
Abstract
Background
Penicillin G amidase/acylases from microbial sources is a unique enzyme that belongs to the N-terminal nucleophilic hydrolase structural superfamily. It catalyzes the selective hydrolysis of side chain amide/acyl bond of penicillins and cephalosporins whereas the labile amide/acyl bond in the β-lactam ring remains intact.
Main body of abstract
This review summarizes the production aspects of PGA from various microbial sources at optimized conditions. The minimal yield from wild strains has been extensively improved using varying strain improvement techniques like recombination and mutagenesis; further applied for the subsequent synthesis of 6-aminopenicillanic acid, which is an intermediate molecule for synthesis of a wide range of novel β-lactam antibiotics. Immobilization of PGA has also been attempted to enhance the durability of enzyme for the industrial purposes.
Short conclusion
The present review provides an emphasis on exploitation of E. coli to enhance the microbial production of PGA. The latest achievements in the production of recombinant enzymes have also been discussed. Besides E. coli, other potent microbial strains with PGA activity must be explored to enhance the yields.
Graphical abstract

Similar content being viewed by others

Background
Penicillin G amidases or acylases (PGA) (penicillin amidohydrolase: EC 3.5.1.11) is one of the principal enzymes at an industrial level used to catalyze the enzymatic hydrolysis of various penicillins by cleaving their amide bond to yield 6-APA and its corresponding organic acid [1]. Penicillin G (PG, also known as benzyl penicillin), penicillin V (PV, also known as phenoxymethylpenicillins), and other bulk penicillins are the substrates which are chemically or enzymatically transformed for synthesis of wide range of novel β-lactam antibiotics such as amoxicillin, ampicillin, and cephalosporins [2]. Due to several disadvantages of chemical synthesis, currently much preference is given to enzymatic hydrolysis than the chemical methods as it is more specific, cheaper, and occurs at milder conditions [3]. The conversion of penicillin G to 6-aminopenicillanic acid (6-APA) and phenyl acetic acid (PAA) occurs via hydrolysis of the amide bond in its side chain catalyzed by PGA under slight alkaline pH resulting in the transfer of phenyl acetyl moiety from 6-APA to water [4] (Fig. 1). PGA is the second most commercially used enzyme worldwide, followed by glucose isomerase [5] and penicillin G was the first β-lactam antibiotic discovered in 1940s which is primarily active against gram-positive bacteria [4]. PGA is widely distributed among microbes like bacteria, fungi, yeasts, and actinomycetes [6], which are often cultivated at temperatures lower than 30 °C [2]. 6-APA further facilitates the route to synthesize a variety of semisynthetic antibiotics with improved antibacterial and pharmacological characteristics [7]. Due to the enormous use of antibiotics, pathogens have developed resistance against them with time. Thus, to overcome this concern, the only method is to synthesize new semisynthetic antibiotics [8]. The recent efforts for the overproduction of PGA are achieved through genetic engineering by constructing novel recombinant host/vector systems [7]. Besides this, the fermentation operations are run by quantitatively analyzing the kinetics to achieve optimum conditions for PGA production.

Hydrolysis of penicillin G by PGA to synthesize 6-APA
To make the process economic and commercial at industrial scale, PGA is immobilized on a matrix by various means like membrane or fiber entrapment, adsorption, and covalent binding to allow re-utilization as well as enhancing the enzymatic conversion of penicillins under acidic or alkaline conditions [9, 10]. The first executed hydrolysis by immobilized PGA involved penicillin G and cephalosporin G as substrates [11]. This method was quickly implemented to be used by the pharmaceutical industry since the mild conditions were appropriate for the stability and activity of PGA [12]. E. coli PGAs generally have a heterodimeric structure as in E. coli ATCC 11105 PGA which is synthesized as a single cytoplasmic precursor with 23-kDa α-subunit and 65-kDa β-subunit [13], maybe due to the unusual protein formation mechanism [14]. The mature E. coli PGA is located in the periplasm [2] which allows a rather simple extraction procedure as only cell permeabilization is required in such case [15]. PGAs belong to the N-terminal nucleophilic hydrolase structural superfamily with a catalytic nucleophile (Ser, Cys, or Thr) as the active site for cleaving an amide bond at the N-terminus [16]. PGAs are potentially useful biocatalysts in various ways such as protection of amino and hydroxyl groups during peptide synthesis and resolution of racemic mixtures of chiral compounds [17, 18]. They are also known to work as a promising candidate for linker cleavage studies of structures for combinatorial chemistry due to their high substrate specificity concerning the acyl residue [19].
Main text
Discovery of penicillin G amidase
Enzymes are the ideal biocatalysts which provide eco-friendly environmental conditions for different processes to occur mildly, thus of great importance to the industries [5]. PGA was firstly reported in the mycelium of Penicillium chrysogenum Q176 in the year 1950 as an enzyme with potential to catalyze the hydrolysis of penicillin G into PAA and an unknown compound called “penicillin” (6-APA) in the form of hygroscopic crystalline needles giving 158–159 °C melting point [20, 21]. There are various microbial strains of bacteria, fungi, yeasts, and actinomycetes like Escherichia coli, Pseudomonas melanogenum, Bacillus megaterium, Streptomyces lavendulae, Achromobacter sp., Bovista plumbea, Kluyvera sp., Penicillium chrysogenum, Actinoplanes sp., Proteus rettgeri, and Mucor griseocyanus [3] available which produce PGA, i.e., exhibit PGA activity either in their natural wild types or after recombinant processing. But the most efficient and highly preferred enzymes are derived from E. coli and Bacillus megaterium strains [22]. In 1960, the hydrolysis of penicillin G using PGA from E. coli was reported by Bayer and Beecham while Kaufmann and Bauer reported the hydrolysis of penicillin to 6-APA [7]. Due to the ability of E. coli PGA to hydrolyze a wide variety of phenyl acetyl substituted compounds, it has been widely used in various formulations [23]. The production kinetics of some PGA producing microorganisms has been presented in the Tables 1 and 2.
