
Abstract
Background
Nitrogen and potassium are crucial supplements for plant development and growth. Plants can detect potassium and nitrate ions in soils and in like way, they modify root-to-shoot transport of these ions to adjust the conveyance among roots and shoots. Transcription factor MYB59 plays essential roles in numerous physiological processes inclusive of hormone response, abiotic stress tolerance, plant development, and metabolic regulation. In this study, we retrieved 56 MYB59 proteins from different plant species. Multiple sequence alignment, phylogenetic tree, conserved motif, chromosomal localization, and cis-regulatory elements of the retrieved sequences were analyzed. Gene structure, protein 3D structure, and DNA binding of OsMYB59 indica were also predicted. Finally, we characterized OsMYB59 and its function under low K+/NO3− conditions in Oryza sativa subsp. indica.
Results
Data analysis showed that MYB59s from various groups separated in terms of conserved functional domains and gene structure, where members of genus Oryza clustered together. Plants showed reduced height and yellowish appearance when grown on K+ and NO3− deficient medium. Quantitative real-time PCR uncovered that the OsMYB59 reacted to abiotic stresses where its expression was increased in BRRI dhan56 but decreased in other varieties on K+ deficient medium. In addition, OsMYB59 transcript level increased on NO3− deficient medium.
Conclusions
Our results can help to explain the biological functions of indica rice MYB59 protein and gave a theoretical premise to additionally describe its biological roles in response to abiotic stresses particularly drought.
Similar content being viewed by others


Background
Abiotic stress, such as drought, submergence, salt, temperature, and other environmental extremes, affects the majority of plants. Plants establish unique ways to adapt to severe climatic conditions in order to compensate for these circumstances. Membrane transport and perception systems play crucial and necessary roles in maintaining cell homeostasis under stressful situations [39]. Drought stress limits the development of roots in plants and the diffusion rates of potassium ions (K+) toward the roots in the soil, restricting the K uptake. During this reduction in K+ levels, plants resilience to drought stress and K+ absorption might be further depressed. Consequently, it is essential to maintain enough plant K for drought tolerance [54]. Also, overexpressing the potassium channels (such as AKT1) in rice has impacts on plants’ tolerance to drought [2]. On the other hand, nitrate (NO3−) is the most common type of nitrogen that plants utilize for development and growth. Plants obtain nitrogen by either atmospheric nitrogen fixation or by using N sources found in soil, such as nitrate, ammonium, urea, and organic forms of N. Nitrogen availability may influence drought tolerance in different ways. Under optimal water conditions and a high nitrate supply, there is enhanced root water absorption. Again, nitrate is associated with stomatal conductance and aquaporin expression, which may regulate plant adaptation to low water supply ([62]; Ding et al. 2016 [8]; Ding et al. 2018 [9]).
Potassium and nitrate ions enter into the plant by different transporters and channels, which convey K+ and NO3− directly into root cells. In Arabidopsis thaliana, K+ transporters such as High Affinity K+ Transporter5 (HAK5) and K+ Absorption Permease7 (KUP7), as well as the K+ channel—Arabidopsis K+ Transporter1 (KT1), are important proteins involved in K+ uptake into the plant cells [17, 45, 48]. On the other hand, the assimilation of NO3− is intervened by the Nitrate Transporter 1/Peptide Transporter (NRT1/PTR) and the transporters of Nitrate Transporter 2 (NRT2) families [27, 55, 63]. After absorption into root cells, K+ and NO3− are released into xylem vessels and therefore transported toward the shoot. Root to shoot K+ transportation, in Arabidopsis, is mediated by the K+ transporter KUP7 and the K+ channel—Stelar K+ Outward Rectifier (SKOR). The Nitrate Transporter 1.5 (NRT1.5) is engaged with root to shoot NO3− transport in Arabidopsis [32], and is currently called as Nitrate Transporter/Peptide Transporter Family 7.3 (NPF7.3) from the NRT1/PTR family. As a proton coupled K+ antiporter, NPF7.3 can also prevent K+ translocation from root to shoot [29]. Thus, in Arabidopsis, NPF7.3 governs the coordinated transport of K+ and NO3− from the root to the shoot [10, 36]. Meanwhile, regulation of SKOR and NPF7.3 mRNA are done at the level of transcription and the inclusion of NO3− upregulates both SKOR and NPF7.3 transcripts. When K+ levels are low, the transcript level of NPF7.3 is reduced, preventing K+/NO3− transport from root to shoot [29]. Thus, transcriptional control of both SKOR and NPF7.3 might accomplish the coordinated transport of K+/NO3− from root to shoot in Arabidopsis. In rice (Oryza sativa), there are several key transporters involved into K+ and NO3− translocation such as OsHAK, OsKAT, OsHKT (for K+ transport) and OsNPF2.4, OsNPF2.2, OsNAR2.1, OsNRT2.3a (for NO3− transport).
Transcription factors regulate gene expression and thereby govern a variety of essential biological processes. They can be classified into different families based on the characteristics of their DNA binding domains [47]. Transcription factors that contain DNA binding MYB domain comprise a large family that have a variety of roles in eukaryotes [15]. MYB type transcription factors share similar domain architecture. At the N-terminus, they have a DNA-binding MYB domain comprising 1 to 4 imperfect repeats residues and at the C-terminus, they have transcription activation or repression domain. Repeats of the MYB domain have a structure of helix-turn-helix (HTH) and each repeat is about 52 amino acids long. Moreover, MYB transcription factors are divided into four types: 1R, R2R3, 3R, and 4R, each with one to four repeats [12]. Although R2R3 MYB proteins are only found in terrestrial plants, they make up the largest subfamily of MYB type transcription factors [15]. In Arabidopsis thaliana, for example, more than 100 R2R3 MYB proteins have been identified. The R2R3-MYB proteins include important roles in cell metabolism, determination of cell fate, growth, and biotic/abiotic stress responses. Many R2R3-MYB proteins, like Arabidopsis AtMYB32 [44], Triticum aestivum TaMYB1D [57], Prunus persica PpMYB18 [67], Chrysanthemum morifolium CmMYB1 [50], and Pinus taeda PtMYB14 [4], are used for flavonoid and lignin synthesis in different plant species. Oryza sativa has 230 MYB proteins where Arabidopsis has 339 MYB proteins [15]. They perform major functions such as hormone response, abiotic stress tolerance, plant growth, and metabolic control in various physiological processes [12]. In Arabidopsis, the MYB59 DNA binding domain interacts with the promoter of NPF7.3, resulting in increased NPF7.3 expression in response to outer environment K+/NO3− levels. NPF7.3 and MYB59 function along the same pathway for directing root to shoot K+/NO3− transport.
