
Abstract
Immunoglobulin κ (IGK) locus rearrangements were analyzed in parallel on cDNA/genomic DNA in 188 κ- and 103 λ-chronic lymphocytic leukemia (CLL) cases. IGKV-KDE and IGKJ-C-intron-KDE rearrangements were also analyzed on genomic DNA. In κ-CLL, only 3 of 188 cases carried double in-frame IGKV-J transcripts: in such cases, the possibility that leukemic cells expressed more than one k chain cannot be excluded. Twenty-eight κ-CLL cases also carried nonexpressed (nontranscribed and/or out-of-frame) IGKV-J rearrangements. Taking IGKV-J, IGKV-KDE, and IGKJ-C-intron-KDE rearrangements together, 38% of κ-CLL cases carried biallelic IGK locus rearrangements. In λ-CLL, 69 IGKV-J rearrangements were detected in 64 of 103 cases (62%); 24 rearrangements (38.2%) were in-frame. Four cases carried in-frame IGKV-J transcripts but retained monotypic light-chain expression, suggesting posttranscriptional regulation of allelic exclusion. In all, taking IGKV-J, IGKV-KDE, and IGKJ-C-intron-KDE rearrangements together, 97% of λ-CLL cases had at least 1 rearranged IGK allele, in keeping with normal cells. IG repertoire comparisons in κ- versus λ-CLL revealed that CLL precursor cells tried many rearrangements on the same IGK allele before they became λ producers. Thirteen of 28 and 26 of 69 non-expressed sequences in, respectively, κ- or λ-CLL had < 100% homology to germline. This finding might be considered as evidence for secondary rearrangements occurring after the onset of somatic hypermutation, at least in some cases. The inactivation of potentially functional IGKV-J joints by secondary rearrangements indicates active receptor editing in CLL and provides further evidence for the role of antigen in CLL immunopathogenesis.
Similar content being viewed by others


Introduction
The most widely accepted model for regulation of isotype exclusion at immunoglobulin (Ig) light-chain loci (ordered model) proposes that immunoglobulin λ (IGL) rearrangements occur only after nonfunctional immunoglobulin k (IGK) rearrangements (1). In this context, single-cell PCR analysis of peripheral blood B cells from healthy individuals has demonstrated that whereas only 2% to 3% of κ-expressing cells carried IGLV-J junctions, nearly all λ-expressing B cells had rearranged IGK genes and bore IGKV-J junctions; furthermore, a significant proportion of these joints (∼30%) were rearranged in-frame (2). The inactivation of functional IGK rearrangements during B-cell development probably permits secondary rearrangements at the light-chain loci.
Antigen encounter in the bone marrow may reinduce RAG gene expression so that autoreactive normal B cells can be rescued through secondary rearrangements and receptor editing (3–5). The IGK locus permits extensive editing because direct V-to-J joining facilitates nested recombination (V-to-J recombination occurs between V and J elements flanking existing V-J joints, thereby inactivating and replacing that rearrangement). Similarly, the strikingly progressive nature of light-chain gene rearrangements, in which the use of upstream IGKJ genes precedes that of downstream genes and κ expression precedes λ, is ideally suited to editing; indeed, κ-chain gene structure does facilitate very efficient tolerance-induced editing (1,4). Occasionally, tolerance-induced editing can also promote the appearance of cells carrying multiple receptors that dilute out the autoreactive specificity (6).
IGK loci are often inactivated in κ and mainly in λ light chain-expressing B cells by rearrangement to the κ-deleting element (KDE), which is located 3′ to the IGKC gene; the KDE is rearranged either to an unrearranged IGKV gene upstream of a IGKV-J junction or to a recombination signal sequence in the IGKJ-C intron (7–9). Because the KDE encodes no protein, rearrangements to KDE are nonproductive; it has been proposed that the KDE has no other purpose than to inactivate IGK genes, many of which were previously functional (4,10–11). Given that KDE rearrangements appear to occur concurrently with IGL locus recombination, they may be predicted to clear the way for λ expression (1,3,11–13).
“Natural”autoantibodies are often found in serum of animals; they are mostly IgM unmutated autoantibodies and independent of T-cell help for secretion (14–16). Such antibodies additionally account for a large proportion of the early human B-cell repertoire (17). Although the escape of autoreactive B cells from clonal deletion or anergy is considered to be detrimental to the host, rescuing of certain autoreactive B cells confers a clear immunological advantage because these cells are also highly protective against microbial pathogens (e.g., S. pneumoniae and filariae) (18–20). Apart from receptor editing, one further mechanism employed by autore-active B cells in an effort to escape clonal deletion is the expression of multiple antigen receptors (“allelic inclusion”) (21).
Allelic inclusion is a frequent feature in the normal marginal zone (6), a “reservoir”of B cells with autoreactive specificities. As shown by analysis of IGH genes, it is also relatively frequent in chronic lymphocytic leukemia (CLL) (22). CLL Ig heavy and light chains have unique and often “autoimmune-like”features (23,24); also, subsets of CLL cases have remarkably similar immunoglobulins (“stereotyped B-cell receptors”), suggesting recognition of individual, discrete antigens or classes of structurally similar epitopes (24–31). Furthermore, CLL cells frequently produce “natural”or polyreactive IgM autoantibodies (32–37). Altogether, the similarity between B cells producing natural autoantibodies and CLL malignant B cells could mean that the process of positive selection of natural autoreactive B cells may carry a risk for malignant transformation (38).