Exploitation of E. coli for penicillin G amidase production
E. coli has been exploited in every way possible, but still it shows to be the best among others in terms of PGA production. The microbial production of PGA, either in its native strain (Table 1) or recombinant hosts (Table 2), has been extensively studied, especially for PGA from E. coli (EcPGA) [2]. PGA is an intracellular enzyme and its production is achieved by growing distinct strains in standard mediums containing carbon, nitrogen sources along with an inducer (mostly, PAA) in appropriate concentrations. The study of substrates, nutrient composition, type of cultivation, bioreactors, etc., plays a key role in optimizing conditions for a biocatalyst to grow and minimize the production of toxic substances. Various purification schemes have been applied in order to get a partially purified or purified PGA enzyme, like ammonium sulfate precipitation followed by desalting which in turn is used as a biocatalyst to produce 6-APA after hydrolyzing penicillins for synthesizing antibiotics [48]. Partially purified PGA can be immobilized for industrial applications without additional chromatographic purification [2]. Chemical deacylation can also be performed for 6-APA production, but it is non-specific, environmentally hazardous, and expensive; thus, it is not preferred much [48]. The designing of industrial plant for large scale production must be done using improvised engineering approaches with highly robust techniques like strain manipulation, cultivation methods, and downstream processing [49]. The overexpression in recombinant E. coli hosts for the industrial usage can be succeeded by optimization of the operational strategies [50].
Production of recombinant strains and PGA purification
The vigorous growth characteristics of E. coli along with its conventional metabolism and physiology make it a simplistic host system. The EcPGA serves as an excellent enzyme for industrial uses but the promoter of its endogenous pac gene is weak and, thus, unsuitable for large scale production. Hence, its manipulation can be done to produce a recombinant PGA (Fig. 2) by overexpressing the native gene in high-copy episomal plasmids to build up gene dosage so as to boost all expression steps under regulation of a strong promoter-operator system, like transcription, translation, translocation, periplasmic processing, and folding. The augmentation of translation can be done by increasing the stability of pac mRNA, modifying the region of ribosome binding site, etc., leading to a mature PGA giving higher levels of activity [15, 51]. The pac genes from bacterial strains other than E. coli have also been heterologously expressed in E. coli like genes from Arthrobacter viscosus [52], B. megaterium [53], Achromobacter xylosoxidans [54], P. rettgeri [55], A. faecalis [56], K. cryocrescens [57], and Thermus thermophilus [43]. Their PGAs can surpass EcPGA when expressed in E. coli in terms of particular enzymatic properties like wide operation range, molecular stability, and environmental tolerance [2]. An overproduction of mature PGA has been performed by cloning pac gene from E. coli ATCC 11105 into pUC 9 rather than pBR 322. The emerged vector (E. coli pUPA-9) when transformed into E. coli 5 K resulted in production of 10 mg PGA/L of cells due to high gene copy number. Hydrophobic interaction chromatography and anion exchange was used to purify the enzyme from periplasmic fraction of E. coli pUPA-9 [58]. The successful expression of pac genes from E. coli and Providencia rettgeri in Saccharomyces cerevisiae revealed that contrary to bacterial hosts, where PGA is retained in the periplasm; the yeast cells secrete enzyme directly into the medium, and E. coli pac is poorly expressed in yeast [33]. A pac gene from parental E. coli ATCC11105 was cloned and expressed in recombinant E. coli χ6212/pRT4 which produced 1000 units PGA/g of cell dry weight that is 23-folds more than the parent strain (43 units PGA/g of cell dry weight). The replacement of original pac promoter by strong ptrc promoter of the vector pYA292 led to this increase in the yield. Later, 16 units/mg of protein was purified near to homogeneity by hydrophobic interaction chromatography giving 60% recovery of PGA by two-step of purification [34].

Strategies adopted by researchers for the production of PGA
However, to enhance PGA production in different recombinant E. coli expression hosts and identify the reason behind decrease in post translational yield, PGA from E. coli ATCC 11105 was used as a model recombinant protein. It was observed that the main processes limiting enzyme production were intracellular proteolytic degradation of the newly synthesized PGA precursor by reduction of the growth rate of cells, protective fusions, and translocation through the plasma membrane. Therefore, to elevate PGA expression at molecular level in recombinant E. coli strains, intracellular proteolysis and translocation were manipulated, and an appropriate host system with suitable cultivation medium was selected. The cultivation of an E. coli strain BL21 (DE3), which is a natural deficient of ATP-dependent proteinase, Lon and outer membrane proteinase, OmpT [59], was done in a medium without a proteinaceous substrate that provided an increased PGA yield by 10-folds [38]. It has been studied earlier that PGA yields can be improved by the use of minimal medium with different carbon sources since it is sensitive to complex proteinaceous substrates [60]. By employing the rhamnose inducible expression system in E. coli, production of recombinant PGA from A. faecalis ATCC19018 was carried out. The rhamnose inducible promoter, also serving as a carbon source, was used to control the cloning of desired gene into a multi copy vector. A PGA activity of 4500 U/L was achieved with 0.96 U/mg of specific activity in the cell free extract [19]. Production of PGA from Kluyvera citrophila was optimized in E. coli BL21 (DE3), and effects of physical parameters such as temperature and pH were investigated. The cells reached their stationary phase after 24 h of growth, and yield was increased by more than 2.4-fold in TB medium. Culture performance was studied, and 9600 U/L activity and 24.4 cell density were achieved [39]. The use of cheese whey as an inducer and carbon source has been done to produce PGA by recombinant E. coli W3110/pPA102. A specific activity of 781 U/g was attained at use of 5 g/L of cheese whey and 3% dissolved oxygen which shows that it can be successfully used as an inducer and carbon source for PGA production using constructions driven by the lac promoter [40]. E. coli HB101 and JM109 are among the host systems which perform well