
Abstract
Ischemia and reperfusion injury (IRI) contributes to the development of chronic interstitial fibrosis/tubular atrophy in renal allograft patients. Cyclooxygenase (COX) 1 and 2 actively participate in acute ischemic injury by activating endothelial cells and inducing oxidative stress. Furthermore, blockade of COX 1 and 2 has been associated with organ improvement after ischemic damage. The aim of this study was to evaluate the role of COX 1 and 2 in the development of fibrosis by performing a COX 1 and 2 blockade immediately before IRI. We subjected C57Bl/6 male mice to 60 min of unilateral renal pedicle occlusion. Prior to surgery mice were either treated with indomethacin (IMT) at days −1 and 0 or were untreated. Blood and kidney samples were collected 6 wks after IRI. Kidney samples were analyzed by real-time reverse transcription-polymerase chain reaction for expression of transforming growth factor β (TGF-β), monocyte chemoattractant protein 1 (MCP-1), osteopontin (OPN), tumor necrosis factor a (TNF-α), interleukin (IL)-1β, IL-10, heme oxygenase 1 (HO-1), vimentin, connective-tissue growth factor (CTGF), collagen I, and bone morphogenic protein 7 (BMP-7). To assess tissue fibrosis we performed morphometric analyses and Sirius red staining. We also performed immunohistochemical analysis of anti-actin smooth muscle. Renal function did not significantly differ between groups. Animals pretreated with IMT showed significantly less interstitial fibrosis than nontreated animals. Gene transcript analyses showed decreased expression of TGF-β, MCP-1, TNF-α, IL-1-β, vimentin, collagen I, CTGF, and IL-10 mRNA (all P < 0.05). Moreover, HO-1 mRNA was increased in animals pretreated with IMT (P < 0.05). Conversely, IMT treatment decreased osteopontin expression and enhanced BMP-7 expression, although these levels did not reach statistical significance when compared with control expression levels. The blockade of COX 1 and 2 resulted in less tissue fibrosis, which was associated with a decrease in proinflammatory cytokines and enhancement of the protective cellular response.
Similar content being viewed by others


Introduction
In renal ischemia and reperfusion injury (IRI), inflammation results in changes in the endothelial cell phenotype. These changes enhance leukocyte adhesion, entrapment, and transmigration, ultimately compromising the regional blood flow (1). In kidney transplantation, IRI is implicated as one of the potential contributors to the development of chronic allograft nephropathy, now recognized as graft tubular atrophy/interstitial fibrosis, the main cause of graft loss after the first year of transplantation (2,3).
In the initial period of reperfusion, there is a rapid increase in proinflammatory cytokines, such as a tumor necrosis factor α (TNF-α) and interleukin (IL)-1β, induced in part by infiltrating mononuclear and endothelial cells (4,5). There are also significant increases in the syntheses of leukotrienes and cyclooxygenase (COX) (3,6).
COX 1 and 2 can be detected in kidney tissue subjected to IRI, and recent data from our group and other investigators have shown that first these molecules were upregulated after IRI, and that their activation has a negative impact on organ function outcome (7,8). Indeed, COX blockade was associated with an improvement in renal function and a decrease in reactive oxygen species (9,10).
More importantly, the inflammation triggered by IRI is not restricted to the early days after the insult. Experimental data have demonstrated that neutrophil activation and CD4+ T-cell infiltration persisted 6 wks after the initial injury, culminating in tissue fibrosis (11,12). Normally, during the process of tubule-interstitial remodeling, fibrosis results from an imbalance between the synthesis and the degradation of proteins of the extracellular matrix (13,14) with gradual loss of renal tubules and their function (15). Several cytokines participate in fibrogenesis, including transforming growth factor β (TGF-β), platelet-derived growth factor (PDGF), connective tissue growth factor (CTGF) (16,17), vimentin (18), collagen I (19–21), IL-1, fibroblast growth factor (FGF), osteopontin (OPN), endothelin, and TNF-α (22). Molecules such as heme oxygenase 1 (HO-1) and bone morphogenic protein-7 (BMP-7), however, are thought to balance the profibrotic effect of the former molecules.
In the current work, we hypothesized that COXs also contribute to this cascade of persistent inflammation. To address this question, we worked with an established model of unilateral IRI. Experimental animals were either pretreated with a COX inhibitor or not pretreated before being subjected to acute injury and followed for 6 wks after injury. We anticipated that the animals treated with the COX inhibitor would exhibit the lowest proinflammatory and profibrotic cytokine levels and have significantly less tissue fibrosis.
