
Abstract
CRISPR-Cas systems are an immune defense mechanism that is widespread in archaea and bacteria against invasive phages or foreign genetic elements. In the last decade, CRISPR-Cas systems have been a leading gene-editing tool for agriculture (plant engineering), biotechnology, and human health (e.g., diagnosis and treatment of cancers and genetic diseases), benefitted from unprecedented discoveries of basic bacterial research. However, the functional complexity of CRISPR systems is far beyond the original scope of immune defense. CRISPR-Cas systems are implicated in influencing the expression of physiology and virulence genes and subsequently altering the formation of bacterial biofilm, drug resistance, invasive potency as well as bacterial own physiological characteristics. Moreover, increasing evidence supports that bacterial CRISPR-Cas systems might intriguingly influence mammalian immune responses through targeting endogenous genes, especially those relating to virulence; however, unfortunately, their underlying mechanisms are largely unclear. Nevertheless, the interaction between bacterial CRISPR-Cas systems and eukaryotic cells is complex with numerous mysteries that necessitate further investigation efforts. Here, we summarize the non-canonical functions of CRISPR-Cas that potentially impact bacterial physiology, pathogenicity, antimicrobial resistance, and thereby altering the courses of mammalian immune responses.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
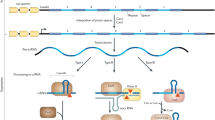
Clustered regularly interspaced short palindromic repeats (CRISPR) and their associated (Cas) proteins possess a unique structural and functional entity in bacterial genomes [1]. Thus far, CRISPR-Cas systems are the sole recognized acquired immunity against invading genetic elements found in many bacterial species and in most archaea. According to the CRISPR database (http://crispr.i2bc.paris-saclay.fr), CRISPR arrays are found in 202 (87%) out of 232 analyzed archaeal species and 3059 (45%) out of 6782 bacterial species by genome sequencing studies [2, 3]. The CRISPR-Cas systems consist of four parts: CRISPR regions, leader sequences, Cas proteins and tracrRNA (for Cas9) [4, 5]. The action steps of CRISPR-Cas systems are generally described as following three stages: adaptation, crRNA expression and interference (Fig. 1). The cas genes locating near the CRISPR locus encode CRISPR-related proteins to exert adaptive immune defense against bacteriophages or other genetic elements [6].

Overview of the CRISPR-Cas systems. CRISPR-Cas systems including leader, spacer and repeats are alternately arranged to form R-S structure and cas locus. The function of CRISPR-Cas systems include three stages: spacer acquisition, crRNA processing and assembly, and target degradation. The first step occurs after the foreign DNA sequence invades bacteria, and the bacterial genome intercepts a sequence from the invaded DNA fragment and integrates it into its CRISPR sequence to become a new spacer. Then, CRISPR clusters are transcribed into pre-crRNA under the initiation of the leader and further processed into mature small crRNAs. The mature crRNAs and Cas proteins assemble to form a CRISPR ribonucleoprotein (crRNP) complex. Once the same foreign DNA invades, the Cas protein complex binds, cleaves and degrades it by the base-pair principle
According to the diversity of cas genes and the mechanism of action for Cas proteins, CRISPR-Cas systems are categorized into 2 classes and further classified into six different types: type I to type VI [7, 8]. Basic CRISPR research only constitutes a relatively small portion, and the majority of work rather focuses on the application of CRISPR-Cas systems in genome-editing, transcriptional control, biotechnology, and agriculture engineering [9,10,11,12,13,14,15]. Furthermore, how CRISPR-Cas systems interact with other genes in the prokaryote, such as endogenous gene targeting and virulence gene regulation, remains largely un-elucidated [16,17,18,19,20].
Apart from providing immunity, CRISPR-Cas systems have been shown to influence the expression of bacterial genes that potentially impact bacterial biofilm formation, quorum sensing, antibiotic resistance, virulence (Fig. 2), adaptability, and viability of bacteria to environmental changes [21,22,23,24,25]. In addition to influencing bacterial behaviors, CRISPR-Cas systems might also have a role in influencing the immune systems of eukaryotic cells, helping bacteria evade host defense mechanisms. In this article, we will discuss some latest advances in the effects of CRISPR-Cas systems on the physiological and virulence alteration of bacteria and potential mechanisms as well as the impact of CRISPR-Cas-mediated gene regulation on mammalian host response, more detailed coverage of other relevant CRISPR-Cas researches may be found elsewhere [26,27,28].

Effect of CRISPR-Cas systems on physiological traits of bacteria. CRISPR-Cas systems affecting multiple bacterial characteristics, such as virulence, bacterial biofilm, QS and antibiotics resistance. CRISPR-Cas systems are associated with the expression of multiple virulence factors, such as vicR, gtfC, smu0630, comDE (Streptococcus sanguinis), esp, hyl, gelE, asa1, ace(E. faecalis). cas3 and cas1 genes directly or indirectly participate in and affect the formation of bacterial biofilm. There is mutual regulation between CRISPR-Cas systems and QS systems, and many genes are involved in this process, such as LasR, SmaI, SmaR cdpR, SsoPox and SmaR. CRISPR-Cas systems regulate the transfer of drug-resistant plasmids
CRISPR-Cas systems regulate bacterial biofilm formation
Biofilms and their function
Biofilms are structural surface materials of microbial cells encased in an extracellular matrix that may substantially decrease the susceptibility to antimicrobial agents, hence the potency for forming biofilms is critical for virulence and drug resistance [29]. Bacteria secrete a large amount of extracellular polymeric substances (EPS) to form biofilm, which can hinder the penetration of antibiotics and produce drug resistance. In addition, the virulence of bacteria in high density increases sharply under the regulation of the sensing system disrupts host immune systems, and gains tolerance to antibiotic therapy, ultimately leading to a variety of severe or persistent infections at the mammalian mucosal surface (e.g., respiratory or gut systems of mice and humans) [30, 31].
CRISPR-Cas systems alter biofilm levels
Recently, growing studies have shown that CRISPR-Cas systems can also regulate the formation of biofilm. CRISPR systems affect the formation of bacterial biofilm, thus affecting the transmission, toxicity and control of pathogenic bacteria. For example, Enterococcus faecalis carrying CRISPR-Cas systems possesses more prominent biofilm forming ability, better resists the destruction of bacteria by external factors, and effectively improves the colonization efficiency of bacteria in the host [32]. The resistance of E. coli to phages in Stx2 phage lysogen is significantly enhanced at a high bacterial density, while the biofilm-forming ability of E. coli is markedly decreased due to the activated CRISPR-Cas system mediated by mutant H-NS (heat-stable nucleoid structuring) protein which exist in a variety of Gram-negative bacterial species and inhibit gene transcription [33].
Another study showed that E. faecalis SK460 strains with strong biofilm forming ability in E. faecalis lack a Cas gene and functional CRISPR system. A possible reason is that the absence of CRISPR-Cas systems promotes the acquisition of antibiotic resistance gene, pheromone response plasmid, prophage, and pathogenic island.
Likewise, clinical studies suggest the same results: a study showed that the E. faecalis root canal isolates without CRISPR-Cas systems exhibited stronger biofilm formation and significant periapical lesions and strains lacking CRISPR1 or CRISPR3 loci produced increased biofilms compared to the strains containing CRISPR1 or CRISPR3 loci [25, 34]. This study suggests that there is an interaction between CRISPR-Cas systems and genes that govern bacterial biofilm formation.
Mechanisms for CRISPR-Cas-mediated biofilm regulation
After phage DMS3 infection, Pseudomonas aeruginosa as one of the lysogenic bacteria could not form biofilm properly. A further study demonstrated that when the bacteria have a complete CRISPR system and the spacer sequence of CRISPR structure cannot effectively combat DMS3-42, a phage that can inhibit the biofilm formation in lysogenesis condition. These results suggest that CRISPR-Cas systems may regulate gene expression through partial base-pairing mechanisms during biofilm formation [35]. This base-pair principle is the key to initiating the activation of CRISPR-Cas activity that may either result in anti-phage immunity or self-targeting that may regulate internal gene expression but potentially cause side effects [36,37,38].
Importantly, the existence of the protospacer-adjacent motif (PAM) in the P. aeruginosa genome is essential for the CRISPR-dependent loss of biofilm features, suggesting that endogenous targeting may share the same mechanism as anti-phage immunity defense. Wild-type (WT) P. aeruginosa PA14 lost the ability of forming biofilm after infected by lysogenic phage DSM3. Interfering with CRISPR2 (CRISPR array 2) or eliminating five of the six cas genes in P. aeruginosa PA14 Type I-F can restore the formation of biofilm. The study strongly suggests that the loss of biofilm is closely related to CRISPR and cas genes [39]. Also, a CRISPR-specific insertion sequence is needed to inhibit biofilm formation of P. aeruginosa as transcription of antisense RNA and silencing of the relevant genes is implicated in altered biofilm formation [39, 40]. Experiments showed that CRISPR-Cas systems in the bacterium were required for inhibiting the formation of biofilm and the ability of bacterial aggregation [39]. In addition, the study found that CRISPR2 instead of CRISPR1 plays a pivotal role in restoring biofilm formation to DMS3 lysogens [40]. These studies support the view that CRISPR-Cas may function in either inhibiting or promoting the formation of biofilms in different bacteria and depending on the conditions. Nevertheless, the detailed mechanisms of how CRISPR-Cas systems affect biofilm formation is currently unclear and needs to be further explored.