for the production of PGA [61]. A pac gene with its promoter was obtained by using SSP-PCR and direct genome sequencing from Bacillus badius and expressed in different E. coli hosts. A recombinant PGA was obtained in E. coli DH5α with maximum 1820 U/L activity. Ni-NTA chromatography was used to purify the PGA which showed stability over a wide pH range of 6.0–8.5 with a maximum activity at pH 7.0 and activity on a wide β-lactam substrate range. The kcat/Km values revealed that enzyme had strongest preference for penicillin G (1025.0 mM/s) followed by cephalothin, cephalexin, and ampicillin [62]. Screening for PGA producing E. coli isolates was carried out to study cloning and recombinant expression. A positive clone gene was cloned in plasmid pGEM-T easy vector, and BL21 host cells were explored for high PGA levels. The comparative study of wild and recombinant strain revealed that in E. coli BL21, inducer IPTG (1 mM) can increase the level of PGA to 150 U/g (wet weight) of recombinant bacteria, i.e., three times more than PGA activity of wild type E. coli strain [63]. A gene homologous to pac from E. coli was isolated from the outer side of cytoplasmic membrane of Thermus thermophilus (Tth) HB27. Its overexpression was difficult in mesophilic host due to complex maturation and nature. Thus, by using chaperone co-expression and calcium supplementation of the culture medium, its PGA was efficiently overexpressed in E. coli. Further, affinity chromatography was performed to obtain recombinant PGA by conformation through SDS-PAGE and MALDI-TOF analysis. It was determined for activity and hydrophobic acyl-chain penicillins were preferred as substrates with more specificity towards penicillin K possess highest specificity constant value (16.12 mM/s) while penicillin G had approx. 1.10−4 mM/s [43]. But this strain can be engineered by mutagenesis of selected active site residues to hydrolyze penicillin G to a definite extent [23]. For efficacious production, PGA was overexpressed in an engineered E. coli and released in medium with low conductivity to allow direct application of the extracellular fraction to the anion-exchange chromatography column. Further, harvested cells were used for purification of enzyme by using strong anion-exchange (Q) column yielding PGA activity of 16.3 U/mg at 871 U/g DCW, i.e., up to 3 folds [44]. Strong cation-exchange membrane adsorbers were used in a one-step purification process to get high productivity of bounded PGA (98%) with a residual enzyme activity of 80–85%. E. coli 5KpHM12 real cell lysate was processed under optimal conditions, and PGA was isolated with a purification factor of 101.3 (4.97 U/mg) [64]. Based on the TFF-AMEC, a single-step downstream process has been reported to purify E. coli PGA with high yield [49].
Mutagenesis for strain improvement
Mutagenesis is another powerful tool used for strain improvement (Scheme 1) for the production of PGA. Different mutants have been produced by treating E. coli with acridine orange yielding high levels of PGA and inactivated β-lactamase [65]. A four times more productive mutant than the parent strain has been produced by chemical mutagenesis of E. coli ATCC 11105 with NTG [66]. UV radiations have also been known to improve the catalytic efficiency and substrate specificity of PGA producing bacterial strains [67]. Locally isolated E. coli strains were mutated using UV radiation by exposing diluted cultures to UV lamps at varying time and distances. BDCS-N-M74 was the hyper-producing mutant exhibiting 3 fold (22.5 mg of 6-APA/h/mg wet cells) increases in PGA activity as compared to that in the parent strain (6.7 mg of 6-APA/h/mg wet cells) with limited expression of β-lactamase. Due to these mutations, the efficiency for microbes to produce enzymes/metabolites increases whereas higher expose can cause lethal changes in the microbe [41]. ep-PCR is a popular method to upgrade and improvise the enzyme properties by increasing frequency of mismatched incorporation of nucleotides into newly synthesized PCR products [68]. ep-PCR coupled with high-throughput screening assay was applied to E. coli pac gene to improve enzyme activity of its PGA. The strain pUC19-pacwt11 with 0.24 U/ml of activity was used as a template for the construction of mutants by ep-PCR. The best mutant PA M2234 had a specific activity 4.0 times higher than that of enzyme from wild strain and also displayed higher stability at pH 10. DEAE-Sepharose column was used for purification providing 33-fold with 1.34 U/mg specific activity for wild strain and 43-fold with 5.51 U/mg specific activity for mutant [17].

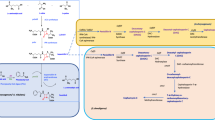
Schematic representation of overall production of PGA, yield improvement strategy and synthesis of 6-APA for the synthesis of β-lactam mediated antibiotics
Immobilization of penicillin G amidase
The commercialization of an enzyme is dependent upon its durability which directly supports product formation by repeating enzyme-reaction cycles. To reduce the process cost of any molecule at industrial scale, the most important factors are an improved log life in growth curve, enzyme stability, and reusability. For all such purposes, immobilization process uses an inert matrix for surface attachment of enzymes, even at extreme of pH and temperature at exceptionally high substrate concentrations in different solvents [10, 69]. Therefore, enzyme immobilization has gained tremendous attraction among scientific communities. Immobilization of PGA has played an important role to make its processing economically feasible at an industrial level [48]. It is due to the successful development of immobilized PGA that replaced the chemical route for 6-APA production [70]. There are several methods such as adsorption, micro-encapsulation, fiber entrapment, copolymerization, cross-linking, and covalent attachment used for immobilization of PGA to increase its stability, facilitate its recovery, and re-use in many hydrolysis cycles. Different methods have been reviewed by Parmar and co-workers [48]. Silica support, glycoxyl agarose, alumina beads, nylon fibers, ethylene glycol dimethacrylate, zerogel, Eupergit C, and sepa beads are some carriers for liquid PGA immobilization [71]. Another methodology known as CLEA is a combination of immobilization as well as purification without much highly purified enzyme. It performs direct immobilization of an enzyme from crude fermentation broth and the first examples of CLEAs were derived from PGA. Due to the limited thermal stability and a low tolerance of free enzymes to organic solvents, PGA stabilization by immobilization becomes an ideal solution for industries [72]. Varying range of other methods like