
Abstract
Background
The cultural connections of the Maidu to the lands they inhabit are profound with burning being a major component of their culture. California black oak plays an important role in the lifeways of many Indigenous tribes and Native peoples tend black oaks. We used multiple lines of evidence to reconstruct Indigenous fire use in a mixed conifer forest in the northern Sierra Nevada. This includes summarizing oral traditions by the Mountain Maidu, quantifying current and historical forest structure, and characterizing fire effects in an area impacted by the 2021 Dixie Fire.
Results
The promotion of fire, both lighting- and human-ignited, has always been part of Mountain Maidu land stewardship. The mixed conifer forests in our focal study site had very high tree densities that were dominated by small, shade-tolerant species and the historic mean fire return interval was 8.6 years. Twenty-three fire intervals of 10 years or less and 11 fire intervals of 5 years or less were recorded. Based on forest inventory data collected in 1924, a majority of the forests in the broader region surrounding our focal site (82%) did not experience high levels of conifer competition. This is a stark contrast to contemporary forest conditions which showed that 73% of forests either were experiencing full site occupancy or were within the zone of imminent mortality. Contemporary forests not experiencing high levels of conifer competition had the highest density of large California black oak while forests in the zone of imminent mortality had the lowest density of large California black oak.
Conclusions
Fire exclusion and suppression led to a slow, albeit catastrophic, shift for California black oaks. The mixed conifer forests sampled in the early 1920s had very low tree densities and were dominated by large ponderosa and sugar pine trees which would have allowed California black oak to thrive. The number of very short fire intervals recorded is difficult to explain by lightning alone, pointing to the management of this area for Indigenous purposes. The 2021 wildfire resulted in 100% mortality of all conifer trees in the area but 61% of the California black oaks resprouted. This provides an opportunity to remove the conifer snags and allow the California black oaks to regain dominance. It would take active stewardship to produce the oak dominance again but it is possible and could be co-led by the Maidu and federal managers.
Resumen
Antecedentes
Las conexiones culturales de los indígenas Maidu de California con las tierras que ellos habitan son muy profundas, siendo las quemas de bosques un componente mayor de su cultura. El roble negro de California juega rol importante en los estilos de vida de muchas tribus, y los pueblos nativos cuidan de los robles negros. Usamos múltiples líneas de evidencias para reconstruir el uso indígena del fuego en un bosque mixto de coníferas en el norte de las Sierras Nevadas. Esto incluye resumir tradiciones orales de los Maidu de las Montañas, cuantificando la estructura actual e histórica, y caracterizamos los efectos del fuego en un área impactada por el incendio de Dixie de 2021.
Resultados
La promoción de los incendios, tanto iniciados por rayos como por la actividad humana, ha sido siempre parte de la administración de tierras por los Maidu. El bosque mixto en nuestro estudio focal tenía una alta densidad de árboles y era dominada por especies pequeñas y tolerantes a la sombra, y su intervalo de retorno del fuego era de 8,6 años. Se registraron veintitrés intervalos de 10 años o menos y 11 intervalos de fuego de 5 o menos años. Basados en datos de inventario forestal colectados en 1924, una gran mayoría de los bosques en una amplia región alrededor de nuestro sitio focal (82%) no experimentó competencia total por parte de las coníferas. Esto representa un contraste muy rígido con las condiciones contemporáneas, que muestran que el 73% del bosque está experimentando o una ocupación total del sitio o están dentro de una zona de inminente mortalidad. Los bosques contemporáneos que no experimentan una competencia total por parte de las coníferas tienen la mayor densidad de robles negros de California, mientras que la zona de inminente mortalidad tiene la menor densidad de estos robles negros.
