
Abstract
Tapuy is an indigenous wine produced in the Philippines. Rice wine fermentation produces a by-product of rice leftover and microbial biomass, collectively called lees, which usually is discarded as a waste product. However, studies have shown the potential value of lees as a healthy food source. We wanted to determine the nutritional profile of tapuy wine and its lees and their abilities to extend lifespan in Caenorhabditis elegans. Tapuy lees (7.65 g/100 g dry weight) was 18.5-fold greater in protein content compared to tapuy wine. Its sugar content (27.66 g/100 g dry weight) is similar to the tapuy wine’s sugar content (23.465 g/100 g dry weight). Through LC–MS/MS, 18.75% and 12.39% of spectral peaks in tapuy wine and lees were matched, respectively, to specific compounds, and several of them are associated with beneficial health effects. Furthermore, the phenolic content in tapuy lees (19,475.526 Gallic Acid Equivalent) is 6.5-fold greater compared to tapuy wine. Surprisingly, the DPPH and FRAP antioxidant assays show that tapuy lees had a lesser antioxidant ability compared to tapuy wine. However, C. elegans fed with supplementation of tapuy lees extract showed a 16.24% increase in mean lifespan, which is higher compared to the 6.10% increase in mean lifespan when supplemented with tapuy wine. Taking all these findings together, our study revealed that tapuy lees has a greater nutritional value than tapuy wine, and this underutilized and wastefully discarded product can be repurposed as a potential functional food.
Graphical Abstract

Similar content being viewed by others

Introduction
Fermentation is a process in which complex macromolecules are converted into simpler molecules (Sharma et al. 2020). It has long been used in human civilizations to change food characteristics such as flavor and aroma, preserve food, and remove unwanted compounds (Allwood et al. 2021; Şanlier et al. 2017; Sharma et al. 2020). A surge in interest in fermented food sources has increased in recent years due to numerous studies pointing to their benefits as functional food or food that have biofunctional properties leading to the improved health of the consumer (Dimidi et al. 2019). Bioactive molecules produced as a by-product of fermentation have been shown to display a wide range of health benefits such as being antihypertensive, prebiotic, antimicrobial, anticarcinogenic, antioxidant, antiallergenic, and an opioid antagonist (Şanlier et al. 2017; Sharma et al. 2020). One such fermentation by-product is whey from the cheese-making process, which has many nutritional benefits such as a source of protein and antioxidants, among others (Mazorra-Manzano et al. 2020). Another fermentation by-product often used is kimchi juice, the liquid by-product of kimchi fermentation. This juice is frequently used in kitchens.
Rice wines such as sake (Japan), cheongju (Korea), and shaoxing (China) are a group of fermented alcoholic beverages that are usually made from a mixture of different cereals, mainly rice, and are grown in different countries in Asia (Kwon et al. 2014). Rice wines have been discovered to have properties that lead to better health. Chinese rice wine has been demonstrated to have antifatigue and antiaging effects (Zhao et al. 2018). Korean makgeolli has also been suggested to be a source of antioxidants, dietary fibers, vitamins, bioactive compounds, and probiotics (Shimoga & Kim 2021). A common by-product of rice wine fermentation is wine lees, the rice leftover after wine fermentation. The wine lees has been proposed to have potential health benefits. Sake lees are frequently used in Japanese cooking after the rice wine fermentation process. Sake lees are a potent source of proteins, fiber, vitamins, and have been shown to decrease serum triacylglycerol concentrations (Tsutsui et al. 1998). It has also been shown to decrease lipid accumulation in adipocytes (Motono et al. 2021).
Tapuy is an indigenous rice wine produced in the northern provinces of the Philippines, particularly in the Cordillera regions. The native tribes produce this alcoholic beverage during special occasions such as weddings, fiestas, harvesting ceremonies, baptism, and as an offering to their deities or anitos. However, appreciation for tapuy and its production has been decreasing due to intense competition from commercially produced rice wines such as sake and soju. Tapuy wine is produced mainly from white and/or black glutinous rice mixed with a starter culture called bubod, a disc made of hardened starch powder packed with various fermenting microorganisms.
Tapuy lees is a product of the tapuy wine fermentation process, and it may contain the same health benefits as the rice wine itself. About 27% of the rice weight used in tapuy fermentation forms wine lees and it is typically discarded (Manaois & Morales 2014; Morales & Manaois 2011). Lees have been shown to have health benefits, as seen in the use of sake lees in Japanese cuisine (Motono et al. 2021; Tsutsui et al. 1998). This study revealed the potential of tapuy lees as a healthy food source by determining its nutritional profile and its effect on lifespan in Caenorhabditis elegans.
Methodology
Nutritional profile analysis of tapuy wine and lees
Mean total sugar content was determined via the Munson-Walker Method at the Intertek Testing and Service Center, Makati, Metro Manila. Mean total fat content was determined via Acid Hydrolysis at the Intertek Testing and Service Center, Makati, Metro Manila. Mean total protein content was determined via the Kjeldahl Method at the Intertek Testing and Service Center, Makati, Metro Manila.
