
Abstract
Background
Acute lymphoblastic leukemia (ALL) is the most common childhood cancer. Glutathione-S-methyl transferase (GSTs) enzymes’ family is known to catalyze carcinogens detoxification. Overexpression of (GSTO) omega class was reported in cancer occurrence. The purpose of the study was to investigate the association of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms with the susceptibility to childhood ALL and to evaluate their prognostic impact. Genotyping was performed using polymerase chain reaction-restriction fragment length polymorphism technique in 96 Egyptian pediatric ALL patients and 99 controls.
Results
No statistically significant different GSTO1*A140D genotype and allele distribution was observed among ALL cases and controls; however, a statistically significant different GSTO1*A140D genotype distribution was found between de novo ALL cases and controls [CC (37% vs. 56.6%), CA (47.8% vs. 40.4%), and AA (15.2% vs. 3.0%), respectively] (0.008). GSTO1*A140D variant genotypes’ frequency was significantly higher in de novo cases than in controls (63% vs. 43.4%) (0.028). The minor allele frequency (MAF) of GSTO1*A140D-A was significantly higher in de novo cases compared to controls (0.39 vs. 0.23) (0.005). Genotyping of GSTO2*N142D revealed a statistically significant difference of genotype distribution between ALL patients and controls [AA (26% vs. 36.3%), AG (62.5% vs. 61.6%), and GG (11.4% vs. 2.0%), respectively] (0.017) and between de novo ALL cases and controls [AA (37% vs. 36.3%), AG (45.7% vs. 61.6%), and GG (17.3% vs. 2.0%), respectively] (0.002). The MAF of GSTO2*N142D-G was significantly higher in ALL patients than in controls (0.42 vs. 0.32) (0.046). The high-risk ALL group had a higher frequency of GSTO1*A140D and GSTO2*N142D variant genotypes compared to corresponding wild genotypes and a higher frequency of combined polymorphisms compared to single polymorphisms and wild genotypes but with no statistically significant difference.
Conclusion
A statistically significant difference of GSTO1*A140D and GSTO2*N142D genotype distribution was detected between de novo ALL cases and controls. Compared to the control group, the MAF of GSTO1*A140D-A was overexpressed in de novo ALL cases and that of GSTO2*N142D-G was significantly higher in ALL patients. These findings suggest that the studied polymorphisms might play a significant role in the susceptibility to de novo childhood ALL in Egypt; however, GSTO1*A140D and/or GSTO2*N142D polymorphisms have no impact on ALL prognosis.
Similar content being viewed by others

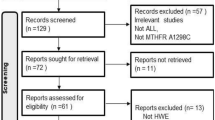
Background
Acute lymphoblastic leukemia (ALL) is the most common type of childhood cancer. Although overall long-term disease-free survival has improved to more than 70% with modern chemotherapy [1], the exact molecular mechanism of this disease remains unknown. The interaction of several environmental and genetic factors has been implicated in cancer occurrence [2]. Moreover, children are exposed to toxic environmental substances in the prenatal period where cell differentiation and growth occur. Upon exposure to drugs or toxic substances, these compounds are metabolized to non-harmful molecules by a biphasic mechanism involving drug detoxification enzymes. These enzymes are essential for the elimination of these toxic substances [3]. They are divided into phase I enzymes, represented by cytochrome P450 isoenzymes (CYP) that catalyze hydrolysis, reduction, and oxidation reactions, and phase II enzymes, which catalyze conjugation reactions, including the glutathione S-transferases (GST) and the N-acetyl transferases (NAT) [4].
Glutathione S-transferases (GSTs) are categorized as phase II drug detoxification enzymes that induce the conjugation of glutathione (GSH) using a number of endogenous or exogenous substances. The genes encoding GSTs were categorized into eight classes: α (GSTA), κ (GSTK), μ (GSTM), ω (GSTO), π (GSTP), ς (GSTS), θ (GSTT), and ζ (GSTZ) [5]. This classification was based on the immunological properties, catalytic activity, amino acid sequence homology, the sequence of bases in genes, and the location of genes on chromosomes [6]. These enzymes have an influence on cell proliferation, apoptosis, neoplastic transformation, tumor metastasis, and development of drug resistance [7]. They are essential for cell protection against cellular stress and toxic compounds [8]. Most often, the polymorphisms that reduce GSTs activity are associated with increased cancer risk and varied response to therapy [9]. Moreover, due to their high level in the plasma from cancer patients and their high expression in tumors compared to normal tissues, these enzymes were identified as cancer markers [10]. For example, GSTM1 and GSTP1 gene polymorphism were known to increase the risk of breast cancer and hepatocellular carcinoma [6]. GSTP1 gene polymorphism and its combination with the GSTM1 and GSTT1 polymorphisms predispose to bladder cancer [11]. Deletion of both alleles of GSTT1 predisposes to lung cancer in Asians [12].
