
Abstract
Background
Shigellosis, also known as bacillary dysentery, is an acute infection of the intestine. The symptoms can vary from mild watery diarrhoea to severe inflammatory bacillary dysentery, which is characterized by fever, intense abdominal cramps, and the presence of blood and mucus in the stools. While the disease typically resolves on its own, it can become life-threatening in immunocompromised individuals or in the absence of adequate medical care.
Main body of the abstract
Shigella is the primary cause of bacillary dysentery worldwide. It is comprised of four distinct species—S. dysenteriae, S. flexneri, S. boydii, and S. sonnei—each with unique genomic characteristics and disease-causing abilities. Shigella spp. have developed resistance to multiple drugs and have also adapted well to the gut environment over time. They have become well-suited to infecting the human gut epithelial cells and causing dysentery. Consequently, numerous studies have investigated the potential application of nanotechnology in the treatment of shigellosis by leveraging its capability for drug delivery and targeted therapy, thereby improving effectiveness while reducing side effects.
Short conclusion
It is crucial to maintain ongoing surveillance and develop new strategies to effectively manage this issue. In this review, we shed light on the present comprehension of distinct Shigella spp. and their potential contribution to the pathogenesis of shigellosis, along with their interaction with the gut microbiota. We also provide insight into how nanotechnology may be a major factor in preventing shigellosis in the future.
Similar content being viewed by others

Background
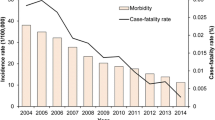
The most prevalent kind of dysentery is shigellosis, often known as bacillary dysentery. Due to their shared genetic and phenotypic traits, the genus Shigella, which is a contemporary member of the Escherichia tribe, is responsible for this enterobacterial illness [1]. Globally, it is estimated that Shigella is responsible for 80–165 million instances of illness and 600,000 fatalities each year, with the majority occurring in children residing in resource poor nations. Of all shigellosis cases worldwide, around 20–119 million illnesses and 6900–30,000 deaths are linked to foodborne transmission [2, 3]. The WHO has identified South Asia and sub-Saharan Africa as the regions with the highest burden [4], albeit regions of heightened activity are also present in Central America, South America, the Ganges–Brahmaputra Delta, and New Guinea island [5]. Shigella spp. is a genus of bacteria that are gram-negative, nonmotile, non-spore-forming, non-lactose fermenting, bacillus-shaped, facultatively anaerobic, and genetically highly correlated with Escherichia coli. There are four species in the genus Shigella (S. dysenteriae, S. boydii, S. sonnei, and S. flexneri), each with numerous serotypes [6]. Shigella boydii is most prevalent in the Indian subcontinent. The two most widespread species worldwide are S. sonnei and S. flexneri. In overcrowded areas with insufficient sanitation and personal cleanliness, the illness is more prevalent [1]. Shigellosis can also arise as an asymptomatic infection to moderate diarrhoea. S. dysenteriae type I represents the most severe and potentially fatal strain of dysentery. Shigellosis brought on by S. dysenteriae type I might occasionally trigger one or more problems [7]. The pathogenesis of shigellosis involves Shigella spp. bacterium. Shigella is typically transmitted via. the faecal–oral route, contaminated food or water sources, and poor hygiene. After being ingested by the host, Shigella starts to invade the intestinal lining, particularly the colon, resulting in inflammation and intestinal mucosal disruption. This results in symptoms like fever, abdominal cramps, watery diarrhoea, and bloody stools [8]. Further insights on how each Shigella spp. involves in propagating shigellosis in humans are discussed under the sub-heading causative microbe’s section.
Common therapy
The first medication, sulphonamides, was introduced in the early 1940s, but their potency had decreased by the late 1940’s. To alleviate this issue, chloramphenicol was recommended followed by tetracycline as a therapy for the management of dysentery. Later research revealed that neither of these medications is efficient. Following that, ampicillin and cotrimoxazole were subsequently made available for purchase. Different antibiotics, including ofloxacin, nalidixic, norfloxacin, and ciprofloxacin, were introduced to the Indian market later in the 1980s for the management of bacillary dysentery and proved to be incredibly effective. Later, more medications like cefotaxime and amikacin entered the market. In 1990, ciprofloxacin was shown to be significantly more effective than drugs like chloramphenicol, ampicillin, and nalidixic acid. Following that, it was reported repeatedly that S. flexneri exhibits 45.6% resistance to ciprofloxacin and 74.1% resistance to nalidixic acid. S. flexneri was discovered to be resistant to fluoroquinolones in 2004; as a result, this medication was no longer favoured for therapy. Following this, the WHO suggested azithromycin, ceftriaxone, and pivmecillinam as substitute medications for treating shigellae that were resistant to fluoroquinolone antibiotics. In recent years, Shigella has grown resistant to these recommended medications. It is alarming that Shigella has developed such extensive resistance to almost all currently used medication classes [9,10,11].
Management of shigellosis
The prevention and control of shigellosis require the implementation of several strategies. Prevention of bloody diarrhoea caused by Shigella relies primarily on measures that prevent the spread of the bacteria within the community, including person-to-person transmission. These measures include ensuring a safe water supply, providing proper sanitation facilities, and practising good personal hygiene and food safety, with a strong emphasis on handwashing. It is crucial to wash hands before eating, before feeding children, after using the toilet, and after handling children's waste. Symptomatic individuals are recommended to avoid sexual contact to reduce transmission. These measures would aid in controlling the disease, but progress is slow in impoverished communities where the disease is most prevalent. Consequently, many individuals believe that the development of a vaccine is the only hope for effectively managing shigellosis [12]. However, creating a vaccine for shigellosis is challenging due to the heterogeneous distribution of Shigella species and serotypes [13]. Nevertheless, progress has been made in developing safe and cost-effective multivalent vaccines. To effectively manage cases of shigellosis, standardized reporting and surveillance practices should be implemented across jurisdictions, taking into account the public health significance of PCR-positive results [8].
Antibiotic treatment is advised for non-resistant shigellosis cases that are moderate to severe. Non-drug-resistant Shigella patients can anticipate clinical improvement within 48 h of antibiotic therapy, leading to reduced chances of severe complications and death, shorter symptom duration, elimination of the bacteria from the stool, and a lower likelihood of transmission to others [13, 14]. Previously, antibiotics such as tetracycline, ampicillin, and cotrimoxazole were highly effective. However, newer fluoroquinolones such as norfloxacin, ciprofloxacin, ofloxacin, azithromycin, and ceftriaxone have proven to be effective as well. Concurrent administration of oral rehydration salt is necessary to prevent or correct dehydration [15]. Overall, the management and prevention of shigellosis depend on an all-encompassing strategy that incorporates effective treatment methods with preventative measures.
Main text
Causative microbes
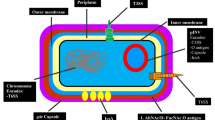
The genus Shigella can be classified into four distinct species, namely S. dysenteriae (serogroup A, which comprises 12 different serotypes); S. flexneri (serogroup B, which consists of 6 distinct serotypes); S. boydii (serogroup C, which encompasses 18 serotypes); and S. sonnei (serogroup D, which consists of a solitary serotype). The physiological makeup of serogroups A, B, and C is extremely similar, although ornithine decarboxylase and positive β-D-galactosidase biochemical responses can distinguish S. sonnei from the other serogroups [16]. Figure 1 shows the general mechanism by which Shigella spp. cause pathogenicity in the host. The following discussion pertains to the four primary categories of Shigella.

Shigella utilizes transcytosis to breach the epithelial cell barrier and interact with macrophage. Proteins (IpaB and IpaC) can be directly injected into host cells in a manner akin to using a syringe owing to the protein secretion mediated by the T3SS. By evoking an apoptotic-like cell death that attracts PMN (polymorphonuclear leukocytes) into the infected tissue and releases pro-inflammatory cytokines that draw neutrophils and trigger innate defences, the bacterium avoids being broken down by the macrophages. Moreover, Shigella effectively penetrates the lamina propria to access the main replicative niche and the cytoplasm of epithelial cells on the basolateral side of the colonic epithelium, from whence it spreads infection from cell to cell. The figure was created using BioRender (trial version) and Wondershare EdrawMax (free version)
Shigella dysenteriae
In 1896, during a major dysentery outbreak in Japan, more than 90,000 cases were reported with a fatality rate close to 30%. Kiyoshi Shiga was the first to isolate Shigella dysenteriae, also known as S. dysenteriae type 1. The next half-century saw the clarification of Shigella spp. microbiology and epidemiology, as well as a thorough investigation of the processes by which the microbe causes illness [17]. One of the main causes of shigellosis or bacillary dysentery is thought to be S. dysenteriae type 1 [18]. There is only one known reservoir for Shigella spp., and that is humans. The bacteria are spread through direct human contact as well as contaminated food and water. In men who have sex with men, there has been evidence of sexual transmission [19]. Lower-middle-class and low-income nations are significantly more likely to have S. dysenteriae infections and intoxications, particularly in children. The loss of intestinal proteins resulting from these diseases might induce chronic diarrhoea and malnourishment [20, 21]. Furthermore, the significance of Shiga toxin [22] in the pathogenesis of dysentery remains unclear, despite the fact that S. dysenteriae type 1 produces this toxin. However, dysentery caused by S. dysenteriae type 1 is generally more severe than that caused by mutants of S. dysenteriae type 1 or other Shigellae that produce little or no Shiga toxin, suggesting that while Shiga toxin is not essential for the pathogenesis of dysentery, it does contribute to the severity of the disease [23]. Shiga toxin produced by S. dysenteriae type 1 predominantly induces haemolytic uraemic syndrome (HUS) in patients, with the secreted toxin resulting in over 2,800,000 acute diseases and approximately 3900 HUS cases each year, while 2–7% of S. dysenteriae infections in humans lead to HUS [24]. Moreover, Shigella dysenteriae type 1 strains are notorious for acquiring resistance against a myriad of antibiotics, rendering previously effective drugs ineffective, as they frequently exhibit resistance to inexpensive and ordinary antibiotics while remaining susceptible to more extravagant or intravenous medications across their distribution [7]. Resistance to antimicrobial agents used to treat shigellosis is growing, leaving only two widely available drugs for outpatient treatment: ciprofloxacin and azithromycin. The increasing resistance to these drugs may lead to a time when there are no effective treatment options for shigellosis [25]. Treatment with the right antibiotics shortens the length of diarrhoea, alleviates symptoms, and expedites healing [7]. The global occurrence of S. dysenteriae type 1 infections has decreased since 2000, but the reasons for this are unknown. S. dysenteriae type 1 has previously become rare for many years before resurfacing in fatal epidemics, a pattern that could potentially happen again. Infections with S. dysenteriae types other than type 1 are even less frequent [25].
