
Abstract
Aflatoxin B1 (AFB1) is a widely spread mycotoxin that poses a threat to the healthy to human and animals. The liver is the main target organ for AFB1-induced damage, primarily causing inflammatory injury and oxidative stress. When AFB1 enters the body, it can damage the intestinal barrier function, and its metabolites are transported to the liver. Therefore, the damage to the liver is closely associated with intestinal barrier impairment. Lactobacillus plays a crucial role in mitigating liver damage by improving the intestinal barrier function. In our previous report, we reported that Lactobacillus reduces liver damage caused by AFB1. However, it is still unclear how the intestinal barrier contributes to the protective effects of Lactobacillus against AFB1. To investigate the protective effects and intestinal barrier mechanisms of Lactobacillus intestinals /rhamnosus against AFB1-induced liver damage, we orally administered AFB1 and Lactobacillus intestinals/rhamnosus to male SD rats. Then the body weight, organ index, histopathological changes in the liver and gut, liver and kidney function indicators, intestinal mucosal barrier indicators, serum AFB1 content and inflammatory factors, liver oxidative stress index, and short-chain fatty acids content were analyzed. Our findings demonstrate that exposure to AFB1 resulted in changes in liver histopathology and biochemical functions, altered inflammatory response and oxidative stress, compromised the intestinal mucosal barrier, and induced the accumulation of inflammatory factor and inflammation in the liver. However, supplementation with Lactobacillus intestinals or Lactobacillus rhamnosus significantly prevented AFB1-induced liver injury, alleviated histopathological changes and hepatic injury by the maintenance of intestinal mucosal barrier integrity.
Similar content being viewed by others

Introduction
Aflatoxin B1 (AFB1) is widely recognized as one of the most toxic mycotoxins [1, 2], carrying significant risks to agriculture, animal husbandry, human health, and food safety [1, 3, 4]. Its high carcinogenic and capacity to induce liver damage through inflammation and oxidative stress make it a major concern. These cytokines stimulate various immune-related cells, leading to local immune responses and inflammatory changes in the liver. When exposed to AFB1, animals produces inflammatory factors such as TNF-α (tumor necrosis factor) and IL-6 (interleukin-6), with TNF-α predominantly released by macrophages [5]. These cytokines stimulate monocytes, Kupffer's cells, T cells, and other immune-related cells to cause local immune response and inflammatory changes in the liver [5]. AFB1 also triggers the production of reactive oxygen species and oxidative stress in the liver, further contributing to its toxicity [6]. Recent research suggests that oxidative stress plays a significant role in aflatoxin's genotoxicity. Regulating the expression of inflammatory factors and oxidative stress factors may help mitigate liver damage [7, 8]. Our previous study demonstrated that Lactobacillus can reduce AFB1-induced hepatocyte vacuolation changes in chicken models. However, the underlying mechanism remains unclear.
The liver and intestineare closely interconnected in terms of structure and function. Liver inflammation is closely associated with intestinal barrier damage [9]. Approximately 75% of the blood supply to the liver comes from the intestine through the portal vein system, carrying nutrients, bacteria, metabolites, and toxins that can affect liver function. Impaired liver function reduces blood flow in the gut-liver axis, resulting in decreased bile and lysozyme bacteria secretion and slowed intestinal peristalsis [10]. This disruption compromises the intestinal mucosal barrier, leading to increased intestinal permeability. Bacterial toxins from the intestinal cavity translocate to the liver via the portal vein system, further exacerbating liver inflammation [11]. Previous studies have highlighted intestinal barrier dysfunction as a key factor in persistent inflammatory liver injury [12].
Lactobacillus exhibits various biological effects, including anti-oxidation and anti-inflammatory properties [13,14,15,16]. It has been shown to significantly reduce liver inflammation reactions in experimental animal models exposed to AFB1, suggesting a potential role in preventing and controlling liver inflammatory changes [5, 17]. Our previous study demonstrated that Lactobacillus rhamnosus and Lactobacillus intestinalis can effectively inhibit the expression of serum pro-inflammatory cytokines TNF-α and IL-6 in AFB1-induced animal models, thereby reducing liver damage. Therefore, Lactobacillus rhamnosus and Lactobacillus intestinalis may serve as ideal products for preventing liver inflammation. However, due to the rapid metabolism and clearance rate of Lactobacillus in the body, achieving effective serum concentrations is challenging, making it difficult to directly attribute the improvement of liver function and regulation of inflammatory factors to Lactobacillus. Further research is needed to fully understand the mechanism by which Lactobacillus prevent liver inflammation.
This study aims to establish an animal model induced by AFB1 and intervene with Lactobacillus rhamnosus and Lactobacillus intestinalis to elucidate their role in improving AFB1-induced liver inflammatory changes. Additionally, we aim to explore the crucial role of the intestinal mucosal barrier in modulating the occurrence and progression of liver inflammation. The findings of this study will provide new preventative strategies for liver inflammation and injury during severe AFB1 exposure.
