
Abstract
Background
A useful technique for growing large amounts of plant material is in vitro propagation of important medicinal plants. The present investigation deals with the enhancement of secondary metabolite production via elicitation using gamma (γ)-radiation and phenylalanine (Phe) precursor feeding in callus cultures of Silybum marianum L.
Results
Seeds were exposed to two doses of γ-radiation (25 and 50 Gy) and the calli derived from stem explants obtained from seedlings of these radiated seeds were treated with different concentrations of Phe. The biosynthesis of phenols and flavonoids was evaluated. It was found that callus cultures derived from explants of the seeds exposed to 25 Gy γ-radiation and treated with 4 mg/l Phe accumulated the maximum phenolic content (34.27±0.02 mg/g d.wt.), while the highest flavonoid content (9.56±0.12 mg/g d.wt.) was found in callus cultures derived from explants of seeds radiated with 25 Gy γ-radiation and subjected to 1 mg/l Phe. Similarly, HPLC quantification revealed that the production of flavonoids was highly accumulated (1343.06 μg/mg d.wt.) in callus cultures from explants of seeds exposed to 25 Gy γ-radiation and grown at 1 mg/l Phe compared to the other treatments. In addition, a total of 11 important flavonoids have been determined in all callus cultures, except for acacetin-7-O-rutinoside, which was not found in the callus culture of the control.
Conclusions
These findings suggest that γ-radiation combined with Phe can improve the metabolism of S. marianum L. and could be used to produce such valuable metabolites on a commercial scale.
Similar content being viewed by others

Background
Milk thistle, Silybum marianum (L.) Gaertn., is among the most ancient of all known herbal medicines. Various preparations of the plant, especially the fruits, have been used medicinally for over 2000 years, mainly for the treatment of liver disorders [1]. Silymarin is a constitutive natural compound that accumulates in the fruits of S. marianum L. is composed of an isomeric mixture of the flavonolignans silychristin, isosilychristin, silydianin, silybin A, silybin B, isosilybin A, and isosilybin B. It is considered to be the principle pharmacologically active constituent in S. marianum L. [1,2,3]. The traditional cultivation of S. marianum L. plants has several limitations that cause a reduction in the total yield of silymarin. Plants of S. marianum L. are difficult to manually manipulate, particularly during harvesting, due to the spiny margins of leaves and flowers [4]. Thus, a possible alternative method for the large-scale production of silymarin is to use plant cell and organ cultures, where the yield depends on the constituents of the culture medium [5]. However, without elicitation, the yield of these compounds is poor [6]. Among different culture systems, callus culture may be considered an important initial biotechnological step for the large-scale production of biomass and a starting material for cell suspension cultures and shoot regeneration, being exploited for the accumulation of active compounds [7].
In vitro culture of important medicinal plants has become a reliable technique for high amounts of plant material production [8]. Production of secondary metabolites via plant cell cultures yields various advantages, including the extensive manipulation of the biosynthesis of bioactive compounds using large quantities of vegetal material under sterile and controlled conditions [9, 10] as well as higher productivity without seasonal harvesting and potential cross-contamination compared to field-grown plants [11]. Flavonoids are a class of secondary metabolites with the basic structure of two aromatic rings (A and B) linked by a heterocyclic pyrane ring (C). They are water-soluble phenolic glycosides that impart color to flowers and fruits of higher plants. They have also been linked to a reduction in the risk of cardiovascular diseases and post-climacteric osteoporosis [12]. Among these, quercetin is regarded as an active component with a variety of biological effects, such as anti-inflammatory, anti-cancer, antibacterial, antiviral, anti-gonadotropic, and anti-hepatotoxic activities [13]. In order to enhance the synthesis of secondary metabolites, several organic compounds have been fortified in the culture medium [14]. The rationale behind this approach is the good possibility that any substance that is an intermediate in or at the start of a secondary metabolite biosynthetic route would increase the yield of the final product [15]. Supplementing precursors, elicitors, and growth promoters have also been shown to increase secondary metabolite content in cell cultures [16].
Electromagnetic radiation, including gamma (γ)-rays, X-rays, visible light, and ultraviolet, is known to influence plant growth and development by inducing morphological, structural, and functional changes in cells and tissues [15]. γ-irradiation is sparsely ionizing electromagnetic radiation that can directly or indirectly affect the growth and physiology of plants. The direct effects of γ-irradiation on plants include induction of DNA mutation and chromosomal aberrations, while the increase in reactive oxygen species (ROS) and oxidative stress (OS) are considered as their indirect effects [17, 18]. For several years, γ-irradiation has been regarded as a new rapid method to change the qualitative and quantitative characteristics of plants [19, 20]. Low-dose ionizing irradiation affects cell growth, proliferation, germination rate, enzyme activity, as well as stress resistance [21]. The γ-irradiation technique is one of the most important methods used to create mutants with desired traits of agricultural significance [22]. γ-rays are highly reactive ionizing radiation that interacts with the atoms and molecules of exposed plant materials to produce free radicals, which are actively participating in various processes within the cells [23].