To further test this hypothesis and define the mechanisms of CRISPR-Cas in regulating biofilms, the impact of CRISPR-Cas is evaluated at the single Cas level. For instance, a cas3 gene deletion mutant of Streptococcus mutans UA159 showed impaired biofilm formation and weakened competition against (co-cultured) Streptococcus sanguinis in the presence of fluoride by up-regulating virulence genes (vicR, gtfC, smu0630 and comDE) [41]. This study illustrates that cas3 of S. mutans may be a positive modulator of biofilm formation and fluoride resistance. Other cas genes, such as cas1 encoded integrase, participate in the acquisition of a new DNA sequence from invading phages and indirectly affecting the formation and aggregation of bacterial biofilm [39]. Alternatively, self-targeting CRISPR arrays in the genome of Streptococcus mutans (a common oral pathogenic bacterium) that can target bacterial virulence genes gtfB or gtfBgtfC may lead to the decrease of EPS and disrupt the biofilm formation [42].
Summary for CRISPR-Cas in biofilm formation
The above studies together indicate that different components of CRISPR-Cas systems could impact the formation of bacterial biofilms by influencing the expression of distinct genes. CRISPR-Cas-mediated regulation of biofilm formation, aggregation, and motility may be critical mechanisms to limit the transmission of phages among bacteria. In other words, phage-infected bacteria may help separate themselves from biofilm and other colonization sites, which may play an important role in preventing large-scale bacterial infection, a potential self-limiting mechanism to help maintain their stable equilibrium [43, 44].
CRISPR-Cas systems interact with quorum sensing systems
Quorum sensing (QS) systems
QS systems are a powerful communication mechanism for adjusting bacterial population density, which relies on signal molecules, such as autoinducers (AI). QS systems are potential gene regulators that powerfully control hundreds of genes to directly or indirectly impact various aspects of bacterial physiology features and virulence potency, including cell adherence, motility, and biofilm formation, which are important factors for bacterial growth and migration in normal or adverse environments [45,46,47]. Sophisticated transcriptome and proteomics analyses suggest that QS systems are not only a local regulatory mechanism but also controlling global bacterial behaviors, including aforementioned virulence characteristics [48]. Similarly, QS also plays an important regulatory role in Gram-negative bacteria, controlling the collective behavior of Gram-negative bacteria by secreting signal molecules [49]. Increasing studies have shown that bacterial biological characteristics are strongly associated with the QS systems; for example, biofilm-forming bacteria use QS to up- or down-regulate gene expression that may be related to CRISPR-Cas systems and will be discussed in more detail in next sections [50, 51] .
CRISPR-Cas systems may sense QS systems
Shortly after discovering Cas9 gene-editing potential, CRISPR-Cas systems (Type I-F) of P. aeruginosa were implicated in impacting various bacterial functions [52, 53]. As biofilm formation is primarily regulated by QS systems, roles of CRISPR-Cas in biofilm formation may be related to QS as reported in QS in Serratia sp. ATCC39006 strain [54, 55], which may be independent of N-acyl homoserine lactone (AHL). In contrast, some homologous proteins of AI receptors LuxIR, such as SmaI and SmaR, influence both the adaption and interference process of CRISPR-Cas as well as expression of cas operons in Serratia sp. [54]. Through the electrically-controlled CRISPR-based transcriptional regulation, the activating effect of CRISPR-Cas by QS is also observed in another bacterium, E. coli, facilitating communication between bacterial cells [56]. In Gram-negative bacteria, such as P. aeruginosa and marine bacterium Chromobacterium violaceum, QS interfering enzymes lactonase SsoPox may down-regulate the expression of CRISPR-Cas, providing indirect evidence for QS reciprocal effects on CRISPR-Cas [57]. Recently, RNA-seq analysis found that the expression of Type I-F CRISPR-Cas was significantly increased in QS mutant strains ΔlitR of Aliivibrio wodanis, implying that LitR, a master QS regulator, negatively affected Type I-F CRISPR [58]. These findings suggest that cas genes are regulated by QS systems when bacteria reach to a high density, then leading to increased integration of spacer sequences of exogenous DNA fragments, and thus heightening the CRISPR-Cas-mediated resistance to invasion of phages and other genetic elements.
Interaction between QS and CRISPR-Cas
Given the above-mentioned discussion, there is likely mutual adjustment between CRISPR-Cas and QS systems. The adaptive immune function of CRISPR-Cas is often inhibited when the QS signaling pathway is absent or blocked, suggesting that QS systems play an indispensable role in CRISPR-Cas activation [59]. Consistent with this conception, cdpR, a QS negative regulatory gene, impeded the transcription of cas operon via interrupting the binding of virulence factor regulator to cas1 promoter induced by LasI/RhlI-mediated autoinducers, dampening Cas3 nuclease activities [60]. Mechanistically, small regulatory RNAs in bacteria may augment CRISPR levels to facilitate the function against phage infection, which is targeted on a leader sequence to negatively affecting Rho-mediated transcription termination [61]. Collectively, these findings indicate that QS-mediated CRISPR-Cas regulation may be a common phenomenon within the bacterial signaling network but much of mechanistic detail remains undetermined and requires further study. The QS regulatory pathway serves as homeostasis mechanism to maintain the balance between phage infection risk and CRISPR-Cas quantities, and contributes to the precise control of the adaptive immunity in bacteria [54, 59].
Based on above-discussed observations, we speculate that QS systems and CRISPR-Cas systems may interact each other to regulate the expression of various genes and alter bacterial behaviors, including immunity and virulence. For example, QS may effectively modulate the expression of CRISPR-Cas systems and thereby altering their function. In return, CRISPR-Cas systems render strong feedback to QS systems by influencing the expression and function of some critical QS regulators, such as the lasR gene, which may have far-reaching effects on the physiology and virulence potency of the involved bacterium [24]. Additionally, we have found that deletion of cas3 in Salmonella enterica induced significant upregulation of QS systems by using the transcriptome-wide screen (RNA-Seq) [62].
While the interaction between CRISPR-Cas and QS systems may be well documented, the regulatory pathways and molecular mechanisms of QS and CRISPR-Cas activation remain to be fully elucidated [63]. Further exploring the mutual regulation mechanism in broader scopes and deeper levels (e.g., diverse species and strains as well as many different signaling pathways) will be necessary to improve our knowledge of how CRISPR-Cas and QS systems are intertwined to control bacterial physiology and virulence.
CRISPR-Cas systems modulate bacterial pathogenicity
CRISPR-Cas systems potentially adjust bacterial pathogenicity
Next, we will discuss how some of the above-mentioned regulations impact bacterial pathogenicity (Table 1). A series of studies indicate a potential link between CRISPR-Cas and bacterial virulence, which may directly or indirectly alter the course or extent of host defense in mammals. A mouse urinary tract infection model with Enterococcus faecalis isolates [32, 73] demonstrated that type II CRISPR-harboring strain manifests increased mortality compared to the CRISPR negative strain as a control. In addition to influencing the genomic differences that lead to bacterial pathogenicity through two non-mutually exclusive processes, CRISPR-Cas also blocked the recognition of exogenous DNA mobile elements carrying virulence factors (endotoxin, exotoxin, or drug resistance genes), thereby reducing bacterial virulence and limiting potential secondary infections [32]. Compared to the attenuated strain of C. jejuni, highly virulent strains bear a shorter or complete absence of CRISPR sequence, indicating that deleting CRISPR increases virulence and leads to severe enteritis and serious complications of infection [70].
CRISPR-Cas systems might either enhance or inhibit the virulence of divergent bacteria depending on different situations. Patients who were infected with Klebsiella pneumoniae bearing the type I-E* CRISPR-Cas (both Cas1 type B and Cas3 type B alleles positive) system showed higher intensive care unit (ICU) occupancy, mortality rates and higher virulence gene clustered in comparison with patients infected with CRISPR-negative isolates [64]. Deletion of cas3 gene (the representative nuclease of the class 1 type I CRISPR-Cas system) in Porphyromonas gingivalis, increased invasive potency in human monocyte/macrophage cell line THP-1 [74]. Furthermore, research illustrated that CRISPR-Cas could prevent the acquisition of certain pathogenic factors in some E. faecalis isolates to influence its pathogenic traits [65]. Collectively, these findings argue that CRISPR-Cas systems indeed shape the host responses through a spectrum of distinct mechanisms in experimental conditions and/or potentially clinical scenarios.
CRISPR-Cas systems are likely associated with virulence regulation
Koonin and colleagues unveiled that prokaryotic toxins are closely associated with CRISPR-Cas, which induce either bacterial death or dormancy in the case of an immune system failure [75]. During the study of 81 strains of Shiga toxin producing Escherichia coli (STEC), Magaly et al. found that the amount of bacterial spacer sequences was inversely correlated with the severity of the disease, the fewer the spacer sequences, the higher the pathogenicity. The reason is that, the less CRISPR-cas system activity, the lower the chance to acquire pathogenicity features. In addition, the author found no relevance between the presence of subtype I-E cas and virulence genes, suggesting that these events may prevent the uptake and acquisition of virulence genes. Therefore, CRISPR array length and virulence genes are likely to be associated with other factors, which need to be further evaluated [76]. Vibrio cholerae in the GI tract could encode CRISPR-Cas genes along with the T6SS (VgrG, Hcp, and PAAR), indicating the mobility of CRISPR-Cas and its relevance to bacterial virulence [77].
However, another study reached conflict conclusions: genotypical screening based on PCR approaches investigated relationship between CRISPR elements and virulence genes in E. faecalis, including ace, asa1, cylA, efaA, ebpR, esp, gelE, and hyl. The researchers found that the presence of CRISPR loci seemed not relevant to the decrease of the number of pathogenic factors and found that CRISPR1-cas deficiency was only related to esp, cylA, asa1 gene deficiency. These discoveries indicate that CRISPR loci may partially repress the expression or function of virulence factors and Pathogenicity Island [65].