Conclusiones
La exclusión o supresión del fuego ha llevado a un lento, aunque catastrófico cambio para los robles negros de California. Los bosques de coníferas muestreados a inicios de los 1920s, tenían una baja densidad de árboles y estaban dominados por pinos ponderosa grandes y por el pino dulce gigante (Pinus lambertiana) que podrían haber permitido prosperar a los robles negros de California. El número de fuegos registrados a intervalos muy cortos es difícil de explicar basado solo en igniciones por rayos, por lo que se deduce se debió al manejo del área para propósitos indígenas. El incendio de 2021 resultó en un 100% de mortalidad para todas las coníferas, mientras que el 61% de los robles rebrotaron. Esto provee una oportunidad para remover todas las coníferas muertas en pie y permitir a los robles reganar su dominancia. Esto necesitará de una gestión activa para producir nuevamente la dominancia de los robles, pero es posible y debe ser co-liderado por los Maudi y los gestores federales.
Similar content being viewed by others

Background
Mixed conifer, ponderosa pine (Pinus ponderosa), and Jeffrey pine (Pinus jeffreyi) forests in California are adapted to frequent, low-moderate-intensity fire regimes (Safford and Stevens 2017). These forests are dominated by conifers but also include hardwood species such as California black oak (Quercus kelloggii). This oak species has the widest distribution and range of all the oaks in the western US, extending over 1250 km from Oregon to Baja California, Mexico (McDonald et al. 1990). California black oak plays an important role in the lifeways of many Indigenous tribes in California (O’Gorman et al. 2022). Acorns produced by trees of Quercus and Notholithocarpus are used as a food source by many Indigenous peoples (Anderson 2005) making them important management priorities.
During the mid-1800s, the opportunity and desire for gold, timber, and land by colonists pushed Indigenous people, including the Maidu, to the brink of extinction and away from their ancestral way of life. Unlike in other parts of the western US, it was found to be much easier to simply remove the Natives from their land by any means necessary, rather than ratify treaties that would create reservations (Kelsey 1973).
Another toll was taken on Indigenous populations with the creation of the national Indian boarding school system (Haag 2007). Located on the north side of what is now called Indian Valley, the Greenville Indian boarding school in Plumas County impacted Native people terribly. Constructed in 1890, the Indian students had finally had enough and took it upon themselves to burn the school to the ground in 1920, leaving behind three decades of abuse and the legacy of separating children from their cultural heritage. The cultural connections of the Maidu to the lands they inhabit are profound with fire being a major component of their culture and the disruption of these linkages has impacted other tribes (Steen-Adams et al. 2019).
Indigenous burning has many objectives and the extent of burning is controlled by careful timing, reduced fuel loads, and the use of fire breaks such as trails, creeks, streams, rivers, and lakes. One of the most important outcomes of Indigenous burning was the intentional creation of vegetation patches with different species composition and successional fuel stages. Across the landscape, the mosaic of differing vegetation patches influenced fuel and flammability characteristics, which would itself strongly limit the extent of subsequent fires (Kimmerer and Lake 2001). This mosaic also promoted food security by ensuring a diverse and productive landscape (Lewis 1985). California black oaks are “cultural keystone” species due to their role in Indigenous peoples’ diet, ceremonial practices, and daily lives (Garibaldi and Turner 2004; Long et al. 2016, 2017). Regarding food sources, California black oak acorns are highly desired for their flavor, processing ease, and their stability for long-term storage (Long et al. 2016).
In California, despite calls to incorporate more fire use in land stewardship, there are only a few examples of restored fire regimes in frequent-fire forest types. These examples consist of a few isolated areas that have implemented wilderness fire programs (managed wildfires for resource objectives ignited by lightning) for several decades (Parks et al. 2015, North et al. 2021, Stephens et al. 2021a). Outside of these isolated areas, fire suppression and timber harvesting that focused on large tree removal have spurred massive infilling of small- and intermediate-sized trees (Knapp et al. 2013; Collins et al. 2017). Fire exclusion has also enabled conifer encroachment in previously hardwood-dominated areas resulting in the development of narrow-crowned trees with high branches (Fig. 1; Cocking et al. 2012) that are less conducive for Indigenous cultural uses (Long et al. 2021).