Flavonoids are the most ubiquitous group of natural polyphenols known for their photoprotective and antioxidative roles in the stress acclimation of plants, especially as UV filters [24]. In comparison to flavonoids generated from intact plant tissues, those derived from plant cell cultures are easier to separate in the polymeric form [25,26,27]. By regulating the in vitro conditions and feeding precursors to induce metabolite synthesis, their concentrations were markedly increased in cell cultures [28]. Phenylalanine (Phe), an upstream metabolic precursor via the phenylpropanoid pathway, is the source of flavonoids. Supplementation of Phe is expected to increase the level of the target compound [29].
Feeding culture media with phenylalanine as a precursor of the phenylpropanoid biosynthetic pathway resulted in a three-to fivefold increase in 5-methoxypodophyllotoxin in suspension culture of Linum flavum [30]. Phenylalanine enhanced the production of Taxol in suspension cell cultures of Taxus baccata [31]. In tissue cultures of Hydrocotyle bonariensis [25] and Silybum marianum L. [16, 32], the impact of Phe on flavonoids has been investigated. It has been found that 20 Gy γ-irradiation increased total phenolic and flavonoid accumulation in rosemary callus culture [17]. In addition, the dose of 10 Gy γ-irradiation increased the phenolic acid content in cinnamon [33]. Likewise, Masoud et al. [34] stated that a dose of 40 Gy increased the accumulation of total phenolic compounds in Cichorium pumilum Jacq. roots, while at 600 Gy the highest phenol content of Abelmoschus moschatus was produced [35]. The importance of phenols for plants is due to a fair correlation between antioxidant/free-radical scavenging activity and its phenolic content. Furthermore, phenol compounds may protect plants from irradiation-induced OS [34]. The main objective of this work was to enhance the production of natural medical components in S. marianum L. callus cultures using phenylalanine and γ-irradiation as elicitors.
Methods
Plant material and callus induction
The seeds of S. marianum L. were collected from its wild habitats, from Gharbia governorate at wheat field edges. The collected seeds were then treated by two doses of γ-irradiation (25 and 50 Gy). Exposure to γ-rays was carried out at the Egyptian Atomic Energy Authority with Mega Gamma-1 type J 6600 cobalt-60 as the irradiation source. The seeds (untreated, exposed to 25 Gy and 50 Gy) were washed under running tap water. They were surface sterilized by dipping in 70% ethanol for 1 min, immersed in 50% Clorox solution containing 5.4% sodium hypochlorite (NaOCl) with one drop of a commercial detergent for 20 min, and rinsed with sterilized distilled water for five times. Surface sterilization of seeds was carried out under complete aseptic conditions in the Laminar Air Flow Hood to prevent bacterial or fungal contamination of the explants. The sterilized seeds were germinated aseptically on half-strength Murashige and Skoog (MS) medium [36], supplemented with 30 g/l sucrose and 10 g/l agar. Cultures were incubated at 25 ± 2°C in the dark to facilitate germination. The clean and sterilized stem explants obtained from germinated seeds were cut into 1 cm segments and inoculated on solid MS medium supplemented with a different concentration of 2,4-dichlorophenoxy acetic acid (2,4-D) alone or in combination with 6-benzyladenine (BA) or kinetin (Kn) to induce callus formation. After 7 weeks, the successfully induced callus was separated from the explants and cultured separately until a sufficient amount of callus was produced.
Culture medium and conditions
The callus (derived from the stem explants) obtained from the three treatments was grown on MS medium supplemented with the best callus induction plant growth regulators (PGRs) combination (1 mg/l 2,4-D and 0.5 mg/l BA), treated with phenylalanine at various concentrations (1, 2, 3, and 4 mg/l) and incubated at 25 ± 2°C in the dark. After 7 weeks, the calli were separated from the medium and dried in the oven at 40°C for 12 h.
Estimation of total phenols
The total phenolic content of S. marianum L. calli was estimated quantitatively using the method described by Jindal and Singh [37]. In brief, 1 ml of the ethanolic extract was mixed with 0.1 ml of Folin reagent and 1 ml of 20% Na2CO3, then completed up to a known volume (5 ml) with distilled water. Thereafter, the absorbance was measured using the UV spectrophotometer, JENWAY (Japan) at 650 nm after 30 min. A standard curve was plotted using different concentrations of gallic acid (GA) for the determination of the total phenolic content (mg/g d.wt).