Materials and Methods
Unilateral Renal IRI Model
Male C57Bl/6 mice, 10 wks of age (25–28 g), were purchased from the Animal Facilities Center from the Institute of Pharmacology at the Universidade Federal de São Paulo (São Paulo, Brazil). The mice were subjected to 60 min unilateral renal pedicle occlusion and were then monitored for 6 wks after ischemia. Animals were anesthetized with intraperitoneal ketamine and xylazine (100 mg/kg and 10 mg/kg, respectively; Agribrands do Brazil, São Paulo, Brazil). After surgery, animals were housed in individual cages with free access to water and food. This project was approved by the Ethics Committee on Human and Animal Research of our institution.
Drug
Mice were treated with IMT (5 mg/kg/d) 1 d before and on the day of surgery (n = 8) or not treated (n = 8) (23,24). Sham-operated animals (n = 5) were used as controls.
Functional Analyses
Blood and kidney samples were collected 6 wks after unilateral IRI.
Assessment of Renal Function
Blood was collected for urea and serum creatinine measurements, and kidneys were harvested for histological and mRNA analyses at the end points. Urea was measured in duplicate by the Urease method and serum creatinine by Jaffe’s method, using a Cobas Mira Plus autoanalyzer (Roche, Mannheim, Germany). The extent of fibrosis was determined by use of Masson Trichrome stain and performed by blinded morphometric assessment using a computer-assisted image system with an Olympus BX40F-3 microscope (Olympus Optical, Tokyo, Japan) and the software program Image Pro Plus for Windows 3.0 (Media Cybernetic, Silver Spring, MD, USA).
Gene Profile
Kidney samples were quickly frozen in liquid nitrogen. Total RNA was isolated from kidney tissues using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) methodology. RNA concentration was determined by spectrophotometer readings at an absorbance of 260 nm. First-strand cDNAs were synthesized using moloney murine leukemia virus reverse transcriptase (Promega, Madison, WI, USA). Reverse transcription-polymerase chain reaction (PCR) was performed using the SYBR Green real-time PCR assay (Applied Biosystems, Foster City, CA, USA) and specific primers for monocyte chemoattractant protein (MCP)-1 (sense: 5′-ACT GCA TCT GCC CTA AGG TCT TCA-3′, antisense: 5′-AGA AGT GCT TGA GGT GGT TGT GGA-3′); IL-1β (sense: 5′-CCT AAA GTA TGG GCT GGA CTG TTT-3′, antisense: 5′-TAG AGA TTG AGC TGT CTG CTC ATT C-3′); TGF-β (sense: 5′-AAC TAT TGC TTC AGC TCC AGA GAG A-3′, antisense: 5′-AGT TGG ATG GTA GCC CTT G-3′); and OPN (sense: 5′-GGC ATT GCC TCC TCC CTC-3′, antisense: 5′ CGA GGC TGT AAA GCT TCT CC-3′). We used a TaqMan real-time PCR assay for TNF-α (Mm00443258_m1), IL-10 (Mm01288386_m1), BMP-7 (Mm00432102_m1), HO-1 (Mm00516007_m1) (a protective gene, vimentin (Mm01333430_m1), CTGF (Mm00515790_g1), and collagen I (Mm01302047_g1). Hypoxanthine phosphoribosyltransferase (HPRT) was used as a housekeeping gene. PCR data were presented as a ratio of genes to HPRT mRNA (sense: 5′-CTC ATG GAC TGA TTA TGG ACA GGA C-3′ and antisense: 5’-GCA GGT CAG CAA AGA ACT TAT AGC C-3’ and Mm03024075_m1) in arbitrary units (AU).
Immunohistochemical Analysis: Smooth Muscle Cell Actin Staining
Paraffin block sections (5µm) were placed on poly-L-lysine-coated slides. After the sections were dried in an oven for 1 h at 60°C, they were dewaxed in xylol for 15 min and hydrated in 3 alcohols (100% for 9 min, 95% for 4 min, and 70% for 2 min). Subsequently, sections were incubated for 10 min in 3% hydrogen peroxide to block endogenous peroxidase. For antigen retrieval, after being washed with phosphate-buffered saline (PBS), each section was unspecifically blocked. Sections were then incubated overnight at 4°C with anti-actin smooth muscle (α-SMA; Calbiochem, Schwalbach, Germany). After a new wash with PBS for 9 min, the sections were incubated in a wet chamber for 30 min with a secondary goat anti-mouse antibody (NCL-GAMP; NovoCastra, Norwell, MA, USA) and washed with PBS. Endogenous peroxidase activity was detected by reacting fixed tissue sections with 3.3′-diaminobenzidine substrate plus 10% hydrogen peroxide for 10 min. After being washed with distilled water, the samples were stained with hematoxylin for 10 s. Dehydration was performed with 3 alcohols (70% for 1 min, 95% for 2 min, and 100% for 6 min) and with xylol for 9 min. All quantification was performed blinded. α-SMA staining was calculated as the percentage of renal cortex and medulla. We used computer image analysis (KS300, Zeiss system) to measure the percentage of area occupied by α-SMA staining per 4 mm2 field at a magnification of 200 and calculate the mean percentage area for each biopsy (25).