To manage stress conditions, many molecular changes can take place by activating and regulating certain stress-related genes [39]. Identifying important and differentially expressed genes in response to stress environments might be a suitable approach toward a better understanding of stress responses and their mechanisms [23]. Past studies indicated that rice MYB59 is positively regulated with drought [46] and there are little studies about the regulation of OsMYB transcripts under K+ and NO3− stress. Transcription factor OsMYB59 indica is significantly less studied and its function is still to be validated. Under water deficient conditions, OsMYB59 may be upregulated and as previously described, K+/NO3− are closely associated with drought. Recently, functional validation and expression of MYB59 in Arabidopsis under K+ and NO3− deficient conditions were studied [11]. So, characterization and expression profiling of OsMYB59 indica were carried out in this study to know more about the role of this transcription factor.
Methods
Sequence retrieval of transcription factor MYB59 in different plant species, conserved motif analysis, multiple sequence alignment, and phylogenetic analysis
The amino acid, cDNA, CDS sequences, and corresponding annotations were downloaded from the NCBI (http://www.ncbi.nih.nim.gov) and EnsemblPlants (http://plants.ensembl.org/index.html) (Supplementary Table S1). Sequences from 56 plant species were retrieved and saved in a text file.
Conserved motifs were identified using the Meme program (http://meme-suite.org/index.html) [3] with statistical significance. The Meme program was run with default settings except that the maximum number of motifs was defined as 3 and the maximum width was set to 50.
Protein sequences of MYB59 identified from 56 plant species were aligned using Clustal Omega [52], with default parameters. The identical and similar residues of the alignment were shaded using the BoxShade Server (https://embnet.vital-it.ch/software/BOX_form.html) by ExPASy.
A multiple alignment was performed with the full-length amino acid sequences of MYB59 proteins using MEGA-X v. 10.1.7 (https://www.megasoftware.net/) [25]. Unrooted trees were constructed by the maximum-likelihood (ML) method with the following parameters: Poisson model; partial deletion; 1000 bootstrap replicates.
Chromosomal location, gene structure, and cis-element analysis of rice MYB59
All required data regarding chromosomal location of rice MYB59 was retrieved from EnsemblPlants (Accession no: BGIOSGA005283-TA > Location > Region in detail) and mapping was prepared using Microsoft Powerpoint (Office 365), while the gene structure was determined using Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/) [21]. We used PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare /html/) [28] to find out cis-elements.
Analysis of protein features
Physico-chemical features of OsMYB59 were analyzed by ProtParam tool (http://web. expasy.org/protparam/) [16] and sub-cellular localization was predicted by CELLO server (http://cello.life.nctu.edu.tw/) [64].
3D models were generated by SWISS-MODEL (https://swissmodel.expasy.org/) [49], refined by ModRefiner (https://zhanglab.ccmb.med.umich.edu/ModRefiner/) [58], validated by Ramachandran plot (https://servicesn.mbi.ucla.edu/PROCHECK/), and visualized by PyMol (https://pymol.org/2/) [7]. Template 6kks.1.A [53] was used because it has an identity of 52.78% with our target sequence. MYB59 protein and DNA (B-DNA) docking was done using HDOCK SERVER (http://hdock.phys.hust.edu.cn/) [60].
Plant materials, growth conditions, and treatments
In this experiment, a total of three indica rice genotypes were used. Mature seeds of genotypes BRRI dhan56, BRRI dhan48, and BRRI dhan71 were collected from Bangladesh Rice Research Institute (BRRI). Healthy and good-quality mature seeds were selected as explants and for treatment. Characteristics of rice varieties are as follows: BRRI dhan56 is medium salt tolerant and drought tolerant; BRRI dhan48 and BRRI dhan71 are both drought tolerant.
The seeds were dehusked, which were meant for germination. The dehusked seeds were soaked in 70% (v/v) ethanol and shaken for 5 min, followed by several washes. Then, the seeds were treated with 2% sodium hypochlorite (NaOCl) for 2-3 min followed by several washes. After that, they were treated with 0.1% (w/v) HgCl2 for 2-3 min with mild shaking and again washed with autoclaved distilled water several times. Finally, 2 drops of Tween 20 along with autoclaved distilled water were added. After mild shaking for 2 min, the solution was discarded and the seeds were subsequently washed with autoclaved distilled water.
Murashige and Skoog (MS) media was modified to make K+ sufficient medium and low K+ medium such as 1.5 mM MgSO4 was unaltered, 1.25 mM KH2PO4 and 2.99 mM CaCl2 were supplanted by 1.25 mM NH4H2PO4 and 2.99 mM Ca(NO3)2, and 20.6 mM NH4NO3 and 18.79 mM KNO3 were withdrawn. The final K+ concentration in the K+ sufficient medium and low-K+ medium were adjusted using KCl to 5 mM and 100 mM, respectively. On the other hand, media were modified from K+ sufficient medium for NO3− treatment such as 1.5 mM MgSO4 and 1.25 mM NH4H2PO4 were unaltered, 2.99 mM Ca(NO3)2 was supplanted by 2.99 mM CaCl2 and afterward either 5 mM KNO3 or 5 mM KCl was included, respectively, representing NO3− sufficient medium or NO3− deficient medium. All the media contained 0.7% (w/v) agar and 3% (w/v) sucrose. Seeds and plantlets were maintained on sufficient or deficient medium at 25 °C under 16-h-light/8-h-dark photoperiod.
Dehusked seeds of all three varieties were grown on K+ and NO3− sufficient media for 21 days. Then, they were transferred to low K+ (LK) and NO3− deficient (LN) media and kept there for 2 days. Finally, plantlets were transferred back to K+ and NO3− sufficient media. Data was taken during the whole process.
Transcription analyses
For both RT-PCR and RT-qPCR analyses, total RNA was extracted from roots (150 mg) by using the TriZOL reagent (Invitrogen) and then treated with DNase I (Invitrogen™ DNA-free™ DNA Removal Kit) to eliminate genomic DNA contamination. The complementary DNA (cDNA) was synthesized by GoScript™ Reverse Transcription System (Promega Corporation, Madison, USA). Both oligo (dT) primers and random hexamer primers were used for RT-PCR and RT-qPCR analyses.