Determination of total flavonoids
The total flavonoids content of S. marianum L. calli was extracted by soaking a known weight (0.1 g) of the dried callus in 10 ml of 95% ethanol in a water bath at 60°C for 4 h. The clear supernatants were diluted to a known volume (10 ml). The aluminum chloride colorimetric method was used for total flavonoids estimation [38]. 0.5 ml of callus ethanolic extract was separately mixed with 1.5 ml of 95% ethanol, 0.1 ml of 10% aluminum chloride, 0.1 ml of 1 M potassium acetate, and 2.8 ml of distilled water. Then, it was incubated at room temperature for 30 min, after which the absorbance of the reaction mixture was measured at 415 nm using the UV spectrophotometer, JENWAY (Japan). The calibration curve was plotted using quercetin as a standard flavonoid. The amount of 10% aluminum chloride was replaced by the same amount of distilled water as the blank. Similarly, 0.5 ml of methanol extract of quercetin was used as a standard. The concentration of total flavonoids was expressed as mg/g d.wt.
Quantification analysis of flavonoids
High performance liquid chromatography (HPLC) was used for the qualitative and quantitative analysis of flavonoids in the calli of S. marianum L. The calli (from different treatments) were oven-dried and 0.1 g of the sample was used for extraction with methanol at 80°C for 4 h. The extracts were dried and re-dissolved in 1 ml of methanol [39]. The HPLC system of Shimadzu LC-10Avp plus with a PDA detector (SPD-M20A) and a C18 column (4.6 mm × 250 mm, 5 μm, Eclipse XDB C18) was used for the qualitative and quantitative analysis of total flavonoids in the methanol extracts. The mobile phase contained acetonitrile (A) and 0.1% trifluoroacetic acid (B). A gradient method was used for the separation of the extracted samples: 0–25 min, 15–40% A; 25– 40 min, 40–100% A. The elution flow rate and detection wavelength were set at 1.0 ml/min and 254 nm, respectively. The identity of peaks separated by HPLC was confirmed by the injection of flavonoid standards obtained from Sigma Aldrich, and UV spectral analysis was carried out to confirm the identification of flavonoids. The HPLC analysis was conducted in the microanalysis laboratory of the Food Technology Research Institute, Agriculture Research Center, Giza, Egypt.
Statistical analysis
The experiments were independently repeated three times under the same conditions, and the concentrations and all analyses were performed in triplicate. Results are expressed as mg/g d.wt. for flavonoids and phenols accumulated in the treated callus cultures compared to untreated calli. The graphs depicting the total flavonoids accumulation were created using Microsoft® Excel. Data were statistically analyzed using one-way ANOVA followed by Duncan’s new multiple range tests as described by Snedecor and Cohchran [40]. Means followed by the same letters are not significantly different at P ≤ 0.05.
Results
Callus induction
The seeds cultured on half strength basal MS (devoid of any PGRs) germinated normally (Fig. 1A) and developed into seedlings with roots and shoots without callus formation. Stem explants derived from 14-days-old seedlings were cultured on MS medium with different concentrations and combinations of 2.4-D and BA or Kn for callus induction (Table 1). The optimum callus induction (about 95%) was obtained at 1 mg/l 2.4-D and 0.5 mg/l BA and the first sign of callus growth from the explants was noticeable between 15-20 days after subculturing in the fresh medium. Callus induction was initiated above the surface of the explant and was friable and granulated (Fig. 1B).

Seed germination of S. marianum L. on half-strength Murashige and Skoog (MS) medium (A), calli formation from stem segments on MS medium supplemented with 1 mg/l 2,4-D and 0.5 mg/l BA (B), and calli from stem explants derived from seeds exposed to 25 Gy (C) or 50 Gy (D)
Combined effects of γ-irradiation and phenylalanine (Phe) on the accumulation of total phenols and flavonoids
Total phenols and flavonoids
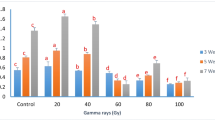
The combinations of γ-radiation (25 and 50 Gy) and different Phe concentrations were found to have astonishing effects on the biosynthesis of phenolic compounds in the callus culture of S. marianum L., where the callus cultures treated with different combinations showed higher levels of phenols and flavonoids than those of the controls. The results indicated that the highest accumulation of total flavonoids of 9.56±0.12 mg/g d.wt. was found in callus cultures subjected to 25 Gy γradiation and 1 mg/l Phe (Fig. 2 and Table 2). This was followed by callus cultures subjected to 50 Gy γ-radiation and 1 mg/l Phe (8.82±0.07 mg/g d.wt.). Likewise, the highest level of total phenolic content (34.27±0.02 mg/g d.wt.) was found in callus cultures subjected to 50 Gy γ-radiation and 4 mg/l Phe in comparison to control (12.10± mg/g d.wt.), which was significantly lower. When applied alone, it was noticed that the content of flavonoids increased more with a 25 Gy dose of γ-radiation (8.08±0.10 mg/g d.wt.) than with a 50 Gy dose (7.43±0.06 mg/g d.wt.). However, both doses (25 and 50 Gy) showed higher phenolic content (18.57 and 18.76 mg/g d.wt., respectively) compared to control (Fig. 2 and Table 2).