Sirius red staining. Four micra formalin fixed slides were stained with Sirius red (F3BA CI 35782; Sigma, St. Louis, MO, USA) 0.1% in saturated picric acid for 60 min at room temperature. Slides were washed twice for 5 min in 0.5% glacial acetic acid and counterstained with Harris hematoxylin. Fibrosis was calculated as the percentage of renal cortex and medulla. We used computer image analysis (KS300, Zeiss system) to measure the percentage of area occupied by Sirius red staining per 4 mm2 field at a magnification of 200 and calculate the mean percentage area calculated for each sample.
Statistics
Data were graphically displayed as mean and standard deviation (SD) or as median (histomorphometric analyses) owing to the Gaussian distribution of variables. Mann-Whitney U test, Student t test, and ANOVA on ranks tests were used to compare the data. PCR data were presented as a ratio of analyzed genes to HPRT mRNA, in arbitrary units (AU). Significant difference was considered when P < 0.05.
Results
Long-term Renal Function Outcomes
Urea and creatinine levels were measured at 6 wks following IRI in animals treated with IMT or otherwise. In this model, renal function was not altered significantly and the previous treatment with IMT did not change these results (Figure 1A). The ischemic kidneys weighted significantly less than the contralateral nonischemic ones, but the IMT pretreatment did not change this relationship (Figure 1B). Therefore, we performed morphometric analyses to quantify tissue fibrosis at this time. Interestingly, animals treated with IMT prior to surgery had less fibrosis compared with nontreated animals (Figure 2). Sham-operated animals presented basal levels similar to normal non-manipulated animals (data not shown).

Renal function and renal fibrosis after unilateral IRI. (A) Blood urea and serum creatinine levels at 6 wks after 60 min of unilateral renal ischemia, in animals treated with IMT or not treated. There were no significant differences between groups (P > 0.05). (B) Kidney weights (in grams) at 6 wks after 60 min of unilateral renal ischemia, in animals treated with IMT or not treated. There were no significant differences between groups (P> 0.05).

Morphometric analyses at 6 wks after unilateral IRI. (A) Histological analyses of kidneys (contralateral (CL) and ischemic kidneys) were performed at 6 wks after 60 min of unilateral renal ischemia in animals not treated or treated with IMT. *P < 0.05.
Cytokine Profiles
As previously seen in renal IRI, as early as 24 h after reperfusion there is strong amplification of proinflammatory cytokines such as IL-1β, TNF-β, and interferon γ. Data from our laboratory and other groups have demonstrated that the Th1 immune response is deleterious to renal tissue after IRI (26,27). Therefore, we investigated the profile of inflammatory cytokines in ischemic kidneys treated with IMT or otherwise. To compare these results, we also amplified the transcripts in the contralateral kidneys, as controls.
Ischemic kidneys presented a proinflammatory phenotype evidenced by the higher expression of MCP-1, IL-1β, and TNF-α (Figure 3). Values for the expression of these cytokines were even higher when we compared them with values for the contralateral kidneys. Interestingly, animals pretreated with IMT showed a significant decrease in mRNA expression consistent with decreased cytokine levels, even in contralateral counterparts.

Proinflammatory cytokine profile at 6 wks after unilateral IRI. mRNA levels of MCP 1 (A, *P < 0.05), IL-1 β (B, *P = 0.013), and TNF-α (C, *P = 0.029 and **P = 0.028) in contralateral and ischemic kidneys in animals treated or not with IMT. mRNAs were standardized for HPRT mRNA and were expressed as AU.
We also measured IL-10, an anti-inflammatory cytokine, and HO-1, considered a protective gene. IMT treatment decreased IL-10 expression compared with nontreatment (Figure 4A). Taking into consideration HO-1 expression, as previously described in acute renal injury, IMT treatment resulted in an impressive amount of HO-1 upregulation in this chronic model (Figure 4B).

Anti-inflammatory profile at 6 wks after unilateral IRI. mRNA levels of IL-10 (A, *P = 0.03) and HO-1 (B, *P < 0.05) in contralateral and ischemic kidneys in animals treated or not with IMT. mRNA was standardized for HPRT mRNA and expressed as AU.