CRISPR-Cas systems impact endogenous genes (bacterial own genome) to Alter host immune responses
Endogenous targeting by type II CRISPR-Cas
In addition to targeting foreign mobile genetic elements, CRISPR spacers also target self-sequences. This self-targeting (autoimmunity) may be harmful to the stability of bacterial host genome, but it is not fatal under specific circumstances [78]. Increasing evidence suggests that Cas9, abundant in the genome of pathogenic bacteria, regulates virulence factors in several bacteria [21]. The Type II CRISPR-Cas systems of Francisella novicida are implicated in influencing bacterial growth, differentiation and virulence, and thereby leading to altered host defense by decreasing the expression of bacterial lipoprotein (BLP), a pathogen-associated molecular pattern (PAMP) antigen (Fig. 3) [21].

Type II and type I CRISPR-Cas systems adjust the expression of endogenous transcripts. TracrRNA and scaRNA (small, CRISPR-Cas-associated RNA) form a dual-RNA complex through sequence recognition to the CRISPR repeats. This dsRNA structure enables the free terminal of the tracrRNA to interact through a non-identity matching mRNA encoding the BLP, the stability of BLP mRNA is impeded and thereby leading to the degradation of mRNA. Type I CRISPR-Cas system of PA14 targets the mRNA of the QS regulator LasR. The crRNA1-12 structure is associated with Cascade (Csy1-4 complex) and interacts with lasR mRNA through a sequence complementing a part of crRNA1-12, and resulting in lowered lasR mRNA stability
This study as a pilot test indicates that CRISPR-Cas systems contribute to virulence regulation in bacteria and hence influence their survival in the natural hosts [66]. Furthermore, Cas9 may influence the adherence of bacteria to the surface of host cells to modulate the composition and structure of bacterial envelope [21]. Recently, another study uncovered the loss of adherence to human nasopharyngeal epithelia by a cas9 deletion meningococcus mutant may be dependent on the indirect regulation instead of influencing adherence genes’ expression [67]. Hence, the meningococcal CRISPR-Cas systems may exhibit novel functions beyond their classical duty in bacterial host defense against foreign DNA [67].
Endogenous targeting by type I CRISPR-Cas
Type I CRISPR-Cas systems of Pseudomonas PA14 were shown to alter the levels of lasR mRNA to influence the immune response of macrophages. This work found that the lasR mRNA region (648 to 687) contains critical homologous sequences to part of spacers (Type I CRISPR array), and that the first 2 nt of the 5′-GGA-3′ site serves as key PAM-like nucleotides for this targeting. As Type I CRISPR-Cas decreased lasR mRNA stability, bacteria evaded the recognition and clearance of TLR4-mediated response by host immune cells, thus resulting in lowered inflammatory response [24] (Fig. 3).
Hence, the type I CRISPR-Cas systems may indirectly influence host defense by influnencing inflammasome activation via regulation of autophagy or other unknown signaling pathways [68]. According to the phenotypic determination of virulence characteristics and transcriptome analysis, it can be speculated that CRISPR-cas9 is involved in regulating a large number of virulence related genes, such as downregulating membrane coding genes, lipoprotein genes and flagellum related genes. Compared to WT bacteria, the Δcas9 mutant showed a decrease in four aspects: mobility, biofilm formation ability, intracellular viability and toxin production ability. Thus, Cas9 contributes to the virulence of C. jejuni [69]. Interestingly, the expression of Cas9 protein in strains lacking CRISPR loci also increased the virulence of bacteria, suggesting that the function of Cas proteins may be exerted through several distinct mechanisms, some of them likely independent of CRISPR transcript levels [70]. It has been reported that cas9 cannot only target DNA, but also cleave endogenous RNAs to influence gene inactivation or activation [79].
CRISPR-Cas endogenous targeting and autoimmunity
CRISPR-Cas systems serve as not only a powerful immune weapon for bacteria to resist foreign invaders, but also an impactful regulator of the expression of endogenous genes, exemplified by the E. coli CRISPR-Cas. CRISPR-Cas systems of E. coli regulate endogenous genes and signaling pathways, which may be active only under specific conditions through a variety of mechanisms, such as DNA interactions, mRNA sequence targeting, and transcription or elongation regulation [80]. Multiple studies demonstrated bacteria containing self-targeting spacers (STS), for instance, CRISPR spacers targeting protospacers on the same genome to exhibit adverse effects. STSs are thought to provoke autoimmunity, an unwanted side effect of CRISPR-Cas defense, while the newer concept points out perhaps a regulatory mechanism for gene expression. Nobrega et al. found the existence of STS in all CRISPR-Cas types and 20% of all CRISPR-carrying bacteria. In particular, up to 40% of I-B and I-F CRISPR-Cas systems in E. coli contained STS.
Intriguingly, although STSs are almost ubiquitous, they hardly cause serious adverse consequences to the bacteria themselves. This may be related to the presence of anti-CRISPR (Acr) proteins, which may serve as a self-regulatory mechanism for the avoidance of autoimmunity [81]. This phenomenon ponders many scientists and encourages further understanding of the underlying regulatory mechanisms critical to maintaining the stable equilibrium of bacteria by countering or fine-tuning the self-targeting elements. Recently, it has been demonstrated that type II-B CRISPR-Cas9 systems and type V-A CRISPR Cas12a systems inhibit transcription and defense through CRISPR RNA (crRNA) guidance and limit complementarity between target genes [82]. The CRISPR systems can target endogenous genes to affect the physiological function of bacteria. CRISPR-Cas systems that target the bacterial genome are thought to have a lethal effect on the host, but a small number of bacteria can escape this CRISPR-Cas attack through genome remodeling. In the model organism Streptococcus thermophilus DGCC710, the escape of self-targeting of endogenous type I and type II CRISPR-Cas systems is primarily the result of the removal of low-frequency defective plasmids in the targeted spacer [83]. This helps explain that side effects are generally limited with CRISPR-Cas self-targeting, leading to the reconciliation that self-attack perhaps in some instances may serve as potential self-conservation or protection mechanisms by removing unimportant or decayed genetic materials in the genome.
To study roles of acetyl-CoA in microbial energy and material metabolism, researchers have used a tunable CRISPR interference (CRISPRi) system for multiplex repression of the expression of endogenous genes (pta, frdA, ldhA, and adhE) in Escherichia coli, leading to reduced end products of carbon metabolism and carbon flux and increased acetyl-CoA. These results demonstrated that the CRISPRi system could serve as a tool for simultaneously regulating bacterial genes to analyze biosynthetic pathways, namely bacterial metabolism. Although this method provides a technical platform for other metabolic research, potentially incomplete silencing CRISPR may partially indicate a possible function attributing to the CRISPR’s natural features, endogenous regulation of metabolisms, which needs to be investigated [84].
Complex roles of CRISPR-Cas in influencing host immunity
cas3 of S. enterica may play an active role in its virulence, including cell penetration and animal pathogenicity, by impacting several genes involved in QS, flagellum, and SPI-1-T3SS systems [62]. Additionally, Cas9 may also promote the invasion and proliferation of Neisseria meningitidis in hosts, suggesting that the role of type II CRISPR-Cas in virulence modulation varies with bacteria and their respective environments [21]. C. jejuni seems to contain a complex CRISPR system in the regulation of pathogenicity; and mutants lacking csn, a type II CRISPR-Cas marker, increase swarming and reduce virulence during infection of the host. The lack of Csn also promoted the binding of antibodies to cell surface molecules, indicating that Csn is involved in regulating the conformation and/or composition of the cell membrane [70]. In addition, CRISPR-Cas systems in Shigella are widespread, and Shigella strains without CRISPR1 (a specific CRISPR array in Shigella genome) exhibit higher pathogenicity and virulence [85].
On the contrary, CRISPR-Cas systems may contribute to increased invasive potency and knockdown of them may lead to lower virulence. In a Salmonella pullorum strain, CRISPR components contributed to the bacterial dispersion and survival in hostile habitats, manifesting increased virulence [86]. Likewise, Legionella pneumophila requires CRISPR-Cas for pathogenesis; for instance, the cas2 gene seemed to be responsible for causing infection in amoebae. However, mutant strains lacking cas9, cas1, and cas4 did not show apparent roles in the infection of macrophages and aquatic amoeba indicating that each of Cas proteins plays a distinct role in pathogenesis [71]. Hence, the detailed mechanisms of Cas2 in regulating bacterial virulence remain unclear and warrant further research. A study showed that RNase activity of Cas2 may be related to the virulence mRNA expression by activating RNase [72]. Wu et al. found that P. aeruginosa type I CRISPR-Cas system affects bacterial virulence, the absence of type I CRISPR-Cas system led to a more serious disease phenotype in the host. Compared with WT P. aeruginosa (PA14), the expression of NLRC4 inflammasome and the level of pro-inflammatory cytokine IL-1β in lung of infected mice significantly increased in total CRISPR-Cas regions (TCR) knockout bacteria group. Concurrently, it was also discovered that type I CRISPR-Cas system involved in the reactive oxygen species of mitochondria and participates in the activation of inflammasome by influencing the release of mtDNA [68] (Fig. 4).