A California black oak tree in our study area overtopped by conifers after the 2021 Dixie Fire, B ponderosa pine fire history sample from the study area with fire dates in white, and C California black oak in our study area that resprouted after the 2021 Dixie Fire
Fire regimes can be reconstructed from fire scars using standard dendro-ecological techniques (Dieterich 1980). Fire scars in coastal California redwood (Sequoia sempervirens) forests are thought to be created by Indigenous burning since lighting-ignited fires are rare in this region (Finney and Martin 1992; Stephens and Fry 2005). In inland areas of California and elsewhere, it is difficult to distinguish Indigenous and lightning-ignited fires in the tree ring record (van Wagtendonk and Cayan 2008). One of the main reasons for this is lightning and Indigenous fires overlap in terms of seasonality in many areas (Anderson 2005).
Two fire history studies outside of coastal California have found direct evidence of the impacts of Indigenous burning. The first study in ponderosa pine forests in the north coast range found a distinct fire seasonality shift before and after 1850 (Skinner et al. 2009). Before 1850, the seasonality of fires, inferred from intra-ring fire scar positions, was 40% earlywood and 50% latewood; after 1850, this changed to 55% latewood and 30% dormant season. Earlywood scar positions tend to correspond with May–July calendar months, latewood positions July–September, and dormant positions September–October, but could also include the winter and early spring (Stephens et al. 2018). Early season fires recorded before 1850 were likely Indigenous due to this site’s proximity to the expansive grass and oak woodlands of the Sacramento Valley. After 1850 when tribes were impacted by colonization, the fires were dominated by lightning ignitions, but were also influenced by intentional ignitions to improve forage for livestock grazing (Whittaker 1960) and ignitions from mining and early timber harvesting activities (Leiberg 1902). Fire frequency in ponderosa pine forests in this area of the north coast range was also impacted by Indigenous ignitions where the study recorded more frequent fires before 1850 (before the gold rush) as compared to other mixed conifer forested areas in northwestern California (Skinner et al. 2009).
The second study that found a fire seasonality shift is in Jeffrey pine-mixed conifer forests in the Sierra San Pedro Martir (SSPM) in northwestern Mexico (Evett et al. 2007). This study demonstrated that contemporary lightning occurrence, which is concentrated in the summer with little spring activity, is insufficient to support the high number and spring fires recorded in the past. Changes in the ignition regime during the last 300 years could have been due to Indigenous burning and climatic factors; however, available evidence suggests that periods of the most frequent fires were dominated by Indigenous ignitions (Evett et al. 2007). This study found that there was an order of magnitude more May and June fires during periods that pre-date colonization. Archaeological evidence indicates there was considerable seasonal use of fire in the SSPM by Native populations (Meigs 1935) and in the surrounding region (Bean and Lawton 1993).
The objective of this study is to characterize and explain the degradation of a forest valued by the Mountain Maidu people, based upon multiple lines of evidence, and suggest appropriate management strategies to reverse that degradation. This includes summarizing oral traditions by the Mountain Maidu, quantifying current and historical forest structure, and characterizing fire effects in an area impacted by the 2021 Dixie Fire. We also integrate these disparate data sources and discuss their implications for restoring California black oak Indigenous stands in the wake of contemporary large-scale wildfires.
Methods
Study site
From time immemorial, the Mountain Maidu people have inhabited northeastern California including the current areas of Butte, Plumas, and Lassen Counties, an area of approximately 10,000 km2. Although the Native population is incredibly difficult to estimate, as many as 4000 Mountain Maidu once lived in this area (Plumas County Museum n.d.) whereas today considerably less than a thousand people claim Mountain Maidu ancestry, either federally recognized or non-federally recognized.