Fibrotic Changes after IMT Treatment
Pro- and antifibrotic factor expression can change during the course of renal IRI. We therefore studied the expression of these molecules in mice at 6 wks after ischemia. We analyzed TGF-β, OPN, vimentin, CTGF, and collagen I. These molecules have previously been shown to be upregulated in fibrosis (28–31). In our model, we observed an increase of mRNA for TGF-β, OPN, vimentin, collagen I, and CTGF in ischemic kidneys compared with contralateral kidneys. After IMT treatment, these increased levels were less evident, but reached statistical significance for expression of TGF-β, vimentin, collagen, and CTFG (Figure 5). The effect of IMT went beyond growth factor expression; compared with nontreated ischemic animals, treated animals showed increases in α-SMA staining (Figure 6A) and mirrored upregulation in BMP-7 expression, although these results were not statistically significant (P > 0.05) (Figure 5C). These quantitative immunohistochemical analyses of α-SMA-positive cells corroborated the effects of COX inhibition on renal tissue fibrosis seen in gene transcript analyses (Figure 6A). Furthermore, IMT-treated animals presented significantly less positive Sirius-red staining than nontreated animals (Figure 6B).

Pro- and antifibrotic profile at 6 wks after unilateral IRI. mRNA levels of TGF-β (A, *P < 0.05), OPN (B, *P < 0.05), BMP-7 (C, *P < 0.05), vimentin (D, *P < 0.05), collagen I (E, *P < 0.05), and CTGF (F, *P< 0.05) in contralateral and ischemic kidneys in animals treated or not with IMT. mRNA was standardized for HPRT mRNA and expressed as AU.

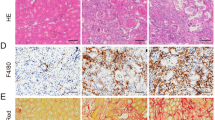
Immunohistochemical analyses of α-SMA and Sirius red staining. (A) Quantification of a-SMA-positive cells in medullar renal tissue 6 wks after 60 min of unilateral ischemia by immunohistochemical analysis (*P < 0.05). (B) Sirius red staining of kidneys (contralateral and ischemic kidneys) was performed 6 wks after animals were subjected to 60 min of unilateral renal ischemia and not treated or treated with IMT.
Discussion
IRI is the foremost antigen-independent factor related to the development of chronic allograft nephropathy, the leading cause of graft loss after the first year of renal transplantation. The inflammation triggered by IRI, especially in the tubule-interstitial compartment, results in fibrosis that is detectable in long-term follow-up (11). The development of alternative strategies to counteract the development of fibrotic tissue is essential for improving survival of patients who receive transplants from deceased individuals.
In this study, we aimed to demonstrate that blockade of the COX pathway could have a beneficial effect in preventing the development of fibrosis after IRI. Of note, we performed the COX blockade prior to acute renal injury. We have previously demonstrated that a single episode of severe unilateral warm ischemic injury to kidney is associated with significant long-term kidney inflammation and development of tissue fibrosis. These data are in accordance with the previously reported finding that a single ischemic episode can lead to maintenance of the inflammatory state, attributable mainly to neutrophils and CD4+ T-cell infiltration. We used a model of severe ischemia to mimic the long-term effects of ischemia seen in recipients of kidney transplants from deceased individuals (32). Lower degrees of ischemia are less likely to be associated with clinically significant long-term problems.
The extension of tissue damage did not alter renal function but did decrease organ size. The histological studies corroborated this finding. At this point, the IMT treatment had no effect on organ size but had a great impact on the development of fibrosis. Histomorphometric analyses showed that less fibrosis was seen in IMT-treated animals. These results indicate that early inflammation induced by ischemic stress leads to longterm fibrosis development, as previously demonstrated (11), and more importantly, the blockade of this early-onset inflammation is beneficial in halting this process.
Because inflammation seems to be linked with fibrogenesis in our model, we opted to quantify a broad range of proinflammatory cytokines/growth factors and of BMP-7, HO-1, and IL-10, molecules that can counteract the actions of proinflammatory cytokines/growth factors.
It is well established that fibrogenesis results from an imbalance between growth factors and cytokines such as TGF-β, PDGF, CTGF, collagen I, OPN, FGF, IL-1α and -1β, and BMP-7 (16–21, 33–35). Proximal epithelial cells as well as infiltrating cells may produce these molecules, stimulated by the reactive oxygen species secreted by the injured cells (36,37). Vimentin has also been linked to fibrogenesis in the kidney (18) and to epithelial-to-mesenchymal transition (EMT) (38,39). These injured proximal tubular cells seem to release chemotactic factors, such as MCP-1, which promotes leukocyte attachment to tubular lumen and infiltration into the interstitium, amplifying this inflammatory cascade (40). Gloria and colleagues have demonstrated in an in vitro cell model of gaseous hypoxia that COX blockade decreased the expression of RANTES (regulated on activation normal T-cell expressed and secreted), another chemokine involved in leukocyte transmigration (41). Our data demonstrated that the animals pretreated with IMT presented less MCP-1 expression, a finding that supports this hypothesis.