Results
Histopathological observations of the liver
The results of liver tissue pathological changes in each group are shown in Fig. 1. In the control group, the liver tissue structure was normal, the liver cell structure was full, the central vein was clearly visible, the hepatic cords were arranged radially along the central vein, and there was no obvious inflammatory cell infiltration in the tissue (Fig. 1a). No abnormal pathological changes were observed in the DMSO (Dimethyl sulfoxide) group (Fig. 1b). However, in the AFB1 group, the hepatocyte structure was loose, extensive water could be seen in the hepatocytes, and some were edematous with vacuolar degeneration, as shown by the yellow arrow in the figure. There was also a small amount of inflammatory cell infiltration in the tissue, as shown by the black arrow (Fig. 1c). Compared with the AFB1 group, the L1 (Lactobacillus rhamnosus) and L2 (Lactobacillus intestinalis) treatment groups showed a reduced extent of liver lesions caused by AFB1. It can be seen from Fig. 1d and e that the degree of liver cell edema was significantly reduced, with only a small amount of inflammatory cell infiltration, especially in the AFB1 + L1 group. These findings suggest that the addition of L1 and L2 can reduce the damage of AFB1 to rat liver.

Histopathological observation of livers after different treatments. a Control group. b Vehicle group (DMSO). c Rat treated with AFB1. d Rat treated with AFB1 and L1. e Rat treated with AFB1 and L2. Note: All of the above observed at 200 ×
Histopathological changes of jejunum in rats
The results of pathological changes in the jejunum for each group are shown in Fig. 2. The intestinal tissue structure of control group rats was normal, the intestinal villi were neatly arranged, the intestinal mucosa epithelial cells showed no obvious dropsy or edema, the goblet cells of intestinal villi were visible (yellow arrow), the intestinal crypts were visible (black arrow), and there was no significant inflammatory cell infiltration (Fig. 2a). No noticeable abnormalities were observed in the DMSO group slices (Fig. 2b). However, compared to the control group, the jejunum tissue structure of the AFB1 group exhibited abnormalities, such as disordered, atrophic and broken intestinal villi, edematous intestinal villi epithelial cells, and exfoliated epithelial cells (yellow arrow). The blood vessels in the submucosa were also hyperemic and dilated (red arrow), and there was obvious inflammatory cell infiltration (black arrow) (Fig. 2c). Compared to the AFB1 group, the overall structure of the intestinal tissue in the AFB1 + L1 and AFB1 + L2 groups appeared to be normal. The intestinal villi were arranged in order and only a few inflammatory cells infiltrated the tissues, as shown by the black arrow in Fig. 2d and e. These findings suggest that the addition of L1 and L2 could alleviate the injury caused by AFB1 to the jejunum.

Histopathological changes of jejunum in each group of rats. a Control group. b Vehicle group (DMSO). c Rat treated with AFB1. d Rat treated with AFB1 and L1. e Rat treated with AFB1 and L2. Note: All of the above observed at 200×
Effects of Lactobacillus intestinals and Lactobacillus rhamnosus on serum liver function indexes in rats with AFB1 poisoning
The serum liver function indices for each group are shown in Fig. 3. There was no significant difference between the Control (CON) and DMSO groups, indicating that the dosage of DMSO used in the experiment did not have any effect on the rats' livers. However, compared to the CON group, the AFB1 group had significantly increased contents of ALT (Alanine aminotransferase), AST (Aspartate aminotransferase), ALP (Alkaline phosphatase), LDH (Lactate dehydrogenase), CHE (Cholinesterase), TBIL (Total bilirubin), DBIL (Direct bilirubin), and IBIL (Indirect bilirubin) in serum while significantly decreasing the contents of Total protein (TP), Albumin (ALB), Globulin (GLB), and Prealbumin (PA) (P < 0.001). On the other hand, both the AFB1 + L1 and AFB1 + L2 groups showed a significant decrease in the contents of ALT, AST, LDH, CHE, TBIL, DBIL, and IBIL compared to the AFB1 group (P < 0.001). Additionally, the content of ALP in the AFB1 + L1 group was significantly lower than that in the AFB1 group (P < 0.01), while the content of ALP in the AFB1 + L2 group was significantly lower than that in the AFB1 group (P < 0.05). Furthermore, the contents of TP, ALB, and GLB in both the AFB1 + L1 and AFB1 + L2 groups were significantly higher than those in the AFB1 group (P < 0.001), and the content of PA in the AFB1 + L1 group was significantly higher than that in the AFB1 group (P < 0.05).