This study is centered in Meadow Valley in the northern Sierra Nevada within the Plumas National Forest at 39856′N, 12183′W. The climate is Mediterranean with a predominance of winter precipitation totaling approximately 1200 mm year–1 (Ansley and Battles 1998). Vegetation is primarily mixed conifer forest including white fir (Abies concolor), Douglas- fir (Pseudotsuga menziesii ), sugar pine (Pinus lambertiana), ponderosa pine, incense-cedar (Calocedrus decurrens), and California black oak.
Indigenous knowledge
Throughout much of the twentieth century, the academic study of ancient America focused on developing models based on archaeology and other fields of anthropology, with only sporadic attention on oral traditions (Echo-Hawk 2000). However, oral traditions provide a viable source of information about historical settings dating back far in time, a fact that has gained increasing recognition in North America (Echo-Hawk 2000). Tribal historians and religious leaders frequently rely on oral traditions as “literal records of ancient history,” while many academically trained scholars respond with skepticism of verbal literature for information over long time spans. In recent decades, however, a dramatic increase in interest among scholars in exploring oral literature for information (Echo-Hawk 2000; Anderson 2005; Ferguson et al. 2015).
The study of oral literature features two major subdivisions: oral history and oral traditions. Oral history is the verbal memoirs of firsthand observers, while oral traditions are verbal memoirs that firsthand observers have passed along to other people (Echo-Hawk 2000). Oral traditions regarding cultural burning practices of the Mountain Maidu in this paper come from a lifetime of conversation among Maidu Elders including Les Hall’s (2nd author of this paper) family. Les’s great-grandmother, Ellen Reaves (Potts, Yemie), born in Genesee Valley in 1889 (died 1977), attended the Greenville Indian school which is approximately 22 km northeast from our study area. Her cultural experiences were passed down through Les’s family and are summarized below. Consideration must be given for the incredible length of time, thousands of years, the Mountain Maidu people inhabited and maintained their place within the balance of nature in this land. This perspective of time is very difficult for non-Natives to come to terms with, given the general written history of man is much shorter. In American terms, it is common that someone with 10,000 h of experience in something is a master. What status do we give to people who have continually learned and passed down this knowledge from generation to generation for 10,000 years?
Local forest structure and fire history
To characterize the conditions of our focal study area, we measured forest structure using eight 0.04-ha circular plots installed west of Spanish Ranch in Meadow Valley, CA, approximately 0.75 km east of Burton Ridge (NE ¼, Section 9, T24N, R8E). This area had been used for 15 years as part of the curriculum at UC Berkley’s Forestry Summer camp by the first and last authors of this paper. The 2009 Silver Fire (122 ha) burned to the west of these inventory plots (Fig. 2). The focal study area also lies within the Pilot Project of the Herger-Feinstein Quincy Library Group (HFQLG) Act (Moghaddas et al. 2010). Several individual projects in the nearby area were implemented under the HFQLG Act, which mainly consisted of shaded fuel breaks and group selection silviculture. However, no activities associated with HFQLG or any other recent treatments were implemented in our focal study site. In 2010, individual plots were established using a systematic 50-m grid with a random starting point along a small ridge. Plot centers were permanently marked with a steel stake and tree species (only conifers) and DBH were recorded for all trees greater than 2.5 cm DBH. Tree density (trees ha−1) and basal area by species (m2 ha−1) were calculated to determine forest structure, which were measured annually from 2010 through 2014. These eight plots burned in the 2021 Dixie Fire and were re-measured in 2022 to assess fire-caused tree mortality. Additionally, we conducted a census of all California black oaks within a 2-ha area that surrounded these eight plots (also in 2022), recording the location (GPS coordinates) and size (DBH) of each tree along with if the tree resprouted 10 months after the 2021 wildfire (Fig. 2).