Total phenolic content under treatment of different combinations of γ-radiation (25 and 50 Gy) and Phe (1, 2, 3, and 4 mg/l) in the callus cultures of S. marianum L. Different letters on the bars indicate a significant difference at P ≤ 0.05
HPLC analysis of total flavonoids
A total of 11 important flavonoids have been determined in callus cultures of S. marianum L. treated with a combination of γ-irradiation (25 and 50 Gy) and 1 mg/l Phe using HPLC (Table 3 and Fig. 3). The raised aggregation of these compounds was noticed in cultures stressed with γ-radiation and Phe compared to controls (Table 3 and Fig. 4). HPLC evaluation showed that the highest accumulation of total flavonoids (1343.06 μg/mg d.wt.) occurred in callus cultures exposed to the dose of (25 Gy) and 1 ml of Phe. In contrast, the least production (763.56 μg/mg d.wt.) was found in callus cultures derived from explants of seeds exposed to 50 Gy (Table 3). Furthermore, the highest levels of kaempferol biosynthesis (37.53±0.35 and 37.63±0.38 μg/mg d.wt.) were observed in callus cultures from explants derived from seeds treated with γ-radiation at a dose of 25 and 50 Gy, respectively. Likewise, the maximum accumulation of quercetin (384.53±6.58 μg/mg d.wt.) and rutin (62.92±0.11 μg/mg d.wt.) were observed with the treatment 25 Gy and 1 mg/l Phe, whereas the highest level of apigenin (47.15±0.78 μg/mg d.wt.) and kaempferol-3-(2"-p-comaroyl) glucoside (469.30±1.02 μg/mg d.wt.) were found in the callus cultures fortified with 1 mg/l of Phe alone. Furthermore, the maximum level of quercitrin (46.43±0.32 μg/mg d.wt.) was found in callus cultures derived from stem explants from seeds exposed to γ-radiation at a dose of 25 Gy (Fig. 4).

Total flavonoid content under treatment of diffrent combinations of γ-radiation (25 and 50 Gy) and Phe (1, 2, 3, and 4 mg/l) in the callus cultures of S. marianum L. Different letters on the bars indicate a significant difference at P ≤ 0.05

Chromatograms of total flavonoids determined in callus cultures of S. marianum L. treated with different combinations of γ-radiation (25 and 50 Gy) and the amino acid precursor Phe (1 mg/l). A: control, B: 1 mg/l Phe, C: 25 Gy γ, D: 25 Gy γ + 1 mg/l Phe, E: 50 Gy γ, F: 50 Gy γ + 1 mg/l Phe
Discussion
Gamma radiation has been considered a rapid and reliable method for inducing variation in a plant’s metabolic and physiological processes [41]. Hence, γ-irradiation has gained popularity in recent decades as a novel elicitation technique for the enhanced production of secondary metabolites in plant organs and cell cultures [42, 43]. The interaction of the applied γ-rays and the free radicals in plant cells activates signal molecules involved in the defense system and synthesizes secondary metabolites [17, 42, 44, 45]. A higher level of different types of metabolites, including phenolics, terpenoids, and alkaloids, has been reported in many medicinal plants by γ-irradiation [46].
Previous studies using γ-radiation showed that it could increase the amount of secondary metabolites in callus and shoot cultures. This was observed in the callus cultures of Lithospermum erythrorhizon [47], Rosmarinus officinalis [17], Panax ginseng [45], Nothapodytes foetida [48], Stevia rebaudiana [7], Sesuvium portulacastrum [49], Rubia cordifolia [43] and shoot cultures of Mucuna pruriens [50]. Gamma radiation could increase radical scavenging activity by induction of phenolic compounds. Previously, it has been shown that phenolic content in Ferula gummosa was increased under γ-irradiation at 20 and 25 Gy, and the best dose was reported to be 20 Gy according to growth response [51].
The present study showed that γ-irradiation alone or combined with Phe increased the phenolic and flavonoid content in S. marianum L. callus cultures. Treatment with 25 and 50 Gy increased phenolic and flavonoid content, where the highest contents were observed at 25 Gy. So, 25 Gy can be used as a tool for secondary metabolite induction in S. marianum L. callus cultures. Based on the data in Table 3, it was noticed that increasing the concentration of γ-radiation resulted in a decrease in the total flavonoid content in the callus cultures of S. marianum L. The reduced levels of phenolic and flavonoid accumulation occurred mostly at higher doses of exposure. The underlying mechanistic reasons include high level of ROS generation, biological membrane disruption, photoinhibition, impairment of lipid metabolism, DNA damage, and photosystem II destruction [52, 53]. Irradiation exhibited a negative impact on plants, including the direct destruction of plastoquinone (PQ) in chloroplasts, disruption of mitochondrial activities, DNA integrity, the production of reactive oxygen species (ROS), and the formation of peroxyl radicals (ROO•) [54].