Changes of serum liver function indexes in rats (CON group, DMSO group, AFB1 group, AFB1 + L1 group, AFB1 + L2 group). The different level changes of liver function indices (A–L). Note: Each value represents mean ± SD. * and *** respectively indicate significant and extremely significant differences compared with the control group (P < 0.05; P < 0.001); #, ## and ### points indicate significant difference compared with the AFB1 group (P < 0.05; P < 0.01; P < 0.001)
The effect of Lactobacillus intestinals and Lactobacillus rhamnosus on serum inflammatory factors in AFB1 poisoned rats
The changes in serum inflammatory factors for each group are shown in Fig. 4. The difference between the control group and the DMSO group was not significant, indicating that the dosage of DMSO used in this experiment did not affect the changes in serum inflammatory factors in rats. Compared to the control group, the AFB1 group showed significantly increased content of TNF-α in serum (P < 0.001). However, the content of TNF-α in the AFB1 + L1 group was significantly higher (P < 0.001), while the content of TNF-α in the AFB1 + L2 group was significantly lower than that in the AFB1 group (P < 0.001). Additionally, the content of TNF-α in the AFB1 + L1 group was significantly lower than that in the AFB1 group (P < 0.05). Regarding IL-1β levels in serum, the AFB1 group exhibited a significantly higher level compared to the control group. However, the levels of IL-1β (interleukin-1β) in both the AFB1 + L1 and AFB1 + L2 groups were significantly higher (P < 0.05), while the level of IL-1β in the AFB1 + L1 group was slightly but not significantly lower than that in the AFB1 group (P > 0.05). Moreover, the level of IL-6 in serum was significantly higher in the AFB1 group (P < 0.01), whereas the level of IL-6 in the AFB1 + L1 group was significantly lower than that in the AFB1 group (P < 0.05), and the level in the AFB1 + L2 group was also lower than that in the AFB1 group although the difference was not significant (P > 0.05). In terms of IL-10 levels in serum, the AFB1 group exhibited a significantly lower level (P < 0.05), while the levels of IL-10 in both the AFB1 + L1 and AFB1 + L2 groups were significantly higher than that in the AFB1 group (P < 0.01). These findings suggested that L1 and L2 can reduce pro-inflammatory factors content and increase anti-inflammatory factor content in serum of AFB1-poisoned rats.

Changes of serum inflammatory factors in rats (CON group, DMSO group, AFB1 group, AFB1 + L1 group, AFB1 + L2 group). A TNF-α level. B IL-1β level. C IL-6 level. D IL-10 level. Note: Each value represents mean ± SD. *, ** and *** respectively indicate significant and extremely significant differences compared with the control group (P < 0.05; P < 0.01; P < 0.001); #, ## and ### points indicate significant difference compared with the AFB1 group (P < 0.05; P < 0.01; P < 0.001)
Effects of Lactobacillus intestinals and Lactobacillus rhamnosus on liver antioxidant status in rats with AFB1 poisoning
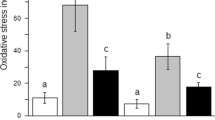
The changes in liver antioxidation for each group are shown in Fig. 5. There was no significant difference between the control group and the DMSO group, indicating that the dosage of DMSO used in this experiment did not affect the changes in liver oxidative stress indexes. Compared to the control group, the AFB1 group showed a significantly increased content in MDA (Malondialdehyde) in the liver tissue (P < 0.01), yet the MDA content in both the AFB1 + L1 and AFB1 + L2 groups was significantly lower than that in the AFB1 group (P < 0.05). Moreover, the content of SOD (Superoxide dismutase) in the liver of the AFB1 group was significantly decreased (P < 0.01); however, the content of SOD in both the AFB1 + L1 and AFB1 + L2 groups was higher than that in the AFB1 group, although the differences were not significant (P > 0.05). Regarding CAT (Catalase) levels in the liver, the AFB1 group yielded a significantly lower content compared to the control group (P < 0.01). However, the CAT content in the AFB1 + L1 group was significantly higher than that in the AFB1 group (P < 0.05), whereas the content in the AFB1 + L2 group was also higher than that in the AFB1 group, though the difference was not significant (P > 0.05). Furthermore, the GSH PX (Glutathione peroxidase) content in the liver tissue of the AFB1 group was significantly lower than that in the control group (P < 0.01). Nevertheless, the GSH PX content in the AFB1 + L1 group was significantly higher than that in the AFB1 group (P < 0.05), and the GSH content in the AFB1 + L2 group was higher than that in the AFB1 group, but the difference was not significant (P > 0.05). Lastly, the GR content in liver tissue of the AFB1 group was significantly lower compared to the control group (P < 0.001); however, the GR content in both the AFB1 + L1 and AFB1 + L2 groups were significantly higher than that in the AFB1 group (P < 0.001). These findings indicated that L1 and L2 can reduce lipid peroxidation in the liver of rats with AFB1 poisoning, with L1 exhibiting a better effect.

Changes in the antioxidant status of rat liver tissues (CON group, DMSO group, AFB1 group, AFB1 + L1 group, AFB1 + L2 group). A SOD level. B CAT level. C GSH level. D GR level. Note: Each value represents mean ± SD. ** and *** respectively indicate significant and extremely significant differences compared with the control group (P < 0.01; P < 0.001); #, ## and ### points indicate significant difference compared with the AFB1 group (P < 0.05; P < 0.01; P < 0.001)
Efects of Lactobacillus intestinals and Lactobacillus rhamnosus on intestinal barrier indexes in rats with AFB1 poisoning
The changes in intestinal mucosal barrier function indexes for each group are shown in Fig. 6. There was no significant difference between the control group and the DMSO group, indicating that the dosage of DMSO used in this experiment did not affect changes in intestinal mucosal barrier indexes in rats. Compared to the control group, the AFB1 group exhibited a significantly increased content of serum DAO (Diamine oxidase) (P < 0.01); however, the DAO content in the AFB1 + L1 group was significantly lower than that in the AFB1 group (P < 0.05), whereas the DAO content in the AFB1 + L2 group was lower than that in the AFB1 group, but the difference was not significant (P > 0.05). Furthermore, the content of d-lac in the serum of rats in the AFB1 group was significantly increased (P < 0.01), yet the content of D-Lac(D-lactic acid) in both the AFB1 + L1 and AFB1 + L2 groups was significantly lower than that in the AFB1 group (P < 0.05). Regarding IFABP (Intestinal fatty acid-binding protein) levels in serum, the AFB1 group yielded a significantly higher level compared to the control group (P < 0.001); however, the IFABP level in the AFB1 + L1 group was significantly lower than that in the AFB1 group (P < 0.001), and the level in the AFB1 + L2 group was also significantly lower than that in the AFB1 group (P < 0.05). These results suggest that L1 and L2 can reduce damage caused by AFB1 to the rat intestinal mucosa, with L1 having a better effect.