The eukaryotic elongation factor 1 alpha (eEF-1α) gene was used as an internal standard for normalization of gene expression levels [22]. Specifically designed primers were used to amplify MYB59 transcript (403 bp): RT-PCR was performed for 35 cycles, each at 94 °C for 30 s, 55.7 °C for 50 s, and 72 °C for 1 min. Three independent experiments were performed. The list of primers with respective Tm values is shown in Supplementary Table S2.
RT-qPCR was conducted using a Maxima SYBR Green qPCR Master Mix (2X) (Thermo Fisher Scientific, Waltham, MA, USA) on a StepOne™ Real-Time PCR System machine (Applied Biosystems). The amplification reactions were performed in a total volume of 25 μl, which contained 12.5 μl SYBR Green Master Mix, 8.45 μl nuclease-free water, 0.75 μl forward and reverse primers (10 mM), and 2.5 μl cDNA. PCR was conducted as follows: 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 55.7 °C for 30 s, and 72 °C for 40 s. Three biological replicates were used in one independent RT-qPCR experiment. Three independent experiments were performed in one RT-qPCR analysis.
A melting curve analysis was performed at the end of the PCR run over a range of 65-95 °C. The ∆CT method using a reference gene was used for evaluating the gene expression levels. Finally, Student’s t test and graphs were plotted using GraphPad Prism 6.0.
Results
Conserved motif analysis
Using the MEME tool, we looked for the three most conserved motifs in 56 MYB59 proteins. Motif 1 was the longest, with 50 amino acid residues, followed by motif 2 with 24 and motif 3 with 29 residues (Supplementary Table S3). Long conserved residues present in the aligned sequences can demonstrate the conserved structures of MYB59 proteins among species.
Multiple sequence alignment
Clustal Omega was used to align the MYB59 protein sequences from 56 plant species using the default parameters, where the residues were shaded as identical and similar. As mentioned earlier, we found 3 most conserved motifs with statistical significance. We highlighted only motif 1 and motif 2 in Supplementary Fig. S1 due to higher significance than motif 3. Motif 2 (LNRTGKSCRLRWVNYLHPGLKRGK) and motif 1 (MTPQEERLVLELHAKWGNRWSRIARKLPGRTDNEIKNYWRTHMRKKAQEK) from Oryza sativa indica were very similar to other plants and suggested that these conserved residues could potentially play a key role in the functionality of MYB59 and could be included in metabolic processes.
Phylogenetic analysis of MYB59 proteins
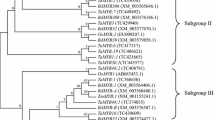
To comprehend the phylogenetic relationship among MYB59s, a maximum likelihood phylogenetic tree was developed, which incorporates both dicotyledonous plants and monocotyledonous plants. The constructed tree was divided into three primary groups based on the tree topology, for example, group A, B, and C. Based on the sub-clustering, group A was further subdivided into two subgroups such as A1 and A2. Group B was likewise subdivided into two subgroups on the basis of sub-clustering (B1 and B2) (Fig. 1). We found 6 homologs of genus Oryza, whereas MYB59s from different groups may differ from one another in terms of gene structure and conserved functional domains. However, even more molecular, genetic, and physiological studies on the functional role/s of MYB59 proteins under various perturbations are needed.

Phylogenetic relationships among MYB59 proteins from 56 plant species with bootstrap values
Gene location and structure of OsMYB59 indica
In this study, the genomic distribution of the OsMYB59 indica in rice plants was investigated through chromosomal mapping. The gene was found on chromosome 1: 47, 136, 245-47, 137, 286 forward strand, as shown by the findings (Supplementary Fig. S2).
To establish the numbers and positions of exons and introns in the OsMYB59 indica gene, an analogy of full-length cDNA sequences with the relevant genomic DNA sequences was made. The coding sequences of the OsMYB59 indica gene were found to be interrupted by an intron (Supplementary Fig. S3).
Cis-element analysis of rice MYB59
5′ UTR sequence (2000bp) of OsMYB59 indica was subjected to classify possible regulatory cis-acting components in PlantCARE database. We found several cis-regulatory elements responsive to abiotic, biotic stresses, and light, as shown in Table 1.
Analysis of protein features
OsMYB59 indica has 276 amino acid residues with a molecular weight of 31.05 kDa and displayed primarily basic characteristics with 6.86 pI value. The subcellular localization of OsMYB59 indica was predicted as nuclear.
3D structure and DNA-binding prediction of OsMYB59 indica
Three-dimensional models were generated by SWISS-MODEL; the model was refined using ModRefiner, and the quality of the model was analyzed using the Ramachandran plot, in which our model showed 98.9% in allowed residues and ERRAT quality factor was A: 100. OsMYB59 indica gene synthesizes a DNA binding R2R3 (Repeat2 Repeat3) transcription factor, containing R2 and R3 amino acid sequence repeats (Fig. 2).

3D model of OsMYB59 indica protein. Models were generated by SWISS-MODEL, refined by ModRefiner, validated by Ramachandran plot, and visualized by PyMol
MYB59 protein and DNA (B-DNA) docking was done using HDOCK SERVER, where the docking score was −217.99 (Fig. 3). After docking, the binding-site residues on the protein (within 3Å) were visualized using PyMOL. We found 9 binding-site residues within 3Å and they were K58, G59, R92, L95, S96, L99, R100, N146, and T154 (Fig. 4).

Predicted structure of the OsMYB59–DNA complex. DNAs are shown as sticks and OsMYB59 is shown as cartoon (a) and as an electrostatic surface potential map (b)

Predicted binding-site residues on protein OsMYB59 within 3Å. OsMYB59 is shown as cartoon-and-stick form and the DNA is shown as spheres. Here, (a) shows K58, G59, and R92, (b) shows L95, (c) shows S96 and L99, (d) shows R100, (e) shows N146, and (f) shows T154 residue
Morphological changes in treatment plants
Plants were germinated on petridish and transferred to K+/NO3− sufficient media or low K+ (LK)/NO3− deficient (LN) media for monitoring the morphological changes. They were transferred after reaching two leaves stage and treated for 7 days. We found yellowish shoots when grown on LK and LN medium. In LK medium, a series of leaf color changes occurred, depending on variety, including yellow and a tan/brown color resulting from leaf death. In severe cases, leaf roll back and die-back (from leaf tips) will proceed. Older leaves or whole plants became yellowish-green in LN medium. Furthermore, older leaves and occasionally all leaves were light green and chlorotic at the tip apart from new greener leaves. The yellowing suggests potassium and nitrate deficiency causing chlorosis, reduced chlorophyll content, and slow growth (Fig. 5).