The results of the current study are in accordance with those of Azeez et al. [42], who concluded that the treatment of callus cultures with 10 Gy γ-radiation dose was effective in stimulating callus biomass production and phytochemical accumulation of Hypericum triquetrifolium Turra. However, higher doses led to a reduction of these compounds, especially with regard to irradiation. Similarly, γ-elicitation is considered a potential technique that enhances the production of secondary metabolites such as alkaloids and terpenoids in the callus of different plants, such as Eurycoma longifolia [55] and Vitis vinifera L. [56].
The outcome of the current research was supported by the study of Patil et al. [57], who declared that γ-irradiation at 5 Gy enhanced the growth of A. annua callus culture, whereas higher doses significantly reduced calli growth as compared to non-irradiated cultures. Similarly, Le et al. [45] documented that mutant adventitious root cultures of Panax ginseng exposed to 20 Gy γ-radiation showed more vigorous growth than the non-irradiated roots. Also, Heydari et al. [58] stated that low doses of γ-radiation favored callus growth in Salvia nemorosa, and a significant increase was found in the biomass growth of cultures that were subjected to 10 and 20 Gy doses. In a similar study, El-Garhy et al. [59] concluded that γ-rays (200 and 600 Gy) and colchicine (0.05%) did not inhibit the formation of S. marianum L. calli and promoted cell culture growth. Thus, callus induction from leaf segments of seedlings obtained from seeds treated with γ-rays (200 and 600 Gy), 0.05% colchicine, or control conditions, showed high biomass production. The viability of the cell suspension cultures was over 90%. The flavonolignan profile of the radiated seeds confirmed the promising impact of the two studied treatments, given that 200 and 600 Gy positively influenced the total silymarin production.
In their study on the dose-and time-dependent effects of γ-irradiation on germination rate, plant growth, chlorophyll content, oxidative stress (OS), gene expression, and antioxidant activity changes, Hong et al. [60] reported that long-term exposure to γ-rays had a significantly negative effect on the seed germination percentage. Also, they observed no differences in the final germination percentage between control and irradiated seeds. However, there was a significant difference in the germination pattern between short-term and long-term irradiation in the early germination stages. Germination-related indicators showed a tendency to decrease as the γ-radiation dose and exposure time increased. Mechanistically, the stress raised by UV light activates defense mechanisms in plants along with producing phytoalexin [61]. The production of these plant defensive compounds, including antioxidative enzymes, secondary metabolites, and cell wall modifications, enables them to cope with the oxidative damage prompted by UV radiation through scavenging lethal ROS [62].
In this study, the incorporation of Phe and γ-radiation increased the yield of total phenols and flavonoids in comparison to the control, whereas Phe alone has a low effect (nearly equals to the control). Alongside, recent approaches include combinatorial use of elicitors for more efficient procedure and increased production of biomass. Different light regimes and ultraviolet-C (UV-C) radiation have been shown to synergistically enhance the production of lignans and neolignas in Linum usitatissimum L. in vitro culture. Application of UV-C radiation and melatonin has been reported to enhance phenolic accumulation and antioxidant activities in callus cultures of Ocimum basilicum L. [63]. The impact of UV-C radiation and melatonin was also affirmed on the biosynthesis of anti-diabetic phytochemicals and antioxidants in in vitro cultures of Lepidium sativum L. by Ullah et al. [64].
Increased total phenols and flavonoids in irradiated plants were also reported by El-Beltagi et al. [17] in rosemary. γ-irradiation can induce the release of phenolic compounds from glycosidic components and degrade the larger phenolic compounds into smaller ones, which finally increases the total phenolic content [65]. Irradiation causes radiolysis of water and helps to produce free radicals such as hydroxyl radical (OH•), hydroperoxyl radical (HOO•), and hydrated electrons. These radicals may break the glycosidic bonds of procyanidin trimer, tetramer, and hexamer that are present in plants, which increases the total phenolic and flavonoid content in irradiated plants [66]. On the other hand, Abou-Zeid and Abdel-Latif [67] found that the increase in phenylalanine ammonia-lyase (PAL) concentration and activity occurred in Triticum aestivum L. seedlings with increasing doses of γ-irradiation. Plant phenylpropanoids or phenolics are synthesized by PAL, a key enzyme in the first step of the phenylpropanoid pathway.
Increased phenolic content under γ-radiation could be related to changes in the enzyme activity of the phenolic biosynthesis pathway [62]. Generally, plants receive various concentrations of radiation that trigger diverse signal transduction pathways. This induces effects on the secondary metabolism of plant species and eventually results in the accumulation of distinct levels of plant metabolites [68].
Conclusions
The results of this study proved that γ-radiation as an elicitor alone or combined with the precursor amino acid Phe could be used to improve secondary metabolite production from S. marianum L. callus cultures, which could be applied on a commercial scale to provide these medicinally important compounds.
Availability of data and materials
The data used in this study are available upon request through the corresponding author.