binding, enzyme crystal or powder packaging, and prefabricated carrier materials have been used to immobilize PGAs from microbes such as Bacillus megaterium, A. faecalis, and E. coli [73]. The cells of E. coli and A. faecalis have also been permeabilized using gelatin, polyvinyl alcohol, and agar matrices giving satisfactory results [15, 55]. Some immobilized preparations of E. coli whole cells and its mutants with PGA activity have been made by entrapping in gluten matrix, open pore gelatin matrix [74], and polymethacrylamide beads [75] proving to be effective in penicillin G hydrolysis. For the hydrolysis as well as synthesis of β-lactam antibiotics, cross-linked enzyme crystals of EcPGA featuring characters of pure enzyme and high organic solvent tolerance are evident to work well [76]. To minimize the decrease in catalytic activity in presence of high concentrations of organic co-solvents, an artificial microenvironment generation around the immobilized enzyme can also be done [77]. Under non-denaturing conditions, CLEAs of EcPGA have been prepared by its physical aggregation and cross-linking with glutaraldehyde [78]. For industrial application of PGA, its covalent binding to commercial epoxy-activated acrylic beads (Eupergit C) has ensured refinement in operational stability [79]. But PGA immobilization on Sepabeads-EP is more stable as compared to Eupergit C [80]. The fabrication of macroporous weak cation-exchange methacrylate polymers was done by Wang et al. [81] and Chen et al. [82] to immobilize PGA using hydrophobic interaction chromatography for concentration and purification of enzyme. Cheng and co-workers [55] used pore matrix cross-linking with glutaraldehyde to immobilize the permeabilized whole-cell PGA from A. faecalis enhancing PGA activity by 7.5-folds and yielding 75% 6-APA by bioconversion of penicillin G. By suspension polymerization, magnetic hydroxyl particles have been activated with epoxyl chloropropane and used for PGA immobilization giving constant activity at ∼94% for up to 80 cycles [11, 81]. A covalent immobilized PGA on glutaraldehyde activated NH2-PVC membranes has been used for 6-APA production giving high catalytic activity up to 4000 μmol min m2 and retaining 45% of activity. The immobilized PGA had KM value (125.8 mM) 23 times higher than that of the free enzyme (5.4 mM) [83]. The immobilization of PGA on magnetic Fe2O3/Fe3O4@SiO2-CHO nanocomposites via the Schiff's reaction also showed excellent pH stability, thermal stability, and reusability as compared to free enzyme. Sixty-seven percent of the initial activity was retained even after 12 cycles of enzyme usage [84]. NIPAM (N-isopropylacrylamide) with active ester groups is a thermo-responsive, biocompatible polymer which has also been used for immobilization of PGA and the resulting enzyme–polymer conjugate possessed a close hydrolytic activity to that of the free enzyme. Thus, it is suitable for synthesis of the semi-synthetic cephalosporin and cephalexin which is also an important β-lactam antibiotic, by the reaction of D-phenylglycine amide with 7-ADCA (aminodeacetoxycephalosporanic acid) [85].
Application of 6-aminopenicillanic acid for the production of β-lactam antibiotics
β-lactam antibiotics are one of the common antibiotics having broad spectrum activity against several Gram-negative and Gram-positive pathogens. For treating various bacterial infections, they have been successfully used as a drug over the past few decades across the globe [86]. For the synthesis of β-lactam antibiotics and their derivatives, 6-APA is an important skeleton molecule (Scheme 1). For the first time in 1959, Batchelor and co-workers discovered 6-APA from fermentation of penicillin. This was an important breakthrough for the discovery of novel semi-synthetic β-lactam antibiotics [87]. Further, scientists attempted for a semi-synthetic route to synthesize other novel antibiotics. The addition of different side chains to 6-APA were also done to generate novel antibiotics with different activity and better pharmacokinetics which served to provide resistance from β-lactamases [88, 89]. The core structure of these antibiotics is 3-carbon and 1-nitrogen ring known as β-lactam ring which is highly reactive. PBP are the enzymes responsible for cross-linkage of peptidoglycan components present in the bacterial cell wall [90]. The mechanism behind the antibiotic effect possessed by β-lactams is that they mimic the structure of natural D-Ala-D-Ala substrate of PBP and inhibit its work. This is due to similarity in the positions of CO-N bond in the β-lactam ring of the penicillin and the CO-N bond in D-alanyl-D-alanine which is the target of transpeptidation. Thus, penicillin binds to the site proposed for D-alanyl-D-alanine and halts the cell wall synthesis causing cell lysis [91, 92], although, due to the increased resistance shown by several bacterial strains towards these antibiotics, their work has been afflicted. These strains show resistance to β-lactams by producing an enzyme known as β-lactamases which cleaves the 4-membered β-lactam ring [93]; therefore, inactivating the drug leading to its inefficiency to bind to the target PBPs. If the antibiotic is unable to bind to the target PBP, it would not be able to hinder in the cell wall synthesis causing no harm to the bacteria. It has been revealed by sequence analysis that PBPs and β-lactamases come from a common ancestor. The first bacterial strain to exhibit resistance to some antibacterial agents like penicillin G, erythromycin, streptomycin, and tetracycline was Staphylococcus aureus as observed in the year 1947. Gram-positive microbes (S. epidermidis, S. aureus, S. pseudintermedius, etc.) produce β-lactamases into their external surroundings as exoenzymes, while in gram-negative microbes (Klebsiella, Pasteurella, Escherichia, Haemophilus, Salmonella, Pseudomonas sp., etc.), it is kept back in their periplasmic space [91]. It has been evident that antibiotic-resistance genes could also pass from bacteria to other strains or species to procure additional gene combinations. Methicillin/oxacillin-resistant Staphylococcus aureus (MRSA), penicillin-resistant Streptococcus pneumonia (PRSP), and vancomycin-resistant Enterococci (VRE) are some multiple drug resistant organisms. For the bacterial resistance, medicinal chemistry approach has been applied for the discovery of novel antibiotics [94, 95]. Several natural antibiotic scaffolds were chemically modified to produce the new antibiotics with higher pharmacological activity. The main motive of semi-synthetic synthesis is to resolve the bacterial resistance issue, improve the spectrum, improve the oral absorption, and enhance pharmacokinetics [87, 96].