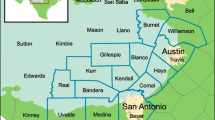
Location of forest inventory plots, fire history sample location, locations of California black oak trees, range of California black oak in California, and Mountain Maidu ancestral homeland. The perimeter of the 2009 Silver Fire is also provided. The forest in the image is before the 2021 Dixie Fire
A ponderosa pine stump with over 25 externally visible fire scars was located at the focal study site (Fig. 2). This sample was collected and sanded to a high sheen so that tree rings and fire scars could be readily distinguished under a microscope. The precise year of each fire was determined by crossdating tree rings using standard dendrochronological techniques (Stokes and Smiley 1977). The position within the ring in which a scar occurred was noted as EE (early earlywood), ME (middle earlywood), LE (late earlywood), LW (latewood), D (dormant), or U (undetermined) to serve as an estimate of the season of fire occurrence (Ahlstrand 1980; Dieterich and Swetnam 1984).
Regional historical and contemporary forest inventory data
To assess how changes in forest structure for a broader study area encompassing our focal site may limit California black oak, we used two datasets to quantify historical and contemporary forest conditions including tree density, size, and competition. Historical conditions were derived from a forest inventory that was conducted in 1924 that only included conifers. This inventory was systematically located using the Public Land Survey System (PLSS), consisting of individual samples from four parallel belt transects that were 40.2 m wide and 402 m long that ran through quarter-quarter sections (QQs; 16.2-ha or 40-acre survey units) within an individual township and range, resulting in a sampling intensity of 40% for each QQ. The archived datasheets from this survey were included in a report prepared by Brown and Brown, Inc. for Curtis, Collins & Holbrook Company, owner of the surveyed timberland in 1924. The report contained inventory data from 1552 QQs that were largely concentrated in Tehama and Plumas Counties. The bulk of these data were summarized in Collins et al. (2021); however, a subset of 71 QQs were excluded because they were geographically distinct from the main inventory area. We used the data from these 71 QQs in this study because they immediately surround our focal study area (Fig. 3). Introductory material provided in the report indicated that no logging occurred prior to the inventory being conducted (Collins et al. 2021).

Map of the study area identifying locations of the historical forest inventory data (pink) and the focal study site with contemporary field data (Silver Fire study area). The photograph to the right was taken at the time the historical inventory was conducted (1924) with the approximate location identified by converging lines from the photo to the map northeast of the Silver Fire study area. The following caption accompanied the photo: “Excellent quality large old-growth [ponderosa] pine”
The data sheets contain summaries of the transect tallies by species for a given QQ section. Our analyses were based on three attributes reported in the historical datasheets—number of trees, average tree diameter, and species. The number of trees was scaled to represent the total for the entire QQ (~16.2 ha), which we converted to trees ha−1 based on the reported area of each QQ section. The average tree diameter was reported as stump diameter, which we adjusted to the diameter at breast height (DBH) using taper equations from Wensel and Olson (1995).
To characterize regional contemporary forest conditions, we obtained plot-level data from the Forest Inventory and Analysis (FIA) program. Because FIA inventories are conducted on 10-year rotations, we subset the data to include plots that were surveyed between 2011 and 2018 (n = 14,130). We then conducted a series of filtering criteria so that FIA plots would align with similar historical site characteristics that our QQs represented. This included selecting plots that were similar in topography and historical temperature and precipitation, which are fundamental drivers of forest vegetation (Collins et al. 2021; Bernal et al. 2022). We first selected plots within the elevation range of the historical data (1104–2121 m) and removed plots where the reported elevation was different from what was extracted using elevation data from LANDFIRE (2022a). This discrepancy is due to “fuzzed” plot coordinates within approximately 0.8 km of the true plot location (Woudenberg et al. 2010), so we retained only those plots with elevation discrepancies less than 150 m (n = 1765). We also retained FIA plots that were characterized by ecological systems similar to the QQ dataset (n = 787). Ecological systems from LANDFIRE (2022b) are a classification scheme that describe the natural range of variation in plant communities based on regional distribution, vegetation physiognomy and composition, environment, and disturbance history (Comer et al. 2003). We then selected FIA plots where historical maximum summer (June–August) temperature and average winter (December–February) precipitation were within the range represented by the QQ dataset (21.4–30.2 °C and 156.8–336.8 mm, respectively) (n = 447). Historical climate data were obtained from the Basin Characterization Model (Flint et al. 2013, 2014) which provides 30-year climate averages (1920–1951) that overlap with the timing of our historical forest inventory. We also retained FIA plots where slopes that were extracted from LANDFIRE (2022c) data were within the range represented by the QQ dataset (2.17–38.36°) (n = 442). Since the lowest resolution of the spatial data we used was 270 m, we resampled datasets accordingly to 270 m.