References
Morazzoni P, Bombardelli E (1995) Silybum marianum (Cardus marianum). Fitoterapia 66:3–42
Kim NC, Graf TN, Sparacino CM, Wani MC, Wall ME (2003) Complete isolation and characterization of silybins and isosilybins from milk thistle (Silybum marianum). Org Biomol Chem 1:1684–1689
Lee DY-W, Liu YJ (2003) Molecular structure and stereochemistry of silybin A, silybin B, Isosilybin A, and Isosilybin B, Isolated from Silybum marianum (Milk Thistle). J Nat Prod 66:1171–1174
Rady M, Saker M, Matter M (2018) In vitro culture, transformation and genetic fidelity of Milk Thistle. J Genet Eng Biotechnol 16:563–572
Akula R, Ravishankar GAJ (2011) Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal 6:1720–1731
Hasanloo T, Khavari-Nejad R, Majidi E, Ardakani M (2008) Flavonolignan production in cell suspension culture of Silybum marianum. Pharm Biol 46:876–882
Khalil SA, Ahmad N, Zamir R (2015) Gamma radiation induced variation in growth characteristics and production of bioactive compounds during callogenesis in Stevia rebaudiana (Bert). New Neg Plant Sci 1-2:1–5
Kolewe ME, Guarav V, Roberts SC (2008) Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol Pharm 5:243–256
Docimo T, Davis AJ, Luck K, Fellenberg C, Reichelt M, Phillips M, Gershenzon J, D’Auria JC (2015) Influerncer of médium and elicitors on the production od cocaine: amino acids and phytohormones by Erythroxylum coca calli. PCTOC 120:1061–1075
Dias IM, Sousa JM, Alves RC, Ferreira ICFR (2016) Exploring plant tissue culture to improve the production of phenolic compounds: a review. Ind Crop Prod 82:9–22
Murch SJ, Saxena P (2006) St. John’s worth (Hypericum perforatum L.): challenges and strategies for production of chemically consistent plants. Can J Plant Sci 86:765–771
Wang HJ, Murphy PA (1994) Isoflavone content in commercial soybean foods. J Agric Food Chem 42:1666–1673
Lamson DW, Brignale MS (2000) Antioxidants and cancer III :quercetin. Altern Med Rev 5:196–208
Namdeo AG, Jadhav TS, Rai PK, Gavali S, Mahadik KR (2007) Precursor feeding for enhanced production of secondary metabolites. Pharmacogn Rev 1(2):227–231
Rao SR, Ravishankar GA (2002) Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol Adv 20:101–153
Preetpal K, Navdeep SB (2012) Extraction of Flavonoids from In vivo and In vitro tissue culture of some important halophytes of western Rajasthan. Int J Pharm Tech Res 4(3):1167–1171
Hossam SEB, Osama KA, Wael ED (2011) Effect of low doses γ-irradiation on oxidative stress and secondary metabolites production of rosemary (Rosmarinus officinalis L.) callus culture. Radiat Phys Chem 80(9):968–976
Ahuja S, Kumar M, Kumar P, Gupta VK, Singhal RK, Yadav A, Singh B (2014) Metabolic and biochemical changes caused by gamma irradiation in plants. J Radioanal Nucl Chem 300(1):199–212
Gudkov SV, Grinberg MA, Sukhov V, Vodeneev V (2019) Effect of ionizing radiation on physiological and molecular processes in plants. J Environ Radioact 202:8–24
Charbaji T, Nabulsi I (1999) Effect of low doses of gamma irradiation on in vitro growth of grapevine. PCTOC 57(2):129–132
Sato Y, Shirasawa K, Takahashi Y, Nishimura M, Nishio T (2006) Mutant selection from progeny of gamma-ray-irradiated rice by DNA heteroduplex cleavage using Brassica petiole extract. Breed Sci 56:179–183
Chakravarty B, Sen S (2001) Enhancement of regeneration potential and variability by γ- irradiation in cultured cells of Scilla indica. Biol Plant 44(2):189–193
Majeed A, Muhammad Z, Ullah R, Ullah Z, Ullah R, Chaudhry Z, Siyar S (2017) Effect of gamma irradiation on growth and postharvest storage of vegetables. PSM Biol Res 2(1):30–35
Landry LG, Chapple CCS, Last RL (1995) Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol 109:1159–1166
Masoumian M, Arbakariya A, Syahida A, Maziah M (2011) Effect of precursors on flavonoid production by Hydrocotyle bonariensis callus tissues. Afr J Biotechnol 10:6021–6029
Ong SL, Ling APK, Poospooragi R, Moosa S (2011) Production of flavonoid compounds in cell cultures of Ficus deltoidea as influenced by medium composition. Int J Med Arom Plants 1:62–74
Bandekar H, Lele SS (2014) Production of flavonol quercetin from cultured plant cells of Banyan (Ficus benghalensis L). Int J Innov Res Sci Eng Technol 3(5):12150–12157