Shigella flexneri
Gram-negative, rod-shaped bacteria make up the genus Shigella, which includes the species of bacterium known as Shigella flexneri (S. flexneri) [26]. S. flexneri is a significant cause of global bacterial diarrhoea, especially in developing nations. It causes a considerable amount of illness, particularly in children under 5 years old [27]. Research in non-human primates has shown that S. flexneri is a pathogen that lives inside colon epithelial cells [28]. It has specialized structures known as Type III secretion systems (T3SS), which function as complex molecular machinery enabling the direct injection of proteins into host cells resembling a syringe-like action [29]. Injected proteins alter the host cell's processes, enabling bacterial invasion, replication, and dissemination within the colon lining [30], causing damage to the intestinal mucosa through the production of Shiga toxins, also known as verotoxin, resulting in inflammation and haemorrhage [31]. This invasion results in the characteristic shigellosis symptoms [32]. The mechanisms implicated entail the initial interaction of S. flexneri with intestinal epithelial cells of the host, wherein the bacteria can attach to the cell surface using different adhesins and outer membrane proteins, ultimately culminating in the assembly of the T3SS of S. flexneri [33]. The bacteria employ a translocator protein known as IpaB, or Invasion plasmid antigen B [34], which creates pores in the membrane of the host cell. This protein is a crucial element of the T3SS. IpaB functions as a conduit through which effector proteins are transported into the cytoplasm of the host cell. Additionally, it collaborates with another translocator protein called IpaC, or Invasion plasmid antigen C. These translocators are secreted by the S. flexneri T3SS apparatus via the needle, forming a complex called IpaB/IpaC translocon [35, 36], which penetrates the host cell membrane and opens a channel to facilitate the transfer of effector proteins from the bacterium into the cytoplasm of the host cell. This process is vital for modifying signalling pathways in host cells, rearranging actin, and creating a space within the cell for S. flexneri [37]. The essential translocator proteins associated with the T3SS of S. flexneri are IpaB and IpaC [37]. Nevertheless, additional effector proteins, such as IpaD [38], VirA [39], and IcsA [40], might also contribute to the effective invasion of the host cell, either through direct or indirect means. Furthermore, S. flexneri causes a severe inflammatory response in the gut mucosa [32]. The infiltration of epithelial cells and subsequent annihilation emancipates pro-inflammatory cytokines, like interleukin-1 (IL-1), interleukin-6 (IL-6), and tumour necrosis factor-alpha (TNF-alpha). These cytokines beckon immune cells, encompassing neutrophils and macrophages, to the location of infection, thus engendering additional detriment to tissues and inflammation [41]. The clinical symptoms of bacillary dysentery caused by S. flexneri, such as severe diarrhoea with abdominal pain, bloody and mucus-filled diarrhoea, fever, and inflammation of the intestinal lining, are a result of the combined action of these mechanisms [27].
Shigella boydii
Shigella boydii (S. boydii) is a gram-negative bacterium that is typically nonmotile and does not produce spores [42]. However, there are some shreds of evidence suggesting the presence of flagella, which are about 10 microns long and 12–14 nm in diameter, primarily located at one end of the cell, although their motility is not essential for intestinal infection [43]. The electrophoretic examination of flagellins demonstrated that the movement of S. boydii C3 flagellin is quicker than that of S. flexneri flagellin despite the fact that the estimated molecular mass of C3 flagellin (58 kD) is higher than that of S. flexneri flagellin (56.6 kD) [44]. The flagella genes in S. dysenteriae, S. flexneri, S. boydii, and S. sonnei exhibit variation, contributing to the genetic diversity within these species [43].
18 serotypes of S. boydii belonging to serogroup C have been documented [16]; however, Akter et al. [45] identified around 20 serotypes, with S. boydii type 1 being the second most commonly observed serotype among the various serotypes of S. boydii in Bangladesh. Within 12–48 h after consuming food tainted with these germs, shigellosis develops. Initial symptoms include fever, pains, weariness, and appetite loss. These symptoms may be accompanied by watery diarrhoea, which can potentially progress into the presence of bloody stools or dysentery. Certain severe cases can lead to the development of a fatal HUS caused by the production of Shiga toxin [46], while the severity of the disease varies for different strains of Shigella bacteria (low in S.sonnei and S. boydii, moderate to severe in S. flexneri, and severe in S. dysenteriae) [47]. In the GEMS study, a more detailed analysis revealed that the majority of the isolates were comprised of S. flexneri accounting for 65.9%, with S. sonnei coming in second with 23.7%, while the percentages for S. dysenteriae and S. boydii were 5.0% and 5.4% [48]. In affluent nations, S. boydii is comparatively uncommon and is usually connected to people who have visited endemic regions [49]. Furthermore, examination of the whole genome sequencing (WGS) information also demonstrated a strong phylogenetic connection between S. boydii serotypes 1 and 20, suggesting that serotype 20 emerged recently from serotype 1. Travel reports from patients to Egypt, Afghanistan, India, and Sierra Leone indicate that this new serotype is widely distributed regionally, even if the PHE (Public Health England) archives' travel data for isolates S. boydii 20 were not fully completed [50]. S. boydii outbreaks are more prevalent in Central and South America compared to other regions [51], and there is limited research available on certain serotypes of S. boydii [45].
Shigella sonnei
Shigella sonnei (S. sonnei), a Gram-negative facultative intracellular pathogen, was named 'Sonne's bacillus' after Carl Olaf Sonne, who identified it as the cause of bacillary dysentery [52]. Watery diarrhoea is the main clinical manifestation in most S. sonnei-infected patients. There is just one serotype of S. sonnei [53], and it can be distinguished from other serogroups by specific biochemical reactions with β-D-galactosidase and ornithine decarboxylase [16]. S. sonnei, originating from Europe, has now spread globally and [54] has emerged as the dominant subgroup in Asian countries [55]. The prevalence of S. sonnei is more common in industrialized nations compared to developing countries and results in less severe illness than S. dysenteriae and S. flexneri [56]. The spread of S. sonnei across borders is often associated with international travel and cross-border food trade [57]. The persistence of S. sonnei is facilitated by water and food contamination, fomites, unsanitary conditions, and ecological factors, leading to the emergence of severe outbreaks [27]. The rapid increase in S. sonnei has been attributed to potential factors such as passive immunization caused by Plesiomonas shigelloides and the favourable environment provided by the widespread amoeba species Acanthamoeba castellanii [58]. Food, water, insects, fauna, birds, and amoeba are among the common reservoirs and modes of transmission that S. sonnei shares with other Shigella species [59]. S. sonnei uses a T3SS to inject effector proteins into host macrophages and epithelial cells after they enter the gastrointestinal system. This is an essential step for tissue invasion and immune response evasion. Like S. flexneri, S. sonnei utilizes IcsA, an adhesin regulated by the T3SS, to adhere to epithelial cells. However, S. sonnei possesses an additional adhesin called multivalent adhesion molecule (MAM SS01327), which also aids in adhesion. These two adhesins work together, along with IcsA, to efficiently attach S. sonnei to host cells [54]. Moreover, S. sonnei has the Type VI Secretion System (T6SS) [60] which helps in host colonization. S. sonnei's possession of T6SS gives it an advantage over S. flexneri, which lacks this system. This advantage could be the reason for S. sonnei's dominance in causing Shigellosis [61]. Colicinogenic plasmids in S. sonnei might also contribute to its dominance [62]. S. sonnei has the group 4 capsule (G4C) which accounts for 8.8% of extracellular capsules. Identical in structure to the O-Antigen (OAg) affixed to the lipid A-core of lipoprotein-associated liposomes (LPS), these capsules, sometimes referred to as OAg capsules, are composed of high molecular weight surface polysaccharides. The bacteria are shielded by the G4C from serum-mediated destruction [63, 64].
Entry of Shigella in the gut and its interaction with the gut microbiome
The human microbiota is made up of trillions of symbiotic microbial cells in each person, mainly in the gut. The human microbiome consists of the genes in these cells [65]. The gut microbiota has important roles in nutrient metabolism, drugs and xenobiotic metabolism, maintaining the integrity of the gut barrier, influencing the immune system, and protecting against pathogens [66]. Shigella has developed the ability to adapt to various environmental conditions (temperature, pH, oxygen or osmolarity) as a result of coevolution, which is facilitated by the expression of specific transcriptional regulators [67]. Shigella species, in contrast to many other bacteria, are highly successful in invasive systems, which allow bacteria to penetrate and grow within the epithelia of the human gut and ultimately cause severe inflammatory colitis, known as shigellosis [68]. Shigella enters the intestinal mucosa after ingestion, causing a severe inflammatory response that causes tissue damage [32].
Shigella secretes neuraminidases and mucinases in the colon to cross the mucus layer and reach the epithelial surface [69], where the iphH gene (encoded by chromosomal DNA and/or recombinant plasmids) allows bacterial cell-to-cell movement and dissemination, while the ial gene which is encoded by plasmids (invasion-associated loci) enables penetration of intestinal epithelial tissues [70, 71]. Oxygen sensing plays a crucial role in infection by priming Shigella at the epithelial surface. Although strict anaerobes are mainly found in the colon, where oxygen levels are low, at the epithelial surface, oxygen concentrations increase as oxygen diffuses from the oxygenated epithelium. Oxygen diffusion may be restricted by the thick mucus layer, but at the epithelial surface oxygen levels remain highest. Shigella senses increasing oxygen content at the epithelial surface to trigger their T3SS and improve invasion, avoiding needless activation of their T3SS while physically distant from their cellular target, even if the transcription of their T3SS is hindered under anaerobic circumstances [72]. T3SS is regarded to be a crucial element for bacterial entrance. It is made up of many proteins, one of which is an oligomer with a needle-like form that is anchored in the protein complex that joins the inner and outer bacterial membranes. An oligomer consisting of the invasion plasmid antigens ipaB, ipaC, and ipaD makes up the needle’s tip [73,74,75]. Conversely, the T3SS regulatory cascade consists of three transcription activators, VirB (ParB family), VirF (AraC-family), and MxiE (AraC family), as well as a repressor called H-NS (histone-like nucleoid structure protein) [76]. Following close bacterial contact with host cells, the T3SS introduces effector proteins into the cells, causing the bacteria to transfer their products into the target cells' cytosol and cause cell invasion, tissue damage, and immune evasion [77, 78], which resembles phagocytosis. Subsequently, Shigella utilizes actin-based motility and effectors to invade neighbouring cells through a process called “cell-to-cell spread”, which shares some similarities with the initial entry [76], and this invasion and spread within the colonic epithelium is the main cause of the severe inflammatory response associated with the infection [79].