Changes of intestinal mucosal barrier function in rats (CON group, DMSO group, AFB1 group, AFB1 + L1 group, AFB1 + L2 group). A DAO level. B D-Lac level. C IFABP level. Note: Each value represents mean ± SD. ** and *** respectively indicate significant and extremely significant differences compared with the control group (P < 0.01; P < 0.001); # and ### points indicate significant difference compared with the AFB1 group (P < 0.05; P < 0.001)
Discussion
AFB1, the most toxic metabolite of aflatoxin B1 in the liver, is known to bind to DNA and RNA, interfere with protein synthesis, and cause liver damage [18]. In this study, hepatocytes of the AFB1-treated rats exhibited extensive edema, vacuolar degeneration, and mild infiltration of inflammatory cells between tissues. However, the degree of hepatocyte edema was significantly reduced in the groups treated with Lactobacillus rhamnoses and Lactobacillus intestinalis, accompanied by a minor amount of inflammatory cell infiltration. These findings suggest that both lactobacilli have inhibitory effects on AFB1-induced liver damage.
AFB1 induces oxidative stress by generating free radicals, which have been identified as a key factor in liver damage [19]. The imbalance between free radicals and antioxidant defense systems leads to a cascade of destructive effects [20]. In vitro studies have reported that AFB1 down-regulates antioxidant enzymes like SOD, GSH-Px, GR, and CAT, resulting in increased lipid peroxidation and decreased levels of exogenous antioxidants such as reduced glutathione (GSH) [21]. Several studies have also demonstrated decreased SOD activity in the liver of broilers fed AFB1-contaminated diets, as well as increased MDA concentration and decreased SOD concentration in AFB1-exposed dairy goats [22]. Moreover, in our study, rats exposed to a dosage of 50 μg/kg body weight of AFB1 exhibited liver and kidney damage, which was associated with AFB1-induced oxidative stress. This oxidative stress increased the production of MDA and NO, while reducing the activity of important antioxidative compounds such as GSH, GSH-Px, SOD and catalase (CAT) [23]. Comparing the contents of SOD, CAT, GSH, and GR in liver tissue of AFB1-treated rats to the control group, we observed a significant decrease. However, the treatment with both Lactobacillus rhamnoses and Lactobacillus intestinalis increased the contents of these important antioxidative compounds (SOD, CAT, GSH, GR) in liver tissue of AFB1-poisoned rats. These findings indicate that Lactobacillus rhamnoses and Lactobacillus intestinalis have the potential to reduce the oxidative stress response in the liver of AFB1-induced rats, ultimately mitigating the damage caused by AFB1.
AFB1 can induce inflammation in various organs and cells, triggering defensive response to damage factors. This leads to excessive expression of chemokines and inflammatory cytokines, such as TNF-α, IL-6, and IL-8 [24]. NTNF-α is a pro-inflammatory cytokine that directly damages the integrity and function of the intestinal mucosa by stimulating the production of chemokines. Conversely, IL-10 is an anti-inflammatory cytokine that reduces the inflammatory response of the intestinal mucosa and enhance local immunity. AFB1, at doses of 10–40 μg/kg, has been shown to increased the release of TNF-α while reducing the release of IL-10 [25]. Consistent with previous studies, our research found a significant increase in the levels of TNF-α, IL-1β, and IL-6, and a decrease in IL-10 levels in the serum of rats exposed to AFB1 compared to the control group. However, treatment with Lactobacillus rhamnoses or Lactobacillus intestinalis reduced TNF-α and IL-6 levels and increased IL-10 content. Although Lactobacillus rhamnoses showed a decreasing trend in IL-1β content, the difference was not significant. Similarly, while Lactobacillus intestinalis reduced IL-1β content, the difference was not significant. In summary, these results indicate that Lactobacillus rhamnoses and Lactobacillus intestinalis have inhibitory effects on kidney damage and inflammation in rats exposed to AFB1, possibly by reducing the expression of pro-inflammatory cytokines and increasing the expression of anti-inflammatory cytokines.
