammation and myofiber injury and promoting muscle regeneration.
Discussion
In this study we performed an analysis to carefully evaluate the efficacy and feasibility of pursuing NBD as a therapeutic for DMD patients. Using a functionally effective, but clinically unsuitable peptide as a template, we were able to formulate a NBD peptide successfully that meets preclinical trial standards (GLP grade) and maintains reasonable stability dissolved in a nontoxic solution (water). Most importantly, the final peptide formulation retained the ability to rescue functional and histological dystrophic pathology in mice. These favorable findings support continued efforts to pursue NBD therapy as a treatment for DMD.
Given the devastating effects compromised diaphragm and limb muscle function have on DMD patients, a therapy capable of improving all skeletal muscles is of paramount importance both for extending life span and for maintaining a good quality of life. Unfortunately, many treatment strategies are being developed specifically to combat limb muscle dysfunction while potentially leaving diaphragm untreated. Our data supports the use of NBD therapy for treatment of both respiratory and limb muscle pathology in DMD. To the best of our knowledge, no other pharmacological therapy has shown the high degree of diaphragm functional rescue that we were able to achieve treating mdx mice with NBD.
NF-κB is a vital signaling pathway relevant at the cellular level for regulating functions such as survival, apoptosis and proliferation as well as for whole body welfare, regulating critical genes involved in immune responses (38). Consequently, inhibiting all signaling through NF-κB likely would result in a therapy where the negative side effects outweigh the desired efficacy of the treatment. A major advantage of using the NBD peptide therapeutically is that it does not interfere with basal NF-κB signaling, only with inflammatory signaling through the IKK complex (13,23). Also important for reducing undesired side effects is a therapy’s functional specificity. NBD has been shown to be specific for inhibiting NF-κB signaling, exhibiting no inhibitory effects toward JNK or p38 MAPK pathways (13). The safety profile for NBD is favorable as well. In vivo, systemic delivery of NBD is not associated with any described toxicity in mice or rats and inhibition of NF-κB has been demonstrated to ameliorate an ever-growing list of inflammatory disease conditions (33,34,37,39–41). Furthermore, in a mouse model of arthritis, systemic administration of NBD for only 5 days was sufficient to maintain clinical improvement for nearly 3 weeks (41), demonstrating that the therapeutic effect of the NBD peptide may outlast the pharmacokinetic properties of this peptide.
In this study, we demonstrate the feasibility of pursuing the NBD peptide as a therapeutic for alleviating DMD pathophysiology. The development of a GLP-approved, water-soluble peptide that ameliorates muscle functional deficits, inflammation and necrosis while promoting muscle regeneration in mdx mice provides preclinical evidence for the efficacy and safety of the NBD peptide.
Disclosure
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
References
Blake DJ, Weir A, Newey SE, Davies KE. (2002) Function and genetics of dystrophin and dystrophin-related proteins in muscle. Physiol. Rev. 82:291–329.
Acharyya S, et al. (2007) Interplay of IKK/NF-kappaB signaling in macrophages and myofibers promotes muscle degeneration in Duchenne muscular dystrophy. J. Clin. Invest. 117:889–901.
Monici MC, Aguennouz M, Mazzeo A, Messina C, Vita G. (2003) Activation of nuclear factor-kappaB in inflammatory myopathies and Duchenne muscular dystrophy. Neurology. 60:993–7.
Chen YW, et al. (2005) Early onset of inflammation and later involvement of TGFbeta in Duchenne muscular dystrophy. Neurology. 65:826–34.
Messina S, et al. (2006) Lipid peroxidation inhibition blunts nuclear factor-kappaB activation, reduces skeletal muscle degeneration, and enhances muscle function in mdx mice. Am. J. Pathol. 168:918–26.
Messina S, et al. (2006) Nuclear factor kappa-B blockade reduces skeletal muscle degeneration and enhances muscle function in Mdx mice. Exp. Neurol. 198:234–41.