CRISPR-Cas impacts host defense by regulating several signals involving innate immunity and inflammatory responses. CRISPR-Cas may participate in the recognition of pattern recognition receptors (PRRs) (e.g., TLR2, TLR4 or other unknown PRRs) and the activation of their downstream signaling pathways. The downstream effects include altered levels of mitochondrion ROS, inflammasome, autophagy, depletion of CRISPR-Cas in P. aeruginosa strain PA14 leads to exaggerated inflammation and organ damage through rapid nuclear translocation of the transcription factor NF-κB after PRRs recognition
Together, these studies suggest that different CRISPR-Cas systems play distinct roles in bacterial pathogenesis upon infecting hosts, which requires continued investigations. Another possibility is that Type I-E CRISPR-Cas systems may only partially modulate the transcription of certain endogenous mRNAs as a means to gauging and modulating network interactions within the bacteria (e.g., E. coli) to combat phages through naïve adaptation. This type of CRISPR self-regulation could cause a bystander effect that may also indirectly change the landscape of host immune response, which may not be true endogenous targeting and impact interpretation of how the host responds to bacterial infection [80]. Overall, although a large number of literature pointed out that the potential endogenous targeting is linked to immune reactions in the host, further experimentation is urgently required to address the complexity and underlying mechanisms with various cas genes in regulating virulence in the broader bacterium kingdom. Collectively, these findings suggest that different CRISPR-Cas systems play distinct roles in bacterial pathogenesis upon infecting hosts, which requires continued and deeper investigations.
Reconciliation with nucleic cleavage in endogenous targeting
The widespread existence of CRISPR-Cas systems in prokaryotes suggests that the possibility of self-targeting to the homologous regions in their genomes may cause potential autoimmunity or modulate defense mechanisms [19, 87]. Sequencing analyses unveil that the homologous regions are not generally long enough to cause extensive self-destruction and unlikely induce organism death. Further, other intrinsic mechanisms, such as homologous recombination, may provide the plasticity and remodeling of bacterial genomes, and thereby leading to evasion of internal genome targeting [83].
Although the evidence for regulating endogenous gene expression by CRISPR-Cas systems to alter bacterial behaviors instead self-killing is mounting, how this occurs is largely unclear. Particularly, the evidence for direct nuclease cleavage of bacterial endogenous genes on genomic loci or transcripts is scarce, and often difficult to gain solid verifications in vivo due to the current technical hindrances.
The most extensively-studied Cas9 was earlier demonstrated to only cleave DNA but is now implicated in cleavage of both DNA and RNA depending on bacterial species and strains as well as the experimental conditions [88]. For example, the C. jejuni Cas9 (CjCas9) is implicated in binding and cleaving complementary endogenous mRNAs in a crRNA-dependent manner by Dugar et al., who recently reported that about hundreds of transcripts were co-immunoprecipitated with CjCas9 through their base-pairing with crRNAs, and, importantly, some of these RNAs could be enzymatically cleaved at the predicted binding sites [79]. Mutational analyses show that the cleavage was crRNA and tracrRNA dependent and requiring the CjCas9 HNH domain.
The study conducted in Type II CRISPR-Cas of F. novicida initially reported cleavage of mRNA of BLP, but later the authors found that it was hard to reproduce the data [18, 21]. The other study performed in our laboratory found the Cas3-mediated cleavage of lasR mRNA in P. aeruginosa strain PA14 by Type I-F CRISPR-Cas systems that were thought to only target and cleave DNA (Fig. 3). However, this cleavage seemed to be irreproducible through the purified protein system by the third party [89], which might be due to the impure protein mixture and the experiment conditions and reagents that were utilized in our initial report [24].
Although Cas9 is shown to cleave endogenous RNA, Wiedenheft and colleagues have indicated the irreproducibility of RNA cleavage by Type I-F CRISPR systems in our early report [79, 89]. We extremely appreciate the challenge to our in vitro RNA cleavage assay, and would like to sincerely apologize to the scientific community for the inconvenience this observation may have caused. Furthermore, the RNA cleavage has never been tested in cells or animal models, which require additional validation for possibilities targeting this RNA transcript or other endogenous RNA (potentially DNA) sequences. Nevertheless, we were able to reproduce the increased expression of lasR in the PA14 strain with deletion of Total CRISPR-Cas regions (TCR) by independent lab members [24, 68]. In addition, the deletion of cas3 in another lab could not detect strong inhibition of lasR mRNA [90], which might be due to several differences, such as the strains used, experimental settings, as well as mutation region (total CRISPR vs. cas3) and approaches used for generating these mutations.
Nevertheless, nuclear acids cleavage assays thus far were only shown in experimentally reconstituted in silico, there was no direct evidence demonstrating that endogenous targeting occurs in vivo in bacteria, in mammalian cells, or whole animals after the bacterial invasion. Therefore, the studies regarding CRISPR-mediated endogenous gene regulation in bacteria or pathogenic effects on mammalian hosts might not be associated with nucleic acid cleavage but are indirectly related to the transcription, expression, or decay of the underlying gene and nearby genes. In addition, endogenous targeting might be attributed to other unrecognized mechanisms. The function and regulation of CRISPR-Cas systems are highly diverse, surprising, and somewhat unpredictable. It is imperative to further dissect the critical mechanisms of how endogenous genes can be targeted, regulated, and/or cleaved to influence bacterial physiology, virulence, and ultimately impact the courses and potencies of mammalian immune responses, such as phagocytosis and inflammatory responses by professional and unprofessional immune cells.
Previous reports on non-canonical roles in pathogenesis were primarily involved in Type I and Type II CRISPR, a recent study revealed that the Type IIIA system in M. tuberculosis (Type III-A system) might be a new player to directly target host responses through secretion of CRISPR-Cas effectors into extracellular compartments. The authors showed that secreted Csm1, 3, 5 and 6, and Cas6 proteins activated immune responses in both cell culture and animal experiments as well as IFN-γ release in T cells and macrophages from active tuberculosis patients [20]. This study and some other observations indicate that pathogens may utilize CRISPR-Cas proteins to exert non-canonical function as virulence factors to retour host immune responses and inflammatory responses to cause more severe host tissue damage [20, 91], which seems irrelevant to endogenous targeting and should be taken into consideration when investigating endogenous CRISPR-Cas targeting for the involvement of host immunity.
CRISPR-Cas systems influence antibiotic resistance
CRISPR-Cas and antibiotic resistance
As the immune apparatus found in prokaryotes, CRISPR-Cas systems represent a powerful weapon to counteract against invasive exogenous genetic elements and phages. Remarkably, the structure and function of CRISPR-Cas are also potentially linked to antibiotic resistance. Acinetobacter baumannii has strong virulence and increasingly serious drug resistance [92]. Recently, Tyumentseva M et al. studied core features of antibiotic resistance genes through sequence analyses of CRISPR arrays in 12 clinical isolates of A. baumannii by using multilocus sequence typing schemes. Their results showed that clinical isolates of strong drug resistance genes contain less CRISPR arrays and active cas genes, suggesting that CRISPR-Cas systems can inhibit the drug resistance in A. baumannii [93].
Cas9 mutations in C. jejuni rendered strong sensitivity to antibiotics compared to WT strains, suggesting that Cas9 likely promotes antibiotic resistance [94]. A recent clinical study with 168 carbapenem-resistant Klebsiella pneumoniae (CRKP) strains from inpatients identified both CRISPR-Cas-positive and CRISPR-negative isolates. The results showed that patients infected by strains harboring the type I-E* CRISPR-Cas system manifested increased mortality vs. patients infected by CRISPR-negative isolates. In the CRISPR-negative strains, the frequency of carbapenemase gene KPC-2, an important drug resistance gene in K. pneumoniae, was the highest compared to CRISPR-containing strains. This study suggested that there is a reverse correlation between CRISPR systems and antibiotic-resistance, providing strong impact on antibiotic resistance in the clinical strains [64].
The possible reason for CRKP strains with positive CRISPR-Cas system still obtain carbapenem gene is that different CRISPR-Cas systems have different spacers, CRISPR-Cas systems interferes with the acquisition of carbapenem gene, which not only depends on the spacer matching in CRISPR-Cas system, but also is affected by the PAM recognition of foreign carbapenem gene. The abuse of carbapenem antibiotics will help bacteria obtain carbapenemase gene [95]. Some CRISPR-Cas positive strains have been inactivated for a long time because of the existence of self-targeting spacers [96]. There are anti-CRISPR proteins (Acrs) in bacteria to inhibit the immune function of CRISPR-Cas systems [97]. This may be associated with one of the common antibiotic resistance mechanisms by hiding bacteria within biofilms (up to 1000 times compared to their planktonic counterparts), often resulting in persistent infection and very difficult to eradicate [98].
From the clinical prospect, the ability to promote antibiotic resistance by CRISPR-Cas is considered as a disadvantage by upregulating biofilm and thereby leading to resistance to phage therapy. It should be noted that phage therapy is becoming an important alternative or auxiliary therapy for multidrug-resistant bacterial infections. A thorough analysis of the relationship between CRISPR-Cas and bacterial antibiotic resistance would improve understanding of drug-resistance mechanisms and potentially reveal new strategies for the prevention and treatment of refractory infection [23]. Recently discovered Acrs can counteract the CRISPR-Cas immunity towards phages and thus hold promise for clinically combatting bacterial infection [99,100,101].
CRISPR-Cas as targets to counteract antibiotic resistance
On the other hand, CRISPR-Cas systems may be exploited to reduce drug resistance and augment the mammalian defense against infection. For instance, targeting CRISPR-Cas DNA sequences interferes with the horizontal transfer of a specific Staphylococcal resistance gene, limiting the spread of drug-resistant pathogens. This may represent a potential drug therapy target and further understanding detailed mechanism is a must for biotechnological development [102, 103]. Interestingly, some bacterial strains lacking CRISPR-Cas are inclined to acquire antibiotics resistance genes, and lack of CRISPR-Cas appears to render bacteria susceptible to bacteriophage attack. This phenomenon is further compounded with variable observations in different types of bacteria (with varying CRISPR systems) and/or environmental settings [60, 90]. The study of phylogenetic distribution in P. aeruginosa showed that CRISPR-Cas systems acquired drug-resistant genes through reconstructing auxiliary genome distribution [104]. The researchers analyzed 75 isolates of Salmonella from chicken farms, found that nearly 80% of the strains were resistant to one or more antibiotics. Through counting the Spearman’s rank correlation of the CRISPR-Cas system and antibiotic resistance genes of the strains, the study unveiled that the CRISPR loci were closely related to the antibiotic resistance genes [105].