The gastrointestinal tract is responsible for nutrient digestion, absorption, and defense against pathogen infections [26]. Due to the area of the intestine and the residence time of food, it is more susceptible to AFB1 toxicity compared to other organs. The integrity of the intestinal barrier is crucial for nutrient absorption and inhibiting pathogen invasion. Long-term exposure to low levels of AFB1 can hinder the growth performance of pigs, reduce their digestibility, damage the integrity of the intestinal barrier, decrease intestinal antioxidant capacity, and increase pro-inflammatory cytokine production. Recent studies have shown that AFB1 can significantly disrupt intestinal barrier function [27]. Diamine oxidase (DAO) is an important intracellular enzyme located inthe cytoplasm of small intestinal cells, with high activity in animal intestinal mucosa, liver, kidney, and other tissues. Its activity reflects the ability of intestinal cells to synthesize protein and maintain cell activity. Healthy organisms show very low DAO activity in the blood, which mainly originates from intestinal cells. However, when the intestinal mucosal layer cells are damaged and necrotic, a large amount of DAO enters the blood circulation, increasing DAO activity in the blood. Therefore, DAO can be used to reflect the activity and integrity of intestinal epithelial cells and evaluate intestinal permeability [28]. Previous studies have shown that intestinal ischemia–reperfusion caused damage to intestinal cells and barrier function, consistent with changes in DAO content in rats' blood [29]. In our study, serum levels of DAO, D-Lac, and IFABP in rats exposed to AFB1 were significantly increased compared to the CON group, indicating that AFB1 damaged the intestinal barrier function in rats. Treatment with Lactobacillus rhamnoses significantly reduced serum levels of DAO, D-Lac, and IFABP in rats exposed to AFB1. Although Lactobacillus intestinalis treatment reduced DAO content, it was not significant, but significantly increased D-Lac and IFABP levels. These results suggest that Lactobacillus rhamnoses and Lactobacillus intestinalis can reduce serum levels of DAO, D-Lac, and IFABP to various degrees, thereby inhibiting intestinal barrier damage caused by AFB1 in rats. The effects of Lactobacillus rhamnoses were more significant than those of Lactobacillus intestinalis.
Several studies have demonstrated that mycotoxins can severely damage the small intestine of chickens, leading to villous epithelial cell atrophy, degeneration, necrosis, submucosal hemorrhage, and infiltration of inflammatory cells. This can cause the proliferation of intrinsic layers and goblet cells in the intestine, resulting in decreased villous height, deepened crypt depth, and reduced villous/crypt ratio in the duodenum, jejunum, and ileum [30]. This study found that Lactobacillus rhamnoses and Lactobacillus intestinalis can mitigate these effects by reducing villous atrophy and rupture caused by AFB1, preventing villous epithelial cell loosening, edema, and shedding, and increasing villous height and villous/crypt ratio. This leads to an improvement in the overall structure of the intestinal tissue, effectively returning it to a normal state (Supplementary materials).
Lactobacillus has been shown to play an important role in maintaining intestinal health by increasing the population of beneficial microorganisms in the intestine. As a functional food with increasing demand, Lactobacillus' ability to bind and remove aflatoxins is also very important as a dietary method for preventing adverse health effects caused by AFB1 exposure [31]. AFB1 is readily absorbed into the blood through the gastrointestinal tract. Some studies have found that Lactobacillus can significantly reduce AFB1 levels [32,33,34]. In our study, the effectiveness of Lactobacillus in reducing AFB1 levels was investigated by measuring serum AFB1 levels in all groups. No AFB1 was detected in the serum of the control group rats. The AFB1 levels in the serum of the Lactobacillus intestinalis and Lactobacillus rhamnoses groups were significantly reduced (Supplementary materials). These results suggest that Lactobacillus intestinalis and Lactobacillus rhamnoses can bind with AFB1 in the intestine, reducing its absorption. This reduces the availability of free AFB1 and subsequently its to the liver for metabolic process. Overall, these findings suggest that Lactobacillus may be a useful dietary supplement for reducing AFB1 levels in the body and mitigating some of the adverse health effects associated with AFB1 exposure.
Conclusions
This study demonstrated that oral administration of Lactobacillus intestinalis and Lactobacillus rhamnosus can reduce the occurrence of liver and kidney injury induced by AFB1 in rats. The protective mechanism is believed to be related to the maintenance of mucosal barriers in the gut. AFB1 can disrupt the integrity of the intestinal barrier and increase the permeability, allowing toxins and harmful substances to enter the bloodstream and affect various organs, including the liver and kidneys. Lactobacillus have been shown to enhance the integrity of the intestinal mucosa, improve the barrier function, and reduce the absorption of AFB1 into the bloodstream. This helps to prevent or mitigate the liver and kidney injury caused by AFB1. The study's findings suggest that Lactobacillus intestinalis and Lactobacillus rhamnosus could be potential dietary supplement for protecting against AFB1-induced liver and kidney damage.
Materials and methods
Main reagents
AFB1 storage solution
Dissolve AFB1 powder in DMSO to prepare a stock solution of AFB1 with a concentration of 1 mg/mL. When using, dilute the stock solution with distilled water to the desired concentration.
Strain cultivation
Lactobacillus rhamnosus was revived according to the instructions, and inoculated into MRS liquid medium, then incubated at 36 ℃ for 16 h in a constant temperature incubator. Lactobacillus intestinalis was stored at −80 ℃ and activated by inoculation into MRS liquid medium and incubation at 36 ℃ for 16 h. The prepared bacterial liquid was centrifuged (4 ℃, 2000 × g, 10 min) in a high-speed freezer, washed twice with phosphate-buffered saline (PBS, pH 7.4), then resuspended in PBS and adjusted to a live bacteria concentration of 1 × 109 CFU/mL using a UV spectrophotometer.