Glutathione S-transferases omega class (GSTO) is encoded by two genes, GSTO1 and GSTO2, located on chromosome 10 (10q24.3) [13]. Unlike other GSTs, it includes an active site cysteine which forms a disulfide bond with GSH and displays glutathione-dependent activities [14]. The widespread distribution of GSTO in normal tissues such as the liver, colon, heart, ovary, pancreas, prostate, and spleen indicates its significant biological role [15]. GSTO participates in cellular signaling [16] and was shown to activate the pro-inflammatory cytokine, interleukin-1b (IL-1b) by post-translational processing [17]. GSTO1 utilizes glutathione in reactions and contributes to the biotransformation and disposition of several drugs, carcinogens, and products of oxidative stress [18]. Reduced thiol-transferase activity may result in defective protection against cellular oxidative stresses [19, 20].
Several studies investigating the association of GSTO gene polymorphism and cancer risk revealed the linkage between GSTO1*A140D and increased risk of breast cancer, hepatocellular carcinoma, cholangiocarcinoma, urothelial cancer, non-small cell lung cancer, and ALL [21]. GSTO2 polymorphism has been identified in ovarian cancer [22, 23]. No evidence of an association of GSTO polymorphism with the increased risk of breast, thyroid, and colon cancer was detected [24]. Previous studies had demonstrated an ethnic variability in GSTO1 and GSTO2 polymorphisms [25,26,27].
This evidence had led us to hypothesize that genetic variation at GSTO1 and GSTO2 genes has been implicated in the occurrence of ALL. In the current study, we aimed to examine the frequency of different genotypes of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms in the Egyptian pediatric ALL patients, to study the possible association of the variant genotypes with the susceptibility to childhood ALL and to evaluate their prognostic implications.
Methods
The study population
This study was conducted on 96 patients diagnosed as ALL. The ALL population was a selection of all patients attending the pediatric hematology clinic, New Children Hospital, Cairo University (NCHCU), during the period from April 2015 to October 2017. Selected patients were Egyptian ALL patients of the pediatric age group. The patients’ ages ranged from 1.5 and 14 years with a mean of 6.95 ± 3.01 years and a median of 7 years. Among the 96 patients, 69 (72%) were males and 27 (28%) were females. The diagnosis of ALL was based on the presence of 20% or more lymphoblasts in the bone marrow according to WHO criteria. Out of the 96 studied ALL patients, 46 (48%) patients were de novo ALL cases. Ninety-nine age- and sex-matched healthy volunteers who did not suffer from any illness were included in the study as a control group, they were 61 (61.6%) males and 38 (38.3%) females, their ages ranged from 1.5 to 12 years with a mean of 5.929 ± 2.84 and a median age of 5.5 years. Controls were selected from Kasr El Aini outpatient clinic, Cairo University Hospitals.
All cases were subjected to careful history-taking, comprehensive clinical examination, and full laboratory investigations including complete blood counts, peripheral blood and bone marrow smear examination, cyto-chemistry, immune-phenotyping, and molecular analysis for t(1;19), t(4;11), t(12;21), t(9;22) p190kD, and t(9;22) p210kD. The clinical and laboratory data of the ALL patients are shown in Table 1. Informed consent was obtained from all subjects’ legal guardians in accordance with the Declaration of Helsinki and the hospital-based ethics committee as well. The study protocol was approved by the Research Ethics Committee of the Clinical Pathology Department, Faculty of Medicine.
GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) genotyping
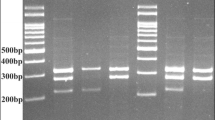
GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) single nucleotide polymorphisms (SNPs) were tested using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) assay. Two milliliters of whole blood samples were collected from each patient and control subject in sterile ethylene di-amine tetra acetic acid (EDTA) vacutainer tubes. Samples were stored at − 20 °C or used directly within 24 h for DNA extraction. Briefly, DNA extraction from peripheral blood samples was carried out using the Biospin Whole Blood Genomic DNA Extraction Kit (Cat# BSC06S1 Bioflux, China). DNA was stored at − 20 °C until use. PCR-RFLP was performed for GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) on Hybaid PCR express thermo-cycler (PCR system 9700) using the 2x Dream Taq Green PCR Master Mix (Cat# MB203-0100, GeneDireX, China). In brief, the 25-μL reaction mixture contains 12.5 μL 2X Dream Taq Green PCR Master Mix (2x), 5 μL DNA, 2 μL primer pair mix, and 5.5 μL sterile DW. The primers used for amplification were GSTO1*A140D (rs4925) Forward 5′-GAACTTGATGCACCCTTGGT-3′; Reverse 5′-TGATAGCTAGGAGAAATAATTAC-3′ and GSTO2*N142D (rs156697) Forward 5′-AGGCAGAACAGGAACTGGAA-3′; Reverse 5′-GAGGGACCCCTTTTTGTACC-3′. The PCR parameters were as follows: initial denaturation at 94 °C for 2 min, 35 cycles of PCR reaction at 95 °C for 30 s, 60 °C for 1 min and 72 °C for 2 min, and then 72 °C for 2 min. Identification of the alleles at each polymorphic site was performed by incubating the PCR products with a restriction enzyme [BstC8I (Cat# E305, SibEnzyme, RUSSIA) and BstMB I (Cat# E120, SibEnzyme, RUSSIA)] for GSTO1*A140D and GSTO2*N142D, respectively. The amplified products were incubated with the restriction enzyme at 37 °C for 16 h in a total reaction volume of 20 μL containing 15 μL of the amplified product and 5 μL of the diluted restriction enzyme. The digested fragments were separated by electrophoresis through a 2% agarose gel. These PCR products and digested fragments were separately visualized in 2% agarose gel with ethidium bromide staining under UV light. The digestion products for GSTO1*A140D (rs4925) revealed the presence of three different patterns: the C/C wild-type homozygote demonstrated by 2 fragments (186 bp and 68 bp), the A/A polymorphic homozygote presented by a single fragment (254 bp), and the C/A heterozygote exhibited 3 fragments (254 bp, 186 bp, and 68 bp) (Fig. 1). Similarly, the digestion products for GSTO2*N142D (rs156697) revealed the presence of three different patterns: the A/A wild-type homozygote demonstrated a single fragment (185 bp), the G/G homozygote presented 2 fragments (122 bp and 63 bp), and the A/G heterozygote exhibited 3 fragments (185 bp, 122 bp, and 63 bp) (Fig. 2).

Agarose gel electrophoresis of GSTO1*A140D (rs4925) SNP RPFP-PCR products. Lanes 1, 2, and 3: homozygous wild genotype C/C (186 bp, 68 bp). Lane 4: homozygous mutant genotype A/A (254 bp). Lanes 5 and 6: heterozygous mutant genotype C/A (254,186 bp, 68 bp)

Agarose gel electrophoresis of GSTO2*N142D (rs156697) SNP RPFP-PCR products. Lanes 1 and 2: heterozygous mutant genotype G/A (185 bp, 122 bp, 63 bp). Lane 3: homozygous wild genotype A/A (185 bp). Lanes 4, 5, and 6: homozygous mutant genotype G/G (122 bp, 63 bp)
Statistical analysis
Data were analyzed using SPSS with statistical package version 15. Numerical data were expressed as the mean and standard deviation (SD) or median and range as appropriate while qualitative data were expressed as frequency and percentage. For the quantitative data, Student’s t test was used to compare the studied groups while the categorical variables were compared using chi-square (χ2) test. Odds ratio (OR) with 95% confidence intervals (CI) was calculated for the studied genes between cases and controls as well as between every 2 groups. A p value < 0.05 was considered as statistically significant.
Results
The characteristics of ALL patients
Our study was performed on 96 ALL patients and 99 age- and sex-matched control subjects. The baseline characteristics of ALL patients are shown in Table 1. The clinical characteristics of the patients were obtained by patient clinical examination and reviewing the medical records. The patients' ages ranged from 1.5 and 14 years with a mean of 6.95 ± 3.01 years. Among the 96 patients, 69 (72%) were males and 27 (28%) were females. In this study, 32 (33.3%) patients had pallor, 15 (15.6%) patients suffered from dyspnea, 49 (51%) patients were feverish, 1 (1.04%) patient had bleeding manifestations, 4 (4.1%) patients had bony pains, 3 (3.1%) patients had lymphadenopathy, and 1 (1.04%) patient had CNS infiltration at presentation. At initial diagnosis, the patients presented with high initial white blood cell count (WBCs) ranging from 2 to 437 × 109/L with a median of 47.9 × 109/L. Blast cell count ranged from 22 to 98% with a median of 60%. Hemoglobin level ranged from 3.3 to 11.8 g/dL with a median of 7.9 g/dL and platelet count ranged from 3 to 118 × 109/L with a median of 39.5 × 109/L. The most commonly encountered FAB subgroup was L2 in 53 (55.2%) patients followed by L1 in 43 (44.8%) patients but none of the patients was classified as FAB L3. 61 (63.5%) patients were classified as B-ALL where 41 (42.7%) patients as pre-B and 20 (20.8%) patients as common B-ALL while 35 (36.4%) patients as T-ALL where 7 (7.3%) patients as early T-ALL, 25 (26%) patients as intermediate T-ALL and 3 (3.1%) patients as late T-ALL. t(1;19) was found in 3 (3.1%) patients while all cases were negative for t(4;11), t(12;21), t(9;22) p190kD, and t(9;22) p210kD. Out of the studied 96 ALL patients, 46 (48%) patients were de novo cases. The clinical and hematological data of the de novo ALL cases are demonstrated in Table 2.