Preparation of tapuy wine and lees extracts
Tapuy wine extracts were prepared by dehydrating the samples at 35 °C. The dried extract was then placed in a screw-cap glass vial. Tapuy lees extract was prepared by first dried and then powdered. The powdered lees were then soaked in 95% ethanol for 96 h in a 1:2 ratio (v/v) and separated from the ethanol via filtration and the filtrate was concentrated using a vacuum concentrator (SpeedVac) for 24 h. The dried lees extract were then resuspended in 0.1% DMSO. High concentrations of tapuy wine or lees were prepared as 100,000 μg/mL. Low concentrations of tapuy wine or lees were prepared as 10,000 μg/mL.
Total phenolic content and antioxidant activity of tapuy wine and lees
Mean total phenolic content was determined via the Folin-Ciocalteu Assay at the Natural Products Laboratory, Department of Molecular Biology and Biochemistry. Total phenolic content was determined using a modified Folin-Ciocalteu Assay from Benabdellah et al. (2016). In brief, 15.4μL of sample and gallic acid standard (at different concentration) were mixed with 61.54μL of Folin-Ciocalteu reagent (in a 1:10 dilution with deionized water). The mixture was neutralized with 1234μL of 7.5% sodium carbonate. The mixtures were left to stand at room temperature for 30 min. Absorbance was then read at 765 nm.
DPPH and FRAP antioxidant assays were performed at the Natural Products Laboratory, Department of Molecular Biology and Biochemistry. Antioxidant potentials were determined using the ferric reducing antioxidant power (FRAP) and 2, 2-diphenyl-1-picrylhydrazyl (DPPH) assays. For the DPPH assay, 10μL of ascorbic acid was used as a standard. 10 μL of tapuy samples were loaded into a 96-well microplate. 140μL of 6.85 × 10−5 M DPPH was added into each well and left to incubate at room temperature and in the dark for 30 min. The solvents used were methanol for DPPH and 0.1% DMSO for the tapuy samples. Absorbance was then read at 517 nm. The absorbance values were plotted against the concentration of the sample. The equation of the line was then used to obtain the actual IC50 value. For the FRAP assay, 70μL of butylated hydroxytoluene (BHT) was used as a standard. Samples were mixed with 176.5μL of 0.2 M, pH 7.4 sodium phosphate buffer and 176.5μL of 1% (K3Fe(CN)6). The resulting mixture was incubated at 50 °C for 20 min. Following this, the mixtures were acidified with 176.5μL of 10% trichloroacetic acid and centrifuged at 650xg for 10 min. An aliquot of 273μL of the supernatant was added to 273μL of deionized water. 55μL of 0.1% FeCl3 was added to this solution. The solvent used was distilled water. Absorbance was read at 700 nm (Bueno et al. 2013). The absorbance values were plotted against the concentration of the sample. The equation of the line was then used to obtain the actual EC50 value.
Liquid chromatography tandem mass spectrometry (LC–MS/MS) was performed at the Molecular Diagnostics and Multi-omics Laboratory, Department of Molecular Biology and Biochemistry, UP Manila.
Lifespan assay
Three plates of thirty age-synchronized (via bleaching) wild-type Caenorhabditis elegans at the L4-stage each were worm-picked and placed in a fresh nematode growth media (NGM) petri dish. All treatment plates had an equal volume of heat-killed Escherichia coli (OP50) solution and either tapuy wine or lees extract solution. The final concentration of the extracts after mixture with OP50 were 50,000 µg/mL for the high concentration and 5,000 µg/mL for the low concentration. The worms were transferred to freshly prepared NGM petri dishes every other day. The proportion of dead, surviving, and missing worms were counted every 12 h until no surviving worm was present on the petri dish. Worms that provided a response to a gentle touch with a platinum wire pick were categorized as alive. Worms that provided no response to a gentle touch with a platinum wire pick were categorized as dead. A control group was set up and fed with E. coli (OP50 strain). Lifespan assays were carried out in three trials and pooled prior to statistical analyses (n = 90). Worms that went missing during the lifespan assay were subtracted from the original sample size.
Statistical analyses
The means and standard deviations were calculated for the total protein, sugar, fat, and phenolic contents. Means and standard deviations were also calculated for the IC50 and EC50 for the antioxidant activities and for the lifespan of C. elegans. A one-way analysis of variance (ANOVA) and Tukey’s honest significance test were conducted using R 4.2.2 and RStudio 2022.02.0 to determine significant differences between the means. A Kaplan–Meier survival plot was generated using the ggplot2 package on R. Finally, all bar graphs were generated using the ggplot2 package on R.
Results and discussion
Tapuy lees has a significantly higher mean total protein content compared to tapuy wine
The difference between the mean protein content of tapuy wines and lees is significant (p < 0.05) as shown in Table 1. There is an 18.5-fold greater protein content in tapuy lees (7.65 g/100 g dry weight) than of tapuy wine (0.41 g/100 g dry weight). This is similar with the nutritional analysis observed in sake lees, which also has a high protein content (Ito et al. 2022; Tsutsui et al. 1998). Black rice, one of the main ingredients used in the production of tapuy wine, has been shown to have increased levels of protein compared to other rice varieties (Peng 2021; Rathna Priya et al. 2019). The mean protein content of tapuy lees is lower than the average protein content of black rice (11.5 g/100 g dry weight) reported by Peng et al. (2021). This may be due to protein breakdown during fermentation, which has been linked to an increased metabolite pool and more available amino acids (Diether & Willing 2019). Furthermore, the fermentation process can increase the digestibility of proteins (Alrosan et al. 2021; Çabuk et al. 2018). The lower mean protein content of tapuy lees compared to plain black rice may be due to some of the rice proteins dissolving into the wine.