Pretreatment with IMT significantly decreased the levels of proinflammatory cytokines, including IL-1β and TNF-α. The production of IL-1β by the glomerular cells is increased in several types of clinical and experimental glomerulopathy in which fibrosis occurs. IL-1β stimulates the proliferation of fibroblasts and possibly the synthesis of extracellular matrix proteins, and it also has a chemotactic effect. Importantly, treatment with IL-1β antagonists reduces fibrosis in rats with glomerulonephritis (42). Furthermore, Stanimirovic et al. demonstrated that in cerebral IRI COX inhibition by IMT reduced neutrophil adhesion and completely inhibited IL-1β mRNA expression (43). Thus, we can infer that the decreased extension of fibrosis may be partly attributable to decreased expression of these molecules owing to the action of IMT.
When we analyzed the profile of pro-and antifibrotic molecules, we observed that after 6 wks of reperfusion ischemic kidney tissue exhibited increased expression of profibrotic factors, namely TGF-β, vimentin, collagen I, CTGF, and OPN. The effect of IMT on TGF-β expression is important given the critical role of TGF-β in renal fibrosis. The EMT (44) may be attributable to TGF-β inducing myofibroblast transformation and increasing collagen synthesis. The present study demonstrated that COX inhibition can actually decrease TGF-β expression, a decrease associated with signs of decreased tissue fibrosis. Additional studies should be carried out to ascertain whether COX accelerates EMT. The expression of OPN, collagen I, and α-SMA were in accordance with that of TGF-β. α-SMA is expressed during EMT and is a marker of “activated” fibroblasts. Therefore, IMT could decrease the fibrosis by reducing these cells in the interstitium. The reduction in OPN expression indicates, beyond the reduction of fibrosis, lower macrophage infiltration, because OPN is considered one of the chemokines that attracts macrophages into the injured renal tissue (45). Macrophage infiltration correlates with the development of fibrosis (46–49). Finally, TGF-β signaling is regulated by BMP-7, also a member of the TGF-β family, through activation of Smad-6. This regulation can inhibit EMT (50).
Vimentin is a transitional filament protein mainly expressed in mesenchymal-derived cells. It can be reexpressed in renal tubular epithelial cells under many pathological conditions such as ischemia. Terzi and colleagues demonstrated that vimentin expression is up-regulated after renal ischemia in proximal tubular cells (51). CTGF seems to be responsible for some TGF-β profibrotic activities (52). Nguyen and Goldschmeding have recently proposed that CTGF inihibition or BMP-7 induction could be a future therapy for renal fibrosis (52). Nguyen and Goldschmeding showed that BMP-7 expression was diminished in the ischemic kidneys compared with the contralateral kidneys and that treatment with IMT reversed BMP-7 expression, albeit without reaching statistical significance, and that IMT significantly decreased CTGF expression. These results could be promising because we have already observed that in animals treated with rapamycin and then subjected to unilateral ureteral obstruction, a model of renal fibrosis, the increased expression of BMP-7 was associated with less renal fibrosis (53). The blockade of CTGF has also been showed to be protective in reducing fibrosis in a unilateral ureteral obstruction model (54).
The action of proinflammatory cytokines could also be antagonized by favoring a Th2 immune response or by enhancing innate cellular protective gene machinery. We analyzed the expression of IL-10, an anti-inflammatory cytokine, and HO-1, a protective gene. Some studies have shown that the increase of IL-10 expression diminishes intercellular adhesion molecule 1 and selectins (55,56), which are both responsible for neutrophil rolling in the endothelium, protecting the kidney from IRI. Furthermore, the inhibition of these molecules by chemical drugs or in knockout animals is associated with tissue protection (57). Consequently, we investigated whether treatment with IMT would have an influence on expression of HO-1 and IL-10. We observed an increase of HO-1 in the ischemic kidneys; however, this increase was visibly higher when the animals were treated with IMT. These data corroborate our observation that pretreatment with IMT in animals was associated with a sustained increase in HO-1 following an acute renal injury (3). Conversely, IL-10 expression was increased in ischemic kidneys but was reduced when the animals were pretreated with-IMT. Although IL-10 and HO-1 can inhibit proinflammatory genes associated with activation of monocytes/macrophages, such as TNF-α, IL-1-β, and granulocyte-macrophage-colony-stimulating factor, in some specific models the anti-inflammatory effects of HO-1 seem to be independent of those of IL-10. Prostaglandin E2 can also have anti-inflammatory properties through induction of IL-10 (58,59), and the data expression found with IL-10 could be a reflection of an already protective response initiated by the organ after the initial injury. This response, however, is not strong enough to halt fibrosis. In this sense, the protection seen with IMT seems to be HO-1 dependent and IL-10 independent.