GSTO1*A140D (rs4925) genotyping
No statistically significant different GSTO1*A140D (rs4925) genotype distribution was observed among ALL cases and controls [CC (44.8% vs. 56.6%), CA (46.9% vs. 40.4%), and AA (8.3% vs. 3.0%), respectively] (0.106). GSTO1*A140D variant genotypes' (CA + AA) frequency did not differ significantly between ALL cases and controls (55.8% vs. 43.4%) [OR (95%CI): 1.643 (0.932–2898)] (0.085). The minor allele frequency (MAF) of GSTO1*A140D-A was comparable between the 2 groups (0.31 vs. 0.23) [OR (95%CI): 1.563 (0.997–2.488)] (0.051) (Table 3).
Comparison of de novo patient group to controls revealed a statistically significant different GSTO1*A140D (rs4925) genotype distribution [CC (37% vs. 56.6%), CA (47.8% vs. 40.4%), and AA (15.2% vs. 3.0%), respectively] (0.008). GSTO1*A140D variant genotypes' (CA+AA) frequency was significantly higher in the de novo group compared to controls (63% vs. 43.4%) [OR (95%CI): 2.222 (1.038–4.558)] (0.028), and the MAF of GSTO1*A140D-A was significantly higher in de novo cases compared to controls (0.39 vs. 0.23) [OR (95%CI): 2.124 (1.24–3.620)] (0.005) (Table 3).
GSTO2*N142D (rs156697) genotyping
A statistically significant different GSTO2*N142D (rs156697) genotype distribution was observed among ALL patients and controls [AA (26% vs. 36.3%), AG (62.5% vs. 61.6%), and GG (11.4% vs. 2.0%), respectively] (0.017). GSTO2*N142D variant genotypes' (AG + GG) frequency did not differ significantly between the 2 groups (73.9% vs. 63.6%) [OR (95%CI): 1.600 (0.866–2955)] (0.132). The MAF of GSTO2*N142D-G was significantly higher in ALL patients than in controls (0.42 vs. 0.32) [OR (95%CI): 1.521 (1.006–2.298)] (0.046) (Table 3).
A statistically significant difference of GSTO2*N142D (rs156697) genotype distribution was observed among de novo ALL patients and controls [AA (37% vs. 36.3%), AG (45.7% vs. 61.6%), and GG (17.3% vs. 2.0%), respectively] (0.002). GSTO2*N142D variant genotypes' (AG + GG) frequency was comparable between the de novo group and controls (63% vs. 63.6%) [OR (95%CI): 0.975 (0.472–2.013)] (0.945), and the MAF of GSTO2*N142D-G was comparable between the 2 groups (0.40 vs. 0.32) [OR (95%CI): 1.37 (0.825–2.296)] (0.220) (Table 3).
The impact of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms on ALL prognosis
ALL patients were categorized into high-risk and standard-risk groups based on the age, WBCs, and immune-phenotypic analysis. The high-risk group is more than 10 years old or had WBCs > 50 × 109/L or T cell immune-phenotype while the standard-risk group is less than 10 years old or had WBCs < 50 × 109/L and has no T cell immune-phenotype. ALL patients with high-risk factors were studied for the frequencies of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms. The present study revealed that GSTO1*A140D and GSTO2*N142D variant genotypes in the high-risk ALL group had a higher frequency than the corresponding wild genotypes but the difference did not reach a statistical significance (Table 4).
Moreover, the combined GSTO1*A140D and GSTO2*N142D polymorphisms in the high risk ALL group showed a higher frequency than single polymorphisms [GSTO1*A140D or GSTO2*N142D] and wild genotypes but the difference did not reach a statistical significance. Thus, we did not find any association of GSTO1*A140D and/or GSTO2*N142D polymorphisms with the poor prognostic factors (Table 4).
Discussion
Acute lymphoblastic leukemia (ALL) is the most common type of childhood cancer [1]. Genetic susceptibility has been studied as a risk factor since environmental exposure to xenobiotics induces the accumulation of somatic mutations [4]. Drug detoxification enzymes are divided into phase I and phase II enzymes [3]. Glutathione S-transferases (GST) are categorized as phase II drug detoxification enzymes that facilitate the conjugation of glutathione (GSH) using several endogenous and exogenous substances. They are indispensable for cellular protection against oxidative stresses [28]. GST gene polymorphisms cause alterations in the enzyme activity that may modify the individual susceptibility to cancer [25].