CRISPR-Cas systems block genetic transfer
Horizontal gene transfer is the main mode of causing bacterial drug resistance, leading to the spread of superbug infection [106, 107]. A study uncovered that multidrug-resistant enterococci lacked CRISPR-Cas elements and showed that the increased resistance of E. faecalis to antibiotic was due to the loss of CRISPR-Cas locus in the gene sequence. At the same time, it was also found that E. faecalis had only CRISPR locus without Cas protein and interestingly, its genome still contains rich antibiotic resistance genes, suggesting that only a certain component of CRISPR-Cas may be sufficient to alter bacterial drug resistance transmission [108].
Another recent study with K. pneumoniae revealed that the antimicrobial resistance of CRISPR-Cas positive strains was lower than those negative strains [109]. Importantly, type I-E systems might alter the spread of blaKPC in K. pneumoniae [110]. CRISPR-Cas systems may inhibit the acquisition of blaKPC plasmids contributing to carbapenem resistance in K. pneumoniae strains because of possessing CRISPR spacers that are in high similarity to known multidrug-resistance plasmids. Correspondingly, resistance to blaKPC plasmid in the K. pneumoniae strain hosting CRISPR-Cas may be abolished through removal of the CRISPR-Cas cassette [109]. These studies further confirm the complexity of CRISPR-Cas function in drug resistance, which warrants further investigation.
Mechanisms of CRISPR-Cas-assisted drug resistance
Interestingly, most non-pathogenic S. epidermidis strains lack CRISPR sequences, while a clinical isolate, RP62a containing CRISPR sequences, showed resistance to the invasion of exogenous genes to a certain extent [111]. To examine the mechanistic feature of CRISPR in drug resistance, the beta-lactam plasmid pG0400 in S. aureus is deactivated and transformed into CRISPR S. epidermidis strains or control strains. The results showed that CRISPR-Cas interference significantly impaired horizontal transfer of drug-resistant plasmids (from S. aureus to S. epidermidis), which may represent a mechanism of blocking drug resistance [102]. Some CRISPR spacers of Mollicutes could target bacterial or phage genes, which may serve as part of integrative and conjugative elements (ICE) [112].
Recently, E. faecalis as a model organism and a natural colonizer of the mammalian intestine that contains pheromone-responsive plasmids (PRPs) could mediate inter- and intra-species transfer of antibiotic resistance genes. WT bacteria containing CRISPR-Cas can block the transfer and dissemination of antibiotic-resistant plasmids but the Δcas9 mutant strain cannot [113]. Overall, we only know the tip of an iceberg about the role of CRISPR-Cas in regulating drug resistance. Critically, we point out that multiple approaches, especially novel ones, are highly desired to further unravel the functions for CRISPR-Cas systems in antibiotic resistance in depth.
Conclusions and perspectives
In summary, the role of CRISPR-Cas systems has gone beyond the canonical function, adaptive immunity. One of these non-canonical functions of the CRISPR-Cas systems in regulating bacterial genes that are involved in invading mammalian hosts is described as an example in Figs. 3 and 4. In addition to adaptive immunity, the function of CRISPR-Cas systems is gradually unfolded by modifying the expression of some critical bacterial genes, thereby impacting their physiological characteristics. Numerous studies have emphatically demonstrated that bacterial functions may be affected by CRISPR-Cas systems. Understanding CRISPR-Cas-mediated gene regulation is mainly derived from unbiased screening for regulators of distinct phenotypes, such as virulence or bacterial physiology behaviors [114]. Importantly, the knowledge of CRISPR-Cas systems is constantly evolving at an enormous pace. Recently, type VI CRISPR-Cas systems and their related protein Csx27 have been implicated in similar regulation of virulence in E. coli. Csx27 forms a membrane channel of solid DNA and may be involved in the ubiquitin signaling pathway, which serves as new machinery for bacterial degradation of exogenous DNA [115]. Therefore, it might be inferred that the ever-expanding CRISPR-Cas systems regulate far more genes than have been identified to date while through a spectrum of different mechanisms.
Dissecting these non-canonical functions of CRISPR-Cas systems has broadened the understanding of bacterial physiological functions and regulatory networks. Furthermore, creative strategies for studying interaction and inter-regulation between CRISPR-Cas systems and bacterial virulence genes can also guide researchers to design experiments for fundamental research or biotechnological application, such as CRISPR-targeting antimicrobial agents. It is worth mentioning that harnessing the native type I-F CRISPR-Cas system has been implemented to delete the AMR (antimicrobial resistance) gene in multidrug-resistant P. aeruginosa [116]. Improved understanding of the influence of CRISPR-Cas systems on bacterial physiology and pathogenicity will have a profound impact on the control of bacterial infection as well as continuously providing new biotechnological tools to control genetic diseases, cancers and so on.
Availability of data and materials
Not applicable.
Abbreviations
- CRISPR-Cas:
-
Clustered regularly interspaced short palindromic repeats-CRISPR associated proteins
- EPS:
-
Extracellular polymeric substances
- PAM:
-
Protospacer-adjacent motif
- sRNAs:
-
Small non-coding RNAs
- c-di-GMP:
-
Cyclic diguanylate monophosphate
- AI:
-
Autoinducers
- AHL:
-
Acyl homoserine lactone
- QS:
-
Quorum sensing
- ICU:
-
Intensive care unit
- BLP:
-
Bacterial lipoprotein
- PAMP:
-
Pathogen-associated molecular pattern
- STS:
-
Self-targeting spacers
- CRKP:
-
Carbapenem-resistant Klebsiella pneumoniae
- Acrs:
-
Anti-CRISPR proteins
- ICE:
-
Integrative and conjugative elements
- PRPs:
-
Pheromone-responsive plasmids
- WT:
-
Wild type
References
Marchfelder A. Special focus CRISPR-Cas. RNA Biol. 2013;10(5):655–8. https://doi.org/10.4161/rna.24687.
Ishino Y, Krupovic M, Forterre P. History of CRISPR-Cas from encounter with a mysterious repeated sequence to genome editing technology. J Bacteriol. 2018;200(7):e00580–17. https://doi.org/10.1128/JB.00580-17.
Bernheim A, Bikard D, Touchon M, Rocha EPC. Atypical organizations and epistatic interactions of CRISPRs and cas clusters in genomes and their mobile genetic elements. Nucleic Acids Res. 2020;48(2):748–60. https://doi.org/10.1093/nar/gkz1091.
Liu Y, Pinto F, Wan X, Yang Z, Peng S, Li M, et al. Reprogrammed tracrRNAs enable repurposing of RNAs as crRNAs and sequence-specific RNA biosensors. Nat Commun. 2022;13(1):1937. https://doi.org/10.1038/s41467-022-29604-x.
Karvelis T, Gasiunas G, Miksys A, Barrangou R, Horvath P, Siksnys V. CrRNA and tracrRNA guide Cas9-mediated DNA interference in Streptococcus thermophilus. RNA Biol. 2013;10(5):841–51. https://doi.org/10.4161/rna.24203.
Kunin V, Sorek R, Hugenholtz P. Evolutionary conservation of sequence and secondary structures in CRISPR repeats. Genome Biol. 2007;8(4):R61. https://doi.org/10.1186/gb-2007-8-4-r61.
Makarova KS, Haft DH, Barrangou R, Brouns SJ, Charpentier E, Horvath P, et al. Evolution and classification of the CRISPR-Cas systems. Nat Rev Microbiol. 2011;9(6):467–77. https://doi.org/10.1038/nrmicro2577.
Koonin EV, Makarova KS, Zhang F. Diversity, classification and evolution of CRISPR-Cas systems. Curr Opin Microbiol. 2017;37:67–78. https://doi.org/10.1016/j.mib.2017.05.008.
Chaudhary M, Mukherjee TK, Singh R, Gupta M, Goyal S, Singhal P, et al. CRISPR/Cas technology for improving nutritional values in the agricultural sector: an update. Mol Biol Rep. 2022. https://doi.org/10.1007/s11033-022-07523-w.
Wong ACS, Massel K, Lam Y, Hintzsche J, Chauhan BS. Biotechnological road map for innovative Weed Management. Front Plant Sci. 2022;13:887723. https://doi.org/10.3389/fpls.2022.887723.
Aregger M, Xing K, Gonatopoulos-Pournatzis T. Application of CHyMErA Cas9-Cas12a combinatorial genome-editing platform for genetic interaction mapping and gene fragment deletion screening. Nat Protoc. 2021;16(10):4722–65. https://doi.org/10.1038/s41596-021-00595-1.
Elliott EK, Haupt LM, Griffiths LR. Mini review: genome and transcriptome editing using CRISPR-cas systems for haematological malignancy gene therapy. Transgenic Res. 2021;30(2):129–41. https://doi.org/10.1007/s11248-020-00232-9.
Lin P, Shen G, Guo K, Qin S, Pu Q, Wang Z, et al. Type III CRISPR-based RNA editing for programmable control of SARS-CoV-2 and human coronaviruses. Nucleic Acids Res. 2022;50(8):e47. https://doi.org/10.1093/nar/gkac016.
Li L, Shen G, Wu M, Jiang J, Xia Q, Lin P. CRISPR-Cas-mediated diagnostics. Trends Biotechnol. 2022;S0167-7799(22):00104. https://doi.org/10.1016/j.tibtech.2022.04.006.