Unknown is the precise way in which Shigella interacts with inherent host elements like the microbiota. Ndungo et al. [80] proposed that Shigella infection could potentially affect the development of the microbial community in infancy, while changes in the gastrointestinal microbiome could make individuals more susceptible to infections. Following Shigella's invasion and replication within host cells, the innate immune system promptly detects DAMPs or PAMPs, transmits warning signals to the immune system as a whole, and eventually starts an inflammatory response [81]. Suryavanshi et al. [82] emphasized the crucial role of the gut microbiome in maintaining health and preventing infection through the production of antimicrobial compounds such as bacteriocins, organic acids like acetic acid and lactic acid, and hydrogen peroxide. Conversely, Oyewale et al. [83] have asserted alterations in the intestinal microbial community have significant implications for human health and disease pathogenesis, which can be attributed to lifestyle and the presence of underlying diseases. When the gut is dysbiotic, disease-causing and pathogenic bacteria proliferate and the protecting bacteria disappear [84]. Additionally, Table 1 provides information on how Shigella might exploit dysbiosis circumstances. In the presence of bile salts, Shigella may develop biofilms, which alter the makeup of the gut microbiota, according to certain studies. Chenodeoxycholate, the prime bile salt, stimulates the synthesis of exopolysaccharides, which in turn encourages the formation of biofilms, facilitating their colonization and propagation [85, 86]. Yang et al. [87] have also mentioned that an oral Shigella infection had a quick and noticeable impact on the gut microbiota, primarily leading to an early stage of infection with an increase in Prevotella and Shigella/Escherichia and a decrease in probiotics like Faecalitalea and Lactobacillus reuteri. On the other hand, a number of strains of Escherichia coli, Lactobacillus, Bifidobacteria, and a recent generation of probiotics, such as Bacteroides thetaiotaomicron and Akkermansia muciniphila, can support intestinal epithelial homeostasis and increase health [88]. Kutshik et al. [89] study demonstrated the potential of probiotics in treating Shigellosis in rats, aligning with the current research that highlights the curative properties of lactic acid bacteria (LAB) against this life-threatening diarrhoea. Lima et al. [90] also mentioned the use of biotherapeutic agents, ideally probiotics, as an alternate therapeutic option for the treatment of infectious gastroenteritis and as a means of preventing antibiotic-induced diarrhoea. It is important to remember, though, that further research in this area is required to determine how probiotics hinder Shigella and boost immunity in the process.
Potential role of nanotechnology in countering the disease
Nanotechnology offers a promising alternative to the conventional treatment methods that usually rely on antibiotics as the main approach for dealing with shigellosis. The excessive use of antibiotics over time can result in the development of antibiotic resistance, making nanomedicine an attractive option. Nanomedicine can be tailored to target infected cells specifically, thereby enhancing treatment effectiveness and reducing potential side effects. Moreover, the unique mechanism of action of nanomedicine may provide a solution to combat Shigella strains that have developed resistance to conventional antibiotics [99, 100]. The potential of nanotechnology in combating shigellosis is being investigated through the utilization of nanoparticles to improve drug delivery, develop targeted antimicrobial agents, and enhance diagnostic techniques, as illustrated in Fig. 2 and extensively discussed below.

This figure demonstrates the diverse functions of nanotechnology in combatting bacterial diseases, emphasizing important mechanisms like targeted drug delivery, nanoscale diagnostics, and antimicrobial nanomaterials. The figure was created using BioRender (trial version) and Wondershare EdrawMax (free version)
Nanomaterials as antibacterial agents
Antimicrobial resistance poses a significant threat to human health at present, which necessitates the use of drugs that are more noxious, costlier, and with low efficiency [99]. To counter antibiotic resistance, nanoparticles have emerged as promising tools that can directly or indirectly combat deadly bacterial infections. Nanomaterials provide a means of accessing novel antibacterial modalities that bacteria do not possess in their natural defence arsenal. The therapeutic impact of nanomaterials largely stems from their confinement at the nanoscale, which allows for multivalent interactions and a high surface-to-volume ratio. Nano-sized metals, organic nanoparticles (NPs), metal oxides, and nanocomposites exhibit potent antibacterial properties and offer strategic advantages for controlling superficial infections and infectious diseases in a safe manner [101]. Silver (Ag) is well recognized among metallic NPs for being the most potent over bacteria and other pathogens. It is also well-suited for usage in medical applications due to its excellent biocompatibility [102]. It has been established as a superior antibacterial agent with the ability to fight bacteria that cause illnesses in vitro as well as in vivo. AgNPs can combat both Gram-positive and Gram-negative bacteria, such as those that are resistant to several drugs [103]. A plethora of research has been conducted to assess and gauge the antibacterial potential of silver and its related compounds. The results have shown that Ag-particles induce protein malfunction, oxidative stress, DNA and membrane damage and ultimately cause harm to microbial cells [104]. Nevertheless, the precise mechanism by which they impede the proliferation of bacteria or exhibit bactericidal action remains unclear. In order to explain how AgNPs work against bacteria, scientific evidence currently endorses three main processes that have been noted either together or independently [103]. These mechanisms encompass the following:
-
(1)
Because AgNPs can pass through outer membranes and accumulate in the inner membranes, where their adherence to the cell causes destabilization and destruction, the particles act at the membrane level, increasing membrane permeability and causing cellular content to leak out and ultimately cause the cell to succumb [102, 105].
-
(2)
Nanoparticles not only possess the ability to breach the cell membrane, altering its structure and permeability, but they can also enter the cell, where it has been suggested that due to their properties, AgNPs are thought to interact with phosphorus and sulphur groups within intracellular components like proteins and DNA, thereby affecting their structure and functions. Similarly, they may also disrupt the respiratory chain in the inner membrane by reacting with thiol groups in enzymes, resulting in reactive free radicals and oxygen species, causing damage to intracellular machinery, and initiating the apoptotic process [106,107,108,109].
-
(3)
The discharge of silver ions resulting from NPs, which, owing to their size and charge, can interact with biological components, thereby affecting membranes, metabolic pathways, and even genetic material [107, 110].
For instance—Gurunathan et al. [111] have previously documented the efficacy of antibiotics, AgNPs, and combinations thereof in fighting pathogenic bacteria such as Staphylococcus aureus, Shigella flexneri, Pseudomonas aeruginosa, and Streptococcus pneumoniae. In comparison with antibiotics or AgNPs alone, the results indicate that the combination of both antibiotics and AgNPs demonstrated notable anti-biofilm and antibacterial properties at the lowest concentration of both substances. Wang et al. [112] recently published a study that unveiled an innovative type of nanomaterial called Bacitracin-AgNCs (Bacitracin-Ag Nanoclusters) that exhibited strong antibacterial efficacy against S. flexneri and exhibited the ability to impede bacterial growth even at low concentrations. The mechanism of action involved rupturing the membrane and causing observable morphological alterations and intracellular nucleotide leakage prior to cell proliferation was obstructed.
Nanoparticle-based vaccines
NPs have emerged as a viable option for targeted delivery of vaccines to immune cells, resulting in enhanced vaccine effectiveness through controlled release, facile antigen absorption, and stimulation of both humoral and cellular immune responses [113]. These NPs can be composed of lipids, inorganic materials such as metals and nonmetals, various polymers, and even virus-like particles, all of which have been extensively investigated in research studies. The properties of the NPs have made it possible to target certain antigen-presenting cells to enhance immunization techniques and provide immunity [114]. In a study conducted by Gilavand et al. [115], recombinant MxiH antigen and purified antigen-loaded into chitosan NPs (CS-MxiH) demonstrated several advantages in terms of vaccine efficacy against Shigella infection. Similarly, the authors deduced that vaccine-based CS-NPs had a substantial immunogenic potential to boost humoral and mucosal immunity based on the elevated levels of IgA and IgG in mice subjected to intranasal injection of CS-MxiH. Despite extensive research and development, no effective or secure vaccine against Shigella has received clinical approval. Adjuvants are necessary to produce adequate immunogenicity in conserved recombinant subunit vaccines, notwithstanding the possibility of cross-protection, which may present safety concerns [116]. Koley et al. [117] discovered next-generation outer membrane vesicles (OMVs)-based antigens derived from Shigella during their research. By disrupting the tolA gene in the Tol-Pal system of the Shigella membrane, the release rate of OMVs was increased by ~ 80%. They revealed that among the 50 circulating Shigella subtypes in mice models, there are four serotype-subtype cross-protection. Consequently, OMVs-based immunogens hold promise as affordable non-living, next-generation candidate vaccination for human shigellosis. Baruah et al. [118] designed biomimetic nanovaccines (NVs) based on a 50:50 poly (lactic-co-glycolide)/PLGA blend. These NVs included stabilized antigens or immunostimulants of S. dysenteriae 1 origin that were surface-modified by basic chemical methods. By administering a large dose of heterologous S. flexneri 2a to immunized groups and keeping an eye out for noticeable symptoms such as weight loss, diarrhoea, and survival rates, the cross-protective effectiveness of these NVs was assessed. When challenged with heterologous Shigella, the immunized groups exhibited ~ 70–80% survival rates, providing protection against weight loss and diarrhoea. Therefore, passive defence in neonates suggests that the immunization of parents might protect newborns, who are the most susceptible group in the event of a Shigella infection. Furthermore, on 23 April 2019, the Indian Council of Medical Research (ICMR) granted a licence for the Shigella vaccine technology to MSD Wellcome Trust Hilleman Laboratories Pvt Ltd for expanded development and commercialization. The Biotech Consortium India Limited (BCIL), New Delhi, facilitated the License Agreement with NICED on behalf of ICMR and Hilleman Labs [119]. The ICMR-NICED-developed Shigella vaccine is anticipated to have enormous potential and to help children residing in low- and middle-income environments.