The present case-control study aimed to investigate the potential association of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms with the susceptibility to childhood ALL and their impact on ALL prognosis in the Egyptian population. Genotyping was performed by PCR amplification of the target gene followed by the RFLP technique. The study included 96 pediatric ALL patients and 99 age- and sex-matched control subjects.
The ALL population was a selection of all patients attending the pediatric hematology clinic, New Children Hospital, Cairo University (NCHCU) during the period from April 2015 to October 2017. Selected patients were Egyptian ALL patients of the pediatric age group. The patients’ ages ranged from 1.5 and 14 years with a mean of 6.95 ± 3.01 years and a median of 7 years. Among the 96 patients, 69 (72%) were males and 27 (28%) were females with a male to female ratio = 2.5:1. The diagnosis of ALL was based on WHO criteria. Sixty-one (63.5%) patients were classified as B-ALL while 35 (36.4%) patients as T-ALL. t(1;19) was found in 3 (3.1%) patients while all cases were negative for t(4;11), t(12;21), t(9;22) p190kD, and t(9;22) p210kD. The number of patients with the most common ALL translocations is very low. That might be attributed to the low sensitivity of the used technique. Out of the studied 96 ALL patients, 46 (48%) patients were de novo ALL cases.
In the present study, GSTO1*A140D (rs4925) genotyping revealed a statistically non-significant distribution among ALL patients and controls [CC (44.8% vs. 56.6%), CA (46.9% vs. 40.4%), and AA (8.3% vs. 3.0%), respectively] (0.106). GSTO1*A140D variant genotypes' (CA + AA) frequency did not differ significantly between ALL cases and controls (55.8% vs. 43.4%) [OR (95%CI): 1.643 (0.932–2898)] (0.085). However, comparing the de novo ALL cases to controls revealed a statistically significant different GSTO1*A140D genotype distribution [CC (37% vs. 56.6%), CA (47.8% vs. 40.4%), and AA: (15.2% vs. 3.0%), respectively] (0.008). GSTO1*A140D variant genotypes' (CA + AA) frequency was significantly higher in the de novo group than in controls (63% vs. 43.4%) [OR (95%CI): 2.222 (1.038–4.558)] (0.028).
The MAF of GSTO1*A140D-A was higher in ALL patients than in controls; however, the difference did not reach a statistically significant level (0.31 vs. 0.23) [OR (95%CI): 1.563 (0.997–2.488)] (0.051). However, in the de novo group, it was significantly higher than controls (0.39 vs. 0.23) [OR (95%CI): 2.124 (1.24–3.620)] (0.005) (Table 3).
GSTO2*N142D (rs156697) genotyping showed a statistically significant different distribution between ALL patients and controls [AA (26% vs. 36.3%), AG (62.5% vs. 61.6%), and GG: (11.4% vs. 2.0%), respectively] (0.017) and between de novo ALL patient group and controls [AA (37% vs. 36.3%), AG (45.7% vs. 61.6%), and GG: (17.3% vs. 2.0%), respectively] (0.002). The MAF of GSTO2*N142D-G was significantly higher in ALL patients than in controls (0.42 vs. 0.32) [OR (95%CI): 1.521 (1.006–2.298)] (0.046) (Table 3).
Based on these findings, we concluded that GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms are associated with an increased risk of de novo childhood ALL in the Egyptian population. Our results are in accordance with those of Pongstaporn et al. [29], who investigated the association of GSTO polymorphisms with the susceptibility to ALL in the pediatric Thai population. They reported that GSTO1*A140D (rs4925) was associated with the susceptibility to ALL [OR (95%CI): 2.24 (1.16–4.35); 0.009] while GSTO2*N142D (rs156697) was associated with the high-risk group of ALL patients [OR (95%CI): 5.52 (1.72–17.71); 0.004], thus, suggesting that GSTO polymorphisms may play a role in the occurrence of ALL.
However, our results were discordant with the findings of Rezazadeh et al. [30] who studied the association of GSTO polymorphisms with the susceptibility to pre-B-ALL in the pediatric Iranian population. They found that GSTO genotype distribution was similar among pre-B-ALL patients and controls. The difference between their results and ours may be attributed to the ethnic variation of the GSTO polymorphisms as variant genotypes are variably expressed among different ethnic populations as shown in previous studies [25,26,27].