In this study, we analyzed productive and nonproductive IGK-locus rearrangements in a series of 291 patients with CLL. Our results indicate that a small subset of CLL patients have cells that may express more than one light-chain allele, indicating that allelic exclusion of light chains is not absolute. IG repertoire comparisons in κ- versus λ-CLL revealed that CLL precursor cells tried many rearrangements on the same IGK allele before they became λ producers. Finally, the inactivation of preexisting functional rearrangements by secondary IGK recombination events could be considered as evidence that, similar to normal B cells, receptor editing contributes significantly to the expressed CLL Ig repertoire.
Patients and Methods
Patient Samples
Peripheral blood (PB) samples were collected from 291 CLL patients. All patients met the diagnostic criteria of the National Cancer Institute Working Group (39). Included in the analysis were 178 men and 113 women with a median age of 65 years (range, 29 to 79 years). Most patients were at early clinical stages by Rai classification (stage 0, 143; I, 54; II, 64; III, 14; and IV, 16 patients). The median follow-up time was 43 months (range, 3 to 261 months). Written informed consent was obtained at study entry. The study was approved by the local Ethics Review Committee of each institution.
Flow Cytometry and Immunohistochemistry
The analysis was performed in whole blood samples, and the monoclonal antibodies were titrated according to manufacturer instructions. Antibodies directed against the following antigens were applied: CD19, CD20, CD5, CD38, CD23, CD43, FMC7, and K/λ/IgM/IgD/IgG/IgA (surface/intracellular). The percentage of positive cells for each CD marker was estimated by gating the CD19+ cluster. A case was considered to be κ- or λ-expressing if the ratio of κ to λ expression on CD19+ cells was greater than 6 or less than 0.3, respectively. Based on the above definitions, 189 cases expressed κ light chain and 103 cases expressed λ light chain. In all cases, the tumor load was at least 70%.
Immunostaining for cytoplasmic κ/λ light chains on formalin-fixed paraffin-embedded bone marrow biopsy sections was performed with the ABC technique using polyclonal antisera raised in rabbit (Dako, Copenhagen, Denmark).
PCR Amplification of IGK Locus Rearrangements and Sequence Analysis
High-molecular-weight DNA was isolated from peripheral blood mononuclear cells with the QIAamp DNA Blood Kit (Qiagen Inc., Valencia, CA, USA). Total cellular RNA isolation, cDNA preparation (after DNAse treatment) and RT-PCR amplification of IGHV-D-J/IGKV-J/IGLV-J rearrangements was performed as previously described (24,31). In all cases, IGKV-J rearrangements were examined on both cDNA and genomic DNA.
IGKV-to-KDE as well as IGKJ-C-intron-to-KDE rearrangements (Figure 1) were amplified by PCR on genomic DNA, using appropriate primers (40). When a case carried two KDE rearrangements, either in the form of coexisting IGKV-KDE and IGKJ-C-intron-KDE junctions or as double IGKV-KDE or IGKJ-C-intron-KDE junctions, the case was considered to have rearranged both IGK alleles.

Schematic overview of potential classical rearrangements in the human IGK locus.
Direct sequence analysis of RT-PCR and PCR products was performed as previously described (24). Ig heavy- and light-chain sequence data were analyzed using the IMGT information system (IMGT, https://doi.org/imgt.cines.fr) (41,42). IGKV/KDE and IGKJ-C-intron-KDE junctions were analyzed using the Basic Local Alignment Search Tool (BLAST) database (https://doi.org/www.ncbi.nlm.nih. gov/Igblast, National Center for Biotechnology Information, Bethesda, MD, USA).
Results
IGKV-J Rearrangements in κ-Expressing CLL
The expressed IGKV-J repertoire of 179 of 188 cases in the present study has been reported previously (24). In κ-CLL, 219 IGKV-J clonal rearrangements were amplified in 188 cases. Thirty-one cases (16.5%) carried double rearrangements; both rearrangements were in-frame in 14 of 188 cases (7.44%). Double in-frame IGKV-J transcripts were detected in only 3 of 188 cases (1.6%).
Twenty-one functional IGKV genes belonging to the IGKV1-4 subgroups were identified in 191 transcribed, in-frame IGKV-J rearrangements. IGKV3-20 was the most frequent IGKV gene (29 of 191 sequences; 15.3%), followed by IGKV1-39/1D-39 (25 of 191; 13.1%), IGKV1-5 (20 of 191; 10.5%), IGKV4-1 (19 of 191; 10%), IGKV2-30 and IGKV3-11 (13 of 191 sequences each; 6.8%) (Supplemental Table). IGKJ2 was the most frequent gene, followed by IGKJ1; collectively, IGKJ3-5 gene usage was observed in 60 of 191 rearrangements (31.4%).
Fifteen different IGKV genes were identified in the remaining 28 IGKV-J rearrangements, which were presumably nonexpressed as they were either nontranscribed (23 of 28 cases; 82%) and/or out-of-frame (18 of 28 cases; 64%); 6 of 18 out-of-frame IGKV-J rearrangements were transcribed. The most frequent genes were IGKV4-1 (9 of 28 sequences; 32%) and IGKV2-30 (3 of 28; 10.7%) (Supplemental Table). Significant differences were identified between expressed versus nonexpressed IGKV-J rearrangements; the IGKV1-39/1D-39 and IGKV1-5 genes were more frequent in the expressed repertoire (χ2 test: P < 0.05 for both), contrasting with the IGKV4-1 gene, which predominated among nonexpressed rearrangements (χ2 test: P = 0.01).