The difference between the mean sugar content of tapuy wines and lees is not significant (p > 0.05) as shown in Table 1. However, the tapuy lees (27.66 g/100 g dry weight) has a slightly higher sugar content compared to the tapuy wine (23.47 g/100 g dry weight), which may be due to the higher content of the mono- and disaccharides after starch breakdown during fermentation. The metabolism of starch by amylolytic organisms involves two steps, the saccharification step in which starch is broken down into constituent mono- and disaccharides, and the fermentation step, in which the sugars are converted into ethanol (Toksoy Oner et al. 2005). We postulate that increasing the time of fermentation would lead to lower sugar content and higher ethanol content. Increasing the time of fermentation would give the microorganisms more time to convert the sugars within the substrate into ethanol. The similar sugar content would lead to a very similar level of sweetness between the tapuy wine and the lees.
Although the difference between the mean sugar content of tapuy wine and lees is not significant, the assay used to determine the sugar content only detects reducing sugars (Jackson & Mcdonald 1941). It may be possible that a large concentration of starch will remain in the tapuy lees and will be broken down inside the body upon consumption. This would effectively increase the total sugars consumed when eating tapuy lees and one should exercise caution in concluding that the total sugar content of the tapuy wine and lees is comparable in this sense. Alternatively, a large percent of the undetected starch is undigestible and may serve a prebiotic function, thus, benefitting the gut microbiota. The small disparity between the mean sugar content between the tapuy wine and lees may be due to the sugars being more diluted in the supernate (liquid wine) than in the precipitate (solid lees). Each grain of rice will have a smaller volume than the entire vat of liquid and thus, would have a more concentrated sugar content in spite of an overall lower amount of sugars inside the grain.
The traditional method of tapuy fermentation is done over a period of one month, which leads to the production of a relatively sweet wine. The soluble mono- and disaccharides from starch breakdown during fermentation give the wine its characteristic sweet taste. The tapuy lees, having both a higher mean sugar content and an overall higher concentration of sugars (by virtue of its smaller and denser size compared to the liquid wine), would be more palatable and appealing to people. The palatability of tapuy lees would make it easier to repurpose it as a functional food. However, tapuy can also be fermented for up to two months, which is enjoyed by the few who prefer a dryer taste with a higher alcohol content.
The difference between the mean fat content of tapuy wines and lees is significant (p < 0.05) as shown in Table 1. However, despite this significant difference, the overall fat content of both tapuy wine and lees is small (0.14 g/100 g dry weight and 1.27 g/100 g dry weight, respectively). Thus, the low-fat content of the tapuy wine and lees contributes minimally to their nutritional value.
Tapuy lees composition is more complex and diverse than tapuy wine
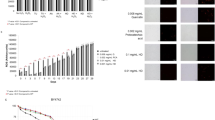
As shown in Figs. 1 and 2, the tapuy lees has 113 spectral peaks while tapuy wine has 43 spectral peaks. The more varied compound composition in the tapuy lees increases the likelihood of it containing compounds that may confer health benefits. Table 2 shows the summarized phenolic compounds identified using LC–MS/MS and screened using the Waters Traditional Chinese Medicine Library. Tapuy wine has 9 compounds identified from 43 spectral peaks (21%) while tapuy lees has 14 compounds identified from 113 spectral peaks (12.4%). There is only one common compound between the two samples (kalmanol), which suggests a clear difference in the phenolic compositions of the tapuy wine and lees. The more varied phenolic compounds in the lees suggests a wider spectrum of antioxidant activities. Of the compounds with identified functions, bakuchiol (Bluemke et al. 2022; Ma et al. 2020), 25S-inokosterone (Liu et al. 2022), and 4-hydroxy ginkgolic acid (Noor-E-Tabassum et al. 2022) are present in the tapuy wine and have antioxidative properties. In the tapuy lees, 6-hydroxykaempferol (Chen & Chen 2014; Ren et al. 2019; Silva dos Santos et al. 2021), cholestenone (Ramalingam & Rajaram 2018), flazin (Fuda et al. 2019; Wu et al. 2022), and ganoderic acid B (Cör et al. 2018; Sheikha 2022) are present and have antioxidative properties. Furthermore, the total number of spectral peaks matched to the database represent only a fraction of the total number of spectral peaks of the two samples. The use of a more exhaustive library or multiple libraries may identify more of the peaks can be identified as to what compounds they are.