The MAF of GSTO1*A140D-A in the Europeans (Italian and Spaniard) and European ancestors (Australian) ranged from 0.302 to 0.399 while in Asians (Chinese, Japanese and Mongolian), it ranged from 0.108 to 0.165. In the Africans (Bantu and Ovambo), it was much lower than those of the Europeans and Asians (ranging from 0.040 to 0.081). In contrast, the Turkish population followed a pattern similar to that of Africans. On the other hand, the MAF of GSTO2*N142D-G in Asians (ranging from 0.173 to 0.285) was lower than that of the Europeans (ranging from 0.310 to 0.341). In the Turkish population, it was similar to that of the Asians while in the Africans, it was higher than that of Asians, ranging from 0.583 to 0.855 [15, 26, 27, 31]. In our Egyptian control group, the MAF of GSTO1*A140D-A was 0.23 while that of GSTO2*N142D-G was 0.32. The latter is similar to that of the Europeans. In conclusion, for GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697), the MAFs were different among different ethnic populations suggesting that these polymorphisms could be useful genetic markers but the association of cancer with a particular mutation in one population might be of limited value as a cancer biomarker into another population. Further detailed studies on different ethnic populations are needed to clarify the geographical distribution of these genetic markers and the possible association between GSTO polymorphism, ethnicity, and the occurrence of ALL. In addition, studying the physiological role of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) will lead to a greater insight into the significance of GSTO polymorphism.
ALL patients were categorized into high-risk and standard-risk groups based on the age, WBCs, and immune-phenotypic analysis. The high-risk group is more than 10 years old or had WBCs > 50 × 109/L or T cell immune-phenotype while the standard-risk group is less than 10 years old or had WBCs < 50 × 109/L or had no T cell immune-phenotype [32]. The present study revealed that the frequency of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) variant genotypes in the high-risk ALL group was higher than the corresponding wild genotypes; moreover, the frequency of the combined GSTO1*A140D and GSTO2*N142D polymorphisms in the high-risk ALL group was higher than single polymorphisms [GSTO1*A140D or GSTO2*N142D] and wild genotypes, but these differences did not reach a statistical significance (Table 4). However, Pongstaporn et al. [29] found that GSTO2*N142D (rs4925) polymorphism was associated with the high-risk group of childhood ALL (0.004), indicating that GSTO2*N142D is associated with the susceptibility to high-risk ALL and can be used as an indicator of the disease severity.
Conclusion
In conclusion, we identified the frequencies of GSTO1*A140D (rs4925) and GSTO2*N142D (rs156697) polymorphisms in the pediatric Egyptian ALL patients. In order to explore the geographic distribution of these markers, the MAF obtained was compared with different ethnic populations as reported in the literature. We also demonstrated a statistically significant difference of GSTO1*A140D and GSTO2*N142D genotype distribution between de novo ALL cases and controls. Compared to the control group, the MAF of GSTO1*A140D-A was overexpressed in de novo ALL cases and that of GSTO2*N142D-G was significantly higher in ALL patients. We did not test more than reported in the manuscript with a p value of 0.05. There was no need to correct the interpretation of p values for the statistical analysis done. These findings suggest that the studied polymorphisms could be genetic risk factors of de novo childhood ALL in the Egyptian population. The present study also showed that the high-risk ALL group had a higher frequency of GSTO1*A140D and GSTO2*N142D variant genotypes compared to the corresponding wild genotypes and a higher frequency of the combined GSTO1*A140D and GSTO2*N142D polymorphisms compared to single polymorphisms (GSTO1*A140D or GSTO2*N142D) and wild genotypes; however, these differences did not reach a statistical significance.
Recommendations
To the best of our knowledge, this is the first study to examine the association of GSTO1*A140D (rs4925) and GSTO2*A142D (rs156697) polymorphisms with the susceptibility to childhood ALL in the Egyptian population. These results must be verified by further studies that enroll a larger population. Moreover, the follow-up of the patients is highly recommended to verify the prognostic implications of the studied polymorphisms. In the present study, since the samples were obtained from de novo ALL cases at the time of diagnosis, contamination by tumor cells cannot be ruled out. The presence of deletion or amplifications in the tumor genome might impact the genotyping and bias the results. So, more studies are recommended on Egyptian pediatric ALL cases when reaching remission to reassess the difference in the studied genotype frequencies in the studied pediatric ALL population.
Availability of data and materials
Not applicable.