IGKV-J Rearrangements in λ-CLL
Clonal IGKV-J rearrangements were amplified in 64 of 103 cases (62%). Five cases had two different rearrangements; in such cases, each rearrangement involved a different IGKV gene. Nineteen different genes of the IGKV1-5 subgroups were used; IGVK4-1 was the most frequent IGKV gene (15 of 69 sequences; 21.7%), followed by 1–33/1D-33 (9 of 69; 13%), 1–39/1D-39, 2–30, and 1–16 (7 of 69 rearrangements each; 10%) (Supplemental Table). Collectively, the aforementioned IGKV genes comprised 65.2% of all IGKV-J rearrangements.
IGKV-J transcripts were detected in 10 of 64 λ-CLL cases with IGKV-J rearrangements; 4 of 10 transcripts were in-frame. In the 4 λ-CLL cases with transcribed, in-frame IGKV-J rearrangements, flow cytometry and immunohistochemistry demonstrated that monotypic IG expression was still maintained: in particular, malignant B cells were negative for either cytoplasmic or surface K light chains. Five of 10 λ-CLL cases with transcribed IGKV-J rearrangements had both KDE sequences in germline configuration; the remainder had PCR evidence for monoallelic KDE rearrangements (2 cases with IGKV-KDE rearrangements and 3 cases with IGKJ-C-intron-KDE rearrangements).
Significant IGKV repertoire differences were identified between expressed versus nonexpressed IGKV-J rearrangements in κ- versus λ-CLL, respectively (Supplemental Table). The IGKV3-20 and IGKV1-5 genes predominated in the expressed repertoire in κ-CLL (χ2 test: P < 0.01). In contrast, the IGKV4-1, IGKV1-16, IGKV1-33/1D-33, and IGKV1-37/1D-37 genes were significantly overrepresented in nonexpressed IGKV-J rearrangements in λ-CLL (χ2 test: P < 0.05).
Complete analysis of the KCDR3 region was possible in 62 of 69 IGKV-J rearrangements. Twenty-four IGKV-J junctions (38.7%) were in-frame. Among in-frame, nontranscribed rearrangements, one utilized the IGKV2-29 pseudogene, whereas three utilized the IGKV1-37/1D-37 gene, with a germline-encoded glycine instead of cysteine at position 104 in FR3-IMGT. Three in-frame rearrangements were functionally inactivated due to extensive 5′ or 3′ exonuclease activity, which led to elimination of cysteine at position 104 in FR3-IMGT or phenylalanine at position 118 in FR4-IMGT. IGKJ4 was the most frequent gene, followed by IGKJ2; collectively, IGKJ3-5 gene usage was observed in 34 of 62 rearrangements (54.8%).
Alignments and Comparisons to Public Database Sequences
IGKV-J sequences from our series were compared with all IGKV-J sequences of the IMGT/LIGM-DB sequence database (https://doi.org/imgt.cines.fr/cgi-bin/IMGTlect.jv?). This analysis identified subsets of very similar sequences with closely homologous CDR3 regions comprising predominantly or exclusively CLL sequences. Four such subsets identified among expressed IGKV-J rearrangements in κ-CLL were recently reported by our group (24).
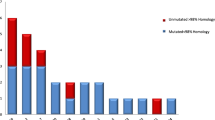
In the present series, we identified two homologous transcribed, in-frame IGKV1-17-IGKJ4 rearrangements. The first was detected in a λ-CLL case with an IGHV6-1-expressing heavy chain. The second was amplified along with a transcribed, inframe IGKV1-39/1D-39-IGKJ1 rearrangement in an IgG+ female CLL patient expressing an unmutated IGHV4-39/IGHD6-13/IGHJ2 heavy chain. IgG+ cases with homologous, unmutated IGHV4-39-IGKV1-39/1D-39 receptors have been reported recently by several groups, including ours (24,27,30). The two homologous IGKV1-17-IGKJ4 rearrangements from our series shared an identical KCDR3 (LQHNSYPLT) with two public sequences (GenBank accession num. L33034 and AF007572) obtained from anti-DNA antibodies with antiplatelet specificity (Figure 2) (43,44). These antibodies utilized IGHV3 subgroup genes (IGHV3-11 and IGHV3-23, respectively) (43,44) and carried unrelated HCDR3s.

Amino acid sequences of two novel subsets of homologous IGKV-J rearrangements identified after alignment of the sequences reported in the present series to public-database sequences. (A) IGKV1-17/IGKJ4 rearrangements. P1597 derives from a λ-CLL case whereas P242 derives from a κ-CLL case, both from the present series; L33034 and AF007572 derive from cells with antiplatelet activity. (B) IGKV1-39-1D-39/IGKVJ2 rearrangements. N1857 derives from a λ-CLL case from the present series; L12108 derives from a cell with anti-TPO activity.
One mutated, nontranscribed, in-frame IGKV1-39/1D-39-IGKJ2 rearrangement from a λ-CLL case of the present series (expressing an IGHV3-30 heavy chain) was homologous to the κ light chain (L12108) of an antithyroid peroxidase (anti-TPO) antibody (Figure 2) (45); this antibody utilized an IGHV1-3-expressing heavy chain (L12109).
IGK Locus Rearrangements to the κ Deleting Element (KDE)
In κ-CLL, 170 cases were examined for IGKV-KDE and IGKJ-C-intron-KDE rearrangements. Fifty cases (29.4%) had PCR evidence for monoallelic KDE rearrangements, leading to IGK locus inactivation on that particular allele. In detail, 24 cases (14%) carried IGKV-KDE rearrangements. Sequencing analysis was successful for 22 of 24 IGKV-KDE rearrangements and revealed frequent usage of the IGKV1D-43 gene (4 of 22 cases; 18.1%), which was rare in either the expressed or nonexpressed CLL repertoire (Supplemental Table). IGKJ-C-intron-KDE rearrangements were amplified in 26 of 170 cases (15.3%). All three κ-CLL cases with double inframe IGKV-J transcripts were negative for KDE rearrangements.