Chromatogram of the base peak intensity vs. retention time of tapuy wine sample. Identified analytes are shown labelled. There are 9 identified analytes from 43 spectral peaks

Chromatogram of the base peak intensity vs. retention time of tapuy lees sample. Identified analytes are shown labelled. There are 14 identified analytes from 113 spectral peaks
The antioxidant assays only measure a fraction of the potential antioxidant activity of tapuy lees
There is a significant difference between the mean phenolic content of tapuy wine and lees as shown in Fig. 3A. The tapuy lees has a 6.5-fold greater mean phenolic content than the tapuy wine. This could be attributed to a higher occurrence of non-polar phenolic compounds (as reported in previous studies) remaining in the tapuy lees, because they have a low dissolution in wine. Phenolic compounds such as triterpenoids (Vrancheva et al. 2021), terpenes (Jiang et al. 2016), and steroids (Hartonen & Riekkola 2017) are non-polar. The high abundance of these compounds in tapuy lees has been shown to have many health benefits such as anti-inflammation activity (Karimi et al. 2012; Kumar et al. 2019), anti-cancer activity (Badhani et al. 2015; Karimi et al. 2012; Kumar et al. 2019; Mori et al. 1999; Rocha et al. 2012), antioxidant activity (Badhani et al. 2015; Karimi et al. 2012; Medina et al. 2007; Middleton et al. 2002), antifungal activity (Badhani et al. 2015), antibacterial activity (Badhani et al. 2015; Bhattacharya et al. 2010; Bodini et al. 2009), antiviral activity (Badhani et al. 2015).

Phenolic content of tapuy wine and lees and their antioxidant activities. A *** indicates a p-value less than 0.001 and an n.s. indicates no significant difference. a Mean phenolic content of tapuy wine and lees. The phenolic content was determined using the Folin-Ciocalteu assay. b DPPH assay of tapuy wine and lees (IC50). IC50 values were determined using a DPPH assay. The IC50 values indicate the minimum concentration of the wine or lees sample that elicits a 50% inhibition of decolorization of DPPH from a purple to a clear solution. c FRAP assay of tapuy wine and lees (EC50). EC50 values were determined using a FRAP assay. The EC50 values indicate the minimum concentration of the wine or lees sample that elicits a 50% change from clear to an intense, blue-colored solution
Figure 3B and C show that according to the positive controls of the DPPH (ascorbic acid) and FRAP (butylated hydroxytoluene) assays, the tapuy lees had higher EC50 and IC50 values than the tapuy wine. Interestingly, higher phenolic content and greater phenolic compound variety in the tapuy lees than the tapuy wine did not translate to better tapuy lees antioxidant activity as measured by the DPPH and FRAP antioxidant assays employed. Phenolic compounds found in the tapuy lees are generally non-polar and are not easily miscible in an aqueous solution. In contrast, the phenolic contents of wine are generally polar and easily mix in aqueous solutions. Thus, the DPPH and FRAP assays measured only the phenolic compounds present in the aqueous extract of the tapuy lees, which may not be indicative of the total phenolic content as most of these would be trapped within the physical tapuy lees. This is in contrast to the tapuy wine in which all of its phenolic contents will already be suspended in solution. An extraction process that will give a phenolic amount in the extracts that is more proportionate of the actual phenolic content of the source may reflect the total antioxidant activities of the tapuy lees more accurately.
Alternatively, it is also possible that the two assays employed only measure the antioxidant activities of a limited class of phenolic compounds. Therefore, the use of other antioxidant assays such as the HORAC, ORAC, TEAC, TAC, TRAP, CUPRAC, TOSC, and ABTS, which measure other classes of phenolic compounds, may yield a more complete and accurate measure of the total antioxidant activity of the samples derived from phenolic compounds. Each antioxidant assay targets a specific method of antioxidant activity such as hydrogen atom transfer (ORAC, TRAP, and HORAC), the transfer of one electron (CUPRAC and FRAP), and mixed tests (ABTS and DPPH) (Munteanu & Apetrei 2021). Each of these methods has its own pros and cons and has the capability to detect different classes of antioxidants based on their mechanism of action. Thus, the usage of only FRAP and DPPH may be a limitation in the determination of the total antioxidant activity of tapuy wine and lees.
Tapuy lees supplementation can moderately increase the mean lifespan of C. elegans
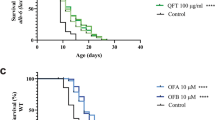
Antioxidants are tightly associated with an increased lifespan (Ding & Zhao 2022; Duangjan et al. 2019; Lin et al. 2019; Rangsinth et al. 2019). We have shown that tapuy lees has high phenolic content and thus, will have a high potential for antioxidant activity. With this, we sought to determine the effect of tapuy supplementation on the lifespan of C. elegans. Since the tapuy wine and lees are rich in potential antioxidants due to its high total phenolic content, we wanted to determine if supplementation with tapuy wine and lees can lead to increased lifespan in C. elegans. Figure 4 shows the Kaplan–Meier survival curves of various experimental groups. Overall, there is no significant difference between high and low concentrations of tapuy wine, lees, and with the no supplementation group (negative control). However, a slight increase in survival is seen in low concentration supplementation of Tapuy wine or lees. Supplementation with high concentration Tapuy wine and lees has slightly worse percent survival compared to the negative control. This could be attributed to two possible reasons. First, the high concentration of antioxidant compounds converts them into functional prooxidants (Sotler et al. 2019). This could explain why the higher concentration of Tapuy wine and lees extracts results in a decreased lifespan compared to the low concentration and the negative control. Second, the high consumption of either tapuy wine or lees may lead to an excessive accumulation of sugars. A high-sugar diet is associated with a decrease in the lifespan of C. elegans (Alcántar-Fernández et al. 2018; Lee et al. 2009). A high-sugar diet in Drosophila melanogaster is also associated with decreased lifespan (Baenas & Wagner 2022). Musselman et al. (2013) report that while triacylglyceride storage in high-sugar diets is a protective mechanism, the synthesis of more triacylglycerides over the capacity of the fly’s fat body would lead to dysregulated lipogenesis. A high-sugar diet is, in general, adverse to the health of mammalians (Kelm et al. 2011). An increase in sugar consumption and thus, lipogenesis, translates into an increased risk for diseases that involve high-fat concentrations, such as cardiovascular diseases and obesity (Simons et al. 2022; Stanhope 2016). Similarities in lipogenesis dysregulation due to high-sugar diets show the translatability of diet-based studies from animal models to humans.