Morphology of the plants grown into K+ sufficient/low K+ medium in BRRI dhan56 (a), BRRI dhan48 (b), BRRI dhan71 (c), and NO3− sufficient/NO3− deficient medium in BRRI dhan56 (d), BRRI dhan48 (e), BRRI dhan71 (f); bar = 1cm
RT-PCR analysis of the target and housekeeping gene
RT-PCR analysis showed the presence of both OsMYB59 indica and eEF-1α bands. We found size specific bands in all three rice varieties and both treatments. Product sizes of MYB59 and eEF-1α were 403 bp and 103 bp, respectively.
Quantitative reverse transcription PCR (RT-qPCR) analysis of OsMYB59 indica gene in responses to various treatments
Transcription factor MYB59 expression levels were evaluated using RT-qPCR after low K+ or NO3− treatment. Plants germinated on K+/NO3− sufficient (5 mM) medium were used as controls. For treatment, they were then relocated to either low K+ (100 mM—LK) or low NO3− (0 mM—LN) medium for 2 days. The plants were then returned to a medium with sufficient K+ or NO3− (5 mM). For the RT-qPCR assay, roots were acquired at different times, as indicated. The assay was repeated for three different rice varieties with different levels of tolerance to abiotic stresses. Melt curve and amplification plots from the StepOneTM software were used to verify the amplification and specificity of the amplicons.
BRRI dhan56 is a moderate salt and drought-tolerant variety [1, 24]. We saw upregulation of OsMYB59 in both LK and LN medium for this variety. There is a significant increase in the OsMYB59 transcript level on the 1st day of treatment which decreases rapidly in the subsequent days of treatment and the return back to sufficient medium (Fig. 6). However, we found different scenarios in the case of BRRI dhan48 and BRRI dhan71. These two varieties are drought tolerant and developed for Barind Tract of Bangladesh. For both BRRI dhan48 and BRRI dhan71, there is a slight decrease in the OsMYB59 transcript level on the 1st and 2nd day of low K+ treatment, which then increases when returned back to sufficient medium. On the other hand, for low NO3− treatment, OsMYB59 transcript level follows, which was seen in the case of BRRI dhan56 (Fig. 6). With the increasing reaction temperature, double-stranded DNA, with dye molecules, is divided into single-stranded DNA that allows fluorescence to shift and a melting curve to form. In our study, the melt curves prove that only MYB59 and eEF-1α were amplified with the melting temperature of ~87 °C for MYB59 and ~79.5 °C for eEF-1α.

OsMYB59 indica transcription responds to external K+ levels in BRRI dhan56 (a), BRRI dhan48 (b), BRRI dhan71 (c), and external NO3− levels in BRRI dhan56 (d), BRRI dhan48 (e), and BRRI dhan71 (f). Plus (+) and minus (−) symbols represent sufficient and deficient conditions, respectively. RT-qPCR data are shown as means ± SE. Differences between the values of treatments and controls were compared using t tests (*p < 0.05, **p < 0.01, and ***p < 0.001)
Discussion
In order to assess the evolutions within the rice and other species MYB59 proteins, we have performed a phylogenetic study. There is a comparatively long evolutionary distance between Arabidopsis and rice, but we found close distance among the members of the genus Oryza. Consequently, its members’ developmental ties are more conservative than those of other subgroups.
The conserved functional domains we found from MEME analysis can have fundamental roles in different groups of MYB59 proteins. The gene structure, however, varied between different plant species. Therefore, they may have separate downstream regulatory genes and take part in various signaling paths. We found that Arabidopsis and japonica rice has no introns in their gene structure, but indica rice has one intron.
OsMYB59 indica functions may be specified by the arrangements and nature of cis elements on the gene promoter. OsMYB59 indica is primarily active in biotic, abiotic stresses, and light-inducing responsiveness, as seen by the findings. To date, only a few researches on functional analysis of the plant transcription factor OsMYB59 have been carried out. Light causes the plant transcriptome to be massively reprogrammed, increases or decreases gene expression and thus regulates the associated signaling pathway. MYB59 in Arabidopsis is classified as an R2R3 transcription factor. An earlier report found that MYB59 contributes to the regulation of cell cycle progression and root formation [38]. Prior studies showed a high resemblance in gene structure and amino-acid sequence between MYB59 and its nearest homolog MYB48 [63]. Furthermore, both play a role in the jasmonic acid signaling network, which indicates their operational redundancy [19]. Rice MYB59 also applies to the R2R3 type MYB protein and includes cis-regulatory elements that are sensitive to methyl jasmonate (MeJA) [46]. This indicates that both Arabidopsis and rice may have common functions for MYB59.
Although the majority of the residues associated with DNA binding are conserved, one residue (Leu versus Glu) of the R2 repeat significantly contrast among plant and animal MYB proteins, which likely contributes to the variation in their target DNAs. 5mC, as well as 6mA modifications, prohibit the interactions between MYB transcription factors and their target DNAs. The key residues responsible for DNA recognition are profoundly preserved in all R2R3-MYB proteins [53]. In addition, MYB transcription factors and DNA 5mC modifications show opposite functions in many plant specific processes, particularly fruit maturing [26, 33]. The buildup of anthocyanins in apple peel is negatively associated with DNA methylation levels, but contrarily it is positively related to MYB proteins [13]. Numerous R2R3-MYB proteins have been shown to increase anthocyanin [5, 6, 30, 41, 68], however, DNA hypermethylation brought about colorless or non-ripening fruits [35]. Significantly, the L→A mutation in WER (a MYB protein) resulted in stronger binding affinity than that of wild-type WER [53]. In addition, the DNA 6mA modification level is higher in plants compared to mammals. In rice, around 0.2% of adenines are modified by 6mA [66] and a comparative level was also seen in Arabidopsis [31]. DNA 5mC and 6mA modifications may direct gene expression by debilitating the interaction between MYB transcription factors and their target DNAs during plant development [53].