Nanoparticle-based drug delivery system
The efficacy of conventional antibiotic therapeutics for shigellosis has been progressively diminished due to the emergence of multidrug-resistant (MDR) strains [120]. In response to this issue, nanoparticle delivery systems have recently emerged as a potential strategy to counteract antibacterial resistance as well as the difficulties in administering antibacterial agents. These challenges include low bioavailability, drug-related toxicities, regular drug dosing schedules, and sub-therapeutic drug buildup in bacterial reservoirs [121]. NP delivery systems offer several advantages, such as enhancing drug solubility, providing immune evasion capabilities, controlling drug release, targeting specific sites, and enabling the delivery of multiple drugs simultaneously. These unique advantages contribute to improving the pharmacokinetic profile and therapeutic index of drug payloads when compared to free drug equivalents. Consequently, several applications focusing on local antimicrobial therapy are seeing a rise in the usage of therapeutic NP, particularly polymeric NP, liposomes, inorganic NP, and dendrimers to increase therapeutic efficacy [122, 123]. According to Mukherjee et al. [124], tetracycline-embedded calcium-phosphate NPs (Tet-CPNPs) can treat mice’s fatal shigellosis, a diarrheal illness brought on by Shigella infection. Tet-CPNP therapy significantly decreased the excretion of mushy stool, weight loss, shortened colon length, and bacterial colonization in the GI tract of mice afflicted with shigellosis, according to their results. Additionally, investigations including immuno- and histological analysis showed that Tet-CPNP administration restored almost normal characteristics to the intestinal tissue of the Shigella-induced mouse model, as well as altered the inflammatory cytokines level of IL-1β, TNF-α, and IFN-γ. In contrast, bulk tetracycline had no effect on shigellosis. NPs serve as carriers (such as solid lipid NPs, liposomes, polymers, etc.), encapsulating antibiotics to extend their half-life [125], and discharge their load in a regulated way, enabling controlled release at the infection site [126]. In addition to reducing the likelihood of side effects and the development of antibiotic resistance since the medication is exposed to fewer non-target bacteria, these nanoparticle-based techniques often improve the physiochemical features of engaged antibiotics, boosting their kinetic rates of absorption and distribution [127, 128].
Biosensors and diagnostics
The field of nanotechnology has made significant contributions to the advancement of biosensors through extensive research on nanomaterials and nanostructures. Various nanomaterials such as carbon nanotubes, graphene quantum dots (GQDs), metal nanoclusters, polymer nanocomposites, metal oxide NPs, plasmonic nanomaterials, and nanogels have been investigated [129]. This research has led to the development of biosensors with immense potential for microbiological diagnosis in real-time. By integrating nanotechnology, biosensors enable rapid, real-time, and accurate detection of molecular biomarkers in actual samples [130]. Typically, biosensors are capable of identifying biomolecules such as nucleic acids, proteins, and cells that are indicative of diseases. This capability is attributed to the three major components of biosensors: the reading device, the physiologically sensitive element, and the detection element [131]. Elahi et al. [132] conducted a study that unveiled an early method of detecting infectious Shigella. The researchers successfully designed a DNA-probe gold NPs (AuNPs)-fluorescence system by immobilizing two DNA probes (sensing element) on the surface of AuNPs. Furthermore, they synthesized iron NPs (Magnetic NPs) that were subsequently altered using Sulfosuccinimidyl 4-Nmaleimidomethyl cyclohexane-1-carboxylate (SMCC). Another system was created by immobilizing a third DNA probe on MNPs to separate the target DNA. The results demonstrated an increase in fluorescence intensity corresponding to an increased concentration of target DNA. Ali et al. [133] devised a DNA biosensor without the need for labels to track S. flexneri. This was achieved by immobilizing the detection probe onto a surface consisting of polyglutamic acid (PGA) and poly-melamine (P-Mel), and using a disuccinimidyl suberate (DSS) functionalized flexible indium tin oxide (ITO) electrode. Signal indication for S. flexneri detection is anthraquinone-2-sulfonic acid monohydrate sodium salt (AQMS). The biosensor demonstrated outstanding recovery rates in detecting S. flexneri within spiked food samples. Various biosensing methods for pathogenic bacteria detection have proven successful and are now under consideration by health authorities and research institutions. This is primarily due to their rapid response, high performance capability, reliable results, and enhanced sensitivity compared to conventional detection methods [129].
Nanofiber wound dressing
The investigation of nanofiber (NF)-based membranes for scaffolds and wound dressings has experienced a rise in recent years due to their notable characteristics such as increased surface area, nano-porosity, and the ability to incorporate drugs or biomolecules [134]. Their size ranges from 50 to 1000 nm and possess low density, high porosity, tiny pore sizes, and substantial surface area. Various methods for nanofiber formulation include thermally induced phase separation, molecular assembly, and electrospinning. Different types of polymers utilized in the production of nanofibers consist of biodegradable hydrophilic polymers, hydrophobic polymers, and amphiphilic polymers [135], all of which are designed to emulate the porous topography of the natural extracellular matrix (ECM) making them advantageous for tissue regeneration [136]. There are two primary categories of nanofibers employed in wound healing to combat Shigella:
-
(1)
NF dressings can be intended to possess a large surface area and the capability to locally administer antibiotics and antibacterial agents to manage infection within the wound milieu [137, 138]. Electrospun NF is considered for pH-mediated and targeted drug delivery to minimize undesirable adverse effects and toxicity in normal tissues [139]. NF prepared through electrospinning are designed for rapid wetting by saliva and will dissolve or disintegrate in the patient's mouth, thereby releasing drugs into the buccal mucosa for immediate absorption without the need to drink or chew. This can be accomplished by employing water-soluble polymers and a significant surface area exposed to the dissolution medium. For controlled release, the drug delivery system must dissolve or disintegrate within a specific time frame. Controlled-release methods, whether oral or transcutaneous, enable the administration of pharmaceutical drugs once or twice daily, enhancing patient compliance and reducing the toxic plasma peak concentrations associated with repeated immediate-release formulations [140].
-
(2)
Due to their distinctive features, NF can be employed to enhance wound healing. Additionally, their spatial structure mimics the ECM [141], a connective network composed of fibrous glycoproteins that coordinate in vivo to furnish the mechanical stability, physical scaffolding, and biochemical cues requisite for tissue morphogenesis and homeostasis [142]. These scaffolds, resembling the ECM, support repair and regeneration, hasten wound healing, and aid in the rapid restoration of functional gastrointestinal tissue [143, 144]. Effective applications typically involve sponges, foams, hydrogels, and nanofibrous networks with highly porous structures as scaffolds [145].
To enhance the treatment of Shigella, the nanofibers can be altered to include probiotics and other active agents, resulting in improved stability of NPs within the host body, aiding in the treatment and prevention, while ensuring that the probiotics stay in the GI tract [87, 138, 146, 147].
The utilization of appropriate nanomaterials and the mitigation of potential adverse effects comprise the essence of nanotechnology. It is vital to acknowledge that the assessment of risks is imperative before authorizing new nano-based products for clinical and commercial utilization, to minimize any potential threats to human health and the environment [148]. Some of the potential drawbacks of nanotechnology in treating shigellosis are given in Table 2. The evaluation of toxicity plays a pivotal role in guaranteeing the safety of nanomedicines and safeguarding public health. Although nanomedicines hold promise in enhancing targeted cellular bioavailability and potency and reducing toxicity, regulatory bodies must comprehend the underlying scientific principles and regulate these products accordingly. Presently, there is a scarcity of testing methodologies available for the assessment of nanotoxicology in clinical translation. Conforming to standard preclinical and clinical protocols is necessary to support the development and approval of nanomedicines, encompassing the evaluation of efficacy, toxicity, and biophysical characteristics. These assessments are pivotal in ensuring the successful introduction of a diverse array of nanodrugs into the market [149,150,151].
Conclusions
Although there is not a vaccination that protects against Shigella at the moment, there are many vaccines that utilize bacterial components or attenuated bacteria, either killed or live, for immunization purposes. Currently, these vaccinations are going through several stages of clinical testing. It is essential to acknowledge that therapeutic interventions aimed at certain microbe components or activities may encounter obstacles due to resistance-causing mutations and selection. Considering this, we ventured into how nanotechnology may be used to create a treatment that effectively combats shigellosis. Nanomaterials might be employed as antibacterial agents, and biosensors could be utilized to deliver vaccinations to immune cells specifically and provide real-time microbiological diagnostics. Nanofiber-based membranes may also be used as wound dressings and scaffolds. The utilization of these tools is crucial in combating the rising prevalence of antimicrobial-resistant Shigella strains. However, it is important to note that nanomedicine, like any medical intervention, must undergo rigorous control and thorough evaluation before it can be used to treat patients to the fullest extent; toxicity assessment and multistage clinical studies must be conducted. The overall aim of this is to provide insight to readers and researchers with an understanding of the role of different Shigella spp. in causing shigellosis in humans, as well as the potential of nanotechnology in addressing this issue.
Availability of data and materials
None.