In λ-CLL, 60 of 100 cases (60%) carried IGKV-KDE rearrangements; 1 case carried biallelic IGKV-KDE rearrangements. Sequencing data were available for 56 of 60 rearrangements. The IGKV3-20 and 2–30 genes predominated (9 of 56 and 8 of 56 cases, respectively) (Supplemental Table), whereas the IGKV4-1 gene was used in only 3 of 56 sequences. IGKJ-C-intron-KDE rearrangements were amplified in 54 of 100 cases (54%); 1 case carried biallelic rearrangements.
Six λ-CLL cases from our series belonged to subsets with stereotyped receptors (BCRs). Four of 6 cases carried IGHV3-21/IGLV3-21 BCRs in a fashion previously reported by several groups, including ours (24,26,31), whereas the remaining 2 cases carried stereotyped IGHV4-b/IGLV1-44 BCRs (24). PCR evidence for at least monoallelic IGK locus rearrangement was obtained in all 6 cases. Specifically, 4 cases carried IGKV-J rearrangements, of which 3 were in-frame; furthermore, KDE rearrangements were also detected in 4 of 6 cases.
Multiple Rearrangements of the IGK Locus in CLL
Among 170 κ-CLL cases analyzed for both IGKV-J and KDE rearrangements, 104 cases (61.2%) carried monoallelic IGK locus rearrangements in the form of a single IGKV-J joint (Table 1). Evidence for biallelic IGK locus rearrangements was obtained in (i) 50 of 170 cases (29.4%) with a KDE rearrangement along with an expressed IGKV-J rearrangement (Table 1) and (ii) 3 of 170 cases (1.8%) with double IGKV-J rearrangements utilizing the same IGKJ gene in both IGKV-J joints. In the remaining 13 κ-CLL cases with double IGKV-J joints, based on IGKV and IGKJ gene repertoire analysis as well as location and mode of recombination (deletional versus inversional) of the IGKV genes identified in both rearrangements, evidence for biallelic IGK locus rearrangements was obtained in 12 of 13 cases; in 1 case, both rearrangements could have occurred sequentially on one or both IGK alleles.
Among 100 λ-CLL cases analyzed for both IGKV-J and KDE rearrangements, only 3 (3%) had no evidence for IGK locus rearrangements; 33 cases carried 1 rearrangement on the IGK locus, whereas 44 and 20 cases, respectively, carried 2 or 3 IGK locus rearrangements (Table 1). In 19 of 64 λ-CLL cases with multiple IGK locus rearrangements, based on IGKV and IGKJ gene usage as well as type of recombination to the KDE (IGKV-KDE versus IGKJ-C-intron-KDE), it would be impossible to define with certainty whether the detected rearrangements occurred on one or both IGK alleles. Following the reasoning described above, the remaining (45 of 64) cases had PCR evidence in support of biallelic IGK locus rearrangements.
In λ-CLL cases with biallelic KDE rearrangements, the IGKV locus was rendered inactive, thus precluding expression of a preexisting, in-frame IGKV-J junction. This might account for failure to express a functional κ light chain at least for the significant proportion of cases (9 of 18) bearing in-frame IGKV-J rearrangements in the context of biallelic KDE rearrangements. None of those IGKV-J rearrangements was transcribed.
Somatic Hypermutation Analysis
In κ-CLL, 62, 36, and 93 transcribed, in-frame IGKV-J rearrangements had, respectively, 100%, 98–99.6%, and < 98% homology to the closest germline gene. Among nonexpressed IGKV-J rearrangements in κ-CLL, 15, 3, and 10 sequences had, respectively, 100%, 98–99.6%, and < 98% homology to the closest germline gene. Four of 6 out-of-frame IGKV-J transcripts in κ-CLL were heavily mutated and often carried deletions/insertions (3 of 6 cases) or one or more stop codons in the coding sequence (4 of 6 cases). High replacement-to-silent mutation ratios (R/S > 2) were observed in the framework regions (FRs), indicating that these rearrangements could not be positively selected.
In λ-CLL 43, 22, and 4 IGKV-J rearrangements had, respectively, 100%, 98–99.6%, and < 98% homology to the closest germline gene. Two of 4 nonexpressed rearrangements with < 98% homology were in-frame; both were homologous to sequences from autoantibodies, as mentioned above.
Please note that supplementary information is available on the Molecular Medicine website (https://doi.org/www.molmed.org).
Discussion
We analyzed IGK locus rearrangements in a series of 291 CLL patients. Four λ-expressing CLL cases carried in-frame IGKV-J transcripts along with in-frame IGLV-J transcripts but still retained monotypic Ig expression: therefore, in these cases allelic exclusion was probably regulated at the posttranscriptional level. In κ-CLL, double in-frame transcripts were detected in only 3 of 188 cases; in such cases, the possibility that leukemic cells expressed more than one κ light chain cannot be excluded. As previously shown, leukemic cells of some CLL patients may express more than one heavy chain due to lack of allelic exclusion (22). Here, we provide evidence that allelic exclusion at the IGK locus may also not be absolute in a minority of CLL cases. This could be interpreted as evidence for continued Ig gene rearrangements either in the context of receptor editing (2–5) or as a result of selection to maintain B cells with multiple binding specificities (21–22).