In 2018, the market size of β-lactams antibiotics and β-lactamase inhibitors was estimated at about $27,126 million and is expected to reach about $34,170 million in 2028 [97]. Market size of the PGA is also increasing with the increasing interest of β-lactam antibiotics in the market. At the international market, few pharmaceutical industries are playing an important role in the production of PGA such as Aumgene Biosciences, Fermenta Biotech Limited, Amicogen, Incorporation, Crawford Wisdom International, LGM Pharma, Hangzhou Biodoor Biotechnology Co. Ltd., Taizhou Doyin, etc., and many others. Due to the increment in drug resistance by pathogens, an emergency for the production of novel antibiotics has been created. These new bacterial infections with high drug resistance are an emerging concern threatening public health and here comes the need of novel antibiotics across the world [92]. Therefore, biochemists are paying more attention in semi-synthesis or synthesizing derivatives of natural drugs [76, 98]. Even though the currently available antibiotics will soon find new drug resistant strains, the present unavailability needs to be satisfied first. Studies on the nature of resistance and other molecular details propose a desire for producing novel agents to overcome bacterial resistance [99].
Conclusion
PGA-mediated transformation is a green process for the synthesis of semisynthetic β-lactam antibiotics which has significant industrial importance. Gene manipulation by recombinant technology, mutagenesis, and immobilization techniques have played an extraordinary role in commercialization of this technology at industrial scale by contribution towards enhancing its properties providing stable and high yield performance strain for the production of 6-APA. Among all the strains, E. coli is the only strain which is extensively exploited for the production of PGA whereas biotechnological application like genetic engineering has played a significant role to overcome the yield limitations of native genes in E. coli. Other potent microbial strains with PGA activity must also be worked upon to enhance the yields. Since the bacterial strains have been known to show resistance against the previously used antibiotics, there is a great need for development of novel β-lactam antibiotics soon.
Availability of data and materials
NA
Abbreviations
- 6-APA:
-
6-Aminopenicillanic acid
- CLEA:
-
Cross-linked enzyme aggregates
- ep-PCR:
-
Error-prone polymerase chain reaction
- IPTG:
-
Isopropyl ß-D-1-thiogalactopyranoside
- MALDI-TOF:
-
Matrix-assisted laser desorption/ionization-time of flight
- NTG:
-
N-methyl-N-nitro-N-nitrosoguanidine
- PAA:
-
Phenyl acetic acid
- PBP:
-
Penicillin-binding proteins
- PGA:
-
Penicillin G amidase/acylases
- SDS-PAGE:
-
Sodium dodecyl sulfate polyacrylamide gel electrophoresis
- SSP-PCR:
-
Single specific primer polymerase chain reaction
- TFF-AMEC:
-
Tangential flow filtration anion-exchange membrane chromatography
- UV:
-
Ultraviolet
References
Wu Z, Liu C, Zhang Z, Zheng R, Zheng Y (2020) Amidase as a versatile tool in amide-bond cleavage: from molecular features to biotechnological applications. Biotechnol Adv 43:107574
Srirangan K, Orr V, Akawi L, Westbrook A, Moo-Young M, Chou CP (2013) Biotechnological advances on penicillin G acylase: pharmaceutical implications, unique expression mechanism and production strategies. Biotechnol Adv 31(8):1319–1332
Martínez-Hernández JL, Mata-Gómez MA, Aguilar-González CN, Ilyina A (2010) A process to produce penicillin G acylase by surface-adhesion fermentation using Mucor griseocyanus to obtain 6-aminopenicillanic acid by penicillin G hydrolysis. Appl Biochem Biotechnol 160(7):2045–2053
Hassan ME (2016) Production, immobilization and industrial uses of penicillin G acylase. Int J Curr Res Rev 8(15):11–22
Silva RG, Souza VR, Nucci ER, Pinotti LM, Cruz AJ, Giordano RC, Giordano RL (2006) Using a medium of free amino acids to produce penicillin G acylase in fed-batch cultivations of Bacillus megaterium ATCC 14945. Braz J Chem Eng 23(1):37–43
Verhaert RM, Riemens AM, Van der Laan JM, Van Duin J, Quax WJ (1997) Molecular cloning and analysis of the gene encoding the thermostable penicillin G acylase from Alcaligenes faecalis. Appl Environ Microbiol 63(9):3412–3418
Buchholz K (2016) A breakthrough in enzyme technology to fight penicillin resistance—industrial application of penicillin amidase. Appl Microbiol Biotechnol 100(9):3825–3839
Cobos-Puc L, Rodríguez-Herrera R, Cano-Cabrera JC, Aguayo-Morales H, Silva-Belmares SY, Gallegos AC, Hernández JL (2020) Classical and new pharmaceutical uses of bacterial penicillin G acylase. Curr Pharm Biotechnol 21(4):287–297
Souza VR, Silva AC, Pinotti LM, Araújo HS, Giordano RD (2005) Characterization of the penicillin G acylase from Bacillus megaterium ATCC 14945. Braz Arch Biol Technol 48:105–111
Li K, Mohammed MA, Zhou Y, Tu H, Zhang J, Liu C, Chen Z, Burns R, Hu D, Ruso JM, Tang Z (2020) Recent progress in the development of immobilized penicillin G acylase for chemical and industrial applications: a mini-review. Polym Adv Technol 31(3):368–388
Chandel AK, Rao LV, Narasu ML, Singh OV (2008) The realm of penicillin G acylase in β-lactam antibiotics. Enzym Microb Technol 42(3):199–207
Volpato G, Rodrigues RC, Fernandez-Lafuente R (2010) Use of enzymes in the production of semi-synthetic penicillins and cephalosporins: drawbacks and perspectives. Curr Med Chem 17(32):3855–3873
Bruns W, Hoppe J, Tsai HS, Brüning HJ, Maywald F, Collins J, Mayer H (1985) Structure of the penicillin acylase gene from Escherichia coli: a periplasmic enzyme that undergoes multiple proteolytic processing. J Mol Appl Genet 3(1):36–44
Erarslan A, Terzi I, Güray A, Bermek E (1991) Purification and kinetics of penicillin G acylase from a mutant strain of Escherichia coli ATCC 11105. J Chem Technol Biotechnol 51(1):27–40
Illanes A, Valencia P (2017) Industrial and therapeutic enzymes: penicillin acylase. In: Current Developments in Biotechnology and Bioengineering. Elsevier, pp 267–305
McVey CE, Walsh MA, Dodson GG, Wilson KS, Brannigan JA (2001) Crystal structures of penicillin acylase enzyme-substrate complexes: structural insights into the catalytic mechanism. J Mol Biol 313(1):139–150
Balci H, Ozturk MT, Pijning T, Ozturk SI, Gumusel F (2014) Improved activity and pH stability of E. coli ATCC 11105 penicillin acylase by error-prone PCR. Appl Microbiol Biotechnol 98(10):4467–4477
Shewale JG, Sivaraman H (1989) Penicillin acylase: enzyme production and its application in the manufacture of 6-APA. Process Biochem 24(4):146–154
Deak PM, Lutz-Wahl S, Bothe H, Fischer L (2003) Bioreactor cultivation of Escherichia coli for production of recombinant penicillin G amidase from Alcaligenes faecalis. Biotechnol Lett 25(5):397–400
Sakaguchi K, Murao S (1950) A preliminary report on a new enzyme, “penicillin-amidase”. Nippon Nōgeikagaku Kaishi 23(9):411
Chisti Y, Moo-Young M (1991) Fermentation technology, bioprocessing, scale-up and manufacture. Biotechnology: the science and the business 167-209.