Abbreviations
- S. dysenteriae :
-
Shigella dysenteriae
- S. flexneri :
-
Shigella flexneri
- S. boydii :
-
Shigella boydii
- S. sonnei :
-
Shigella sonnei
- HUS:
-
Haemolytic uraemic syndrome
- T3SS:
-
Type III secretion systems
- IpaC:
-
Invasion plasmid antigen C
- IpaB:
-
Invasion plasmid antigen B
- ipaD:
-
Invasion plasmid antigen D
- IL-1:
-
Interleukin-1
- IL-6:
-
Interleukin-6
- IFN-γ:
-
Interferon-gamma
- TNF-alpha:
-
Tumour necrosis factor-alpha
- GEMS:
-
Global Enteric Multicenter Study
- PHE:
-
Public Health England
- MAM:
-
Multivalent adhesion molecule
- T6SS:
-
Type VI Secretion System
- G4C:
-
Group 4 capsule
- OAg:
-
O-Antigen
- LPS:
-
Lipoprotein-associated liposomes
- PMN:
-
Polymorphonuclear leukocytes
- H-NS:
-
Histone-like nucleoid structure protein
- DAMPs:
-
Damage-associated molecular patterns
- PAMPs:
-
Pathogen-associated molecular patterns
- LAB:
-
Lactic acid bacteria
- NPs:
-
Nanoparticles
- Ag:
-
Silver
- Bacitracin-AgNCs:
-
Bacitracin-Ag Nanoclusters
- CS-Nps:
-
Chitosan nanoparticles
- IgA:
-
Immunoglobulin A
- IgG:
-
Immunoglobulin B
- OMVs:
-
Outer membrane vesicles
- NVs:
-
Nanovaccines
- ICMR:
-
Indian Council of Medical Research
- BCIL:
-
Biotech Consortium India Limited
- NICED:
-
National Institute of Cholera and Enteric Diseases
- MDR:
-
Multidrug-resistant
- Tet-CPNPs:
-
Tetracycline-embedded calcium-phosphate nanoparticles
- GQDs:
-
Graphene quantum dots
- AuNPs:
-
Gold nanoparticles
- MNPs:
-
Magnetic nanoparticles
- NF:
-
Nanofiber
- ECM:
-
Extracellular matrix
References
Nicolas X, Granier H, Le Guen P (2007) Shigellosis or bacillary dysentery. Presse Med 36(11 Pt 2):1606–1618. https://doi.org/10.1016/j.lpm.2007.03.001
https://wwwnc.cdc.gov/travel/yellowbook/2024/infections-diseases/shigellosis
Tickell KD, Brander RL, Atlas HE, Pernica JM, Walson JL, Pavlinac PB (2017) Identification and management of Shigella infection in children with diarrhoea: a systematic review and meta-analysis. Lancet Glob Health 5(12):e1235–e1248. https://doi.org/10.1016/S2214-109X(17)30392-3
https://www.who.int/teams/immunization-vaccines-and-biologicals/diseases/shigella
Badr HS, Colston JM, Nguyen NLH, Chen YT, Burnett E, Ali SA et al (2023) Spatiotemporal variation in risk of Shigella infection in childhood: a global risk mapping and prediction model using individual participant data. Lancet Glob Health 11(3):e373–e384. https://doi.org/10.1016/S2214-109X(22)00549-6
Jandu N, Goldberg MB (2013) Dysentery. In: The prokaryotes: human microbiology, pp 309–17. https://doi.org/10.1007/978-3-642-30144-5
Bhattacharya SK, Sur D (2017) Shigellosis. In: Second E (ed) Quah SRBTIE of PH. International Encyclopedia of Public Health. Elsevier, Oxford, pp 499–501. https://doi.org/10.1016/B978-0-12-803678-5.00406-9
Prabhurajeshwar C, Chandrakanth KR (2018) Shigellosis: its prevention and management issues. Concepts Dairy Vet Sci 1(5):125–134. https://doi.org/10.32474/cdvs.2018.01.000121
Notes from the Field: Outbreak of Infections Caused by Shigella sonnei with Decreased Susceptibility to Azithromycin—Los Angeles, California (2012). https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6209a4.htm
Kotloff KL, Winickoff JP, Ivanoff B, Clemens JD, Swerdlow DL, Sansonetti PJ et al (1999) Global burden of Shigella infections: implications for vaccine development and implementation of control strategies. Bull World Health Organ 77(8):651–666
He F, Han K, Liu L, Sun W, Zhang L, Zhu B et al (2012) Shigellosis outbreak associated with contaminated well water in a rural elementary school: Sichuan Province, China, June 7–16, 2009. PLoS ONE 7(10):e47239. https://doi.org/10.1371/journal.pone.0047239
Emch M, Ali M, Yunus M (2008) Risk areas and neighborhood-level risk factors for Shigella dysenteriae 1 and Shigella flexneri. Health Place 14(1):96–105. https://doi.org/10.1016/j.healthplace.2007.05.004
Von Seidlein L, Kim DR, Ali M, Lee H, Wang X, Thiem VD et al (2006) A multicentre study of Shigella diarrhoea in six Asian countries: disease burden, clinical manifestations, and microbiology. PLoS Med 3(9):e353. https://doi.org/10.1371/journal.pmed.0030353
Gu B, Cao Y, Pan S, Zhuang L, Yu R, Peng Z et al (2012) Comparison of the prevalence and changing resistance to nalidixic acid and ciprofloxacin of Shigella between Europe-America and Asia-Africa from 1998 to 2009. Int J Antimicrob Agents 40(1):9–17. https://doi.org/10.1016/j.ijantimicag.2012.02.005
Bhattacharya SK, Sur D (2003) An evaluation of current shigellosis treatment. Expert Opin Pharmacother 4(8):1315–1320. https://doi.org/10.1517/14656566.4.8.1315
Hale TL, Keush GT (1996) Chapter-22 Shigella. In: Baron S (ed) Medical microbiology, 4th edn. University of Texas Medical Branch, Galveston, pp 684–741
Ballal M (2016) Chapter 4—trends in antimicrobial resistance among enteric pathogens: a global concern. In: Kon K, Rai MBTAR (eds) Antibiotic resistance: mechanisms and new antimicrobial approaches, pp 63–92. https://doi.org/10.1016/B978-0-12-803642-6.00004-6
Nasser A, Mosadegh M, Azimi T, Shariati A (2022) Molecular mechanisms of Shigella effector proteins: a common pathogen among diarrheic pediatric population. Mol Cell Pediatr 9(1):12. https://doi.org/10.1186/s40348-022-00145-z
Drancourt M (2017) Acute diarrhea. In: Cohen J, Powderly WG (eds) Infectious diseases, 4th edn. Elsevier, Amsterdam, pp 335–340. https://doi.org/10.1016/B978-0-7020-6285-8.00038-1
Mark Taylor C (2008) Enterohaemorrhagic Escherichia coli and Shigella dysenteriae type 1-induced haemolytic uraemic syndrome. Pediatr Nephrol 23(9):1425–1431. https://doi.org/10.1007/s00467-008-0820-3
Nabatanzi M, Kwesiga B, Bulage L, Lubwama B, Ario R, Harris J (2022) Epidemiology of dysentery, Uganda, 2014–2018. J Interv Epidemiol Public Heal 5(4):2014–2018
Weil AA, Harris JB, LaRocque RC (2020) Approach to the patient with diarrhea. In: Ryan ET, Hill DR, Solomon T, Aronson NE, Endy T (eds) Hunter’s tropical medicine and emerging infectious diseases, 10th edn. Elsevier, London, pp 172–177. https://doi.org/10.1016/B978-0-323-55512-8.00022-3
Holmes RK, Jobling MG, Schmitt MP (1999) Phage toxins and disease. In: Granoff A (ed) Encyclopedia of virology, 2nd edn. Elsevier, Amsterdam, pp 1228–1234. https://doi.org/10.1006/rwvi.1999.0025
Baker S, The HC (2018) Recent insights into Shigella: a major contributor to the global diarrhoeal disease burden. Curr Opin Infect Dis 31(5):449–454. https://doi.org/10.1097/QCO.0000000000000475
Bennish ML, Ahmed S (2020) Shigellosis. In: Ryan ET, Hill DR, Solomon T, Aronson NE, Endy T (eds) Hunter’s tropical medicine and emerging infectious diseases. Elsevier, London, pp 492–499. https://doi.org/10.1016/B978-0-323-55512-8.00048-X
Juneja VK, Sehgal S, Singh (2024) Bacteria: Shigella. In: Encyclopedia of Food safety, 2nd edn. Academic Press, London, pp 293–302. https://doi.org/10.1016/B978-0-12-822521-9.00091-5
Taneja N, Mewara A (2016) Shigellosis: epidemiology in India. Indian J Med Res 143(5):565–576. https://doi.org/10.4103/0971-5916.187104
Takeuchi A, Formal SB, Sprinz H (1968) Exerimental acute colitis in the Rhesus monkey following peroral infection with Shigella flexneri. An electron microscope study. Am J Pathol 52(3):503–529
Muthuramalingam M, Whittier SK, Picking WL, Picking WD (2021) The Shigella type III secretion system: an overview from top to bottom. Microorganisms 9(2):1–26. https://doi.org/10.3390/microorganisms9020451
Perdomo OJ, Cavaillon JM, Huerre M, Ohayon H, Gounon P, Sansonetti PJ (1994) Acute inflammation causes epithelial invasion and mucosal destruction in experimental shigellosis. J Exp Med 180(4):1307–1319. https://doi.org/10.1084/jem.180.4.1307
Lingwood CA, Mahfoud R (2010) Bacterial toxins and their carbohydrate receptors at the host–pathogen interface. In: Holst O, Brennan PJ, von Itzstein M, Moran AP (eds) Microbial glycobiology structures, relevance and applications. Academic Press, San Diego, pp 599–621. https://doi.org/10.1016/B978-0-12-374546-0.00030-4
Carayol N, Tran Van Nhieu G (2013) The inside story of Shigella invasion of intestinal epithelial cells. Cold Spring Harb Perspect Med 3(10):a016717. https://doi.org/10.1101/cshperspect.a016717
Brotcke Zumsteg A, Goosmann C, Brinkmann V, Morona R, Zychlinsky A (2014) IcsA is a Shigella flexneri adhesion regulated by the type III secretion system and required for pathogenesis. Cell Host Microbe 15(4):435–445. https://doi.org/10.1016/j.chom.2014.03.001
Thirumalai K, Kim KS, Zychlinsky A (1997) IpaB, a Shigella flexneri invasin, colocalizes with interleukin-1 beta-converting enzyme in the cytoplasm of macrophages. Infect Immun 65(2):787–793. https://doi.org/10.1128/iai.65.2.787-793.1997
Dickenson NE, Choudhari SP, Adam PR, Kramer RM, Joshi SB, Middaugh CR et al (2013) Oligomeric states of the Shigella translocator protein IpaB provide structural insights into formation of the type III secretion translocon. Protein Sci 22(5):614–627. https://doi.org/10.1002/pro.2245
Epler CR, Dickenson NE, Olive AJ, Picking WL, Picking WD (2009) Liposomes recruit IpaC to the Shigella flexneri type III secretion apparatus needle as a final step in secretion induction. Infect Immun 77(7):2754–2761. https://doi.org/10.1128/IAI.00190-09