Animal ethics
All animals procedures were conducted in accordance with the Guidelines for the Care and Use of Experimental Animals at Heilongjiang Bayi Agricultural University (Daqing, China) (DWKJXY2023063).
Animals
Fifty male SD rats were purchased from the Experimental Animal Center of Harbin Medical University and used for animal testing in the animal house of the College of Animal Science and Technology at Heilongjiang Bayi Agricultural University.
Experimental animal grouping and management
After a 7-day period of adaptive feeding in the animal room, rats were randomly assigned to five groups of ten each and subjected to different treatments: group I served as a blank control (CON); group II was treated with DMSO solvent as a control (DMSO) by gavage at a dose equivalent to AFB1 treatment, approximately 250 μg/kg body weight; group III was exposed to AFB1 toxin (AFB1) at a dose of 250 μg/kg body weight by gavage; group IV was treated with Lactobacillus rhamnosus (AFB1 + L1), each rat receiving 250 μg/kg body weight of AFB1 and 1 × 109 CFU of Lactobacillus rhamnosus by gavage, and group V was treated with Lactobacillus intestinals (AFB1 + L2), each rat receiving 250 μg/kg body weight of AFB1 and 1 × 109 CFU of Lactobacillus intestinals by gavage. AFB1 and Lactobacillus were administered by gavage at 12-h intervals once a day. The experiment lasted for 8 weeks. During the experiment, all groups of rats were fed the same regular diet and had free access to drinking water under a 12-h light/dark cycle.
Sample collection
After the trial, all rats were fasted for 12 h. The rats were restrained with a glass beaker, and a cotton ball soaked in ether was placed in the beaker and keep the room maintain ventilation. Blood samples were collected via abdominal aortic puncture and transferred into 10 ml centrifuge tubes. The tubes were then centrifuged at 1000 g for 10 min at 4 ℃, and the supernatant was collected and stored at −80 ℃. Immediately after blood collection, the rats were euthanized by neck dislocation, and the organs including the heart, liver, spleen, lungs, and kidneys were collected and washed with normal saline to remove blood. The organs were blotted dry with filter paper and weighed on an electronic balance. Portions of the liver, duodenum, jejunum, ileum, cecum, colon, and rectum were selected and fixed with 4% paraformaldehyde. Some liver tissue was snap-frozen in liquid nitrogen and stored at −80 ℃ for subsequent analysis.
Histological section observation
The collected rat liver and intestinal tissues should be fixed in 4% paraformaldehyde solution for at least 24 h. After that, the fixed tissue blocks should be embedded in paraffin to make 5 μm sections, which can then be stained with hematoxylin–eosin or other staining methods. Finally, observe the histopathological changes of tissues using an optical microscope and capture images for analysis purposes.
Serum liver function index determination
The serum liver function indexes are assessed by using a fully automated biochemical analyzer to detect the levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), lactate dehydrogenase (LDH), cholinesterase (CHE), total protein (TP), albumin (ALB), globulin (GLB), prealbumin (PA), total bilirubin (TBIL), direct bilirubin (DBIL) and indirect bilirubin (IBIL) in rat serum samples.
Measurement of pro-inflammatory cytokines in serum
Using enzyme-linked immunosorbent assay (ELISA) according to the instructions of Nanjing Jiancheng ELISA kit, detect the expression levels of IL-1β, TNF-α, IL-6 and IL-10 in serum.
Measurement of liver tissue lipid peroxidation product MDA
The liver tissue sample taken out from −80 ℃ freezer was cut into small pieces with scissors, homogenized in phosphate-buffered saline (PBS, pH 7.4), and centrifuged at 1700 rpm for 10 min at 4 ℃. The supernatant was collected and the malondialdehyde (MDA) content was determined according to the instructions of Nanjing Jiancheng ELISA kit.
Measurement of liver tissue antioxidant indices
After homogenization of the liver sample, the supernatant was further diluted with phosphate-buffered saline (PBS, pH 7.4) and the total protein level was determined using BCA protein assay kit (China Beyotime). The levels of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px), and glutathione reductase (GR) were then determined according to the instructions of Nanjing Jiancheng ELISA kit.
Detection of intestinal mucosal barrier function indices in rat serum
Detection of intestinal mucosal barrier function indicators in rat serum. According to the instructions of Nanjing Jiancheng assay kit, detect the levels of diamine oxidase (DAO), D-lactic acid (D-Lac) and intestinal fatty acid-binding protein (ILABP) in serum.
Measurement of AFB1 levels in serum
Measurement of AFB1 content in rat serum. The concentration of AFB1 in rat serum was measured using a commercially available ELISA kit, COKAQ8000 (provided by Romer Labs, Beijing, China).