Recent data have established that IRI is an inflammatory syndrome and, more importantly, that the inflammatory response of IRI is persistent, culminating in the development of fibrosis. Renal fibrosis is the final pathway of many diseases, and no known treatment can halt or regress it. Our results demonstrated that COX 1 and 2 play an important role in the development of sustained inflammation. Their blockade prior to an acute injury could be beneficial in limiting the renal damage caused by fibrosis.
Disclosure
The authors have no conflicts of interest.
References
Molitoris BA. (1991) New insights into the cell biology of ischemic acute renal failure. J. Am. Soc. Nephrol. 1:1263–70.
Nankivell BJ, Borrows RJ, Fung CL, O’Connell PJ, Allen RD, Chapman JR. (2003) The natural history of chronic allograft nephropathy. N. Engl. J. Med. 349:2326–33.
Feitoza CQ et al. (2007) A role for HO-1 in renal function impairment in animals subjected to ischemic and reperfusion injury and treated with immunosuppressive drugs. Transplant Proc. 39:424–6.
Jaeschke H, Farhood A. (1991) Neutrophil and Kupffer cell-induced oxidant stress and ischemia-reperfusion injury in rat liver. Am. J. Physiol. 260: G355–62.
Colletti LM, Remick DG, Burtch GD, Kunkel SL, Strieter RM, Campbell DA Jr. (1990) Role of tumor necrosis factor-alpha in the pathophysiologic alterations after hepatic ischemia/reperfusion injury in the rat. J. Clin. Invest. 85:1936–43.
Karasawa A, Guo JP, Ma XL, Tsao PS, Lefer AM. (1991) Protective actions of a leukotriene B4 antagonist in splanchnic ischemia and reperfusion in rats. Am. J. Physiol. 261: G191–8.
Harris RC. (2000) Cyclooxygenase-2 in the kidney. J. Am. Soc. Nephrol. 11:2387–94.
Hausknecht B, Voelkl S, Riess R, Gauer S, Goppelt-Struebe M. (2003) Expression of cyclooxygenase-2 in biopsies obtained from human transplanted kidneys undergoing rejection. Transplantation 76:109–14.
Feitoza CQ et al. (2005) Cyclooxygenase 1 and/or 2 blockade ameliorates the renal tissue damage triggered by ischemia and reperfusion injury. Int. Immunopharmacol. 5:79–84.
Feitoza CQ, Sanders H, Cenedeze M, Camara NO, Pacheco-Silva A. (2002) Pretreatment with indomethacin protects from acute renal failure following ischemia-reperfusion injury. Transplant Proc. 34:2979–80.
Burne-Taney MJ, Yokota N, Rabb H. (2005) Persistent renal and extrarenal immune changes after severe ischemic injury. Kidney Int. 67:1002–9.
Friedewald JJ, Rabb H. (2004) Inflammatory cells in ischemic acute renal failure. Kidney Int. 66:486–91.
Azuma H, Nadeau K, Takada M, Mackenzie HS, Tilney NL. (1997) Cellular and molecular predictors of chronic renal dysfunction after initial ischemia/reperfusion injury of a single kidney. Transplantation 64:190–7.
Williams P, Lopez H, Britt D, Chan C, Ezrin A, Hottendorf R. (1997) Characterization of renal ischemia-reperfusion injury in rats. J. Pharmacol. Toxicol. Methods 37:1–7.
Remuzzi G, Bertani T. (1998) Pathophysiology of progressive nephropathies. N. Engl. J. Med. 339:1448–56.
Clarkson MR, Gupta S, Murphy M, Martin F, Godson C, Brady HR. (1999) Connective tissue growth factor: a potential stimulus for glomerulosclerosis and tubulointerstitial fibrosis in progressive renal disease. Curr. Opin. Nephrol. Hypertens. 8:543–8.
Ito Y et al. (1998) Expression of connective tissue growth factor in human renal fibrosis. Kidney Int. 53:853–861.