Matsumoto K (1993) Production of 6-APA, 7-ACA and 7-ADCA by immobilized penicillin and cephalosporin amidases. Industrial application of immobilized biocatalysts 67-88.
Avinash VS, Pundle AV, Ramasamy S, Suresh CG (2016) Penicillin acylases revisited: importance beyond their industrial utility. Crit Rev Biotechnol 36(2):303–316
Self DA, Kay G, Lilly MD, Dunnill P (1969) The conversion of benzyl penicillin to 6-aminopenieillanie acid using an insoluble derivative of penieillin amidase. Biotechnol Bioeng 11(3):337–348
Robas N, Zouheiry H, Branlant G, Branlant C (1993) Improved penicillin amidase production using a genetically engineered mutant of Escherichia coli ATCC 11105. Biotechnol Bioeng 41(1):14–24
Rajendhran J, Krishnakumar V, Gunasekaran P (2002) Optimization of a fermentation medium for the production of penicillin G acylase from Bacillus sp. Lett Appl Microbiol 35(6):523–527
José L, Martinez H, Iliyná A, Malfavon LD, Sánchez O, Dustet MJ (2003) Partial characterization of penicillin acylase from fungi Aspergillus fumigatus and Mucor gryseocianum. Mosc Univ Chem Bull 44:53–56
Das S, Gayen JR, Pal A, Ghosh K, Rosazza JP, Samanta TB (2004) Purification, substrate specificity, and N-terminal amino acid sequence analysis of a β-lactamase-free penicillin amidase from Alcaligenes sp. Appl Microbiol Biotechnol 65(3):281–286
Chakraborty J, Hossain MZ, Tripathi A, Ghosh W, Samanta TB (2012) Taxonomical characterization of a penicillin amidase producing novel strain of Achromobacter xylosoxidans: proposal of A. xylosoxidans subsp. indiges subsp. nov. Dev Microbiol Mol Biol 3:55–65
Priya S, Suganya R (2013) Production and partial characterization of penicillin amidase produced by Bacillus megaterium from Temple Puja Wastes. IJAAEEE 2(1):44–47
Cano-Cabrera JC, Palomo-Ligas L, Flores-Gallegos AC, Martínez-Hernández JL, Rodríguez-Herrera R (2020) Penicillin G acylase production by Mucor griseocyanus and the partial genetic analysis of its pga gene. Int Microbiol:1–9
Bigot Y, Lutcher F, Hamelin MH, Périquet G (1992) The 28S ribosomal RNA-encoding gene of Hymenoptera: inserted sequences in the retrotransposon-rich regions. Gene 121(2):347–352
Ljubijankić G, Storici F, Glišin V, Bruschi CV (1999) Synthesis and secretion of Providencia rettgeri and Escherichia coli heterodimeric penicillin amidases in Saccharomyces cerevisiae. Gene 228(1-2):225–232
Vohra PK, Sharma R, Kashyap DR, Tewari R (2001) Enhanced production of penicillin G acylase from a recombinant Escherichia coli. Biotechnol Lett 23(7):531–535
Marešová H, Štěpánek V, Kyslik P (2001) A chemostat culture as a tool for the improvement of a recombinant E. coli strain over-producing penicillin G acylase. Biotechnol Bioeng 75(1):46–52
Ševo M, Degrassi G, Skoko N, Venturi V, Ljubijankić G (2002) Production of glycosylated thermostable Providencia rettgeri penicillin G amidase in Pichia pastoris. FEMS Yeast Res 1(4):271–277
Kasche V, Galunsky B, Ignatova Z (2003) Fragments of pro-peptide activate mature penicillin amidase of Alcaligenes faecalis. Eur J Biochem 270(23):4721–4728
Ignatova Z, Mahsunah A, Georgieva M, Kasche V (2003) Improvement of posttranslational bottlenecks in the production of penicillin amidase in recombinant Escherichia coli strains. Appl Environ Microbiol 69(2):1237–1245
Wen Y, Feng M, Yuan Z, Zhou P (2005) Expression and overproduction of recombinant penicillin G acylase from Kluyvera citrophila in Escherichia coli. Enzym Microb Technol 37(2):233–237
De León-Rodríguez A, Rivera-Pastrana D, Medina-Rivero E, Flores-Flores JL, Estrada-Baltazar A, Ordóñez-Acevedo LG, de la Rosa AP (2006) Production of penicillin acylase by a recombinant Escherichia coli using cheese whey as substrate and inducer. Biomol Eng 23(6):299–305
Arshad R, Farooq S, Ali SS (2010) Improvement of penicillin G acylase expression in Escherichia coli through UV induced mutations. Braz J Microbiol 41(4):1133–1141
Karthikeyan R, Surianarayanan M, Sudharshan S, Gunasekaran P, Baran MA (2011) Biocalorimetric and respirometric studies on production of Penicillin G acylase from Bacillus badius pac in E. coli DH5α. Biochem Eng J 55(3):223–239
Torres LL, Ferreras ER, Cantero Á, Hidalgo A, Berenguer J (2012) Functional expression of a penicillin acylase from the extreme thermophile Thermus thermophilus HB27 in Escherichia coli. Microb Cell Factories 11(1):1–2
Orr V, Scharer J, Moo-Young M, Honeyman CH, Fenner D, Crossley L, Suen SY, Chou CP (2012) Integrated development of an effective bioprocess for extracellular production of penicillin G acylase in Escherichia coli and its subsequent one-step purification. J Biotechnol 161(1):19–26
Marešová H, Palyzová A, Plačková M, Grulich M, Rajasekar VW, Štěpánek V, Kyslíková E, Kyslík P (2017) Potential of Pichia pastoris for the production of industrial penicillin G acylase. Folia Microbiol 62(5):417–424
Pan X, Yu Q, Chu J, Jiang T, He B (2018) Fitting replacement of signal peptide for highly efficient expression of three penicillin G acylases in E. coli. Appl Microbiol Biotechnol 102(17):7455–7464