Shaikh SB, Bhandary YP. CRISPR/Cas9 genome editing tool: a promising tool for therapeutic applications on respiratory diseases. Curr Gene Ther. 2020;20(5):333–46. https://doi.org/10.2174/1566523220666201012145731.
Klose SM, Wawegama N, Sansom FM, Marenda MS, Browning GF. Efficient disruption of the function of the mnuA nuclease gene using the endogenous CRISPR/Cas system in mycoplasma gallisepticum. Vet Microbiol. 2022;269:109436. https://doi.org/10.1016/j.vetmic.2022.109436.
Kong L, Jiang D, He C, Xia J, Wei H, Zhou L, et al. TgROP18 targets IL20RB for host-defense-related-STAT3 activation during toxoplasma gondii infection. Parasit Vectors. 2020;13(1):400. https://doi.org/10.1186/s13071-020-04251-7.
Ratner HK, Escalera-Maurer A, Le Rhun A, Jaggavarapu S, Wozniak JE, Crispell EK, et al. Catalytically active Cas9 mediates transcriptional interference to facilitate bacterial virulence. Mol Cell. 2019;75(3):498–510. https://doi.org/10.1016/j.molcel.2019.05.029.
Muller-Esparza H, Randau L. Commentary: type I CRISPR-Cas targets endogenous genes and regulates virulence to evade mammalian host immunity. Front Microbiol. 2017;8:319. https://doi.org/10.3389/fmicb.2017.00319.
Jiao J, Zheng N, Wei W, Fleming J, Wang X, Li Z, et al. M. Tuberculosis CRISPR/Cas proteins are secreted virulence factors that trigger cellular immune responses. Virulence. 2021;12(1):3032–44. https://doi.org/10.1080/21505594.2021.2007621.
Sampson TR, Saroj SD, Llewellyn AC, Tzeng YL, Weiss DS. A CRISPR/Cas system mediates bacterial innate immune evasion and virulence. Nature. 2013;497(7448):254–7. https://doi.org/10.1038/nature12048.
Miyoshi-Akiyama T, Tada T, Ohmagari N, Viet Hung N, Tharavichitkul P, Pokhrel BM, et al. Emergence and spread of epidemic multidrug-resistant Pseudomonas aeruginosa. Genome Biol Evol. 2017;9(12):3238–45. https://doi.org/10.1093/gbe/evx243.
Fouts DE, Matthias MA, Adhikarla H, Adler B, Amorim-Santos L, Berg DE, et al. What makes a bacterial species pathogenic?:comparative genomic analysis of the genus Leptospira. PLoS Negl Trop Dis. 2016;10(2):e0004403. https://doi.org/10.1371/journal.pntd.0004403.
Li R, Fang L, Tan S, Yu M, Li X, He S, et al. Type I CRISPR-Cas targets endogenous genes and regulates virulence to evade mammalian host immunity. Cell Res. 2016;26(12):1273–87. https://doi.org/10.1038/cr.2016.135.
Tong Z, Du Y, Ling J, Huang L, Ma J. Relevance of the clustered regularly interspaced short palindromic repeats of enterococcus faecalis strains isolated from retreatment root canals on periapical lesions, resistance to irrigants and biofilms. Exp Ther Med. 2017;14(6):5491–6. https://doi.org/10.3892/etm.2017.5205.
Sampson TR, Weiss DS. Cas9-dependent endogenous gene regulation is required for bacterial virulence. Biochem Soc Trans. 2013;41(6):1407–11. https://doi.org/10.1042/BST20130163.
Koonin EV, Zhang F. Coupling immunity and programmed cell suicide in prokaryotes: life-or-death choices. Bioessays. 2017;39(1):1–9. https://doi.org/10.1002/bies.201600186.
Guzman NM, Esquerra-Ruvira B, Mojica FJM. Digging into the lesser-known aspects of CRISPR biology. Int Microbiol. 2021;24(4):473–98. https://doi.org/10.1007/s10123-021-00208-7.
Venkatesan N, Perumal G, Doble M. Bacterial resistance in biofilm-associated bacteria. Future Microbiol. 2015;10(11):1743–50. https://doi.org/10.2217/fmb.15.69.
Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis. 2002;8(9):881–90. https://doi.org/10.3201/eid0809.020063.
Del Pozo JL. Biofilm-related disease. Expert Rev Anti-Infect Ther. 2018;16(1):51–65. https://doi.org/10.1080/14787210.2018.1417036.
Bourgogne A, Garsin DA, Qin X, Singh KV, Sillanpaa J, Yerrapragada S, et al. Large scale variation in enterococcus faecalis illustrated by the genome analysis of strain OG1RF. Genome Biol. 2008;9(7):R110. https://doi.org/10.1186/gb-2008-9-7-r110.
Fu Q, Li S, Wang Z, Shan W, Ma J, Cheng Y, et al. H-NS mutation-mediated CRISPR-Cas activation inhibits phage release and toxin production of Escherichia coli Stx2 phage lysogen. Front Microbiol. 2017;8:652. https://doi.org/10.3389/fmicb.2017.00652.
Suryaletha K, Chandrika SK, Thomas S. Comprehensive genomics depict accessory genes encoding pathogenicity and biofilm determinants in enterococcus faecalis. Future Microbiol. 2021;16(3):175–84. https://doi.org/10.2217/fmb-2020-0111.
Cady KC, O'Toole GA. Non-identity-mediated CRISPR-bacteriophage interaction mediated via the Csy and Cas3 proteins. J Bacteriol. 2011;193(14):3433–45. https://doi.org/10.1128/JB.01411-10.
Barrangou R. The roles of CRISPR-Cas systems in adaptive immunity and beyond. Curr Opin Immunol. 2015;32:36–41. https://doi.org/10.1016/j.coi.2014.12.008.
Jiang F, Doudna JA. CRISPR-Cas9 structures and mechanisms. Annu Rev Biophys. 2017;46:505–29. https://doi.org/10.1146/annurev-biophys-062215-010822.
Hidalgo-Cantabrana C, Barrangou R. Characterization and applications of type I CRISPR-Cas systems. Biochem Soc Trans. 2020;48(1):15–23. https://doi.org/10.1042/BST20190119.
Zegans ME, Wagner JC, Cady KC, Murphy DM, Hammond JH, O'Toole GA. Interaction between bacteriophage DMS3 and host CRISPR region inhibits group behaviors of Pseudomonas aeruginosa. J Bacteriol. 2009;191(1):210–9. https://doi.org/10.1128/JB.00797-08.
Palmer KL, Whiteley M. DMS3-42: the secret to CRISPR-dependent biofilm inhibition in Pseudomonas aeruginosa. J Bacteriol. 2011;193(14):3431–2. https://doi.org/10.1128/JB.05066-11.
Tang B, Gong T, Zhou X, Lu M, Zeng J, Peng X, et al. Deletion of cas3 gene in Streptococcus mutans affects biofilm formation and increases fluoride sensitivity. Arch Oral Biol. 2019;99:190–7. https://doi.org/10.1016/j.archoralbio.2019.01.016.
Gong T, Tang B, Zhou X, Zeng J, Lu M, Guo X, et al. Genome editing in Streptococcus mutans through self-targeting CRISPR arrays. Mol Oral Microbiol. 2018;33(6):440–9. https://doi.org/10.1111/omi.12247.
Chang RYK, Wallin M, Lin Y, Leung SSY, Wang H, Morales S, et al. Phage therapy for respiratory infections. Adv Drug Deliv Rev. 2018;133:76–86. https://doi.org/10.1016/j.addr.2018.08.001.
Kortright KE, Chan BK, Koff JL, Turner PE. Phage therapy: a renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe. 2019;25(2):219–32. https://doi.org/10.1016/j.chom.2019.01.014.
Hover T, Maya T, Ron S, Sandovsky H, Shadkchan Y, Kijner N, et al. Mechanisms of bacterial (Serratia marcescens) attachment to, migration along, and killing of fungal hyphae. Appl Environ Microbiol. 2016;82(9):2585–94. https://doi.org/10.1128/AEM.04070-15.
Chung CH, Fen SY, Yu SC, Wong HC. Influence of oxyR on growth, biofilm formation, and mobility of Vibrio parahaemolyticus. Appl Environ Microbiol. 2016;82(3):788–96. https://doi.org/10.1128/AEM.02818-15.
Azimi S, Klementiev AD, Whiteley M, Diggle SP. Bacterial quorum sensing during infection. Annu Rev Microbiol. 2020;74:201–19. https://doi.org/10.1146/annurev-micro-032020-093845.
Zaitseva IV, Popova AA, Khmel IA. Quorum sensing regulation in bacteria of the family Enterobacteriaceae. Genetika. 2014;50(4):373–91. https://doi.org/10.1134/S1022795414030120.
Papenfort K, Bassler BL. Quorum sensing signal-response systems in gram-negative bacteria. Nat Rev Microbiol. 2016;14(9):576–88. https://doi.org/10.1038/nrmicro.2016.89.
Warrier A, Satyamoorthy K, Murali TS. Quorum-sensing regulation of virulence factors in bacterial biofilm. Future Microbiol. 2021;16:1003–21. https://doi.org/10.2217/fmb-2020-0301.
Mukherjee S, Bassler BL. Bacterial quorum sensing in complex and dynamically changing environments. Nat Rev Microbiol. 2019;17(6):371–82. https://doi.org/10.1038/s41579-019-0186-5.