Kaplan–Meier survival curves for Caenorhabditis elegans fed with varying amounts of tapuy wine or lees or fed without tapuy supplementation. A risk table showing the percentage of the population at risk is included below the Kaplan–Meier survival curves. Results of lifespan assays of three trials were pooled together to form one survival curve per experimental group. Sample sizes are n = 80 for the tapuy lees high, n = 83 for the tapuy lees low, n = 82 for the tapuy wine high, n = 77 for the tapuy wine low, and n = 82 for the no supplementation group. The concentrations of tapuy wine and lees high are 50,000 µg/mL. The concentrations of tapuy wine and lees low are 5,000 µg/mL
Our experiments demonstrate that high tapuy lees supplementation increases the mean lifespan of C. elegans by 16.24% compared to no supplementation, albeit below statistical significance. However, the trend suggests that if we increase the tapuy lees dosage in the C. elegans diet, we will eventually reach statistical significance (Fig. 5). This is consistent with the observed lifespan extension seen in Drosophila melanogaster (Dela Cruz, in preparation) fed with tapuy lees. Though high sugar consumption is associated with high supplementation of either tapuy wine or lees, there were no significant adverse effects on the mean lifespan of C. elegans. The other metabolites present in the tapuy lees (especially the high mean phenolic content) may contribute enough health benefits to counteract the potential lifespan-shortening effects of a high-sugar content with tapuy lees. The high sugar content may even be desirable due to the increase in palatability conferred by a sweeter taste. Since both C. elegans and humans follow the same trend of decreased lifespan induced by a high-sugar diet, C. elegans is a good model for lifespan-affecting diet assays. Therefore, it is likely that the moderate increase in the lifespan of C. elegans induced by high tapuy lees supplementation may also be observed in humans. It can be observed that in the Kaplan–Meier survival curve, a few C. elegans were able to survive when supplemented with low dosage of tapuy wine or lees as compared to C. elegans supplemented with high dosage of tapuy wine or lees. The few individuals that did survive longer in the Kaplan–Meier survival curve may have been outliers. However, when determining the overall effect on lifespan of tapuy supplementation on all the C. elegans in each treatment group by taking the mean lifespan, it was revealed that the high dosage of tapuy lees slightly increases the lifespan the most compared to other treatment groups and the negative control. The mean lifespan of the C. elegans supplemented with a high dosage of tapuy lees had a 16.24% increase in lifespan compared to C. elegans not fed with either tapuy wine or lees (i.e. negative control).

Mean lifespan of Caenorhabditis elegans supplemented with varying amounts of either tapuy wine or lees extracts. Only high supplementation of tapuy lees led to a moderate increase in mean lifespan over other worms. The concentrations of tapuy wine and lees high are 50,000 µg/mL. The concentrations of tapuy wine and lees low are 5,000 µg/mL
Conclusion
Through fermentation technology, we were able to enhance the quality of the nutrients of a substrate (rice) in the form of tapuy wine and lees. Our experiments have revealed the nutritional value of tapuy lees, with a high protein content, low-fat content, moderate sugar content, and high phenolic content (Table 1 and Fig. 3). We found that tapuy lees has a wider variety of compounds identified via LC–MS/MS than tapuy wine and many of these compounds have been linked to beneficial health effects. We recommend that further analysis be done to identify unidentified peaks and conduct functional screening of these putative compounds to identify their biofunctionalities. The two antioxidant assays employed in the study may not be indicative of the actual antioxidant activity of tapuy lees. There are two options to address this limitation. First, by employing a more comprehensive analysis of its antioxidant activities through a wider range of antioxidant assays. Second, to utilize a more efficient extraction technique for tapuy lees that would also efficiently extract the non-polar phenolic components from the substrate. Interestingly, even though the measured antioxidant activity of the tapuy lees is lower than the wine, albeit having a higher phenolic content than the tapuy wine, functional lifespan assays performed on C. elegans show that tapuy lees supplementation led to a higher increase in lifespan than tapuy wine supplementation. Furthermore, worms supplemented with high amounts of tapuy lees had a slightly higher mean lifespan than both tapuy wine supplementation and with no supplementation. Establishing dose dependency in increasing percent survival and mean lifespan may be of great value in recommending an ideal amount for consumption. Overall, we show that tapuy lees, although heavily underutilized and wasted, is a healthy food source. Repurposing the tapuy lees into a food product and its consumption may nutritionally enhance consumer diet.
Availability of data and materials
Data is available upon reasonable request.
References
Alcántar-Fernández, J., et al. (2018). Caenorhabditis elegans respond to high-glucose diets through a network of stress-responsive transcription factors. PLoS ONE, 13(7), e0199888.
Allwood, J. G., Wakeling, L. T., & Bean, D. C. (2021). Fermentation and the microbial community of Japanese koji and miso: A review. Journal of Food Science, 86(6), 2194–2207.