Mendell JR, et al. (1989) Randomized, doubleblind six-month trial of prednisone in Duchenne’s muscular dystrophy. N. Engl. J. Med. 320:1592–7.
De Bosscher K, et al. (1997) Glucocorticoid-mediated repression of nuclear factor-kappaB-dependent transcription involves direct interference with transactivation. Proc. Natl. Acad. Sci. U. S. A. 94:13504–9.
De Bosscher K, et al. (2000) Glucocorticoids repress NF-kappaB-driven genes by disturbing the interaction of p65 with the basal transcription machinery, irrespective of coactivator levels in the cell. Proc. Natl. Acad. Sci. U. S. A. 97:3919–24.
Karin M, Ben-Neriah Y. (2000) Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu. Rev. Immunol. 18:621–63.
Perkins ND. (2007) Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell. Biol. 8:49–62.
Hayden MS, Ghosh S. (2008) Shared principles in NF-kappaB signaling. Cell. 132:344–62.
Baima ET, et al. (2010) Novel insights into the cellular mechanisms of the anti-inflammatory effects of NF-kappaB essential modulator binding domain peptides. J. Biol. Ghem. 285:13498–506.
Akers WS, Cross A, Speth R, Dwoskin LP, Cassis LA. (2000) Renin-angiotensin system and sympathetic nervous system in cardiac pressure-overload hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 279:H2797–806.
Archer JD, Vargas CC, Anderson JE. (2006) Persistent and improved functional gain in mdx dystrophic mice after treatment with L-arginine and deflazacort. Faseb. J. 20:738–0.
Barton ER, Morris L, Kawana M, Bish LT, Toursel T. (2005) Systemic administration of L-arginine benefits mdx skeletal muscle function. Muscle Nerve. 32:751–60.
Carlson CG, Samadi A, Siegel A. (2005) Chronic treatment with agents that stabilize cytosolic IkappaB-alpha enhances survival and improves resting membrane potential in MDX muscle fibers subjected to chronic passive stretch. Neurobiol. Dis. 20:719–30.
Hnia K, et al. (2008) L-arginine decreases inflammation and modulates the nuclear factor-kappaB/matrix metalloproteinase cascade in mdx muscle fibers. Am. J. Pathol. 172:1509–19.
Pan Y, et al. (2008) Curcumin alleviates dystrophic muscle pathology in mdx mice. Mol. Cells. 25:531–7.
Siegel AL, et al. (2009) Treatment with inhibitors of the NF-kappaB pathway improves whole body tension development in the mdx mouse. Neuromuscul. Disord. 19:131–9.
Voisin V, et al. (2005) L-arginine improves dys-trophic phenotype in mdx mice. Neurobiol. Dis. 20:123–30.
Stedman HH, et al. (1991) The mdx mouse diaphragm reproduces the degenerative changes of Duchenne muscular dystrophy. Nature. 352:536–9.
May MJ, et al. (2000) Selective inhibition of NF-kappaB activation by a peptide that blocks the interaction of NEMO with the IkappaB kinase complex. Science. 289:1550–4.
Mulieri LA, Hasenfuss G, Ittleman F, Blanchard EM, Alpert NR. (1989) Protection of human left ventricular myocardium from cutting injury with 2,3-butanedione monoxime. Circ. Res. 65:1441–9.
Chandrasekharan K, et al. (2010) A human-specific deletion in mouse Cmah increases disease severity in the mdx model of Duchenne muscular dystrophy. Sci. Transl. Med. 2:42ra54.
Janssen PML, Hiranandani N, Mays TA, Rafael-Fortney JA. (2005) Utrophin deficiency worsens cardiac contractile dysfunction present in dystrophin-deficient mdx mice. Am. J. Physiol. Heart Circ. Physiol. 289:H2373–8.
Diaz PT, Brownstein E, Clanton TL. (1994) Effects of N-acetylcysteine on in vitro diaphragm function are temperature dependent. J. Appl. Physiol. 77:2434–9.