Heussler GE, Cady KC, Koeppen K, Bhuju S, Stanton BA, O'Toole GA. Clustered regularly interspaced short palindromic repeat-dependent, biofilm-specific death of Pseudomonas aeruginosa mediated by increased expression of phage-related genes. mBio. 2015;6(3):e00129–15. https://doi.org/10.1128/mBio.00129-15.
Gholizadeh P, Aghazadeh M, Asgharzadeh M, Kafil HS. Suppressing the CRISPR/Cas adaptive immune system in bacterial infections. Eur J Clin Microbiol Infect Dis. 2017;36(11):2043–51. https://doi.org/10.1007/s10096-017-3036-2.
Patterson AG, Jackson SA, Taylor C, Evans GB, Salmond GPC, Przybilski R, et al. Quorum sensing controls adaptive immunity through the regulation of multiple CRISPR-Cas systems. Mol Cell. 2016;64(6):1102–8. https://doi.org/10.1016/j.molcel.2016.11.012.
Fu Q, Su Z, Cheng Y, Wang Z, Li S, Wang H, et al. Clustered, regularly interspaced short palindromic repeat (CRISPR) diversity and virulence factor distribution in avian Escherichia coli. Res Microbiol. 2017;168(2):147–56. https://doi.org/10.1016/j.resmic.2016.10.002.
Tsao CY, Hooshangi S, Wu HC, Valdes JJ, Bentley WE. Autonomous induction of recombinant proteins by minimally rewiring native quorum sensing regulon of E. coli. Metab Eng. 2010;12(3):291–7. https://doi.org/10.1016/j.ymben.2010.01.002.
Mion S, Plener L, Remy B, Daude D, Chabriere E. Lactonase SsoPox modulates CRISPR-Cas expression in gram-negative proteobacteria using AHL-based quorum sensing systems. Res Microbiol. 2019;170(6-7):296–9. https://doi.org/10.1016/j.resmic.2019.06.004.
Maharajan AD, Hjerde E, Hansen H, Willassen NP. Quorum sensing controls the CRISPR and type VI secretion systems in Aliivibrio wodanis 06/09/139. Front Vet Sci. 2022;9:799414. https://doi.org/10.3389/fvets.2022.799414.
Hoyland-Kroghsbo NM, Paczkowski J, Mukherjee S, Broniewski J, Westra E, Bondy-Denomy J, et al. Quorum sensing controls the Pseudomonas aeruginosa CRISPR-Cas adaptive immune system. Proc Natl Acad Sci U S A. 2017;114(1):131–5. https://doi.org/10.1073/pnas.1617415113.
Lin P, Pu Q, Shen G, Li R, Guo K, Zhou C, et al. CdpR inhibits CRISPR-Cas adaptive immunity to lower anti-viral defense while avoiding self-reactivity. iScience. 2019;13:55–68. https://doi.org/10.1016/j.isci.2019.02.005.
Lin P, Pu Q, Wu Q, Zhou C, Wang B, Schettler J, et al. High-throughput screen reveals sRNAs regulating crRNA biogenesis by targeting CRISPR leader to repress rho termination. Nat Commun. 2019;10(1):3728. https://doi.org/10.1038/s41467-019-11695-8.
Cui L, Wang X, Huang D, Zhao Y, Feng J, Lu Q, et al. CRISPR-cas3 of Salmonella upregulates bacterial biofilm formation and virulence to host cells by targeting quorum-sensing systems. Pathogens. 2020;9(1):53. https://doi.org/10.3390/pathogens9010053.
Semenova E, Severinov K. Come together: CRISPR-Cas immunity senses the quorum. Mol Cell. 2016;64(6):1013–5. https://doi.org/10.1016/j.molcel.2016.11.037.
Liao W, Liu Y, Chen C, Li J, Du F, Long D, et al. Distribution of CRISPR-Cas systems in clinical carbapenem-resistant Klebsiella pneumoniae strains in a Chinese tertiary hospital and its potential relationship with virulence. Microb Drug Resist. 2020;26(6):630–6. https://doi.org/10.1089/mdr.2019.0276.
Gholizadeh P, Aghazadeh M, Ghotaslou R, Ahangarzadeh Rezaee M, Pirzadeh T, Kose S, et al. CRISPR-cas system in the acquisition of virulence genes in dental-root canal and hospital-acquired isolates of enterococcus faecalis. Virulence. 2020;11(1):1257–67. https://doi.org/10.1080/21505594.2020.1809329.
Sampson TR, Napier BA, Schroeder MR, Louwen R, Zhao J, Chin CY, et al. A CRISPR-Cas system enhances envelope integrity mediating antibiotic resistance and inflammasome evasion. Proc Natl Acad Sci U S A. 2014;111(30):11163–8. https://doi.org/10.1073/pnas.1323025111.
Heidrich N, Hagmann A, Bauriedl S, Vogel J, Schoen C. The CRISPR/Cas system in Neisseria meningitidis affects bacterial adhesion to human nasopharyngeal epithelial cells. RNA Biol. 2019;16(4):390–6. https://doi.org/10.1080/15476286.2018.1486660.
Wu Q, Wang B, Zhou C, Lin P, Qin S, Gao P, et al. Bacterial type I CRISPR-Cas systems influence inflammasome activation in mammalian host by promoting autophagy. Immunology. 2019;158(3):240–51. https://doi.org/10.1111/imm.13108.
Shabbir MAB, Tang Y, Xu Z, Lin M, Cheng G, Dai M, et al. The involvement of the cas9 gene in virulence of campylobacter jejuni. Front Cell Infect Microbiol. 2018;8:285. https://doi.org/10.3389/fcimb.2018.00285.
Louwen R, Horst-Kreft D, de Boer AG, van der Graaf L, de Knegt G, Hamersma M, et al. A novel link between campylobacter jejuni bacteriophage defence, virulence and Guillain-Barre syndrome. Eur J Clin Microbiol Infect Dis. 2013;32(2):207–26. https://doi.org/10.1007/s10096-012-1733-4.
Gunderson FF, Cianciotto NP. The CRISPR-associated gene cas2 of legionella pneumophila is required for intracellular infection of amoebae. mBio. 2013;4(2):e00074–13. https://doi.org/10.1128/mBio.00074-13.
Beloglazova N, Brown G, Zimmerman MD, Proudfoot M, Makarova KS, Kudritska M, et al. A novel family of sequence-specific endoribonucleases associated with the clustered regularly interspaced short palindromic repeats. J Biol Chem. 2008;283(29):20361–71. https://doi.org/10.1074/jbc.M803225200.
Frimodt-Moller N. The mouse peritonitis model: present and future use. J Antimicrob Chemother. 1993;31:55–60. https://doi.org/10.1093/jac/31.suppl_d.55.
Solbiati J, Duran-Pinedo A, Godoy Rocha F, Gibson FC 3rd, Frias-Lopez J. Virulence of the pathogen Porphyromonas gingivalis is controlled by the CRISPR-Cas protein cas3. mSystems. 2020;5(5):e00852–20. https://doi.org/10.1128/mSystems.00852-20.
Koonin EV, Makarova KS. CRISPR-Cas: evolution of an RNA-based adaptive immunity system in prokaryotes. RNA Biol. 2013;10(5):679–86. https://doi.org/10.4161/rna.24022.
Toro M, Cao G, Ju W, Allard M, Barrangou R, Zhao S, et al. Association of clustered regularly interspaced short palindromic repeat (CRISPR) elements with specific serotypes and virulence potential of Shiga toxin-producing Escherichia coli. Appl Environ Microbiol. 2014;80(4):1411–20. https://doi.org/10.1128/AEM.03018-13.
Labbate M, Orata FD, Petty NK, Jayatilleke ND, King WL, Kirchberger PC, et al. A genomic island in vibrio cholerae with VPI-1 site-specific recombination characteristics contains CRISPR-Cas and type VI secretion modules. Sci Rep. 2016;6:36891. https://doi.org/10.1038/srep36891.
Louwen R, Staals RH, Endtz HP, van Baarlen P, van der Oost J. The role of CRISPR-Cas systems in virulence of pathogenic bacteria. Microbiol Mol Biol Rev. 2014;78(1):74–88. https://doi.org/10.1128/MMBR.00039-13.
Dugar G, Leenay RT, Eisenbart SK, Bischler T, Aul BU, Beisel CL, et al. CRISPR RNA-dependent binding and cleavage of endogenous RNAs by the campylobacter jejuni Cas9. Mol Cell. 2018;69(5):893–905. https://doi.org/10.1016/j.molcel.2018.01.032.
Bozic B, Repac J, Djordjevic M. Endogenous gene regulation as a predicted main function of type I-E CRISPR/Cas system in E. coli. Molecules. 2019;24(4):784. https://doi.org/10.3390/molecules24040784.
Nobrega FL, Walinga H, Dutilh BE, Brouns SJJ. Prophages are associated with extensive CRISPR-Cas auto-immunity. Nucleic Acids Res. 2020;48(21):12074–84. https://doi.org/10.1093/nar/gkaa1071.
Ratner HK, Weiss DS. CrRNA complementarity shifts endogenous CRISPR-Cas systems between transcriptional repression and DNA defense. RNA Biol. 2021;18(11):1560–73. https://doi.org/10.1080/15476286.2021.1878335.
Canez C, Selle K, Goh YJ, Barrangou R. Outcomes and characterization of chromosomal self-targeting by native CRISPR-Cas systems in Streptococcus thermophilus. FEMS Microbiol Lett. 2019;366(9):fnz105. https://doi.org/10.1093/femsle/fnz105.
Kim SK, Seong W, Han GH, Lee DH, Lee SG. CRISPR interference-guided multiplex repression of endogenous competing pathway genes for redirecting metabolic flux in Escherichia coli. Microb Cell Factories. 2017;16(1):188. https://doi.org/10.1186/s12934-017-0802-x.