Borčinová M, Raschmanová H, Zamora I, Looser V, Marešová H, Hirsch S, Kyslík P, Kovar K (2020) Production and secretion dynamics of prokaryotic Penicillin G acylase in Pichia pastoris. Appl Microbiol Biotechnol 104(13):5787–5800
Parmar A, Kumar H, Marwaha SS, Kennedy JF (2000) Advances in enzymatic transformation of penicillins to 6-aminopenicillanic acid (6-APA). Biotechnol Adv 18(4):289–301
Nandi A, Pan S, Potumarthi R, Danquah MK, Sarethy IP (2014) A proposal for six sigma integration for large-scale production of penicillin G and subsequent conversion to 6-APA. J Anal Methods Chem:413616. https://doi.org/10.1155/2014/413616
Wilms B, Hauck A, Reuss M, Syldatk C, Mattes R, Siemann M, Altenbuchner J (2001) High-cell-density fermentation for production of L-N-carbamoylase using an expression system based on the Escherichia coli rhaBAD promoter. Biotechnol Bioeng 73(2):95–103
Viegas SC, Schmidt D, Kasche V, Arraiano CM, Ignatova Z (2005) Effect of the increased stability of the penicillin amidase mRNA on the protein expression levels. FEBS Lett 579(22):5069–5073
Ohashi H, Katsuta Y, Nagashima M, Kamei T, Yano M (1989) Expression of the Arthrobacter viscosus penicillin G acylase gene in Escherichia coli and Bacillus subtilis. Appl Environ Microbiol 55(6):1351–1356
Kang JH, Hwang Y, Yoo OJ (1991) Expression of penicillin G acylase gene from Bacillus megaterium ATCC 14945 in Escherichia coli and Bacillus subtilis. J Biotechnol 17(2):99–108
Cai G, Zhu S, Yang S, Zhao G, Jiang W (2004) Cloning, overexpression, and characterization of a novel thermostable penicillin G acylase from Achromobacter xylosoxidans: probing the molecular basis for its high thermostability. Appl Environ Microbiol 70(5):2764–2770
Cheng S, Wei D, Song Q, Zhao X (2006) Immobilization of permeabilized whole cell penicillin G acylase from Alcaligenes faecalis using pore matrix crosslinked with glutaraldehyde. Biotechnol Lett 28(14):1129–1133
Wang T, Zhu H, Ma X, Fei Z, Ma Y, Wei D (2006) Enhancing enzymatic activity of penicillin G acylase by coexpressing pcm gene. Appl Microbiol Biotechnol 72(5):953–958
Jiang YM, Tong WY, Wei DZ (2007) Effects of induction starting time and Ca2+ on expression of active penicillin G acylase in Escherichia coli. Biotechnol Prog 23(5):1031–1037
Hunt PD, Tolley SP, Ward RJ, Hill C, Dodson GG (1990) Expression, purification and crystallization of penicillin G acylase from Escherichia coli ATCC 11105. Protein Eng Des Sel 3(7):635–639
Gottesman S (1996) Proteases and their targets in Escherichia coli. Annu Rev Genet 30(1):465–506
Liu YC, Liao LC, Wu WT (2000) Cultivation of recombinant Escherichia coli to achieve high cell density with a high level of penicillin G acylase activity. Proceedings of the National Science Council, Republic of China. Part B Life Sci 24(4):156–160
Narayanan N, Xu Y, Chou CP (2006) High-level gene expression for recombinant penicillin acylase production using the araB promoter system in Escherichia coli. Biotechnol Prog 22(6):1518–1523
Rajendhran J, Gunasekaran P (2007) Molecular cloning and characterization of thermostable β-lactam acylase with broad substrate specificity from Bacillus badius. J Biosci Bioeng 103(5):457–463
Kafshnochi M, Farajnia S, Aboshof R, Babaei H, Aminolroayaee M (2010) Cloning and over-expression of Penicillin G acylase in Escherichia coli BL21. Afr J Biotechnol 9(18):2697–2701
Mönster A, Villain L, Scheper T, Beutel S (2013) One-step-purification of penicillin G amidase from cell lysate using ion-exchange membrane adsorbers. J Membr Sci 444:359–364
Arshad R, Farooq S, Iqbal N, Ali SS (2006) Mutagenic effect of acridine orange on the expression of penicillin G acylase and β-lactamase in Escherichia coli. Lett Appl Microbiol 42(2):94–101
Erarslan A, Güray A (1991) Kinetic investigation of penicillin G acylase from a mutant strain of Escherichia coli ATCC 11105 immobilized on oxirane–acrylic beads. J Chem Technol Biotechnol 51(2):181–195
Forney LJ, Wong DC (1989) Alteration of the catalytic efficiency of penicillin amidase from Escherichia coli. Appl Environ Microbiol 55(10):2556–2560
Xu H, Petersen EI, Petersen SB, El-Gewely MR (1999) Random mutagenesis libraries: optimization and simplification by PCR. Biotechniques 27(6):1102–1108
Hanefeld U, Gardossi L, Magner E (2009) Understanding enzyme immobilisation. Chem Soc Rev 38(2):453–468
Illanes A, Wilson L (2014) Synthesis of b-lactam antibiotics with penicillin acylases. In: Illanes A,(Eds). Enzyme biocatalysis: principles and applications. Springer, United Kingdom, pp 273–292
Kallenberg AI, van Rantwijk F, Sheldon RA (2005) Immobilization of penicillin G acylase: the key to optimum performance. Adv Synth Catal 347(7-8):905–926
Sheldon RA, van Pelt S (2013) Enzyme immobilisation in biocatalysis: why, what and how. Chem Soc Rev 42(15):6223–6235
De Vroom E, Gist Brocades BV (2009) Penicillin G acylase immobilized with a crosslinked mixture of gelled gelatin and amino polymer. United States Patent US 6(060):268
Norouzian D, Javadpour S, Moazami N, Akbarzadeh A (2002) Immobilization of whole cell penicillin G acylase in open pore gelatin matrix. Enzym Microb Technol 30(1):26–29