In plant’s responses to the K+ deficiency, transcriptional control is a very essential method. Under K+ deficient conditions, for example, the transcript levels of K+ transporter genes are upregulated to improve high affinity K+ intake of the plant roots [54]. Simultaneously, K+ transport from root to shoot should be decreased to retain enough K+ in the root. This represents an effective adaptive mechanism for maintaining root operation and the root to shoot K+ equilibrium. In order to preserve the required membrane potential and osmotic balance, the root must maintain a minimum concentration of K+ in the event of K+ deficiency in the soil. In comparison, the stem and young leaves reflect a large sink that needs a significant amount of K+ if adequate K+ is provided for plants’ development. As such, more K+ is carried to the shoots to promote plant growth. The root-to-shoot transportation of K+ then must be fine-tuned to external K+ levels. The yield and quality of crops depend on the sufficient supply of potassium and nitrogen. In certain agricultural regions like Asia, however, existing fertilization activity limits agricultural production and leads to several challenges.
In addition to a substantial reduction in nitrogen utilization efficiencies, the excessive usage of nitrogen fertilizer and the absence of potash fertilizers often lead to air/water/soil pollution and deterioration of the environment [18, 43, 65]. The coordination of the K/N use is the molecular mechanism behind this phenomenon. A previous study of Arabidopsis MYB59 showed that under sufficient K+/NO3−, MYB59 associates directly with the NPF7.3 promoter and enables root to shoot K+/NO3− transportation via NPF7.3. If plants suffer from a K+/NO3− deficiency, MYB59 will be downregulated and the NPF7.3 transcript level will be reduced. The root to shoot transport of K+/NO3− is therefore limited [11]. In our study, we found that under low K+ stress, two out of three selected varieties show downregulation of MYB59 transcripts. These two varieties (BRRI dhan48 and BRRI dhan71) are drought tolerant. On the other hand, BRRI dhan56, which is moderate salt and drought tolerant, showed upregulation of MYB59. This suggests MYB59 transcript level under low K+ also depends on the genotype and level of stress tolerance of the variety. Again, under low NO3− stress, all three varieties showed upregulation of MYB59. Therefore, MYB59 possibly regulate one or more of K+ transporters like OsHAK, OsKAT, OsHKT [20, 40, 42, 51, 61], and NO3− transporters like OsNPF2.4, OsNPF2.2, OsNAR2.1, and OsNRT2.3a [14, 34, 56, 59] in rice under stresses. In summary, our findings have helped to clarify the biological functions of rice MYB59 indica and established a theoretical framework to characterize its biological functions further in exposure to abiotic stress.
Conclusions
As the world population increases, rice consumption has placed agriculture on top of the international agenda. To meet the dual challenge of producing enough food and alleviating poverty, more rice needs to be produced at a low unit cost so that the environment and ecosystem services can be safeguarded. At the same time, increased water scarcity in irrigated systems, as well as droughts, salinity, submergence, and global warming, are putting the capacity of rice-producing productive environments in jeopardy.
In our study, we characterized transcription factor MYB59 and analyzed its expression under low K+ and NO3− stress. Transcription factor MYB59 participates in many plant biological processes but has not been systematically studied in rice. We found many cis-regulatory elements that confirm its participation under biotic and abiotic stresses. MYB59s from different groups diverged in terms of gene structure and conserved domains. The prediction of conserved motifs and domains, chromosomal and subcellular localization, and their sequence homology with others provided insight into the structure and putative functions. Real-time quantitative PCR analysis of the OsMYB59 gene subjected to various stressors revealed that they are induced in response to external K+ and NO3− levels and depend on the stress responsiveness of the drought and salt-tolerant varieties. During K+ stress, we found that OsMYB59 is upregulated in BRRI dhan56 (about 3.1-fold increase vs control at 1st day of treatment), but downregulated in BRRI dhan48 (0.37-fold decrease vs control at 1st day of treatment), and BRRI dhan71 (0.3-fold decrease vs control at 1st day of treatment). On the other hand, OsMYB59 is upregulated under NO3− stress in all the three varieties (1.72-fold, 0.19-fold, and 0.33-fold increase vs control at 1st day of treatment in BRRI dhan56, BRRI dhan48, and BRRI dhan71, respectively). Previous studies showed only downregulation of OsMYB and AtMYB59 transcription factors under low K+, and upregulation of AtMYB59 under NO3− deficient conditions. However, these responses are also regulated by light and individual OsMYB59 expressions differed with the presence or absence of light. The availability of this information might encourage researchers for further functional validation.
Availability of data and materials
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
Abbreviations
- BRRI:
-
Bangladesh Rice Research Institute
- SE:
-
Standard error
- bp:
-
Base pair
- PCR:
-
Polymerase chain reaction
- RT-PCR:
-
Reverse transcriptase polymerase chain reaction
- RT-qPCR:
-
Quantitative reverse transcription polymerase chain reaction
- MeJA:
-
Methyl jasmonate
References
Ahmed M, Haq ME, Hossain MM, Md S-a-M, Hasan MM (2017) Performance of four different rice cultivars under drought stress in the north-western part of Bangladesh. Int J Agric Forestry 7(6):134–139. https://doi.org/10.5923/j.ijaf.20170706.03