Hong L, Zhang B, Duan G, Liang W, Wang Y, Chen S, et al. Role of CRISPR/Cas systems in drugresistance and virulence and the effect of IS600 on the expression of cse2 in Shigella. Wei Sheng Wu Xue Bao. 2016;56(12):1912–23. https://doi.org/10.13343/j.cnki.wsxb.20160135.
Guo R, Li Z, Zhou X, Huang C, Hu Y, Geng S, et al. Induction of arthritis in chickens by infection with novel virulent Salmonella Pullorum strains. Vet Microbiol. 2019;228:165–72. https://doi.org/10.1016/j.vetmic.2018.11.032.
Marraffini LA, Sontheimer EJ. Self versus non-self discrimination during CRISPR RNA-directed immunity. Nature. 2010;463(7280):568–71. https://doi.org/10.1038/nature08703.
Strutt SC, Torrez RM, Kaya E, Negrete OA, Doudna JA. RNA-dependent RNA targeting by CRISPR-Cas9. Elife. 2018;7:e32724. https://doi.org/10.7554/eLife.32724.
Buyukyoruk M, Wiedenheft B. Type I-F CRISPR-Cas provides protection from DNA, but not RNA phages. Cell Discov. 2019;5:54. https://doi.org/10.1038/s41421-019-0123-9.
Hoyland-Kroghsbo NM, Munoz KA, Bassler BL. Temperature, by controlling growth rate, regulates CRISPR-Cas activity in Pseudomonas aeruginosa. mBio. 2018;9(6):e02184–18. https://doi.org/10.1128/mBio.02184-18.
Yang F, Xu L, Liang L, Liang W, Li J, Lin D, et al. The involvement of Mycobacterium type III-A CRISPR-Cas system in oxidative stress. Front Microbiol. 2021;12:774492. https://doi.org/10.3389/fmicb.2021.774492.
Ibrahim S, Al-Saryi N, Al-Kadmy IMS, Aziz SN. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol Biol Rep. 2021;48(10):6987–98. https://doi.org/10.1007/s11033-021-06690-6.
Tyumentseva M, Mikhaylova Y, Prelovskaya A, Tyumentsev A, Petrova L, Fomina V, et al. Genomic and phenotypic analysis of multidrug-resistant Acinetobacter baumannii clinical isolates carrying different types of CRISPR/Cas systems. Pathogens. 2021;10(2):205. https://doi.org/10.3390/pathogens10020205.
Shabbir MA, Wu Q, Shabbir MZ, Sajid A, Ahmed S, Sattar A, et al. The CRISPR-cas system promotes antimicrobial resistance in campylobacter jejuni. Future Microbiol. 2018;13(16):1757–74. https://doi.org/10.2217/fmb-2018-0234.
Lin TL, Pan YJ, Hsieh PF, Hsu CR, Wu MC, Wang JT. Imipenem represses CRISPR-Cas interference of DNA acquisition through H-NS stimulation in Klebsiella pneumoniae. Sci Rep. 2016;6:31644. https://doi.org/10.1038/srep31644.
Shen J, Zhou J, Xu Y, Xiu Z. Prophages contribute to genome plasticity of Klebsiella pneumoniae and may involve the chromosomal integration of ARGs in CG258. Genomics. 2020;112(1):998–1010. https://doi.org/10.1016/j.ygeno.2019.06.016.
Pawluk A, Davidson AR, Maxwell KL. Anti-CRISPR: discovery, mechanism and function. Nat Rev Microbiol. 2018;16(1):12–7. https://doi.org/10.1038/nrmicro.2017.120.
Grassi L, Maisetta G, Esin S, Batoni G. Combination strategies to enhance the efficacy of antimicrobial peptides against bacterial biofilms. Front Microbiol. 2017;8:2409. https://doi.org/10.3389/fmicb.2017.02409.
Bondy-Denomy J, Pawluk A, Maxwell KL, Davidson AR. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature. 2013;493(7432):429–32. https://doi.org/10.1038/nature11723.
Bondy-Denomy J, Garcia B, Strum S, Du M, Rollins MF, Hidalgo-Reyes Y, et al. Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins. Nature. 2015;526(7571):136–9. https://doi.org/10.1038/nature15254.
Hwang S, Maxwell KL. Meet the anti-CRISPRs: widespread protein inhibitors of CRISPR-Cas systems. CRISPR J. 2019;2(1):23–30. https://doi.org/10.1089/crispr.2018.0052.
Marraffini LA, Sontheimer EJ. CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science. 2008;322(5909):1843–5. https://doi.org/10.1126/science.1165771.
Marraffini LA. Impact of CRIPSR immunity on the emergence of bacterial pathogens. Future Microbiol. 2010;5(5):693–5. https://doi.org/10.2217/fmb.10.38.
van Belkum A, Soriaga LB, LaFave MC, Akella S, Veyrieras JB, Barbu EM, et al. Phylogenetic distribution of CRISPR-Cas systems in antibiotic-resistant Pseudomonas aeruginosa. mBio. 2015;6(6):e01796–15. https://doi.org/10.1128/mBio.01796-15.
Li C, Wang Y, Gao Y, Li C, Ma B, Wang H. Antimicrobial resistance and CRISPR typing among Salmonella isolates from poultry farms in China. Front Microbiol. 2021;12:730046. https://doi.org/10.3389/fmicb.2021.730046.
Um MM, Barraud O, Kerouredan M, Gaschet M, Stalder T, Oswald E, et al. Comparison of the incidence of pathogenic and antibiotic-resistant Escherichia coli strains in adult cattle and veal calf slaughterhouse effluents highlighted different risks for public health. Water Res. 2016;88:30–8. https://doi.org/10.1016/j.watres.2015.09.029.
Stalder T, Barraud O, Casellas M, Dagot C, Ploy MC. Integron involvement in environmental spread of antibiotic resistance. Front Microbiol. 2012;3:119. https://doi.org/10.3389/fmicb.2012.00119.
Palmer KL, Gilmore MS. Multidrug-resistant enterococci lack CRISPR-cas. mBio. 2010;1(4):e00227–10. https://doi.org/10.1128/mBio.00227-10.
Mackow NA, Shen J, Adnan M, Khan AS, Fries BC, Diago-Navarro E. CRISPR-Cas influences the acquisition of antibiotic resistance in Klebsiella pneumoniae. PLoS One. 2019;14(11):e0225131. https://doi.org/10.1371/journal.pone.0225131.
Zhou Y, Tang Y, Fu P, Tian D, Yu L, Huang Y, et al. The type I-E CRISPR-Cas system influences the acquisition of blaKPC-IncF plasmid in Klebsiella pneumonia. Emerg Microbes Infect. 2020;9(1):1011–22. https://doi.org/10.1080/22221751.2020.1763209.
Gill SR, Fouts DE, Archer GL, Mongodin EF, Deboy RT, Ravel J, et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J Bacteriol. 2005;187(7):2426–38. https://doi.org/10.1128/JB.187.7.2426-2438.2005.
Ipoutcha T, Tsarmpopoulos I, Talenton V, Gaspin C, Moisan A, Walker CA, et al. Multiple origins and specific evolution of CRISPR/Cas9 systems in minimal bacteria (Mollicutes). Front Microbiol. 2019;10:2701. https://doi.org/10.3389/fmicb.2019.02701.
Price VJ, McBride SW, Hullahalli K, Chatterjee A, Duerkop BA, Palmer KL. Enterococcus faecalis CRISPR-Cas is a robust barrier to conjugative antibiotic resistance dissemination in the murine intestine. mSphere. 2019;4(4):e00464–19. https://doi.org/10.1128/mSphere.00464-19.
Martins AFM, Fontana H, Moreira BM, Bonelli RR. Draft genome sequence of a tetracycline-resistant Plesiomonas shigelloides strain isolated from aquaculture-reared tilapia. Microbiol Resour Announc. 2018;7(2):e00832–18. https://doi.org/10.1128/MRA.00832-18.
Makarova KS, Gao L, Zhang F, Koonin EV. Unexpected connections between type VI-B CRISPR-Cas systems, bacterial natural competence, ubiquitin signaling network and DNA modification through a distinct family of membrane proteins. FEMS Microbiol Lett. 2019;366(8):fnz088. https://doi.org/10.1093/femsle/fnz088.
Xu Z, Li M, Li Y, Cao H, Miao L, Xu Z, et al. Native CRISPR-Cas-mediated genome editing enables dissecting and sensitizing clinical multidrug-resistant P. aeruginosa. Cell Rep. 2019;29(6):1707–17. https://doi.org/10.1016/j.celrep.2019.10.006.
Acknowledgements
Figure 1 and Figure 3 were created with BioRender.com. Figure 2 and Figure 4 were drawn by Adobe Illustrator 2022.
Code availability in declaration
Not applicable.
Funding
This work was supported by the National Institutes of Health 5R01AI138203-01 to MW. This work was also funded by the Wang Kuancheng Medical Award Fund of Shanghai Jiao Tong University to QW and the National Key R&D Program of China (grant number. 2017YFC1600100) to MD.
Author information
Authors and Affiliations
Contributions
QW and LC performed the literature search and drafted the manuscript, QW and LC prepared the Figures, LC prepared the Table. YL and RL advised on the contents and revised the manuscript. MD, ZX, and MW revised the manuscript. MW provided overall supervision. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of interest with the contents of this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, Q., Cui, L., Liu, Y. et al. CRISPR-Cas systems target endogenous genes to impact bacterial physiology and alter mammalian immune responses. Mol Biomed 3, 22 (2022). https://doi.org/10.1186/s43556-022-00084-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43556-022-00084-1