Alrosan, M., et al. (2021). Effects of fermentation on the quality, structure, and nonnutritive contents of lentil (Lens culinaris) Proteins. Journal of Food Quality, 2021, 1–7.
Badhani, B., Sharma, N., & Kakkar, R. (2015). Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Advances, 5(35), 27540–27557.
Baenas, N., & Wagner, A. E. (2022). Drosophila melanogaster as a model organism for obesity and type-2 Diabetes Mellitus by applying high-sugar and high-fat diets. Biomolecules, 12(2), 307.
Benabdallah, A., et al. (2016). Total phenolic content and antioxidant activity of six wild Mentha species (Lamiaceae) from northeast of Algeria. Asian Pacific Journal of Tropical Biomedicine, 6(9), 760–766.
Bhattacharya, A., Sood, P., & Citovsky, V. (2010). The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Molecular Plant Pathology, 11(5), 705–719.
Bluemke, A., et al. (2022). Multidirectional activity of bakuchiol against cellular mechanisms of facial ageing - Experimental evidence for a holistic treatment approach. International Journal of Cosmetic Science, 44(3), 377–393.
Bodini, S. F., et al. (2009). Quorum sensing inhibition activity of garlic extract and p-coumaric acid. Letters in Applied Microbiology, 49(5), 551–555.
Bueno, P., et al. (2013). Antioxidant activity of Ficus pseudopalma Blanco and its cytotoxic effect on hepatocellular carcinoma and peripheral blood mononuclear cells. Current Research in Biological and Pharmaceutical Sciences, 2, 14–21.
Çabuk, B., et al. (2018). Effect of fermentation on the protein digestibility and levels of non-nutritive compounds of pea protein concentrate. Food Technology and Biotechnology, 56(2), 257–264.
Chaudhuri, R. K., & Bojanowski, K. (2014). Bakuchiol: A retinol-like functional compound revealed by gene expression profiling and clinically proven to have anti-aging effects. International Journal of Cosmetic Science, 36(3), 221–230.
Chen, S. (2020). The pharmacological effects of triterpenoids from Ganoderma lucidum and the regulation of its biosynthesis. Advances in Biological Chemistry, 10, 55–65.
Chen, A. Y., & Chen, Y. C. (2014). A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chemistry, 138(4), 2099–2107.
Cör, D., Ž. Knez, and M. Knez Hrnčič (2018). Antitumour, antimicrobial, antioxidant and antiacetylcholinesterase effect of ganoderma lucidum terpenoids and polysaccharides: A review. Molecules, 23(3), 649.
Dhaliwal, S., et al. (2019). Prospective, randomized, double-blind assessment of topical bakuchiol and retinol for facial photoageing. British Journal of Dermatology, 180(2), 289–296.
Diether, N. E., & Willing, B. P. (2019). Microbial fermentation of dietary protein: An important factor in diet–microbe–host interaction. Microorganisms, 7(1), 19.
Dimidi, E., et al. (2019). Fermented foods: definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients, 11(8), 1806.
Ding, F., & Zhao, Y. (2022). Astaxanthin induces transcriptomic responses associated with lifespan extension in caenorhabditis elegans. Antioxidants, 11(11), 2115.
Duangjan, C., et al. (2019). Lifespan extending and oxidative stress resistance properties of a leaf extracts from. Oxidative Medicine and Cellular Longevity, 2019, 9012396.
Dutra, L., et al. (2014). Ent-Kaurane diterpenes from the stem bark of Annona vepretorum (Annonaceae) and cytotoxic evaluation. Bioorganic & Medicinal Chemistry Letters, 24, 3315–3320.
El Sheikha, A. F. (2022). Nutritional Profile and Health Benefits of Ganoderma lucidum “Lingzhi, Reishi, or Mannentake” as functional foods: Current scenario and future perspectives. Foods, 11(7), 1030.
Fuda, H., et al. (2019). Flazin as a promising Nrf2 pathway activator. Journal of Agricultural and Food Chemistry, 67(46), 12844–12853.
Hartonen, K., & Riekkola, M.-L. (2017). Chapter 2 - water as the first choice green solvent. In F. Pena-Pereira & M. Tobiszewski (Eds.), The application of green solvents in separation processes. (pp. 19–55). Elsevier.
Hua, Z., et al. (2017). The antibacterial activity and mechanism of ginkgolic acid C15:1. BMC Biotechnology, 17(1), 5.
Huang, P., et al. (2022). The Akebia Genus as a Novel Forest Crop: A review of its genetic resources, nutritional components, biosynthesis, and biological studies. Frontiers in Plant Science, 13, 936571.
Ito, K. R., et al. (2022). Utilization of sake lees as broiler feedstuff and its effects on growth performance and intestinal immunity. The Journal of Poultry Science, 59(3), 247–259.
Jackson, R. F., & Mcdonald, E. J. (1941). Errors of Munson and Walker’s reducing-sugar tables and the precision of their method. Journal of Association of Official Agricultural Chemists, 24(3), 767–788.
Jiang, Z., Kempinski, C., & Chappell, J. (2016). Extraction and analysis of terpenes/terpenoids. Current Protocols in Plant Biology, 1, 345–358.