Hsiau LT, Lee WC, Wang FS (1997) Immobilization of whole-cell penicillin G acylase by entrapping within polymethacrylamide beads. Appl Biochem Biotechnol 62(2):303–315
Arroyo M, De la Mata I, Acebal C, Castillón MP (2003) Biotechnological applications of penicillin acylases: state-of-the-art. Appl Microbiol Biotechnol 60(5):507–514
Abian O, Mateo C, Fernández-Lorente G, Palomo JM, Fernández-Lafuente R, Guisán JM (2001) Stabilization of immobilized enzymes against water-soluble organic cosolvents and generation of hyper-hydrophilic micro-environments surrounding enzyme molecules. Biocatal Biotransfor 19(5-6):489–503
Cao L, van Rantwijk F, Sheldon RA (2000) Cross-linked enzyme aggregates: a simple and effective method for the immobilization of penicillin acylase. Org Lett 2(10):1361–1364
Katchalski-Katzir E, Kraemer DM (2000) Eupergit® C, a carrier for immobilization of enzymes of industrial potential. J Mol Catal B-Enzym 10(1-3):157–176
Mateo C, Abian O, Fernandez-Lafuente R, Guisan JM (2000) Increase in conformational stability of enzymes immobilized on epoxy-activated supports by favoring additional multipoint covalent attachment. Enzym Microb Technol 26(7):509–515
Wang W, Deng L, Peng ZH, Xiao X (2007) Study of the epoxydized magnetic hydroxyl particles as a carrier for immobilizing penicillin G acylase. Enzym Microb Technol 40(2):255–261
Chen CI, Chen CW, Huang CW, Liu YC (2007) Simultaneous purification and immobilization of penicillin G acylase using bifunctional membrane. J Membr Sci 298(1-2):24–29
Mohy Eldin MS, El Enshasy HA, Hassan ME, Haroun B, Hassan EA (2012) Covalent immobilization of penicillin G acylase onto amine-functionalized PVC membranes for 6-APA production from penicillin hydrolysis process. II. Enzyme immobilization and characterization. J Appl Polym Sci 125(5):3820–3828
Liu R, Huang W, Pan S, Li Y, Yu L, He D (2020) Covalent immobilization and characterization of penicillin G acylase on magnetic Fe2O3/Fe3O4 heterostructure nanoparticles prepared via a novel solution combustion and gel calcination process. Int J Biol Macromol 162:1587–1596
Ivanov AE, Edink E, Kumar A, Galaev IY, Arendsen AF, Bruggink A, Mattiasson B (2003) Conjugation of penicillin acylase with the reactive copolymer of N-isopropylacrylamide: a step toward a thermosensitive industrial biocatalyst. Biotechnol Prog 19(4):1167–1175
Coleman K (2011) Diazabicyclooctanes (DBOs): a potent new class of non-β-lactam β-lactamase inhibitors. Curr Opin Microbiol 14(5):550–555
Batchelor FR, Doyle FP, Nayler JH, Rolinson GN (1959) Synthesis of penicillin: 6-aminopenicillanic acid in penicillin fermentations. Nature 183(4656):257–258
Rolinson GN (1952) Respiration of Penicillium chrysogenum in penicillin fermentations. Microbiol 6(3-4):336–343
Rolinson GN (1998) Forty years of beta-lactam research. J Antimicrob Chemother 41(6):589–603
Tipper DJ, Strominger JL (1965) Mechanism of action of penicillins: a proposal based on their structural similarity to acyl-D-alanyl-D-alanine. Proc Natl Acad Sci U S A 54(4):1133
Kong KF, Schneper L, Mathee K (2010) Beta-lactam antibiotics: from antibiosis to resistance and bacteriology. Apmis 118(1):1–36
Worthington RJ, Melander C (2013) Overcoming resistance to β-lactam antibiotics. J Organomet Chem 9:4207–4213
Abraham EP, Chain E (1940) An enzyme from bacteria able to destroy penicillin. Nature 146(3713):837
Wright PM, Seiple IB, Myers AG (2014) The evolving role of chemical synthesis in antibacterial drug discovery. Angew Chem Int 53(34):8840–8869
Pawlowski AC, Johnson JW, Wright GD (2016) Evolving medicinal chemistry strategies in antibiotic discovery. Curr Opin Biotechnol 42:108–117
Nayler JH (1991) Semi-synthetic approaches to novel penicillins. Trends Biochem Sci 16:234–237
Beta-lactam and Beta-lactamase Inhibitors Market by Drug Class, Disease and Route of Administration: Global Opportunity Analysis and Industry Forecast 2019-2028. Report Linker. ID: 5816614. https://www.reportlinker.com/p05816614/?utm_source=PRN. Accessed 9 June 2020
Penicillin G Acylase Market - Global Industry Analysis, Size, Share, Growth, Trends, and Forecast 2019 – 2027. Transparency market research. https://www.transparencymarketresearch.com/penicillin-g-acylase-market.html. Accessed 9 June 2020
Gaude GS, Hattiholli J (2013) Rising bacterial resistance to beta-lactam antibiotics: Can there be solutions. J NTR Univ Health Sci 2(1):4
Acknowledgements
The authors do not acknowledge anyone for the present work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
KS wrote the MS, and RVS conceptualized, reviewed, edited, and supervised. Both the authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
NA
Consent for publication
NA
Competing interests
KS and RVS declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sambyal, K., Singh, R.V. Exploitation of E. coli for the production of penicillin G amidase: a tool for the synthesis of semisynthetic β-lactam antibiotics. J Genet Eng Biotechnol 19, 156 (2021). https://doi.org/10.1186/s43141-021-00263-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-021-00263-7