Blocker A, Gounon P, Larquet E, Niebuhr K, Cabiaux V, Parsot C et al (1999) The tripartite type III secreton of Shigella flexneri inserts IpaB and IpaC into host membranes. J Cell Biol 147(3):683–693. https://doi.org/10.1083/jcb.147.3.683
Arizmendi O, Picking WD, Picking WL (2016) Macrophage apoptosis triggered by IpaD from Shigella flexneri. Infect Immun 84(6):1857–1865. https://doi.org/10.1128/iai.01483-15
Zhu Z, Wang W, Cao M, Zhu Q, Ma T, Zhang Y et al (2021) Virulence factors and molecular characteristics of Shigella flexneri isolated from calves with diarrhea. BMC Microbiol 21(1):214. https://doi.org/10.1186/s12866-021-02277-0
Brotcke Zumsteg A, Goosmann C, Brinkmann V, Morona R, Zychlinsky A (2014) IcsA is a Shigella flexneri adhesin regulated by the type III secretion system and required for pathogenesis. Cell Host Microbe 15(4):435–445. https://doi.org/10.1016/j.chom.2014.03.001
Kany S, Vollrath JT, Relja B (2019) Cytokines in inflammatory disease. Int J Mol Sci 20(23):1–31. https://doi.org/10.3390/ijms20236008
Shigella boydii—microbewiki. https://microbewiki.kenyon.edu/index.php/Shigella_boydii
Girón JA (1995) Expression of flagella and motility by Shigella. Mol Microbiol 18(1):63–75. https://doi.org/10.1111/j.1365-2958.1995.mmi_18010063.x
Tominaga A, Mahmoud MAH, Al Mamun AAM, Mukaihara T (2001) Characterization of cryptic flagellin genes in Shigella boydii and Shigella dysenteriae. Genes Genet Syst 76(2):111–120. https://doi.org/10.1266/ggs.76.111
Akter M, Brown N, Clokie M, Yeasmin M, Tareq TM, Baddam R et al (2019) Prevalence of Shigella boydii in Bangladesh: isolation and characterization of a rare phage MK-13 that can robustly identify shigellosis caused by Shigella boydii type 1. Front Microbiol 10(2461):1–11. https://doi.org/10.3389/fmicb.2019.02461
Labbé R, García S (2013) Guide to foodborne pathogens. Wiley, New York, pp 1–488. https://doi.org/10.1002/9781118684856
Hamer DH, Gill CJ, Chilengi R (2017) Intestinal infections: overview. In: Reference module in biomedical sciences: international encyclopedia of public health, 2nd edn. Academic Press, London, pp 322–335. https://doi.org/10.1016/B978-0-12-803678-5.00240-X
Livio S, Strockbine NA, Panchalingam S, Tennant SM, Barry EM, Marohn ME et al (2014) Shigella isolates from the global enteric multicenter study inform vaccine development. Clin Infect Dis an Off Publ Infect Dis Soc Am 59(7):933–941. https://doi.org/10.1093/cid/ciu468
Woodward DL, Clark CG, Caldeira RA, Ahmed R, Soule G, Bryden L et al (2005) Identification and characterization of Shigella boydii 20 serovar nov., a new and emerging Shigella serotype. J Med Microbiol 54(8):741–748. https://doi.org/10.1099/jmm.0.46095-0
Todkill D, Pudney R, Terrell A, Tuck J, Hutley E, Dallman TJ et al (2018) An outbreak of Shigella boydii serotype 20 in January 2015 amongst United Kingdom healthcare workers involved in the Ebola response in Sierra Leone. J Med Microbiol 67(11):1596–1600. https://doi.org/10.1099/jmm.0.000832
Todd ECD (2014) Foodborne Diseases: overview of biological hazards and foodborne diseases. In: Fahey JW (ed) Reference module in food science: encyclopedia of food safety. Academic Press, Waltham, pp 221–242. https://doi.org/10.1016/B978-0-12-378612-8.00071-8
Torraca V, Holt K, Mostowy S (2020) Shigella sonnei. Trends Microbiol 28(8):696–697. https://doi.org/10.1016/j.tim.2020.02.011
Thompson CN, Duy PT, Baker S (2015) The rising dominance of Shigella sonnei: an intercontinental shift in the etiology of bacillary dysentery. PLoS Negl Trop Dis 9(6):1–13. https://doi.org/10.1371/journal.pntd.0003708
Mahmoud RY, Stones DH, Li W, Emara M, El-domany RA, Wang D et al (2016) The multivalent adhesion molecule SSO1327 plays a key role in Shigella sonnei pathogenesis. Mol Microbiol 99(4):658–673. https://doi.org/10.1111/mmi.13255
Zhu Z, Shi Y, Zhou X, Li B, Zhang J (2018) Molecular characterization of fluoroquinolone and/or cephalosporin resistance in Shigella sonnei isolates from yaks. BMC Vet Res 14(1):1–9. https://doi.org/10.1186/s12917-018-1500-6
Nandy S, Dutta S, Ghosh S, Ganai A, Rajahamsan J, Theodore RB et al (2011) Foodborne associated Shigella sonnei, India, 2009 and 2010. Emerg Infect Dis 17(11):2072–2074. https://doi.org/10.3201/eid1502.080988
Filliol-Toutain I, Chiou CS, Mammina C, Gerner-Smidt P, Thong KL, Phung DC et al (2011) Global distribution of Shigella sonnei clones. Emerg Infect Dis 17(10):1910–1912. https://doi.org/10.3201/eid1710.101486
Kozyreva VK, Jospin G, Greninger AL, Watt JP, Eisen JA, Chaturvedi V (2016) Recent outbreaks of shigellosis in california caused by two distinct populations of Shigella sonnei with either increased virulence or fluoroquinolone resistance. mSphere 1(6):e00344-16. https://doi.org/10.1128/mSphere.00344-16
Shad AA, Shad WA (2021) Shigella sonnei: virulence and antibiotic resistance. Arch Microbiol 203(1):45–58. https://doi.org/10.1007/s00203-020-02034-3
Anderson MC, Vonaesch P, Saffarian A, Marteyn BS, Sansonetti PJ (2017) Shigella sonnei encodes a functional T6SS used for interbacterial competition and niche occupancy. Cell Host Microbe 21(6):769-776.e3. https://doi.org/10.1016/j.chom.2017.05.004
Allsopp LP, Bernal P, Nolan LM, Filloux A (2020) Causalities of war: the connection between type VI secretion system and microbiota. Cell Microbiol 22(3):e13153. https://doi.org/10.1111/cmi.13153
Holt KE, Thieu Nga TV, Thanh DP, Vinh H, Kim DW, Vu Tra MP et al (2013) Tracking the establishment of local endemic populations of an emergent enteric pathogen. Proc Natl Acad Sci U S A 110(43):17522–17527. https://doi.org/10.1073/pnas.1308632110
Caboni M, Pédron T, Rossi O, Goulding D, Pickard D, Citiulo F et al (2015) An O antigen capsule modulates bacterial pathogenesis in Shigella sonnei. PLoS Pathog 11(3):e1004749. https://doi.org/10.1371/journal.ppat.1004749
Rendueles O, Garcia-Garcerà M, Néron B, Touchon M, Rocha EPC (2017) Abundance and co-occurrence of extracellular capsules increase environmental breadth: implications for the emergence of pathogens. PLoS Pathog 13(7):e1006525. https://doi.org/10.1371/journal.ppat.1006525
Turnbaugh PJ, Ley RE, Hamady M, Fraser-Liggett CM, Knight R, Gordon JI (2007) The human microbiome project. Nature 449(7164):804–810. https://doi.org/10.1038/nature06244
Jandhyala SM, Talukdar R, Subramanyam C, Vuyyuru H, Sasikala M, Nageshwar Reddy D (2015) Role of the normal gut microbiota. World J Gastroenterol 21(29):8787–8803. https://doi.org/10.3748/wjg.v21.i29.8787
Marteyn B, West NP, Browning DF, Cole JA, Shaw JG, Palm F et al (2010) Modulation of Shigella virulence in response to available oxygen in vivo. Nature 465(7296):355–358. https://doi.org/10.1038/nature08970
Shi R, Yang X, Chen L, Chang H, Liu H, Zhao J et al (2014) Pathogenicity of Shigella in chickens. PLoS ONE 9(6):e100264. https://doi.org/10.1371/journal.pone.0100264
Haider K, Hossain A, Wanke C, Qadri F, Ali S, Nahar S (1993) Production of mucinase and neuraminidase and binding of Shigella to Intestinal mucin. J Diarrhoeal Dis Res 11(2):88–92
Phantouamath B, Sithivong N, Insisiengmay S, Ichinose Y, Higa N, Song T et al (2005) Pathogenicity of Shigella in healthy carriers: a study in Vientiane, Lao People’s Democratic Republic. Jpn J Infect Dis 58(4):232–234
Ashida H, Sasakawa C (2015) Shigella IpaH family effectors as a versatile model for studying pathogenic bacteria. Front Cell Infect Microbiol 6(5):100. https://doi.org/10.3389/fcimb.2015.00100
Schnupf P, Sansonetti PJ (2019) Shigella pathogenesis: new insights through advanced methodologies. Microbiol Spectr 7(2):1–24. https://doi.org/10.1128/microbiolspec.bai-0023-2019
Büttner D, Bonas U (2006) Who comes first? How plant pathogenic bacteria orchestrate type III secretion. Curr Opin Microbiol 9(2):193–200. https://doi.org/10.1016/j.mib.2006.02.006
Faruque SM, Khan R, Kamruzzaman M, Yamasaki S, Ahmad QS, Azim T et al (2002) Isolation of Shigella dysenteriae type 1 and S. flexneri strains from surface waters in Bangladesh: comparative molecular analysis of environmental Shigella isolates versus clinical strains. Appl Environ Microbiol 68(8):3908–3913. https://doi.org/10.1128/AEM.68.8.3908-3913.2002
Marteyn B, Gazi A, Sansonetti P (2012) Shigella: a model of virulence regulation in vivo. Gut Microbes 3(2):104–120. https://doi.org/10.4161/gmic.19325
Bajunaid W, Haidar-Ahmad N, Kottarampatel AH, Ourida Manigat F, Silué N, Tchagang CF et al (2020) The T3SS of Shigella: expression, structure, function, and role in vacuole escape. Microorganisms 8(12):1933. https://doi.org/10.3390/microorganisms8121933
Bohn E, Sonnabend M, Klein K, Autenrieth IB (2019) Bacterial adhesion and host cell factors leading to effector protein injection by type III secretion system. Int J Med Microbiol 309(5):344–350. https://doi.org/10.1016/j.ijmm.2019.05.008
Puhar A, Sansonetti PJ (2014) Type III secretion system. Curr Biol 24(17):R784–R791. https://doi.org/10.1016/j.cub.2014.07.016
Killackey SA, Sorbara MT, Girardin SE (2016) Cellular aspects of Shigella pathogenesis: focus on the manipulation of host cell processes. Front Cell Infect Microbiol 6:1–11. https://doi.org/10.3389/fcimb.2016.00038