Karimi, E., et al. (2012). Phenolic compounds characterization and biological activities of Citrus aurantium bloom. Molecules, 17(2), 1203–1218.
Kelm, D. H., et al. (2011). High activity enables life on a high-sugar diet: Blood glucose regulation in nectar-feeding bats. Proceedings of the Royal Society b: Biological Sciences, 278(1724), 3490–3496.
Kim, J. W., et al. (2016). Anti-neuroinflammatory ent-kaurane diterpenoids from pteris multifida roots. Molecules, 22, 27.
Kobayashi, J., et al. (2021). Cholestenone functions as an antibiotic against. Proceedings of the National Academy of Sciences U S A, 118(16), e2016469118.
Kong, Y., et al. (2021). Immunomodulatory effects of flazin from Crassostrea sikamea on splenic lymphocytes of Sprague-Dawley rats. Chinese Journal of Natural Medicines, 19(11), 836–843.
Kumar, N., et al. (2019). Extrapolation of phenolic compounds as multi-target agents against cancer and inflammation. Journal of Biomolecular Structure & Dynamics, 37(9), 2355–2369.
Kwon, D. Y., Nyakudya, E., & Jeong, Y. S. (2014). Fermentation: Food products. In N. K. Van Alfen (Ed.), Encyclopedia of agriculture and food systems (pp. 113–123). Academic Press.
Kwon, M., et al. (2020). Kushenol E inhibits autophagy and impairs lysosomal positioning via VCP/p97 inhibition. Biochemical Pharmacology, 175, 113861.
Lee, S.-J., C.T. Murphy, and C. Kenyon (2009). Glucose shortens the life span of C. elegans by downregulating DAF-16/FOXO activity and aquaporin gene expression. Cell Metabolism, 10(5), 379–391.
Lin, C., et al. (2019). Carnosol improved lifespan and healthspan by promoting antioxidant capacity in. Oxidative Medicine and Cellular Longevity, 2019, 5958043.
Liu, Y., et al. (2022). Inokosterone from Gentiana rigescens Franch extends the longevity of yeast and mammalian cells via antioxidative stress and mitophagy induction. Antioxidants (Basel), 11(2), 214.
Ma, W., et al. (2020). Bakuchiol alleviates hyperglycemia-induced diabetic cardiomyopathy by reducing myocardial oxidative stress via activating the SIRT1/Nrf2 signaling pathway. Oxidative Medicine and Cellular Longevity, 2020, 1–15.
Maciąg, D., et al. (2021). Akebia quinata and Akebia trifoliata - a review of phytochemical composition, ethnopharmacological approaches and biological studies. Journal of Ethnopharmacology, 280, 114486.
Manaois, R. V., & Morales, A. V. (2014). Evaluation of tapuy lees as a functional ingredient in the snack food polvoron. Journal of Food Quality, 37(3), 196–202.
Mango, D., Weisz, F., & Nisticò, R. (2016). Ginkgolic acid protects against Aβ-induced synaptic dysfunction in the hippocampus. Frontiers in Pharmacology, 7, 401.
Mazorra-Manzano, M. A., et al. (2020). Cheese whey fermentation by its native microbiota: proteolysis and bioactive peptides release with ACE-inhibitory activity. Fermentation, 6(1), 19.
Medina, I., et al. (2007). Effect of molecular structure of phenolic families as hydroxycinnamic acids and catechins on their antioxidant effectiveness in minced fish muscle. Journal of Agriculture and Food Chemistry, 55(10), 3889–3895.
Middleton, E., Kandaswami, C., & Theoharides, T. (2002). The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease and cancer. Pharmacological Reviews, 52, 103–108.
Morales, A. and R. Manaois (2011). Rice wine lees, a by-product of tapuy fermentation, for rice cookies. Philippine Journal of Crop Science (Philippines).
Mori, H., et al. (1999). Chemopreventive effects of ferulic acid on oral and rice germ on large bowel carcinogenesis. Anticancer Research, 19(5A), 3775–3778.
Motono, Y., et al. (2021). Effect of sake lees on the inhibition of lipid accumulation in adipocytes. Fermentation, 7(3), 145.
Munteanu, I. G., & Apetrei, C. (2021). Analytical methods used in determining antioxidant activity: a review. International Journal of Molecular Sciences, 22(7), 3380.
Musselman, L. P., et al. (2013). Role of fat body lipogenesis in protection against the effects of caloric overload in Drosophila. Journal of Biological Chemistry, 288(12), 8028–8042.
Nagao, K., et al. (2022). The cholesterol metabolite cholest-5-en-3-one alleviates hyperglycemia and hyperinsulinemia in obese (db/db) mice. Metabolites, 12(1), 26.
Neuvonen, M., et al. (2014). Enzymatic oxidation of cholesterol: Properties and functional effects of cholestenone in cell membranes. PLoS ONE, 9(8), e103743.
Nhiem, N. X., et al. (2014). New ent-kauranes from the fruits of Annona glabra and their inhibitory nitric oxide production in LPS-stimulated RAW264.7 macrophages. Bioorganic & Medicinal Chemistry Letters, 25, 254–258.
Noor-E-Tabassum, et al. (2022). Ginkgo biloba: A treasure of functional phytochemicals with multimedicinal applications. Evidence-based Complementary and Alternative Medicine, 2022, 8288818.