In λ-CLL, in keeping with normal B cells (2), nearly all cases (97%) carried IGK locus rearrangements. IGKV-J junctions were detected in 64 of 103 cases (62%). This is probably an underestimate, as some IGKV-J junctions may not have been successfully amplified. Furthermore, preexisting IGKV-J joints may have been deleted by IGKV-KDE rearrangements on the same allele: this could be the case for 16 of 100 λ-CLL cases carrying only IGKV-KDE rearrangements. The possibility that rearrangements from contaminating nonleukemic cells were amplified cannot be formally excluded. Nevertheless it seems very remote on the following grounds: (i) the tumor load was always > 70%; (ii) all experiments were repeated at least 3 times with identical results; and (iii) identical results were obtained by analysis of blood samples obtained at different times (repeat samples were available in 180 of 291 cases).
Similar to normal λ-expressing B cells (2), a significant percentage (38.7%) of IGKV-J rearrangements in λ-CLL cases of our series were in-frame. In a previous analysis of IGK locus rearrangements in λ-expressing chronic B cell leukemias (including CLL, prolymphocytic leukemia, hairy cell leukemia), IGKV-J junctions were detected in only 7 of 60 cases (12%) (32). The discrepant results between that study and our series as regards the percentage of IGKV-J-bearing cases could probably be attributed to differences in methodology (Southern blotting versus PCR), sample size, and homogeneity of patient groups.
Three λ-CLL cases of the present series carried in-frame IGKV-J rearrangements utilizing the defective IGKV1-37/1D-37 gene. This gene has been reported in the repertoire of a patient with systemic lupus erythematosus (SLE) (46). However, that study was DNA-based and would discriminate only between inframe vs. out-of-frame rearrangements, hindering definitive conclusions about the expressed repertoire.
Several IGKV genes utilized in nonexpressed IGKV-J rearrangements in either κ- or λ-CLL cases of the present series have been found as a component of autoantibodies (e.g., IGKV1-33/1D-33, 1–39/1D-39, 3–20, 1–17) (although this may merely reflect their overall frequency in the repertoire [47]). It is perhaps relevant that 2 IGKV1-17 and 1 IGKV1-39/1D-39 in-frame rearrangements from our series shared identical KCDR3s with public autoantibody sequences (43,44). The IGKV1-17 gene, rarely expressed by normal cells, is critically implicated in the development of SLE-nephritis (48). It has been argued that normal B cells may edit IGKV1-17 rearrangements by receptor editing so as to avoid self-reactivity, whereas SLE B cells may have a defect in this mechanism (48). In this context, it would perhaps be possible that primary IGK rearrangements with autoreactive potential in CLL clonogenic cells were followed by a secondary light-chain rearrangement in the context of a receptor “dilution”(21) or “editing”process (1,49–51), which would eventually lead either to expression of more than one κ light chain or to a shift to λ production.
An intriguing finding of this study was the frequent usage of downstream IGKJ genes (IGKJ3-5) in λ-CLL, contrasting with both normal (47,52) and CLL (24) repertoires. In autoantibody transgenic mice, usage of downstream IGKJ genes is a hallmark of autoreactive cells that escape deletion by secondary rearrangements that alter specificity (53). The significant bias for downstream IGKJ genes in λ-CLL might be considered an indication that the clonogenic cells tried many rearrangements on the same IGK allele before they became λ producers (2,14). Evidence for prolonged light-chain recombination in our series is also provided by the increased usage of more IGKJ-distal genes in IGKV-KDE versus IGKV-J rearrangements (especially in λ-CLL). This may be attributed to a shift in accessibility to the recombinase from the J-proximal to the J-distal parts of the IGK locus at more advanced stages in B-cell ontogeny (54,55).
In both κ- and λ-CLL, a minority of nonexpressed (in-frame/out-of-frame) IGKV-J rearrangements were < 100% homologous to germline. The existence of mutated, in-frame, nonexpressed IGKV-J rearrangements might be considered as indirect evidence for secondary rearrangements after the onset of somatic hypermutation (SHM). Chronic antigen stimulation may trigger secondary rearrangements at the IGH or the IGK/IGL loci (53,56–66). Although the occurrence of secondary rearrangements in normal peripheral B cells was considered as doubtful or—at best—limited (67), several studies indicate that secondary rearrangements may occur following SHM, particularly in a setting of autoimmunity or neoplasia (58,60,62,66,68). Nonetheless, SHM features in nonexpressed rearrangements of the present series (high R/S mutation ratios in FRs, stop codons, insertions/deletions) could be more plausibly attributed to a “bystander mutagenesis”effect, whereby nonexpressed rearrangements are mutated without selection for expression of a functional antigen receptor, as previously reported for either normal or neoplastic cells (69–71).
Defective mRNA transcripts are normally detected and destroyed by nonsense-mediated decay (NMD) (72,73). B and T cells have very efficient NMD mechanisms to avoid production of truncated IG and TCR chains (74,75). Occasionally, defective transcripts may persist: this has been reported previously in a setting of autoimmunity (76) or immunodeficiency (77). The results of this series show that defective Ig transcripts may also be detected in CLL. Nonetheless, their low frequency (12 of 291 cases, 4%) indicates operation of active mRNA surveillance mechanisms (NMD, other?) in CLL cells.
In conclusion, the present study confirms IGK locus rearrangements in the vast majority of λ-CLL cases and indicates that the light-chain rearrangement “hierarchy”in CLL is not inherently different from that in normal cells. The results reported here suggest prolonged IGK locus recombination before CLL precursors became λ-producers. Finally, the inactivation of preexisting functional IGKV-J joints by secondary rearrangements indicates active receptor editing in CLL and provides further evidence for the role of antigen in the emergence and maintenance of the leukemic clone, at least in a proportion of CLL cases.
References
Nemazee D. (2000) Receptor selection in B and T lymphocytes. Annu. Rev. Immunol. 18:19–51.
Brauninger A, Goossens T, Rajewsky K, Kuppers R. (2001) Regulation of immunoglobulin light chain gene rearrangements during early B cell development in the human. Eur. J. Immunol. 31:3631–7.