Abbreviations
- ALL:
-
Acute lymphoblastic leukemia
- CYP:
-
Cytochrome P450 isoenzymes
- GSH:
-
Glutathione
- GSTA:
-
Glutathione S-methyl transferase α
- GSTK:
-
Glutathione S-methyl transferase κ
- GSTM:
-
Glutathione S-methyl transferase µ
- GSTO:
-
Glutathione S-methyl transferase omega class
- GSTO1:
-
Glutathione S-methyl transferase omega class 1
- GSTO2:
-
Glutathione S-methyl transferase omega class 2
- GSTP:
-
Glutathione S-methyl transferase π
- GSTs:
-
Glutathione S-methyl transferase enzymes
- GSTS:
-
Glutathione S-methyl transferase ς
- GSTT:
-
Glutathione S-methyl transferase θ
- GSTZ:
-
Glutathione S-methyl transferase ζ
- IL-1b:
-
Interleukin-1b
- MAF:
-
Minor allele frequency
- NAT:
-
N-acetyl transferases
- PCR-RFLP:
-
Polymerase chain reaction – restriction fragment length polymorphism
- SNP:
-
Single nucleotide polymorphism
- WBCs:
-
White blood cell count
References
Abdelmabood S, Fouda AE, Boujettif F, Mansour A (2020) Treatment outcomes of children with acute lymphoblastic leukemia in a middle-income developing country: high mortalities, early relapses, and poor survival. Pediatr (Rio J) 96:108–116
Perera FP (1996) Molecular epidemiology: insights into cancer susceptibility, risk assessment, and prevention. J Natl Cancer Inst 88:496–509
Vineis P, Malats N, Lang M, d'Errico A, Caporaso N, Cuzick J (1999) Metabolic polymorphisms and susceptibility to cancer. IARC press, Lyon
Brisson GD, Alves LR, Pombo-de-Oliveira MS (2015) Genetic susceptibility in childhood acute leukemia: a systematic review. Ecancermedicalscience 9:539
He HR, Zhang XX, Sun JY, Hu SS, Ma Y, Dong YL et al (2014) Glutathione S-transferase gene polymorphisms and susceptibility to chronic myeloid leukemia. Tumor Biol 35(6):6119–6125
Song K, Yi J, Shen X, Cai Y (2012) Genetic polymorphisms of glutathione S-transferase genes GSTM1, GSTT1 and risk of hepatocellular carcinoma. PLoS One 7(11):e48924
Wu B, Dong D (2012) Human cytosolic glutathione transferases: structure, function, and drug discovery. Trends Pharmacol Sci 33(12):656–668
Autrup H (2004) Gene-environment interaction in environment carcinogens. Cancer Environ Dis 20:71–101
Traverso N, Ricciarelli R, Nitti M, Marengo B, Furfaro AL, Pronzato MA et al (2013) Role of glutathione in cancer progression and chemoresistance. Oxid Med Cellul Longev 2013:972913 10 pages
Gate L, Tew KD (2001) Glutathione S-transferases as emerging therapeutic targets. Expert Opin Ther Targets 5:477–489
Safarinejad MR, Safarinejad S, Shafiei N, Safarinejad S (2013) Association of genetic polymorphism of glutathione S-transferase (GSTM1, GSTT1, GSTP1) with bladder cancer susceptibility. Urol Oncol 31(7):1193–1203
Yang H, Shen X, Li B, Ma R (2014) Association between glutathione S-transferase T1 null genotype and risk of lung cancer: a meta-analysis of 55 studies. Tumor Biol 35(3):2359–2366
Frova C (2006) Glutathione transferases in the genomic era: new insights and perspectives. Biomol Eng 23:149–169
Girardini J, Amirante A, Zemzoumi K, Serra E (2002) Characterization of an omega-class glutathione S-transferase from Schistosoma mansoni with glutaredoxin-like dehydroascorbate reductase and thiol transferase activities. Eur J Biochem 269:5512–5521
Whitbread AK, Tetlow N, Eyre HJ, Sutherland GR, Board PG (2003) Characterization of the human omega class glutathione transferase genes and associated polymorphisms. Pharmacogenetics 13:131–144
Wang L, Xu J, Ji C, Gu S, Lv Y, Li S et al (2005) Cloning, expression and characterization of human glutathione S-transferase omega 2. Int J Mol Med 16:19–27
Laliberte RE, Perregaux DG, Hoth LR, Rosner PJ, Jordan CK, Peese KM et al (2003) Glutathione s-transferase omega 1-1 is a target of cytokine release inhibitory drugs and may be responsible for their effect on interleukin-1beta posttranslational processing. J Biol Chem 278(19):16567–16578
Board PG (2011) The omega-class glutathione transferases: structure, function, and genetics. Drug Metab Rev 43(2):226–235
Li YJ, Oliveira SA, Xu P, Martin ER, Stenger JE, Scherzer C et al (2003) Glutathione S-transferase omega-1 modifies the age of onset of Alzheimer disease and Parkinson disease. Hum Mol Genet 12:3259–3267
Tanaka-Kagawa T, Jinno H, Hasegawa T, Makino Y, Seko Y, Hanioka N et al (2003) Functional characterization of two variant human GSTO1K1s (Ala140Asp and Thr217Asn). Biochem Biophys Res Commun 301:516–520