Ndungo E, Holm JB, Gama S, Buchwald AG, Tennant SM, Laufer MK et al (2022) Dynamics of the gut microbiome in Shigella-infected children during the first two years of life. mSystems 7(5):e0044222. https://doi.org/10.1128/msystems.00442-22
Ashida H, Mimuro H, Sasakawa C (2015) Shigella manipulates host immune responses by delivering effector proteins with specific roles. Front Immunol 6:219. https://doi.org/10.3389/fimmu.2015.00219
Suryavanshi MV, Bhute SS, Jadhav SD, Bhatia MS, Gune RP, Shouche YS (2016) Hyperoxaluria leads to dysbiosis and drives selective enrichment of oxalate metabolizing bacterial species in recurrent kidney stone endures. Sci Rep 6:34712. https://doi.org/10.1038/srep34712
Ogunrinola GA, Oyewale JO, Oshamika OO, Olasehinde GI (2020) The human microbiome and its impacts on health. Int J Microbiol 2020:8045646. https://doi.org/10.1155/2020/8045646
Artemev A, Naik S, Pougno A, Honnavar P, Shanbhag NM (2022) The association of microbiome dysbiosis with colorectal cancer. Cureus 14(2):e22156. https://doi.org/10.7759/cureus.22156
Köseoğlu VK, Hall CP, Rodríguez-López EM, Agaisse H (2019) The autotransporter IcsA promotes Shigella flexneri biofilm formation in the presence of bile salts. Infect Immun 87(7):e00861-e918. https://doi.org/10.1128/IAI.00861-18
Nickerson KP, Chanin RB, Sistrunk JR, Rasko DA, Fink PJ, Barry EM, Nataro JP, Faherty CS (2017) Analysis of Shigella flexneri resistance, biofilm formation, and transcriptional profile in response to bile salts. Infect Immun 85(6):e01067-e1116. https://doi.org/10.1128/IAI.01067-16
Yang J, Chen W, Xia P, Zhang W (2020) Dynamic comparison of gut microbiota of mice infected with Shigella flexneri via two different infective routes. Exp Ther Med 19(3):2273–2281. https://doi.org/10.3892/etm.2020.8469
Liu Q, Yu Z, Tian F, Zhao J, Zhang H, Zhai Q, Chen W (2020) Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb Cell Fact 19(23):1–11. https://doi.org/10.1186/s12934-020-1289-4
Kutshik RJ, Longdet IY, Egbere O, Atuluku VA (2012) Lactobacillus casei isolates from kunun zaki: the probiotic potential against induced shigellosis in experimental rats. J Med Appl Biosci 4:46–53
Filho-Lima JV, Vieira EC, Nicoli JR (2000) Antagonistic effect of Lactobacillus acidophilus, Saccharomyces boulardii and Escherichia coli combinations against experimental infections with Shigella flexneri and Salmonella enteritidis subsp. typhimurium in gnotobiotic mice. J Appl Microbiol 88(3):365–370. https://doi.org/10.1046/j.1365-2672.2000.00973.x
Levy M, Kolodziejczyk AA, Thaiss CA, Elinav E (2017) Dysbiosis and the immune system. Nat Rev Immunol 17(4):219–232. https://doi.org/10.1038/nri.2017.7
Humphreys C (2020) Intestinal permeability. In: Pizzorno JE, Murray MT (eds) Textbook of natural medicine, 5th edn. Churchill Livingstone, St. Louis, pp 166–177
Anderson M, Sansonetti PJ, Marteyn BS (2016) Shigella diversity and changing landscape: insights for the twenty-first century. Front Cell Infect Microbiol 6(45):1–9. https://doi.org/10.3389/fcimb.2016.00045
Khan MF, Wang H (2020) Environmental exposures and autoimmune diseases: contribution of gut microbiome. Front Immunol 10(3094):1–11. https://doi.org/10.3389/fimmu.2019.03094
Fiorentino M, Levine MM, Sztein MB, Fasano A (2014) Effect of wild-type Shigella species and attenuated Shigella vaccine candidates on small intestinal barrier function, antigen trafficking, and cytokine release. PLoS ONE 9(1):e85211. https://doi.org/10.1371/journal.pone.0085211
Weiss GA, Hennet T (2017) Mechanisms and consequences of intestinal dysbiosis. Cell Mol Life Sci 74(16):2959–2977. https://doi.org/10.1007/s00018-017-2509-x
Ducarmon QR, Zwittink RD, Hornung BVH, van Schaik W, Young VB, Kuijper EJ (2019) Gut microbiota and colonization resistance against bacterial enteric infection. Microbiol Mol Biol Rev 83(3):e00007-19. https://doi.org/10.1128/MMBR.00007-19
DeGruttola AK, Low D, Mizoguchi A, Mizoguchi E (2016) Current understanding of dysbiosis in disease in human and animal models. Inflamm Bowel Dis 22(5):1137–1150. https://doi.org/10.1097/MIB.0000000000000750
Anand U, Carpena M, Kowalska-Góralska M, Garcia-Perez P, Sunita K, Bontempi E, Dey A, Prieto MA, Prockow J, Simal-Gandara J (2022) Safer plant-based nanoparticles for combating antibiotic resistance in bacteria: a comprehensive review on its potential applications, recent advances, and future perspective. Sci Total Environ 821:153472. https://doi.org/10.1016/j.scitotenv.2022.153472
Huang Y, Guo X, Wu Y, Chen X, Feng L, Xie N (2024) Nanotechnology’s frontier in combatting infectious and inflammatory diseases: prevention and treatment. Signal Transduct Target Ther 9(1):1–50. https://doi.org/10.1038/s41392-024-01745-z
Chakraborty N, Jha D, Roy I, Kumar P, Gaurav SS, Marimuthu K, Ng OT, Lakshminarayanan R, Verma NK, Gautam HK (2022) Nanobiotics against antimicrobial resistance: harnessing the power of nanoscale materials and technologies. J Nanobiotechnology 20(1):375. https://doi.org/10.1186/s12951-022-01573-9
Ivask A, Elbadawy A, Kaweeteerawat C, Boren D, Fischer H, Ji Z, Chang CH, Liu R, Tolaymat T, Telesca D, Zink JI, Cohen Y, Holden PA, Godwin HA (2014) Toxicity mechanisms in Escherichia coli vary for silver nanoparticles and differ from ionic silver. ACS Nano 8(1):374–386. https://doi.org/10.1021/nn4044047
Bruna T, Maldonado-Bravo F, Jara P, Caro N (2021) Silver nanoparticles and their antibacterial applications. Int J Mol Sci 22(13):7202. https://doi.org/10.3390/ijms22137202
Anees Ahmad S, Sachi Das S, Khatoon A, Tahir Ansari M, Afzal M, Saquib Hasnain M, Nayak AK (2020) Bactericidal activity of silver nanoparticles: a mechanistic review. Mater Sci Energy Technol 3:756–769. https://doi.org/10.1016/j.mset.2020.09.002
Seong M, Lee DG (2017) Silver nanoparticles against Salmonella enterica serotype typhimurium: role of inner membrane dysfunction. Curr Microbiol 74(6):661–670. https://doi.org/10.1007/s00284-017-1235-9
Seil JT, Webster TJ (2012) Antimicrobial applications of nanotechnology: methods and literature. Int J Nanomed 7:2767–2781. https://doi.org/10.2147/IJN.S24805
Li WR, Xie XB, Shi QS, Zeng HY, Ou-Yang YS, Chen YB (2010) Antibacterial activity and mechanism of silver nanoparticles on Escherichia coli. Appl Microbiol Biotechnol 85(4):1115–1122. https://doi.org/10.1007/s00253-009-2159-5
Sondi I, Salopek-Sondi B (2004) Silver nanoparticles as antimicrobial agent: a case study on E. coli as a model for Gram-negative bacteria. J Colloid Interface Sci 275(1):177–182. https://doi.org/10.1016/j.jcis.2004.02.012
Quinteros MA, Cano Aristizábal V, Dalmasso PR, Paraje MG, Páez PL (2016) Oxidative stress generation of silver nanoparticles in three bacterial genera and its relationship with the antimicrobial activity. Toxicol In Vitro 36:216–223. https://doi.org/10.1016/j.tiv.2016.08.007
Agnihotri S, Mukherji S, Mukherji S (2013) Immobilized silver nanoparticles enhance contact killing and show highest efficacy: elucidation of the mechanism of bactericidal action of silver. Nanoscale 5(16):7328–7340. https://doi.org/10.1039/C3NR00024A
Gurunathan S, Han JW, Kwon DN, Kim JH (2014) Enhanced antibacterial and anti-biofilm activities of silver nanoparticles against Gram-negative and Gram-positive bacteria. Nanoscale Res Lett 9(1):373. https://doi.org/10.1186/1556-276X-9-373
Wang L, Liu L, Zhou X (2021) Bacitracin-Ag nanoclusters as a novel antibacterial agent combats Shigella flexneri by disrupting cell membrane and inhibiting biofilm formation. Nanomaterials 11(11):2928. https://doi.org/10.3390/nano11112928
Lugade AA, Bharali DJ, Pradhan V, Elkin G, Mousa SA, Thanavala Y (2013) Single low-dose un-adjuvanted HBsAg nanoparticle vaccine elicits robust, durable immunity. Nanomedicine 9(7):923–934. https://doi.org/10.1016/j.nano.2013.03.008
Diaz-Arévalo D, Zeng M (2020) Nanoparticle-based vaccines: opportunities and limitations. Nanopharmaceuticals. https://doi.org/10.1016/B978-0-12-817778-5.00007-5
Gilavand F, Marzban A, Ebrahimipour G, Soleimani N, Goudarzi M (2020) Designation of chitosan nano-vaccine based on MxiH antigen of Shigella flexneri with increased immunization capacity. Carbohydr Polym 15(232):115813. https://doi.org/10.1016/j.carbpol.2019.115813
Kaveti B (2022) Single dose nanovaccine developed to help tackle Shigella. https://www.azonano.com/news.aspx?newsID=39715
Koley H, Bhaumik U, Baruah N, Mitra S, Maiti S, Nag D (2023) A story of Shigella vaccine development in ICMR-NICED involving multidimensional approaches. Indian J Physiol Allied Sci 75(2):58–75. https://doi.org/10.55184/ijpas.v75i02.08
Baruah N, Ahamad N, Halder P, Koley H, Katti DS (2023) Facile synthesis of multi-faceted, biomimetic and cross-protective nanoparticle-based vaccines for drug-resistant Shigella: a flexible platform technology. J Nanobiotechnol 21(1):34. https://doi.org/10.1186/s12951-023-01780-y
2019. ICMR distributes a license for Shigella vaccine. https://main.icmr.nic.in/content/icmr-distributes-license-shigella-vaccine-april2019