Nakatsuji S, Yamate J, Sakuma S. (1998) Relationship between vimentin expressing renal tubules and interstitial fibrosis in chronic progressive nephropathy in aged rats. Virchows Arch. 433:359–67.
Chai Q, Krag S, Chai S, Ledet T, Wogensen L. (2003) Localisation and phenotypical characterisation of collagen-producing cells in TGF-beta 1-induced renal interstitial fibrosis. Histochem. Cell Biol. 119:267–80.
Zeisberg M et al. (2001) Renal fibrosis: collagen composition and assembly regulates epithelial-mesenchymal transdifferentiation. Am. J. Pathol. 159:1313–21.
Cooker LA et al. (2007) TNF-αlpha, but not IFN-gamma, regulates CCN2 (CTGF), collagen type I, and proliferation in mesangial cells: possible roles in the progression of renal fibrosis. Am. J. Physiol. Renal Physiol. 293: F157–65.
Hogaboam CM, Steinhauser ML, Chensue SW, Kunkel SL. (1998) Novel roles for chemokines and fibroblasts in interstitial fibrosis. Kidney Int. 54:2152–9.
Kondo F, Kondo Y, Gomez-Vargas M, Ogawa N. (1998) Indomethacin inhibits delayed DNAfragmentation of hippocampal CA1 pyramidal neurons after transient forebrain ischemia in gerbils. Brain Res. 791:352–6.
Ko JK, Tang F, Cho CH. (1997) Co-regulation of mucosal prostanoids and substance P by in-domethacin in rat stomachs. Life Sci. 60:PL 277–81.
Mazzali M, Jefferson JA, Ni Z, Vaziri ND, Johnson RJ. (2003) Microvascular and tubulointerstitial injury associated with chronic hypoxia-induced hypertension. Kidney Int. 63:2088–93.
Yokota N, Burne-Taney M, Racusen L, Rabb H. (2003) Contrasting roles for STAT4 and STAT6 signal transduction pathways in murine renal ischemia-reperfusion injury. Am. J. Physiol. Renal Physiol. 285:F319–25.
Marques VP et al. (2006) Influence of TH1/TH2 switched immune response on renal ischemia-reperfusion injury. Nephron Exp. Nephrol. 104:e48–56.
Okada H, Danoff TM, Kalluri R, Neilson EG. (1997) Early role of Fsp1 in epithelial-mesenchymal transformation. Am. J. Physiol. 273:F563–74.
Mazzali M et al. (2002) Effects of cyclosporine in osteopontin null mice. Kidney Int. 62:78–85.
Badid C et al. (2002) Interstitial expression of alpha-SMA: an early marker of chronic renal allograft dysfunction. Nephrol. DM. Transplant. 17:1993–8.
Bottinger EP. (2007) TGF-beta in renal injury and disease. Semin. Nephrol. 27:309–20.
van Es A, Hermans J, van Bockel JH, Persijn GG, van Hooff JP, de Graeff J. (1983) Effect of warm ischemia time and HLA (A and B) matching on renal cadaveric graft survival and rejection episodes. Transplantation 36:255–8.
Zeisberg M, Soubasakos MA, Kalluri R. (2005) Animal models of renal fibrosis. Methods Mol. Med. 117:261–72.
Kalluri R, Sukhatme VP. (2000) Fibrosis and angiogenesis. Curr. Opin. Nephrol. Hypertens. 9:413–8.
Okada H, Strutz F, Danoff TM, Kalluri R, Neilson EG. (1996) Possible mechanisms of renal fibrosis. Contrib. Nephrol. 118:147–54.
Yokoyama H, Wada T, Furuichi K. (2003) Chemokines in renal fibrosis. Contrib. Nephrol. 139:66–89.
Furuichi K, Wada T, Yokoyama H, Kobayashi KI. (2002) Role of Cytokines and Chemokines in Renal Ischemia-Reperfusion Injury. Drug News Perspect. 15:477–82.
Kokkinos MI, Wafai R, Wong MK, Newgreen DF, Thompson EW, Waltham M. (2007) Vimentin and epithelial-mesenchymal transition in human breast cancer: observations in vitro and in vivo. Cells Tissues Organs 185:191–203.
Bravo J et al. (2003) Vimentin and heat shock protein expression are induced in the kidney by angiotensin and by nitric oxide inhibition. Kidney Int. Suppl. S46-51.
Ruster M, Sperschneider H, Funfstuck R, Stein G, Grone HJ. (2004) Differential expression of beta-chemokines MCP-1 and RANTES and their receptors CCR1, CCR2, CCR5 in acute rejection and chronic allograft nephropathy of human renal allografts. Clin. Nephrol. 61:30–9.