Prezant DJ, et al. (1990) Temperature dependence of rat diaphragm muscle contractility and fatigue. J. Appl. Physiol. 69:1740–5.
Rucker M, et al. (2004) Rescue of enzyme deficiency in embryonic diaphragm in a mouse model of metabolic myopathy: Pompe disease. Development. 131:3007–19.
Acharyya S, et al. (2005) Dystrophin glycoprotein complex dysfunction: a regulatory link between muscular dystrophy and cancer cachexia. Cancer Cell. 8:421–32.
Guttridge DC, Albanese C, Reuther JY, Pestell RG, Baldwin AS Jr. (1999) NF-kappaB controls cell growth and differentiation through transcriptional regulation of cyclin D1. Mol. Cell. Biol. 19:5785–99.
Chapoval SP, et al. (2007) Inhibition of NF-kappaB activation reduces the tissue effects of transgenic IL-13. J. Immunol. 179:7030–41.
Dasgupta S, et al. (2004) Antineuroinflammatory effect of NF-kappaB essential modifier-binding domain peptides in the adoptive transfer model of experimental allergic encephalomyelitis. J. Immunol. 173:1344–54.
di Meglio P, Ianaro A, Ghosh S. (2005) Amelioration of acute inflammation by systemic administration of a cell-permeable peptide inhibitor of NF-kappaB activation. Arthritis Rheum. 52:951–8.
Ghosh A, et al. (2007) Selective inhibition of NF-kappaB activation prevents dopaminergic neuronal loss in a mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. U. S. A. 104:18754–9.
Shibata W, et al. (2007) Cutting edge: The IkappaB kinase (IKK) inhibitor, NEMO-binding domain peptide, blocks inflammatory injury in murine colitis. J. Immunol. 179:2681–5.
Tas SW, et al. (2006) Local treatment with the selective IkappaB kinase beta inhibitor NEMO-binding domain peptide ameliorates synovial inflammation. Arthritis Res. Ther. 8:R86.
Hayden MS, Ghosh S. (2004) Signaling to NF-kappaB. Genes. Dev. 18:2195–224.
Clohisy JC, Yamanaka Y, Faccio R, Abu-Amer Y. (2006) Inhibition of IKK activation, through sequestering NEMO, blocks PMMA-induced osteo-clastogenesis and calvarial inflammatory osteolysis. J. Orthop. Res. 24:1358–65.
Dai S, Hirayama T, Abbas S, Abu-Amer Y. (2004) The IkappaB kinase (IKK) inhibitor, NEMO-binding domain peptide, blocks osteoclastogenesis and bone erosion in inflammatory arthritis. J. Biol. Chem. 279:37219–22.
Jimi E, et al. (2004) Selective inhibition of NF-kappa B blocks osteoclastogenesis and prevents inflammatory bone destruction in vivo. Nat. Med. 10:617–24.
Acknowledgments
This study was supported by a grant from the National Institutes of Health U01 NS058451 (to DC Guttridge, DJ Ricca, PML Janssen, JA Rafael-Fortney, JR Mendell, PR Clemens, PD Robbins, JN Kornegay), as well as support from the Muscular Dystrophy Association (to DC Guttridge and JM Peterson), and a K02 HL08357 award (to PML Janssen). We thank JI Elliott at Yale University for assistance with NBD peptide. The authors take full responsibility for the contents of this paper, which do not represent the views of the Department of Veterans Affairs or the United States Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
JMP, WK, and BDC all contributed equally to this paper.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Peterson, J.M., Kline, W., Canan, B.D. et al. Peptide-Based Inhibition of NF-κB Rescues Diaphragm Muscle Contractile Dysfunction in a Murine Model of Duchenne Muscular Dystrophy. Mol Med 17, 508–515 (2011). https://doi.org/10.2119/molmed.2010.00263
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/molmed.2010.00263