Resistance D, Ranjbar R, Farahani A (2019) Shigella: antibiotic-resistance mechanisms and new horizons for treatment. Infect Drug Resist 12:3137–3167. https://doi.org/10.2147/IDR.S219755
Zong TX, Silveira AP, Morais JAV, Sampaio MC, Muehlmann LA, Zhang J, Jiang CS, Liu SK (2022) Recent advances in antimicrobial nano-drug delivery systems. Nanomater (Basel, Switzerland) 12(11):1855. https://doi.org/10.3390/nano12111855
Gao W, Thamphiwatana S, Angsantikul P, Zhang L (2014) Nanoparticle approaches against bacterial infections. Wiley Interdiscip Rev Nanomed Nanobiotechnol 6(6):532–547. https://doi.org/10.1002/wnan.1282
Gao W, Chen Y, Zhang Y, Zhang Q, Zhang L (2018) Nanoparticle-based local antimicrobial drug delivery. Adv Drug Deliv Rev 127:46–57. https://doi.org/10.1016/j.addr.2017.09.015
Mukherjee R, Dutta D, Patra M, Chatterjee B, Basu T (2019) Nanonized tetracycline cures deadly diarrheal disease ‘shigellosis’ in mice, caused by multidrug-resistant Shigella flexneri 2a bacterial infection. Nanomed Nanotechnol Biol Med 18:402–413. https://doi.org/10.1016/j.nano.2018.11.004
El-Sherbiny IM, El-Baz NM, Yacoub MH (2015) Inhaled nano- and microparticles for drug delivery. Glob Cardiol Sci Pract 1(2):1–14. https://doi.org/10.5339/gcsp.2015.2
Yusuf A, Almotairy ARZ, Henidi H, Alshehri OY, Aldughaim MS (2023) Nanoparticles as drug delivery systems: a review of the implication of nanoparticles’ physicochemical properties on responses in biological systems. Polymers 15(7):1596. https://doi.org/10.3390/polym15071596
Bhattacharjee R, Negi A, Bhattacharya B, Dey T, Mitra P, Preetam S, Kumar L, Kar S, Das SS, Iqbal D, Kamal M, Alghofaili F, Malik S, Dey A, Jha SK, Ojha S, Paiva-Santos AC, Kesari KK, Jha NK (2023) Nanotheranostics to target antibiotic-resistant bacteria: strategies and applications. OpenNano 11:100138. https://doi.org/10.1016/j.onano.2023.100138
Yeh YC, Huang TH, Yang SC, Chen CC, Fang JY (2020) Nano-based drug delivery or targeting to eradicate bacteria for infection mitigation: a review of recent advances. Front Chem 8:286. https://doi.org/10.3389/fchem.2020.00286
Castillo-Henríquez L, Brenes-Acuña M, Castro-Rojas A, Cordero-Salmerón R, Lopretti-Correa M, Vega-Baudrit JR (2020) Biosensors for the detection of bacterial and viral clinical pathogens. Sensors 23:6926
Ramesh M, Janani R, Deepa C, Rajeshkumar L (2022) Nanotechnology-enabled biosensors: a review of fundamentals, design principles, materials, and applications. Biosensors 13(1):40. https://doi.org/10.3389/fchem.2020.00286
Choi C (2012) Integrated nanobiosensor technology for biomedical application. Nanobiosens Dis Diagn 2012(1):1–4. https://doi.org/10.2147/NDD.S26422
Elahi N, Kamali M, Baghersad MH, Amini B (2019) A fluorescence nano-biosensors immobilization on Iron (MNPs) and gold (AuNPs) nanoparticles for detection of Shigella spp. Mater Sci Eng C 105:110113. https://doi.org/10.1016/j.msec.2019.110113
Ali MR, Bacchu MS, Das S, Akter S, Rahman MM, Saad Aly MA (2023) Label free flexible electrochemical DNA biosensor for selective detection of Shigella flexneri in real food samples. Talanta 253:123909. https://doi.org/10.1016/j.talanta.2022.123909
Abrigo M, McArthur SL, Kingshott P (2014) Electrospun nanofibers as dressings for chronic wound care: advances, challenges, and future prospects. Macromol Biosci 14(6):772–792. https://doi.org/10.1002/mabi.201300561
Nunes CS, Philipps-Wiemann P (2018) Formulation of enzymes. In: Nunes CS, Kumar V (eds) Enzymes in human and animal nutrition: principles and perspectives. Academic Press, London, pp 429–440
Ghajarieh A, Habibi S, Talebian A (2021) Biomedical applications of nanofibers. Russ J Appl Chem 94(7):847–872. https://doi.org/10.1134/S1070427221070016
Homaeigohar S, Boccaccini AR (2020) Antibacterial biohybrid nanofibers for wound dressings. Acta Biomater 107:25–49. https://doi.org/10.1016/j.actbio.2020.02.022
Park JM, Kim M, Park HS, Jang A, Min J, Kim YH (2013) Immobilization of lysozyme-CLEA onto electrospun chitosan nanofiber for effective antibacterial applications. Int J Biol Macromol 54:37–43. https://doi.org/10.1016/j.ijbiomac.2012.11.025
Samadi S, Moradkhani M, Beheshti H, Irani M, Aliabadi M (2018) Fabrication of chitosan/poly(lactic acid)/graphene oxide/TiO(2) composite nanofibrous scaffolds for sustained delivery of doxorubicin and treatment of lung cancer. Int J Biol Macromol 110:416–424. https://doi.org/10.1016/j.ijbiomac.2017.08.048
Torres-Martinez EJ, Cornejo Bravo JM, Serrano Medina A, Pérez González GL, Villarreal Gómez LJ (2018) A summary of electrospun nanofibers as drug delivery system: drugs loaded and biopolymers used as matrices. Curr Drug Deliv 15(10):1360–1374. https://doi.org/10.2174/1567201815666180723114326
Jiang Z, Zheng Z, Yu S, Gao Y, Ma J, Huang L, Lei Y (2023) Nanofiber scaffolds as drug delivery systems promoting wound healing. Pharmaceutics 15(7):1829. https://doi.org/10.3390/pharmaceutics15071829
Kim Y, Ko H, Kwon IK, Shin K (2016) Extracellular matrix revisited: roles in tissue engineering. Int Neurourol J 20(Suppl 1):S23-29. https://doi.org/10.5213/inj.1632600.318
Zhang X, Chen X, Hong H, Hu R, Liu J, Liu C (2022) Decellularized extracellular matrix scaffolds: recent trends and emerging strategies in tissue engineering. Bioact Mater 10:15–31. https://doi.org/10.1016/j.bioactmat.2021.09.014
Hussey GS, Cramer MC, Badylak SF (2018) Extracellular matrix bioscaffolds for building gastrointestinal tissue. Cell Mol Gastroenterol Hepatol 5(1):1–13. https://doi.org/10.1016/j.jcmgh.2017.09.004
Nikolova MP, Chavali MS (2019) Recent advances in biomaterials for 3D scaffolds: a review. Bioact Mater 4:271–292. https://doi.org/10.1016/j.bioactmat.2019.10.005
Mojaveri SJ, Hosseini SF, Gharsallaoui A (2020) Viability improvement of Bifidobacterium animalis Bb12 by encapsulation in chitosan/poly (vinyl alcohol) hybrid electrospun fiber mats. Carbohydr Polym 241:116278. https://doi.org/10.1016/j.carbpol.2020.116278
Latif A, Shehzad A, Niazi S, Zahid A, Ashraf W, Iqbal MW, Rehman A, Riaz T, Aadil RM, Khan IM, Ozogul F, Rocha JM, Esatbeyoglu T, Korma SA (2023) Probiotics: mechanism of action, health benefits and their application in food industries. Front Microbiol 14:1–15. https://doi.org/10.3389/fmicb.2023.1216674
Sim S, Wong NK (2021) Nanotechnology and its use in imaging and drug delivery (Review). Biomed Rep 14(5):42. https://doi.org/10.3892/br.2021.1418
Bansal V, Katiyar SS, Dora CP (2021) Toxicity aspects: crucial obstacles to clinical translation of nanomedicines. In: Pardeshi CV, Souto E (eds) Direct nose-to-brain drug delivery. Elsevier, Amsterdam, pp 485–494
Manandhar S, Sankhe R, Harish Kumar B, Pai KSR (2021) Clinical protocols for nanopharmacokinetics assessment. In: Thorat ND, Kumar N (eds) Nano-pharmacokinetics and theranostics. Elsevier, Amsterdam, pp 193–220
Ahmad A, Imran M, Sharma N (2022) Precision nanotoxicology in drug development: current trends and challenges in safety and toxicity implications of customized multifunctional nanocarriers for drug-delivery applications. Pharmaceutics 14(11):2463. https://doi.org/10.3390/pharmaceutics14112463
Soliman S (2023) Nanomedicine: advantages and disadvantages of nanomedicine. Rev J Nanomed Nanotechnol 14(3):1–2. https://doi.org/10.35248/2157-7439.23.14.666
Stater EP, Sonay AY, Hart C, Grimm J (2021) The ancillary effects of nanoparticles and their implications for nanomedicine. Nat Nanotechnol 16(11):1180–1194. https://doi.org/10.1038/s41565-021-01017-9
A matter of scale. Nat Nanotechnol 11(9):733. https://doi.org/10.1038/nnano.2016.180
Akhter MH, Khalilullah H, Gupta M, Alfaleh MA, Alhakamy NA, Riadi Y, Md S (2021) Impact of protein corona on the biological identity of nanomedicine: understanding the fate of nanomaterials in the biological milieu. Biomedicines 9(10):1496. https://doi.org/10.3390/biomedicines9101496
Xuan L, Ju Z, Skonieczna M, Zhou PK, Huang R (2023) Nanoparticles-induced potential toxicity on human health: applications, toxicity mechanisms, and evaluation models. MedComm 4(4):e327. https://doi.org/10.1002/mco2.327
Acknowledgements
El Bethel Lalthavel Hmar expresses gratitude to the University Grant Commission and Ministry of Tribal Affairs, Government of India, for awarding UGC-JRF fellowship (Award no. 202223-NFST-ASS-00796) supporting his Ph.D. project.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
Dr. H.K Sharma conceptualized the idea, synthesized the review technique, and reviewed the manuscript. El Bethel Lalthavel Hmar and Sujata Paul contributed to the content significantly and edited the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
None.
Consent for publication
We, the undersigned authors of this article, formally agree to the publications of our work in this reputed journal.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hmar, E.B.L., Paul, S. & Sharma, H.K. The role of Shigella spp. in propagating bacillary dysentery in humans and the prominence of nanotechnology in disease prevention. Futur J Pharm Sci 10, 97 (2024). https://doi.org/10.1186/s43094-024-00676-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43094-024-00676-4