Prak EL, Weigert M. (1995) Light chain replacement: a new model for antibody gene rearrangement. J. Exp. Med. 182:541–8.
Retter MW, Nemazee D. (1998) Receptor editing occurs frequently during normal B cell development. J. Exp. Med. 188:1231–8.
Ghia P, Gratwohl A, Signer E, Winkler TH, Melchers F, Rolink AG. (1995) Immature B cells from human and mouse bone marrow can change their surface light chain expression. Eur. J. Immunol. 25:3108–14.
Li Y, Li H, Weigert M. (2002) Autoreactive B cells in the marginal zone that express dual receptors. J. Exp. Med. 195:181–8.
Siminovitch KA, Moore MW, Durdik J, Selsing E. (1987) The human kappa deleting element and the mouse recombining segment share DNA sequence homology. Nucleic Acids Res. 15:2699–705.
Klobeck HG, Zachau HG. (1986) The human CK gene segment and the kappa deleting element are closely linked. Nucleic Acids Res. 14:4591–603.
Durdik J, Moore MW, Selsing E. (1984) Novel kappa light-chain gene rearrangements in mouse lambda light chain producing B lymphocytes. Nature 307:749–52.
Dunda O, Corcos D. (1997) Recombining sequence recombination in normal kappa-chain-expressing B cells. J. Immunol. 159:4362–6.
Daitch LE, Moore MW, Persiani DM, Durdik JM, Selsing E. (1992) Transcription and recombination of the murine RS element. J. Immunol. 149:83–40.
Muller B, Reth M. (1988) Ordered activation of the Ig lambda locus in Abelson B cell lines. J. Exp. Med. 168:2131–7.
Shimizu T, Iwasato T, Yamagishi H. (1991) Deletions of immunoglobulin C kappa region characterized by the circular excision products in mouse splenocytes. J. Exp. Med. 173:1065–72.
Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig MC. (2003) Predominant autoantibody production by early human B cell precursors. Science. 301:1374–7.
Hardy RR, Wei CJ, Hayakawa K. (2004) Selection during development of VH11+ B cells: a model for natural autoantibody-producing CD5+ B cells. Immunol. Rev. 197:60–74.
Bendelac A, Bonneville M, Kearney J. (2001) Autoreactivity by design: innate B and T lymphocytes. Nat. Rev. Immunol. 1:77–86.
Hayakawa K, Asano M, Shinton SA, et al. (1999) Positive selection of natural autoreactive B cells. Science. 285:113–6.
Kruetzmann S, Rosado MM, Weber H, et al. (2003) Human immunoglobulin M memory B cells controlling Streptococcus pneumoniae infections are generated in the spleen. J. Exp. Med. 197:939–45.
MacLennan IC, Toellner KM, Cunningham AF, et al. (2003) Extrafollicular antibody responses. Immunol. Rev. 194:8–18.
Pillai S, Cariappa A, Moran ST. (2005) Marginal zone B cells. Annu. Rev. Immunol. 23:161–96.
Kenny JJ, Rezanka LJ, Lustig A, Fischer RT, Yoder J, Marshall S, Longo DL. (2000) Autoreactive B cells escape clonal deletion by expressing multiple antigen receptors. J. Immunol. 164:4111–9.
Rassenti LZ, Kipps TJ. (1997) Lack of allelic exclusion in B cell chronic lymphocytic leukemia. J. Exp. Med. 185:1435–45.
Chiorazzi N, Rai KR, Ferrarini M. (2005) Chronic lymphocytic leukemia. N. Engl. J. Med. 352:804–15.
Stamatopoulos K, Belessi C, Hadzidimitriou A, et al. (2005) Immunoglobulin light chain repertoire in chronic lymphocytic leukemia. Blood 106:3575–83.
Widhopf GF, Kipps TJ. (2001) Normal B cells express 51p1-encoded Ig heavy chains that are distinct from those expressed by chronic lymphocytic leukemia B cells. J. Immunol. 166:95–102.
Tobin G, Thunberg U, Johnson A, et al. (2003) Chronic lymphocytic leukemias utilizing the VH3-21 gene display highly restricted Vλ2-14 gene use and homologous CDR3s: implicating recognition of a common antigen epitope. Blood 101:4952–7.
Ghiotto F, Fais F, Valetto A, et al. (2004) Remarkably similar antigen receptors among a subset of patients with chronic lymphocytic leukemia. J. Clin. Invest. 113:1008–16.
Widhopf GF, Rassenti LZ, Toy TL, Gribben JG, Wierda WG, Kipps TJ. (2004) Chronic lymphocytic leukemia B cells of more than 1% of patients express virtually identical immunoglobulins. Blood 104:2499–504.
Messmer BT, Albesiano E, Efremov DG, et al. (2004) Multiple distinct sets of stereotyped antigen receptors indicate a role for antigen in promoting chronic lymphocytic leukemia. J. Exp. Med. 200:519–25.
Tobin G, Thunberg U, Karlsson K, et al. (2004) Subsets with restricted immunoglobulin gene rearrangement features indicate a role for antigen selection in the development of chronic lymphocytic leukemia. Blood 104:2879–85.
Ghia P, Stamatopoulos K, Belessi C, et al. (2005) Geographic patterns and pathogenetic implications of IGHV gene usage in chronic lymphocytic leukemia: the lesson of the IGHV3-21 gene. Blood 105:1678–85.
Broker BM, Klajman A, Youinou P, et al. (1988) Chronic lymphocytic leukemic (CLL) cells secrete multispecific autoantibodies. J. Autoimmun. 1:469–81.