Ahmad I, Mian A, Maathuis FJM (2016) Overexpression of the rice AKT1 potassium channel affects potassium nutrition and rice drought tolerance. J Exp Bot 67:2689–2698. https://doi.org/10.1093/jxb/erw103
Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS (2009) MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res 37(suppl_2):W202–W208
Bedon F, Bomal C, Caron S, Levasseur C, Boyle B, Mansfield SD, Schmidt A, Gershenzon J, Grima-Pettenati J, Séguin A, MacKay J (2010) Subgroup 4 R2R3-MYBs in conifer trees: gene family expansion and contribution to the isoprenoid-and flavonoid-oriented responses. J Exp Bot 61(14):3847–3864. https://doi.org/10.1093/jxb/erq196
Chen K, Liu H, Lou Q, Liu Y (2017) Ectopic expression of the grape hyacinth (Muscari armeniacum) R2R3-MYB transcription factor gene, MaAN2, induces anthocyanin accumulation in tobacco. Front Plant Sci 8:965. https://doi.org/10.3389/fpls.2017.00965
Dasgupta K, Thilmony R, Stover E, Oliveira ML, Thomson J (2017) Novel R2R3-MYB transcription factors from Prunus americana regulate differential patterns of anthocyanin accumulation in tobacco and citrus. GM Crops Food 8(2):85–105. https://doi.org/10.1080/21645698.2016.1267897
DeLano WL (2002) The PyMOL molecular graphics system. http://www.pymol.org
Ding L, Li Y, Wang Y, Gao L, Wang M, Chaumont F, Shen Q, Guo S (2016) Root ABA accumulation enhances rice seedling drought tolerance under ammonium supply: interaction with aquaporins. Front. Plant Sci 7:1206.https://doi.org/10.3389/fpls.2016.01206
Ding L, Lu Z, Gao L, Guo S, Shen Q (2018) Is nitrogen a key determinant of water transport and photosynthesis in higher plants upon drought stress? Front. Plant Sci 9:1143. https://doi.org/10.3389/fpls.2018.01143
Drechsler N, Zheng Y, Bohner A, Nobmann B, von Wiren N, Kunze R, Rausch C (2015) Nitrate-dependent control of shoot K homeostasis by the nitrate transporter1/peptide transporter family member NPF7.3/NRT1.5 and the stelar K+ outward rectifier SKOR in Arabidopsis. Plant Physiol 169(4):2832–2847. https://doi.org/10.1104/pp.15.01152
Du XQ, Wang FL, Li H, Jing S, Yu M, Li J, Wu WH, Kudla J, Wang Y (2019) The transcription factor MYB59 regulates K+/NO3- translocation in the Arabidopsis response to low K+ stress. Plant Cell 31(3):699–714. https://doi.org/10.1105/tpc.18.00674
Dubos C, Stracke R, Grotewold E, Weisshaar B, Martin C, Lepiniec L (2010) MYB transcription factors in Arabidopsis. Trends Plant Sci 15(10):573–581. https://doi.org/10.1016/j.tplants.2010.06.005
El-Sharkawy I, Liang D, Xu K (2015) Transcriptome analysis of an apple (Malus× domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J Exp Bot 66(22):7359–7376. https://doi.org/10.1093/jxb/erv433
Fan X, Naz M, Fan X, Xuan W, Miller AJ, Xu G (2017) Plant nitrate transporters: from gene function to application. J Exp Bot 68(10):2463–2475. https://doi.org/10.1093/jxb/erx011
Feller A, MacHemer K, Braun EL, Grotewold E (2011) Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J 66(1):94–116. https://doi.org/10.1111/j.1365-313X.2010.04459.x
Gasteiger E, Hoogland C, Gattiker A, Wilkins MR, Appel RD, Bairoch A (2005) Protein identification and analysis tools on the ExPASy server. The Proteomics Protoc Handb 571–607. https://doi.org/10.1385/1-59259-890-0:571
Gierth M, Mäser P, Schroeder JI (2005) The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol 137(3):1105–1114. https://doi.org/10.1104/pp.104.057216
Guo JH, Liu XJ, Zhang Y, Shen JL, Han WX, Zhang WF, Christie P, Goulding KWT, Vitousek PM, Zhang FS (2010) Significant acidification in major chinese croplands. Sci 327(5968):1008–1010. https://doi.org/10.1126/science.1182570
Hickman R, van Verk MC, van Dijken AJH, Mendes MP, Vroegop-Vos IA, Caarls L, Steenbergen M, van der Nagel I, Wesselink GJ, Jironkin A, Talbot A, Rhodes J, de Vries M, Schuurink RC, Denby K, Pieterse CMJ, van Wees SCM (2017) Architecture and dynamics of the jasmonic acid gene regulatory network. Plant Cell 29(9):2086–2105. https://doi.org/10.1105/tpc.16.00958
Horie T, Brodsky DE, Costa A, Kaneko T, Lo Schiavo F, Katsuhara M, Schroeder JI (2011) K+ transport by the OsHKT2;4 transporter from rice with atypical Na+ transport properties and competition in permeation of K+ over Mg2+ and Ca2+ ions. Plant Physiol 156(3):1493–1507. https://doi.org/10.1104/pp.110.168047
Hu B, Jin J, Guo A-Y, Zhang H, Luo J, Gao G (2015) GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 31(8):1296–1297. https://doi.org/10.1093/bioinformatics/btu817
Jain M, Nijhawan A, Tyagi AK, Khurana JP (2006) Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem Biophys Res Commun 345:646–651. https://doi.org/10.1016/j.bbrc.2006.04.140
Javadi SM, Shobbar Z-S, Ebrahimi A, Shahbazi M (2021) New insights on key genes involved in drought stress response of barley: gene networks reconstruction, hub, and promoter analysis. J Genet Eng Biotechnol 19(1):2. https://doi.org/10.1186/s43141-020-00104-z
Kader MA, Aditya TL, Majumder RR, Hore TK, Shalahuddin AKM, Amin A (2020) Development of drought tolerant rice variety BRRI dhan66 for rainfed lowland ecosystem of Bangladesh. Bangladesh Rice J 23(1):45–55. https://doi.org/10.3329/brj.v23i1.46080
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35(6):1547–1549. https://doi.org/10.1093/molbev/msy096
Lang Z, Wang Y, Tang K, Tang D, Datsenka T, Cheng J, Zhang Y, Handa AK, Zhu J-K (2017) Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proc Natl Acad Sci 114(22):E4511–E4519. https://doi.org/10.1073/pnas.1705233114
Léran S, Varala K, Boyer JC, Chiurazzi M, Crawford N, Daniel-Vedele F, David L, Dickstein R, Fernandez E, Forde B, Gassmann W, Geiger D, Gojon A, Gong JM, Halkier BA, Harris JM, Hedrich R, Limami AM, Rentsch D, Seo M, Tsay YF, Zhang M, Coruzzi G, Lacombe B (2014) A unified nomenclature of nitrate transporter 1/peptide transporter family members in plants. Trends Plant Sci 19(1):5–9. https://doi.org/10.1016/j.tplants.2013.08.008