Gloria MA, Cenedeze MA, Pacheco-Silva A, Camara NO. (2006) The blockade of cyclooxygenases-1 and -2 reduces the effects of hypoxia on endothelial cells. Braz. J. Med. Biol. Res. 39:1189–96.
Lonnemann G, Shapiro L, Engler-Blum G, Muller GA, Koch KM, Dinarello CA. (1995) Cytokines in human renal interstitial fibrosis; I, Interleukin-1 is a paracrine growth factor for cultured fibrosis-derived kidney fibroblasts. Kidney Int. 47:837–44.
Stanimirovic D, Shapiro A, Wong J, Hutchison J, Durkin J. (1997) The induction of ICAM-1 in human cerebromicrovascular endothelial cells (HCEC) by ischemia-like conditions promotes enhanced neutrophil/HCEC adhesion. J. Neuroimmunol. 76:193–205.
Stahl PJ, Felsen D. (2001) Transforming growth factor-beta, basement membrane, and epithelial-mesenchymal transdifferentiation: implications for fibrosis in kidney disease. Am. J. Pathol. 159:1187–92.
Crisman JM, Richards LL, Valach DP, Franzoni DF, Diamond JR. (2001) Chemokine expression in the obstructed kidney. Exp. Nephrol. 9:241–8.
Pichler R et al. (1994) Tubulointerstitial disease in glomerulonephritis. Potential role of osteopontin (uropontin). Am. J. Pathol. 144:915–26.
Eddy AA. (1995) Interstitial macrophages as mediators of renal fibrosis. Exp. Nephrol. 3:76–9.
Diamond JR, Kees-Folts D, Ricardo SD, Pruznak A, Eufemio M. (1995) Early and persistent up-regulated expression of renal cortical osteopontin in experimental hydronephrosis. Am. J. Pathol. 146:1455–66.
Ophascharoensuk V et al. (1999) Obstructive uropathy in the mouse: role of osteopontin in interstitial fibrosis and apoptosis. Kidney Int. 56:571–80.
Zeisberg M et al. (2003) Bone morphogenic protein-7 inhibits progression of chronic renal fibrosis associated with two genetic mouse models. Am. J. Physiol. Renal Physiol 285: F1060–7.
Terzi F et al. (1997) Normal tubular regeneration and differentiation of the post-ischemic kidney in mice lacking vimentin. Am. J. Pathol. 150:1361–71.
Nguyen TQ, Goldschmeding R. (2008) Bone morphogenetic protein-7 and connective tissue growth factor: novel targets for treatment of renal fibrosis? Pharm Res. 25:2416–26.
Damiao MJ et al. (2007) The effects of rapamycin in the progression of renal fibrosis. Transplant Proc 39:457–9.
Yokoi H et al. (2004) Reduction in connective tissue growth factor by antisense treatment ameliorates renal tubulointerstitial fibrosis. J. Am. Soc. Nephrol. 15:1430–40.
Deng J et al. (2001) Interleukin-10 inhibits ischemic and cisplatin-induced acute renal injury. Kidney Int. 60:2118–28.
Takahira R, Yonemura K, Fujise Y, Hishida A. (2001) Dexamethasone attenuates neutrophil infiltration in the rat kidney in ischemia/reperfusion injury: the possible role of nitroxyl. Free Radic. Biol. Med. 31:809–15.
Salahudeen AK, Haider N, May W. (2004) Cold ischemia and the reduced long-term survival of cadaveric renal allografts. Kidney Int. 65:713–8.
Harizi H, Gualde N. (2006) Pivotal role of PGE2 and IL-10 in the cross-regulation of dendritic cell-derived inflammatory mediators. Cell Mol. Immunol. 3:271–7.
Treffkorn L, Scheibe R, Maruyama T, Dieter P. (2004) PGE2 exerts its effect on the LPS-induced release of TNF-αlpha, ET-1, IL-1alpha, IL-6 and IL-10 via the EP2 and EP4 receptor in rat liver macrophages. Prostaglandins Other Lipid Mediat. 74:113–23.
Acknowledgments
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico and Fundação de Amparo à Pesquisa of São Paulo (Grant numbers: 04/08311-4, 04/13449-7, 06/03982-5, and 07/07139-3).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Feitoza, C.Q., Gonçalves, G.M., Semedo, P. et al. Inhibition of COX 1 and 2 prior to Renal Ischemia/Reperfusion Injury Decreases the Development of Fibrosis. Mol Med 14, 724–730 (2008). https://doi.org/10.2119/2008-00064.Feitoza
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/2008-00064.Feitoza