Lila MA, Yousef GG, Jiang Y, Weaver CM (2005) Flavonoid mixtures from cell cultures. J Nutr 135:1231–1235
Shinde AN, Malpathak N, Fulzele DP (2009) Enhanced production of phytoestrogenic isoflavones from hairy root cultures of Psoralea corylifolia L. using elicitation and precursor feeding. Biotechnol Bioprocess Eng 14:288–294
Uden WV, Pars N, Vossebeld EM, Jos NM (1990) Production of 5- metoxypodophyllotoxin in cell suspension culture of Linum flavum L. PCTOC 20:81–87
Yari-Khosroushahi A, Valizade M, Ghasempour A, Khosrowshahli M, Naghdibadi H, Dadpour MR, Omidi Y (2006) Improved Taxol production by combination of inducing factors in suspension cell culture of Taxus baccata. Cell Biol Int 30:262–269
Rahimi S, Hasanloo T, Najafi F, Nejad RAK (2011) Enhancement of silymarin accumulation using precursor feeding in Silybum marianum hairy root cultures. Plant Omics J 4:34–39
Variyar PS (1998) Effect of gamma-irradiation on the phenolic acids of some Indian spices. Int J Food Sci Technol 33(6):533–537
Masoud M, Zayed MA, Gad D, Elhaak MA (2018) Effect of gamma irradiation on some metabolites of Cichorium pumilum Jacq. Egypt J Exp Biol 14(1):153–159
Suneetha S, Priya BT, Shekar MR, Sadarunnisa S, Latha P (2018) Effect of gamma rays treatments on percentage germination, morphological variation and chlorophyll mutation in musk okra (Abelmoschus moschatus L.). Int J Curr Microbiol App Sci 6:2875–2884
Murashige T, Skoog F (1962) Revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Jindal KK, Singh RN (1975) Phenolic content in male and female Carica papaya: A possible physiological marker for sex identification of vegetable seedlings. Physiol Plant 33:104–107
Chang C, Yang M, Wen H, Chern J (2002) Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J Food Drug Anal 10:178–182
Sánchez-Sampedro MA, Fernandez-Tarago J, Corchete P (2005) Yeast extract and methyl jasmonate induced silymarin production in cell culture of Silybum marianum L. Gaerth J Biotechnol 119:60–69
Snedecor GM, Cohchran WG (1967) Statistical Methods 5th Ed. Press, Amcr Iowa, USA, Iowa State Collage
Parchin RE, Ghomi AAN, Badi HN, Eskandari A, Navabpour S, Ali M (2019) Growth characteristics and phytochemical responses of Iranian fenugreek (Trigonella foenum-graecum L.) exposed to gamma irradiation. Ind Crops Prod 139(36):111593
Azeez H, Ibrahim K, Pop R, Pamfil D, Hârţa M, Bobiș O (2017) Changes induced by gamma ray irradiation on biomass production and secondary metabolites accumulation in Hypericum triquetrifolium Turra callus cultures. Ind Crop Prod 108:183–189
Mariadoss A, Satdive R, Fulzele DP, Ramamoorthy S, Georgepriyadoss C, Zayed H, Younes S, Rajasekaran C (2020) Enhanced production of anthraquinones by gamma-irradiated cell cultures of Rubia cordifolia in a bioreactor. Ind Crop Prod 14(5):111987
Esnault M-A, Legue F, Chenal C (2010) Ionizing radiation: advances in plant response. Environ Exp Bot 68(3):231–237
Le K-C, Ho T-T, Paek K-Y, Park S-Y (2019) Low dose gamma radiation increases the biomass and ginsenoside content of callus and adventitious root cultures of wild ginseng (Panax ginseng Mayer). Ind Crop Prod 130:16–24
Vardhan PV, Shukla LI (2017) Gamma irradiation of medicinally important plants and the enhancement of secondary metabolite production. Int J Radiat Biol 93(9):967–979
Chung BY, Lee YB, Baek MH, Kim JH, Wi SG, Kim JS (2006) Effects of low dose gamma-irradiation on production of shikonin derivatives in callus cultures of Lithospermum erythrorhizon S. Radiat Phys Chem 75:1018–1023
Fulzele DP, Satdive R, Kamble S, Singh S, Singh S (2015) Improvement of anticancer drug camptothecin production by gamma irradiation on callus cultures of Nothapodytes foetida. Int J Pharm Res Allied Sci 4(1):19–27
Kapare V, Satdive R, Fulzele DP, Malpathak N (2017) Impact of gamma irradiation induced variation in cell growth and phytoecdysteroid production in Sesuvium portulacastrum. J Plant Growth Regul 36(4):919–930
Detmontree W, Sorrapetch M, Subhadhcha P, Suwimol J, Worapan S (2021) Productivity of L-DOPA in in vitro shoots of Mucuna pruriens var. utilis enhanced by gamma radiation. J App Pharm Sci 11(01):84–88
Ashouri SA, Hassanpour H, Jonoubi P, Ghorbani Nohooji M, Nadimifar MS (2016) The Effect of Gamma Irradiation on In vitro Total Phenolic Content and Antioxidant Activity of Ferula gummosa Bioss. J Med Plants 15(59):122–131