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, van de Peer Y, Rouzé P, Rombauts S (2002) PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 30(1):325–327. https://doi.org/10.1093/nar/30.1.325
Li H, Yu M, Du XQ, Wang ZF, Wu WH, Quintero FJ, Jin XH, Li HD, Wang Y (2017) NRT1.5/NPF7.3 functions as a proton-coupled H+/K+ antiporter for K+ loading into the xylem in arabidopsis. Plant Cell 29(8):2016–2026. https://doi.org/10.1105/tpc.16.00972
Li S, Wang W, Gao J, Yin K, Wang R, Wang C, Petersen M, Mundy J, Qiu J-L (2016) MYB75 phosphorylation by MPK4 is required for light-induced anthocyanin accumulation in Arabidopsis. Plant Cell 28(11):2866–2883. https://doi.org/10.1105/tpc.16.00130
Liang Z, Shen L, Cui X, Bao S, Geng Y, Yu G, Liang F, Xie S, Lu T, Gu X (2018) DNA N6-adenine methylation in Arabidopsis thaliana. Dev Cell 45(3):406–416. https://doi.org/10.1016/j.devcel.2018.03.012
Lin SH, Kuo HF, Canivenc G, Lin CS, Lepetit M, Hsu PK, Tillard P, Lin HG, Wang YY, Tsai CB, Gojon A, Tsay YF (2008) Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 20(9):2514–2528. https://doi.org/10.1105/tpc.108.060244
Liu R, How-Kit A, Stammitti L, Teyssier E, Rolin D, Mortain-Bertrand A, Halle S, Liu M, Kong J, Wu C (2015) A DEMETER-like DNA demethylase governs tomato fruit ripening. Proc Natl Acad Sci 112(34):10804–10809. https://doi.org/10.1073/pnas.1503362112
Luo B, Chen J, Zhu L, Liu S, Li B, Lu H, Ye G, Xu G, Fan X (2018) Overexpression of a high-affinity nitrate transporter OsNRT2.1 increases yield and manganese accumulation in rice under alternating wet and dry condition. Frontiers in. Plant Sci 9:1192. https://doi.org/10.3389/fpls.2018.01192
Manning K, Tör M, Poole M, Hong Y, Thompson AJ, King GJ, Giovannoni JJ, Seymour GB (2006) A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat Genet 38(8):948–952. https://doi.org/10.1038/ng1841
Meng S, Peng JS, He YN, Zhang GB, Yi HY, Fu YL, Gong JM (2016) Arabidopsis NRT1.5 mediates the suppression of nitrate starvation-induced leaf senescence by modulating foliar potassium level. Mol Plant 9(3):461–470. https://doi.org/10.1016/j.molp.2015.12.015
Maruyama K, Todaka D, Mizoi J, Yoshida T, Kidokoro S, Matsukura S, Takasaki H, Sakurai T, Yamamoto YY, Yoshiwara K, Kojima M, Sakakibara H, Shinozaki K, Yamaguchi-Shinozaki K (2012) Identification of cis-acting promoter elements in cold-and dehydration-induced transcriptional pathways in Arabidopsis, rice, and soybean.DNA Res 19:37–49. https://doi.org/10.1093/dnares/dsr040
Mu RL, Cao YR, Liu YF, Lei G, Zou HF, Liao Y, Wang HW, Zhang WK, Ma B, Du JZ, Yuan M, Zhang JS, Chen SY (2009) An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis. Cell Res 19(11):1291–1304. https://doi.org/10.1038/cr.2009.83
Muppala S, Gudlavalleti PK, Malireddy KR, Puligundla SK, Dasari P (2021) Development of stable transgenic maize plants tolerant for drought by manipulating ABA signaling through Agrobacterium-mediated transformation. J Genet Eng Biotechnol 19(1):96. https://doi.org/10.1186/s43141-021-00195-2
Obata T, Kitamoto HK, Nakamura A, Fukuda A, Tanaka Y (2007) Rice shaker potassium channel OsKAT1 confers tolerance to salinity stress on yeast and rice cells. Plant Physiol 144(4):1978–1985. https://doi.org/10.1104/pp.107.101154
Oglesby L, Ananga A, Obuya J, Ochieng J, Cebert E, Tsolova V (2016) Anthocyanin accumulation in muscadine berry skins is influenced by the expression of the MYB transcription factors, MybA1, and MYBCS1. Antioxidants 5(4):35. https://doi.org/10.3390/antiox5040035
Okada T, Nakayama H, Shinmyo A, Yoshida K (2008) Expression of OsHAK genes encoding potassium ion transporters in rice. Plant Biotechnology 25(3):241–245. https://doi.org/10.5511/plantbiotechnology.25.241
Pettigrew WT (2008) Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol Plant 133(4):670–681. https://doi.org/10.1111/j.1399-3054.2008.01073.x
Preston J, Wheeler J, Heazlewood J, Song FL, Parish RW (2004) AtMYB32 is required for normal pollen development in Arabidopsis thaliana. Plant J 40(6):979–995. https://doi.org/10.1111/j.1365-313X.2004.02280.x
Pyo YJ, Gierth M, Schroeder JI, Cho MH (2010) High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and postgermination growth under low-potassium conditions. Plant Physiol 153(2):863–875. https://doi.org/10.1104/pp.110.154369
Quan R, Hu S, Zhang Z, Zhang H, Zhang Z, Huang R (2010) Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance. Plant Biotechnol J 8(4):476–488. https://doi.org/10.1111/j.1467-7652.2009.00492.x
Riechmann JL, Heard J, Martin G, Reuber L, Jiang CZ, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR, Creelman R, Pilgrim M, Broun P, Zhang JZ, Ghandehari D, Sherman BK, Yu GL (2000) Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Sci 290(5499):2105–2110. https://doi.org/10.1126/science.290.5499.2105
Rubio F, Guillermo SM, Rodríguez-Navarro A (2000) Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol Plant 109(1):34–43. https://doi.org/10.1034/j.1399-3054.2000.100106.x
Schwede T, Kopp J, Guex N, Peitsch MC (2003) SWISS-MODEL: an automated protein homology-modeling server. Nucleic Acids Res 31(13):3381–3385. https://doi.org/10.1093/nar/gkg520
Shan H, Chen S, Jiang J, Chen F, Chen Y, Gu C, Li P, Song A, Zhu X, Gao H, Zhou G, Li T, Yang X (2012) Heterologous expression of the chrysanthemum R2R3-MYB transcription factor CmMYB2 enhances drought and salinity tolerance, increases hypersensitivity to ABA and delays flowering in Arabidopsis thaliana. Mol Biotechnol 51(2):160–173. https://doi.org/10.1007/s12033-011-9451-1
Shankar A, Singh A, Kanwar P, Srivastava AK, Pandey A, Suprasanna P, Kapoor S, Pandey GK (2013) Gene expression analysis of rice seedling under potassium deprivation reveals major changes in metabolism and signaling components. PLoS One 8(7):e70321. https://doi.org/10.1371/journal.pone.0070321
Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H, Remmert M, Söding J (2011) Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7(1):539. https://doi.org/10.1038/msb.2011.75
Wang B, Luo Q, Li Y, Yin L, Zhou N, Li X, Gan J, Dong A (2020) Structural insights into target DNA recognition by R2R3-MYB transcription factors. Nucleic Acids Res 48(1):460–471. https://doi.org/10.1093/nar/gkz1081
Wang Y, Wu W-H (2013) Potassium transport and signaling in higher plants. Annu Rev Plant Biol 64(1):451–476. https://doi.org/10.1146/annurev-arplant-050312-120153
Wang YY, Hsu PK, Tsay YF (2012) Uptake, allocation and signaling of nitrate. Trends Plant Sci 17(8):458–467. https://doi.org/10.1016/j.tplants.2012.04.006
Wei J, Zheng Y, Feng H, Qu H, Fan X, Yamaji N, Ma JF, Xu G (2018) OsNRT2.4 encodes a dual-affinity nitrate transporter and functions in nitrate-regulated root growth and nitrate distribution in rice. J Exp Bot 69(5):1095–1107. https://doi.org/10.1093/jxb/erx486
Wei Q, Zhang F, Sun F, Luo Q, Wang R, Hu R, Chen M, Chang J, Yang G, He G (2017) A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants. Plant Sci 265:112–123. https://doi.org/10.1016/j.plantsci.2017.09.020
Xu D, Zhang Y (2011) Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys J 101(10):2525–2534. https://doi.org/10.1016/j.bpj.2011.10.024
Yan M, Fan X, Feng H, Miller AJ, Shen Q, Xu G (2011) Rice OsNAR2.1 interacts with OsNRT2.1, OsNRT2.2 and OsNRT2.3a nitrate transporters to provide uptake over high and low concentration ranges. Plant Cell Environ 34(8):1360–1372. https://doi.org/10.1111/j.1365-3040.2011.02335.x
Yan Y, Zhang D, Zhou P, Li B, Huang S-Y (2017) HDOCK: a web server for protein–protein and protein–DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res 45(W1):W365–W373. https://doi.org/10.1093/nar/gkx407
Yang T, Zhang S, Hu Y, Wu F, Hu Q, Chen G, Cai J, Wu T, Moran N, Yu L, Xu G (2014) The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol 166(2):945–959. https://doi.org/10.1104/pp.114.246520
Yang X, Li Y, Ren B, Ding L, Gao C, Shen Q, Guo S (2012) Drought-induced root aerenchyma formation restricts water uptake in rice seedlings supplied with nitrate. Plant Cell Physiol 53:495–504. https://doi.org/10.1093/pcp/pcs003
Yanhui C, Xiaoyuan Y, Kun H, Meihua L, Jigang L, Zhaofeng G, Zhiqiang L, Yunfei Z, Xiaoxiao W, Xiaoming Q, Yunping S, Li Z, Xiaohui D, Jingchu L, Xing-Wang D, Zhangliang C, Hongya G, Li-Jia Q (2006) The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family. Plant Mol Biol 60(1):107–124. https://doi.org/10.1007/s11103-005-2910-y
Yu C, Chen Y, Lu C, Hwang J (2006) Prediction of protein subcellular localization. Proteins: Structure, Function, and Bioinformatics 64(3):643–651. https://doi.org/10.1002/prot.21018
Zhang X (2017) Biogeochemistry: a plan for efficient use of nitrogen fertilizers. Nature 543(7645):322–323. https://doi.org/10.1038/543322a
Zhou C, Wang C, Liu H, Zhou Q, Liu Q, Guo Y, Peng T, Song J, Zhang J, Chen L (2018) Identification and analysis of adenine N6-methylation sites in the rice genome. NaturePlants 4(8):554–563. https://doi.org/10.1038/s41477-018-0214-x
Zhou H, Lin-Wang K, Wang F, Espley RV, Ren F, Zhao J, Ogutu C, He H, Jiang Q, Allan AC, Han Y (2019) Activator-type R2R3-MYB genes induce a repressor-type R2R3-MYB gene to balance anthocyanin and proanthocyanidin accumulation. New Phytol 221(4):1919–1934. https://doi.org/10.1111/nph.15486
Zhou H, Peng Q, Zhao J, Owiti A, Ren F, Liao L, Wang L, Deng X, Jiang Q, Han Y (2016) Multiple R2R3-MYB transcription factors involved in the regulation of anthocyanin accumulation in peach flower. Front Plant Sci 7:1557. https://doi.org/10.3389/fpls.2016.01557
Zhu ZF, Sun CQ, Fu YC, Qian XY, Yang JS, Wang XK (2005) Isolation and analysis of a novel MYC gene from rice. Acta Genet Sin 32:393–398
Acknowledgements
The authors thank Plant Genetic Engineering (PGE) Laboratory, Department of Genetic Engineering and Biotechnology, Shahjalal University of Science and Technology, Sylhet-3114, for providing opportunity to conduct the research.
Funding
This study was financially supported by the US Department of Agriculture (USDA) and The University Grants Commission, People’s Republic of Bangladesh. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
MQI and MNH conceived and designed the research. SHP supervised the experiments. MQI performed the experiments. MQI, MNH, and NAJ analyzed the data. MQI wrote the manuscript. MNH, HH, and NAJ provided guidance and revised the manuscript. MFHB assisted in the bioinformatics work. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Table S1.
List of retrieved MYB59 protein sequences from 56 plants species using NCBI and EnsemblPlants. Supplementary Table S2. List of primers with Tm and product size used in this study. Supplementary Table S3. Most conserved three motifs of MYB59 proteins in 56 plant species detected by using MEME tool. Supplementary Fig. S1. Multiple sequence alignments of the MYB59 proteinsobtained with Clustal Omega. Identical and similar residues were shaded as black and grey, respectively. Shading of the multiple-alignment file was done using BoxShade by ExPASy. Supplementary Fig. S2. Location of rice MYB59 gene on rice chromosome 1. Supplementary Fig. S3. Gene structure of rice MYB59 indica. Exons, introns, and untranslated regions are marked by round red rectangles, black lines, and blue rectangles, respectively. The scale bar at the bottom estimates the lengths of the exons, introns, and untranslated regions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Islam, M.Q., Hasan, M.N., Hoque, H. et al. Characterization of transcription factor MYB59 and expression profiling in response to low K+ and NO3− in indica rice (Oryza sativa L.). J Genet Eng Biotechnol 19, 167 (2021). https://doi.org/10.1186/s43141-021-00248-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-021-00248-6