Kipps TJ, Robbins BA, Kuster P, Carson DA. (1988) Autoantibody-associated cross-reactive idiotypes expressed at high frequency in chronic lymphocytic leukemia relative to B-cell lymphomas of follicular center cell origin. Blood 72:422–8.
Kipps TJ, Tomhave E, Chen PP, Carson DA. (1988) Autoantibody-associated kappa light chain variable region gene expressed in chronic lymphocytic leukemia with little or no somatic mutation: implications for etiology and immunotherapy. J. Exp. Med. 167:840–52.
Sthoeger ZM, Wakai M, Tse DB, et al. (1989) Production of autoantibodies by CD5-expressing B lymphocytes from patients with chronic lymphocytic leukemia. J. Exp. Med. 169:255–68.
Borche L, Lim A, Binet JL, Dighiero G. (1990) Evidence that chronic lymphocytic leukemia B lymphocytes are frequently committed to production of natural autoantibodies. Blood 76:562–9.
Rassenti LZ, Pratt LF, Chen PP, Carson DA, Kipps TJ. (1991) Autoantibody-encoding kappa L chain genes frequently rearranged in lambda L chain-expressing chronic lymphocytic leukemia. J. Immunol. 147:1060–6.
Widhopf GF, Brinson DC, Kipps TJ, Tighe H. (2004) Transgenic expression of a human polyreactive Ig expressed in chronic lymphocytic leukemia generates memory-type B cells that respond to nonspecific immune activation. J. Immunol. 172:2092–9.
Cheson BD, Bennett JM, Grever M, Kay N, Keating MJ, O’Brien S, Rai KR. (1996) National Cancer Institute-sponsored Working Group guidelines for chronic lymphocytic leukemia: revised guidelines for diagnosis and treatment. Blood 87:4990–7.
van der Burg M, Tumkaya T, Boerma M, de Bruin-Versteeg S, Langerak AW, van Dongen JJ. (2001) Ordered recombination of immunoglobulin light chain genes occurs at the IGK locus but seems less strict at the IGL locus. Blood 97:1001–8.
Lefranc MP, Giudicelli V, Kaas Q, et al. (2005) IMGT, the international ImMunoGeneTics information system. Nucleic Acids Res. 33Database Issue: D593–7.
Giudicelli V, Chaume D, Lefranc MP. (2004) IMGT/V-QUEST, an integrated software for immunoglobulin and T cell receptor V-J and V-D-J rearrangement analysis. Nucleic Acids Res. 32:W435–40.
Lampman GW, Furie B, Schwartz RS, Stollar BD, Furie BC. (1989) Amino acid sequence of a platelet-binding human anti-DNA monoclonal autoantibody. Blood 74:262–9.
Rioux JD, Zdarsky E, Newkirk MM, Rauch J. (1995) Anti-DNA and anti-platelet specificities of SLE-derived autoantibodies: evidence for CDR2H mutations and CDR3H motifs. Mol. Immunol. 32:683–96.
Portolano S, McLachlan SM, Rapoport B. (1993) High affinity, thyroid-specific human autoantibodies displayed on the surface of filamentous phage use V genes similar to other autoantibodies. J. Immunol. 151:2839–51.
Dorner T, Foster SJ, Farner NL, Lipsky PE. (1998) Immunoglobulin kappa chain receptor editing in systemic lupus erythematosus. J. Clin. Invest. 102:688–94.
Foster SJ, Brezinschek HP, Brezinschek RI, Lipsky PE. (1997) Molecular mechanisms and selective influences that shape the kappa gene repertoire of IgM+ B cells. J. Clin. Invest. 99:1614–27.
Suzuki N, Harada T, Mihara S, Sakane T. (1996) Characterization of a germline Vk gene encoding cationic anti-DNA antibody and role of receptor editing for development of the autoantibody in patients with systemic lupus erythematosus. J. Clin. Invest. 98:1843–50.
Tiegs SL, Russell DM, Nemazee D. (1993) Receptor editing in self reactive bone marrow B cells. J. Exp. Med. 177:1009–20.
Casellas R, Shih TA, Kleinewietfeld M, et al. (2001) Contribution of receptor editing to the antibody repertoire. Science. 291:1541–4.
Chen C, Prak EL, Weigert M. (1997) Editing disease-associated autoantibodies. Immunity 6:97–105.
Juul L, Hougs L, Andersen V, Svejgaard A, Barington T. (1997) The normally expressed kappa immunoglobulin light chain gene repertoire and somatic mutations studied by single-sided specific polymerase chain reaction (PCR); frequent occurrence of features often assigned to autoimmunity. Clin. Exp. Immunol. 109:194–203.
Radic MZ, Erikson J, Litwin S, Weigert M. (1993) B lymphocytes may escape tolerance by revising their antigen receptors. J. Exp. Med. 177:1165–73.
Martin D, Huang RQ, LeBien T, Van Ness B. (1991) Induced rearrangement of kappa genes in the BLIN-1 human pre-B cell line correlates with germline J-C kappa and V kappa transcription. J. Exp. Med. 173:639–45.
Goebel P, Janney N, Valenzuela JR, Romanow WJ, Murre C, Feeney AJ. (2001) Localized gene-specific induction of accessibility to V(D)J recombination induced by E2A and early B cell factor in nonlymphoid cells. J. Exp. Med. 194: 645–56.
Zhang Z, Burrows PD, Cooper MD. (2004) The molecular basis and biological significance of VH replacement. Immunol. Rev. 197:231–42.
Chen C, Nagy Z, Prak EL, Weigert M. (1995) Immunoglobulin heavy chain gene replacement: a mechanism of receptor editing. Immunity 3:747–55.