Sharma K, Li Y, Schenk PM (2014) UV-C-mediated lipid induction and settling, a step change towards economical microalgal biodiesel production. Green Chem 16:3539–3548
Waszczak C, Carmody M, Kangasjärvi J (2018) Reactive oxygen species in plant signaling. Annu Rev Plant Biol 69:209–236
Urban L, Sari DC, Orsal BP, Lopes M (2018) UV-C light and pulsed light as alternatives to chemical and biological elicitors for stimulating plant natural defenses against fungal diseases. Sci Hortic 235:452–459
Parikrama R, Esyanti RR (2014) Effect of UV Elicitation on Callus Growth, Alkaloid and Terpenoid Contents in Eurycoma longifolia Jack. Int J Adv Chem Eng Biolog Sci 1(1):12–15
Cetin ES (2014) Induction of secondary metabolite production by UV-C radiation in Vitis vinifera L. Öküzgözü callus cultures. Biological Research 47(1):37
Patil A, Suryavanshi P, Fulzele D (2018) In vitro regeneration of gamma irradiated callus of Artemisia annua and evaluation of increase artemisinin content by HPLC analysis. J Anal Pharm Res 7(5):569–573
Heydari HR, Chamani E, Esmaielpour B (2020) Cell line selection through gamma irradiation combined with multi-walled carbon nanotubes elicitation enhanced phenolic compounds accumulation in Salvia nemorosa cell culture. PCTOC 142:353–367
El-Garhy HAS, Sherif SAH, Soliman SM, Haredy AH (2021) Effect of gamma rays and colchicine on silymarin production in cell suspension cultures of Silybum marianum: A transcriptomic study of key genes involved in the biosynthetic pathway. Gene 790:145700
Hong MG, Kim DY, Jo YD, Choi H, Ahn J-W, Kwon S-J, Kim SH, Seo YW, Kim J-B (2022) Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat. Appl Sci 12:3208. https://doi.org/10.3390/app12063208
Yin X, Singer SD, Qiao H, Liu Y, Jiao C, Wang H, Li Z, Fei Z, Wang Y, Fan C (2016) Insights into the mechanisms underlying ultraviolet-C induced resveratrol metabolism in grapevine (V. amurensis Rupr.) cv.“Tonghua-3”. Front Plant Sci 7:503
Anjum S, Abbasi BH, Doussot J, Favre-Réguillon A, Hano C (2017) Effects of photoperiod regimes and ultraviolet-C radiations on biosynthesis of industrially important lignans and neolignans in cell cultures of Linum usitatissimum L. (Flax). J Photochem Photobiol B Biol 167:216–227
Nazir M, Asad Ullah M, Mumtaz S, Siddiquah A, Shah M, Drouet S, Hano C, Abbasi BH (2020) Interactive effect of melatonin and UV-C on phenylpropanoid metabolite production and antioxidant potential in callus cultures of purple basil (Ocimum basilicum L. var. s purpurascens). Molecules 25:1072
Ullah MA, Tungmunnithum D, Garros L, Drouet S, Hano C, Abbasi BH (2019) Effect of ultraviolet-C radiation and melatonin stress on biosynthesis of antioxidant and antidiabetic metabolites produced in in vitro callus cultures of Lepidium sativum L. Int J Mol Sci 20:1787
Freitas A, Moldão-Martins M, Costa HS, Albuquerque TG, Valente A, Sanches-Silva A (2015) Effect of UV-C radiation on bioactive compounds of pineapple (Ananas comosus L. Merr.) by-products. J Sci Food Agric 95:44–52
Li D, Luo Z, Mou W, Wang Y, Ying T, Mao L (2014) ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol Technol 90:56–62
Abou-Zeid HM, Abdel-Latif SA (2014) Effects of gamma irradiation on biochemical and antioxidant defense system in wheat (Triticum aestivum L.) seedlings. Int J Adv Res 2(8):287–300
Matsuura HN, de Costa F, Yendo ACA, Fett-Neto AG (2013) Biotechnology for Medicinal Plants. In: Chandra S, Lata H, Varma A (eds) Photoelicitation of bioactive secondary metabolites by ultraviolet radiation: mechanisms, strategies, and applications. Springer-Verlag, Berlin Heidelberg, pp 171–190
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
RG and AK conceived and designed the research. AK collected the data. EA-E and RA-K performed data analyses. RG and AK participated in data analysis and wrote the manuscript. RG and RA-K revised the final version of manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Khalifa, A.M., Abd-ElShafy, E., Abu-Khudir, R. et al. Influence of gamma radiation and phenylalanine on secondary metabolites in callus cultures of milk thistle (Silybum marianum L.). J Genet Eng Biotechnol 20, 166 (2022). https://doi.org/10.1186/s43141-022-00424-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-022-00424-2