Data analysis and statistics
All mentioned experiments were performed in triplicate to ensure accuracy and reproducibility. Excel was used for preliminary statistical analysis of experimental data, and SPSS 17.0 software was used to process the data. One-way ANOVA was used to determine the significance between groups, with P < 0.05 indicating statistical significance. The results were expressed as "mean ± standard deviation". GraphPad Prism 7.0 software was used to create bar graphs and line graphs to visually display differences and trends in the experimental data. In addition, more in-depth data analysis and interpretation can be performed using statistical methods such as multi-factor ANOVA, t-test, and correlation analysis with SPSS 17.0 software. Finally, scientific conclusions and summaries were drawn based on the experimental results.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AFB1:
-
Aflatoxin B1
- DMSO:
-
Dimethyl sulfoxide
- AFB1 + L1:
-
Lactobacillus rhamnosus
- AFB1 + L2:
-
Lactobacillus intestinals
- CON:
-
Control
- ALT:
-
Alanine aminotransferase
- AST:
-
Aspartate aminotransferase
- ALP:
-
Alkaline phosphatase
- LDH:
-
Lactate dehydrogenase
- CHE:
-
Cholinesterase
- TP:
-
Total protein
- ALB:
-
Albumin
- GLB:
-
Globulin
- PA:
-
Prealbumin
- TBIL:
-
Total bilirubin
- DBIL:
-
Direct bilirubin
- MDA:
-
Malondialdehyde
- SOD:
-
Superoxide dismutase
- CAT:
-
Catalase
- GSH-Px:
-
Glutathione peroxidase
- GR:
-
Glutathione reductase
- DAO:
-
Diamine oxidase
- D-Lac:
-
D-lactic acid
- ILABP:
-
Intestinal fatty acid-binding protein
References
Williams JH, Phillips TD, Jolly PE, Stiles JK, Jolly CM, Aggarwal D. Human aflflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. Am J Clin Nutr. 2004;80:1106–22.
Bandyopadhyay R, Ortega-Beltran A, Akande A, Mutegi C, Atehnkeng J, Kaptoge L, et al. Biological control of aflflatoxins in Africa: current status and potential challenges in the face of climate changes. World Mycotoxin J. 2016;9:771–89.
Benkerroum N. Chronic and acute toxicities of aflatoxins: mechanisms of action. Int J Environ Res Public Health. 2020;17(2):423–51.
Ajmal M, Bedale W, Akram A, Yu J-H. Comprehensive review of aflatoxin contamination, impact on health and food security, and management strategies in Pakistan. Toxins. 2022;14:845–62.
Chen Y, Li R, Chang Q, Dong Z, Yang H, Xu C. Lactobacillus bulgaricus or Lactobacillus rhamnosus suppresses NF-κB signaling pathway and protects against AFB1-induced hepatitis: a novel potential preventive strategy for aflatoxicosis? Toxins. 2019;11(1):17–40.
Cao W, Yu P, Yang K, Cao D. Aflatoxin B1: metabolism, toxicology, and its involvement in oxidative stress and cancer development. Toxicol Mech Methods. 2022;32(6):395–419.
Guan X, Zhao Y, Liu X, Shang B, Xing F, Zhou L, et al. The bZIP transcription factor Afap1 mediates the oxidative stress response and aflatoxin biosynthesis in Aspergillus flavus. Rev Argent Microbiol. 2019;51(4):292–301.
Wan XL, Li N, Chen YJ, Chen XS, Yang Z, Xu L, et al. Protective effects of lycopene on mitochondrial oxidative injury and dysfunction in the liver of aflatoxin B1-exposed broilers. Poult Sci. 2021;100(11):101441.
Madnawat H, Welu AL, Gilbert EJ, Taylor DB, Jain S, Manithody C, et al. Mechanisms of parenteral nutrition-associated liver and gut injury. Nutr Clin Pract. 2020;35(1):63–71.
Albillos A, de Gottardi A, Rescigno M. The gut-liver axis in liver disease: pathophysiological basis for therapy. J Hepatol. 2020;72(3):558–77.
Mandato C, Delli Bovi AP, Vajro P. The gut-liver axis as a target of liver disease management. Hepatobiliary Surg Nutr. 2021;10(1):100–2.
Fan SX, Wang J, Li Q, Li YS, Guan WX, Li JS. Mechanism of gut-microbiota-liver axis in the pathogenesis of intestinal failure-associated liver disease. Zhonghua Wei Chang Wai Ke Za Zhi. 2021;24(1):94–100.
Wang L, Zhao Z, Zhao L, Zhao Y, Yang G, Wang C, et al. Lactobacillus plantarum DP189 reduces α-SYN aggravation in MPTP-induced Parkinson’s Disease mice via regulating oxidative damage, inflammation, and gut microbiota disorder. J Agric Food Chem. 2022;70(4):1163–73.
Dias AMM, Douhard R, Hermetet F, Regimbeau M, Lopez TE, Gonzalez D, et al. Lactobacillus stress protein GroEL prevents colonic inflammation. J Gastroenterol. 2021;56(5):442–55.
Rastogi S, Singh A. Gut microbiome and human health: exploring how the probiotic genus Lactobacillus modulate immune responses. Front Pharmacol. 2022;13:1042189.
Iyer N, Williams MA, O’Callaghan AA, Dempsey E, Cabrera-Rubio R, Raverdeau M, et al. Lactobacillus salivarius UCC118™ dampens inflammation and promotes microbiota recovery to provide therapeutic benefit in a DSS-induced colitis model. Microorganisms. 2022;10(7):1383.
Huang L, Zhao Z, Duan C, Wang C, Zhao Y, Yang G, et al. Lactobacillus plantarum C88 protects against aflatoxin B1-induced liver injury in mice via inhibition of NF-κB-mediated inflammatory responses and excessive apoptosis. BMC Microbiol. 2019;19(1):170.