Ada TG, Ada AO, Kunak SC, Alpar S, Gulhan M, Iscan M (2013) Association between glutathione S-transferase omega 1 A140D polymorphism in the Turkish population and susceptibility to non-small cell lung cancer. Arh Hig Rada Toksikol 64:61–67
Pongstaporn W, Rochanawutanon M, Wilailak S, Linasamita V, Weerakiat S, Petmitr S (2006) Genetic alterations in chromosome 10q24.3 and glutathione S-transferase omega 2 gene polymorphism in ovarian cancer. J Exp Clin Cancer Res 25:107–114
Morari EC, Lima ABC, Bufalo NE, Leite JL, Granja F, Ward LS (2006) Role of glutathione-S-transferase and codon 72 of p53 genotypes in epithelial ovarian cancer patients. J Clin Res Clin Oncol 132:521–528
Andonova IE, Justenhoven C, Winter S, Hamann U, Baisch C, Rabstein S et al (2010) No evidence for glutathione S-transferases GSTA2, GSTM2, GSTO1, GSTO2, and GSTZ1 in breast cancer risk. Breast Cancer Res Treat 121(2):497–502
Marahatta SB, Punyarit P, Bhudisawasdi V, Paupairoj A, Wongkham S, Petmitr S (2006) Polymorphism of glutathione S-transferase omega gene and risk of cancer. Cancer Lett 236:276–281
Fujihara J, Kunito T, Takeshita H (2007) Frequency of two human glutathione-S-transferase omega-1 polymorphisms (E155 deletion and E208K) in Ovambo and Japanese populations using the PCR-based genotyping method. Clin Chem Lab Med 45:621–624
Takeshita H, Fujihara J, Takastuka H, Agusa T, Yasuda T, Kunito T (2009) Diversity of glutathione S-transferase OMEGA 1 (A140D) and 2 (N142D) gene polymorphisms in worldwide populations. Clin Exp Pharmacol Physiol 36:283–286
Townsend DM, Tew KD (2003) The role of glutathione-S transferase in anticancer drug resistance. Oncogene 22:7369–7375
Pongstaporn W, Pakakasama S, Sanguansin S, Hongeng S, Petmitr S (2009) Polymorphism of glutathione S-transferase omega gene: association with risk of childhood acute lymphoblastic leukemia. J Cancer Res Clin Oncol 135(5):673–678
Rezazadeh D, Moradi MT, Kazemi A, Mansouri K (2015) Childhood Pre-B acute lymphoblastic leukemia and glutathione S-transferase omega 1 and 2 polymorphisms. Int J Lab Hematol 37:530–535
Paiva L, Marcos R, Creus A, Coggan M, Oakley AJ, Board PG (2008) Polymorphism of glutathione transferase omega 1 in a population exposed to a high environmental arsenic burden. Pharmacogenet Genomics 18:1–10
Smith M, Arthur D, Camitta B, Carroll AJ, Crist W, Gaynon P et al (1996) Uniform approach to risk classification and treatment assignment for children with acute lymphoblastic leukemia. J Clin Oncol 14:18–24
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Dr. “EO”: conceptualization and methodology; Dr. “WM”: data curation and formal analysis; Dr. “SB”: visualization and investigation; Dr. “DM”: writing - original draft preparation; Dr. “MY”: writing, reviewing, and editing; and Dr. “HA”: supervision, validation, and final approval of the version to be submitted for publication. We affirm that the submission represents original work that has not been published previously and is not currently being considered or submitted to another journal until a decision has been made. Also, we confirm that the manuscript has been read and approved by all the authors and each author believes that the manuscript represents honest work and be able to verify the validity of the reported results. There is no published work that concerns the same patients included in the current study. This work received no specific grant from any funding source. The authors have no conflict of interest to declare.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study protocol was approved by the Research Ethics Committee of the Clinical Pathology Department, Faculty of Medicine, Cairo University.
An informed written consent was obtained from all subjects’ legal guardians in accordance with the Declaration of Helsinki and the hospital-based ethics committee as well.
Consent for publication
Not applicable.
Competing interests
The authors have no conflicts of interest to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yassa, M.E., Arnaout, H.H., Botros, S.K. et al. The role of glutathione S-transferase omega gene polymorphisms in childhood acute lymphoblastic leukemia: a case-control study. Egypt J Med Hum Genet 22, 17 (2021). https://doi.org/10.1186/s43042-020-00128-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43042-020-00128-7