Peng, B., et al. (2021). The nutritional value and application of black rice-a review. Journal of Biotechnology Research, 7(4), 63–72.
Ramalingam, V., & Rajaram, R. (2018). 2-Ethoxycarbonyl-2-β-hydroxy-a-nor-cholest-5-ene-4one: Extraction, structural characterization, antimicrobial, antioxidant, anticancer and acute toxicity studies. Steroids, 140, 11–23.
Rangsinth, P., et al. (2019). Leaf extract of Caesalpinia mimosoides enhances oxidative stress resistance and prolongs lifespan in Caenorhabditis elegans. BMC Complementary and Alternative Medicine, 19(1), 164.
Rathna Priya, T. S., et al. (2019). Nutritional and functional properties of coloured rice varieties of South India: A review. Journal of Ethnic Foods, 6(1), 11.
Reddy, D., et al. (2020). Anticancer and antiviral properties of cardiac glycosides: a review to explore the mechanism of actions. Molecules, 25(16), 3596.
Ren, J., et al. (2019). Recent progress regarding kaempferol for the treatment of various diseases (Review). Experimental and Therapeutic Medicine, 18(4), 2759–2776.
Rocha, L., Monteiro, M., & Teodoro, A. (2012). Anticancer properties of hydroxycinnamic acids -a review. Cancer and Clinical Oncology, 1, 1–11.
Şanlier, N., Gökcen, B. B., & Sezgin, A. C. (2017). Health benefits of fermented foods. Critical Reviews in Food Science and Nutrition, 59(3), 506–527.
Shahrajabian, M. H., Sun, W., & Cheng, Q. (2021). Improving health benefits with considering traditional and modern health benefits of Peganum harmala. Clinical Phytoscience, 7(1), 18.
Sharma, R., et al. (2020). Microbial fermentation and its role in quality improvement of fermented foods. Fermentation, 6(4), 106.
Shimoga, G. and S.-Y. Kim (2021). Makgeolli - The traditional choice of korean fermented beverage from cereal: An overview on its composition and health benefits. Food Science and Technology, 42, e43920.
Silva dos Santos, J., et al. (2021). The pharmacological action of kaempferol in central nervous system diseases: a review. Frontiers in Pharmacology, 11, 565700.
Simons, P. I. H. G., et al. (2022). Association between de novo lipogenesis susceptibility genes and coronary artery disease. Nutrition, Metabolism and Cardiovascular Diseases, 32(12), 2883–2889.
Sotler, R., et al. (2019). Prooxidant activities of antioxidants and their impact on health. Acta Clinica Croatica, 58(4), 726–736.
Stanhope, K. L. (2016). Sugar consumption, metabolic disease and obesity: The state of the controversy. Critical Reviews in Clinical Laboratory Sciences, 53(1), 52–67.
Tang, J.-G., et al. (2008). Synthesis of analogues of flazin, in particular, flazinamide, as promising anti-HIV agents. Chemistry & Biodiversity, 5(3), 447–460.
Toksoy Oner, E., S.G. Oliver, and B. Kirdar (2005). Production of ethanol from starch by respiration-deficient recombinant Saccharomyces cerevisiae. Applied and Environmental Microbiology, 71(10), 6443–5.
Tsutsui, N., Yamamoto, Y., & Iwami, K. (1998). Protein-nutritive assessment of sake lees obtained by brewing from liquefied rice. J Nutr Sci Vitaminol (tokyo), 44(1), 177–186.
Vrancheva, R., et al. (2021). Triterpenoids and other non-polar compounds in leaves of wild and cultivated. Plants (Basel), 10(1), 94.
Wu, X., et al. (2022). Flazin as a lipid droplet regulator against lipid disorders. Nutrients, 14(7), 1501.
Zhao, P., et al. (2018). Antifatigue and antiaging effects of Chinese rice wine in mice. Food Science & Nutrition, 6(8), 2386–2394.
Acknowledgements
We thank the following research assistants: Ismael H. Velasco III, Jury Rex Villafuerte-Flor, John Christian H. Arrogante, Carlito M. Lamarca, Charles Jason Jimenez, Paolo Vidal, and Joseph Conrad M. Libunao for their help in the completion of this research.
We would like to thank the Philippine Institute of Traditional and Alternative Health Care (PITAHC) for funding this research project.
Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40OD010440).
Compliance with ethical standards
All authors comply with BMC’s ethical policies regarding conflicts of interest, informed consent, and ethical treatment of humans and animals in research.
Funding
This study was funded by the Philippine Institute of Traditional and Alternative Health Care (PITAHC).
Author information
Authors and Affiliations
Contributions
Paul Mark B. Medina contributed the following: Conceptualized the research project. Managed the administration of the research project. Implemented the research project. Analyzed the data. Wrote the manuscript. Sean Philippe L. Chua contributed the following: Analyzed the data. Wrote the manuscript. Lesley Dale Umayat contributed the following: Conceptualized the research project. Managed the administration of the research project. Implemented the research project.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
The authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
Cite this article
Chua, S.P.L., Umayat, L.D. & Medina, P.M.B. Phenolic-rich lees from Philippine rice wine (tapuy) increases Caenorhabditis elegans lifespan in spite of having low antioxidant activity. Food Prod Process and Nutr 6, 1 (2024). https://doi.org/10.1186/s43014-023-00181-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43014-023-00181-w