Rotimi OA, De Campos OC, Adelani IB, Olawole TD, Rotimi SO. Early-life AFB1 exposure: DNA methylation and hormone alterations. Vitam Horm. 2023;122:237–52.
Bueno DJ, Casale CH, Pizzolitto RP, Salvano MA, Oliver G. Physical adsorption of aflatoxin B1 by lactic acid bacteria and Saccharomyces cerevisiae: a theoretical model. J Food Protect. 2007;70(9):2148–54.
Chen SW, Hsu JT, Chou YA, Wang HT. The application of digestive tract lactic acid bacteria with high esterase activity for zearalenone detoxification. J Sci Food Agric. 2018;98(10):3870–9.
Sadeghi N, Bahadori R, Ojagh SM, Salamroodi E. Effect of dietary Lactobacillus rhamnosus on blood biochemical indices and some digestive enzymes activity in rainbow trout (Oncorhynchus mykiss) fed with aflatoxin B1 infected diet. J Anim Environ. 2020;12(2):151–60.
Karlovsky P, Suman M, Berthiller F, De Meester J, Eisenbrand G, Perrin I, et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016;32(4):179–205.
Kabak B, Dobson ADW, Var I. Strategies to prevent mycotoxin contamination of food and animal feed: a review. Crit Rev Food Sci Nutr. 2006;46(8):593–619.
Pál L, Dublecz K, Weber M, Balogh K, Erdélyi M, Szigeti G, et al. Effect of combined treatment with aflatoxin B1 and T-2 toxin and metabolites on some production traits and lipid peroxide status parameters of broiler chickens. Acta Vet Hung. 2009;57(1):75–84.
Shen HM, Shi CY, Shen YI, Ong CN. Detection of elevated reactive oxygen species level in cultured rat hepatocytes treated with aflatoxin B1. Free Radic Biol Med. 1996;21(2):139–46.
Levrat MA, Rémésy C, Demigné C. High propionic acid fermentations and mineral accumulation in the cecum of rats adapted to different levels of inulin. J Nutr. 1991;121(11):1730–7.
Topping DL, Illman RJ, Clarke JM, Trimble RP, Jackson KA, Marsono Y. Dietary fat and fiber alter large bowel and portal venous volatile fatty acids and plasma cholesterol but not biliary steroids in pigs. J Nutr. 1993;123(1):133–43.
Ratajczak W, Rył A, Mizerski A, Walczakiewicz K, Sipak O, Laszczyńska M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim Polonica. 2019;66(1):1–12.
Kumagai S. Intestinal absorption and excretion of aflatoxin in rats. Toxicol Appl Pharmacol. 1989;97(1):88–97.
Bonnefont-Rousselot D, Bastard JP, Jaudon MC, et al. Consequences of the diabetic status on the oxidant/antioxidant balance. Diabetes Metab. 2000;26(3):163–77.
Liew WP, Nurul-Adilah Z, Than LTL, Mohd-Redzwan S. The Binding Efficiency and Interaction of Lactobacillus casei Shirota Toward Aflatoxin B1. Front Microbiol. 2018;10(9):1503.
Zhang LY, Liu S, Zhao XJ, Wang N, Jiang X, Xin HS, et al. Lactobacillus rhamnosus GG modulates gastrointestinal absorption, excretion patterns, and toxicity in Holstein calves fed a single dose of aflatoxin B1. J Dairy Sci. 2019;102(2):1330–40.
Śliżewska K, Cukrowska B, Smulikowska S, Cielecka-Kuszyk J. The effect of probiotic supplementation on performance and the histopathological changes in liver and kidneys in broiler chickens fed diets with aflatoxin B1. Toxins (Basel). 2019;11(2):112.
Chang J, Wang T, Wang P, Yin Q, Liu C, Zhu Q, et al. Compound probiotics alleviating aflatoxin B1 and zearalenone toxic effects on broiler production performance and gut microbiota. Ecotoxicol Environ Saf. 2020;194:110420.
Acknowledgements
We are grateful to all the personnel at College of Animal Science and Veterinary Medicine in Heilongjiang Bayi Agricultural University involved in the care of the rats and in the collection samples.
Funding
This research were funded by National Natural Science Foundation of China (Beijing, China; grant no. 32125038 and U20A2062).
Author information
Authors and Affiliations
Contributions
C.Y.Y. participated to the study design and prepared the final manuscript, A.T.H. and L.H.L. prepared the draft, Z.H.T. and W.Q. carried out the experiment and conducted the data analysis, J.L. made critical revision the paper, C.J.X., W.J.Q., S.J. made grammer revision. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animals procedures were conducted in accordance with the Guidelines for the Care and Use of Experimental Animals at Heilongjiang Bayi Agricultural University (Daqing, China) (DWKJXY2023063).
Consent for publication
I would like to declare on behalf of my co-authors that the work described was original research that has not been published previously, and not under consideration for publication elsewhere, in whole or in part. All the authors listed have approved the manuscript.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, Y., Liu, H., An, T. et al. Lactobacillus intestinalis/Lactobacillus rhamnosus protects against AFB1-induced liver damage: involvement of intestinal mucosal barrier. One Health Adv. 1, 24 (2023). https://doi.org/10.1186/s44280-023-00023-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/s44280-023-00023-x