Itoh K, Meffre E, Albesiano E, et al. (2000) Immunoglobulin heavy chain variable region gene replacement as a mechanism for receptor revision in rheumatoid arthritis synovial tissue B lymphocytes. J. Exp. Med. 192:1151–64.
Dorner T, Farner NL, Lipsky PE. (1999) Ig lambda and heavy chain gene usage in early untreated systemic lupus erythematosus suggests intensive B cell stimulation. J. Immunol. 163:1027–36.
Diaw L, Siwarski D, DuBois W, Jones G, Huppi K. (2000) Double producers of kappa and lambda define a subset of B cells in mouse plasmacytomas. Mol. Immunol. 37:775–81.
Meffre E, Davis E, Schiff C, et al. (2000) Circulating human B cells that express surrogate light chains and edited receptors. Nat. Immunol. 1:207–13.
Zhang Z, Wu X, Limbaugh BH, Bridges SL Jr. (2001) Expression of recombination-activating genes and terminal deoxynucleotidyl transferase and secondary rearrangement of immunoglobulin κ light chains in rheumatoid arthritis synovial tissue. Arthritis Rheum. 44:2275–84.
Kaschner S, Hansen A, Jacobi A, et al. (2001) Immunoglobulin Vλ light chain gene usage in patients with Sjogren’s syndrome. Arthritis Rheum. 44: 2620–32.
Meffre E, Schaefer A, Wardemann H, Wilson P, Davis E, Nussenzweig MC. (2004) Surrogate light chain expressing human peripheral B cells produce self-reactive antibodies. J. Exp. Med. 199:145–50.
Stamatopoulos K, Kosmas C, Stavroyianni N, Loukopoulos D. (1996) Evidence for immunoglobulin heavy chain variable region gene replacement in a patient with B cell chronic lymphocytic leukemia. Leukemia 10:1551–6.
Lenze D, Greiner A, Knorr C, Anagnostopoulos I, Stein H, Hummel M. (2003) Receptor revision of immunoglobulin heavy chain genes in human MALT lymphomas. Mol. Pathol. 56:249–55.
Goossens T, Brauninger A, Klein U, Kuppers R, Rajewsky K. (2001) Receptor revision plays no major role in shaping the receptor repertoire of human memory B cells after the onset of somatic hypermutation. Eur. J. Immunol. 31:3638–48.
Magari M, Sawatari T, Kawano Y, et al. (2002) Contribution of light chain rearrangement in peripheral B cells to the generation of high affinity antibodies. Eur. J. Immunol. 32:957–66.
Goossens T, Klein U, Kuppers R. (1998) Frequent occurrence of deletions and duplications during somatic hypermutation: implications for oncogene translocations and heavy chain disease. Proc. Natl. Acad. Sci. U S A 95:2463–8.
Klein U, Goossens T, Fischer M, Kanzler H, Braeuninger A, Rajewsky K, Kuppers R. (1998) Somatic hypermutation in normal and transformed human B cells. Immunol. Rev. 162:261–80.
Kuppers R, Goossens T, Klein U. (1999) The role of somatic hypermutation in the generation of deletions and duplications in human Ig V region genes and chromosomal translocations. Curr. Top. Microbiol. Immunol. 246:193–8.
Frischmeyer PA, Dietz HC. (1999) Nonsense-mediated mRNA decay in health and disease. Hum. Mol. Genet. 8:1893–900.
Iborra FJ, Escargueil AE, Kwek KY, Akoulitchev A, Cook PR. (2004) Molecular cross-talk between the transcription, translation, and nonsense-mediated decay machineries. J. Cell Sci. 117:899–906.
Wang J, Vock VM, Li S, Olivas OR, Wilkinson MF. (2002) A quality control pathway that down-regulates aberrant T-cell receptor (TCR) transcripts by a mechanism requiring UPF2 and translation. J. Biol. Chem. 277:18489–93.
Buhler M, Paillusson A, Muhlemann O. (2004) Efficient downregulation of immunoglobulin mu mRNA with premature translation-termination codons requires the 5′-half of the VDJ exon. Nucleic Acids Res. 32:3304–15.
Benito C, Gomis R, Fernandez-Alvarez J, Usac EF, Gallart T. (2003) Transcript expression of two Igλ rearrangements and RAG-1/RAG-2 in a mature human B cell producing IgMλ islet cell autoantibody. J. Clin. Immunol. 23:107–18.
Darlow JM, Farrell AM, Stott DI. (2004) Non-functional immunoglobulin G transcripts in a case of hyper-immunoglobulin M syndrome similar to type 4. Immunology 111:212–22.
Acknowledgements
We wish to thank Prof. Marie-Paule Lefranc and Dr. Veronique Giudicelli (Laboratoire d’Immunogenetique Moleculaire, LIGM, Université Montpellier II, UPR CNRS) for their support and generosity to share with us their insight on immunoglobulin gene analysis; Dr. Paolo Ghia, Department of Oncology, Università Vita Salute-San Raffaele, Milano, Italy, and Dr. Fred Davi, Laboratory of Hematology and University Paris 6, Hôpital Pitié-Salpètrière, Paris, France, for many stimulating talks and helpful comments; and Dr. Anton Langerak, Department of Immunology Erasmus MC, University Medical Center Rotterdam, The Netherlands, for critically reading the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Belessi, C., Stamatopoulos, K., Hadzidimitriou, A. et al. Analysis of Expressed and Non-Expressed IGK Locus Rearrangements in Chronic Lymphocytic Leukemia. Mol Med 11, 52–58 (2005). https://doi.org/10.2119/2005-00044.Belessi
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/2005-00044.Belessi