To ensure that the FIA plots were an appropriate comparison to the QQ dataset, we continued filtering plots based on dissimilarities between the datasets. Since we wanted to compare FIA plots that represent unmanaged and undisturbed sites, we further removed an additional 62 plots for having more than one “condition” to describe the site. This would indicate that the plot is not characterized by similar age, species composition, and disturbance history (n = 380). We also removed 17 plots that had recent wildfire activity and 2 plots where the stand age was recorded as zero (only seedlings present [n = 361]). Based on the data provided for the 71 QQs, the survey was limited to trees >30.5 cm (12 in) DBH, which we presume to be the merchantable threshold at the time the inventory was conducted. The inventory was also limited to conifers and specific criteria for determining what was merchantable was not provided. As such, we removed all dead trees, as well as live trees that were <30.5 cm DBH. While no plots were entirely made up of dead trees or trees <30.5 cm DBH, we did remove 1 plot that consisted of hardwoods only (n = 360). To balance the datasets between FIA plots and the QQ dataset, our final FIA sample consisted of 71 FIA plots that were the closest to the QQ sections (Fig. 4).

Locations and overall extent of historical timber inventory conducted in 1924. The inventory was based on the Public Land Survey System, with individual quarter-quarter sections (red) as the observational unit for sampling. The dots represent Forest Inventory and Analysis plots that were surveyed between 2011 and 2018, which were used as the regional contemporary forest conditions in our analysis. They are colored based on whether black oaks (BO) were absent (blue) or present (orange) in the plot. Locations are superimposed on a topographic map provided by ESRI in ArcMap 10.7.1
For the QQ and FIA datasets, we estimated conifer density (trees ha−1) and average DBH to calculate the stand density index (SDI). SDI is a common metric in forest management that indicates stocking levels and can be used to measure inter-tree competition (North et al. 2022). We estimated SDI using the summation method:
where TPHi is the trees ha−1 represented by ploti and Di represents the average DBH (cm) of ploti. While the summation method uses individual tree diameters instead of average DBH, comparing both approaches with other historical datasets that have individual tree data yielded similar results (North et al. 2022). We then compared each QQ section and FIA plot’s absolute SDI to a theoretical maximum value that was estimated by Zhang et al. (2013, 2019) describing pine-mixed conifer forests (902 trees ha−1; 365 trees ac−1) to calculate relative SDI (% of maximum SDI). While a majority of QQ sections and FIA plots would historically be described as pine-mixed conifer, a lower maximum SDI value would yield a conservative estimate of relative SDI for plots that were not characterized by this forest type. Based on relative SDI, we assigned competition “zones” to each plot using benchmarks (Long 1985; Long and Shaw 2005) that indicate it is “free” from competition (≤25% of maximum SDI), under partial competition (25–35%), full site occupancy (35–60%), or within the “zone of imminent mortality” that is driven by density-dependent competition (≥60%).
Results
Indigenous knowledge
The promotion of fire, both natural and human-made, has always been part of Mountain Maidu land stewardship. Living in and as part of the natural environment, a symbiotic relationship between the human and non-human world was the basis for longevity lasting thousands of years. Since it is not sensible to diminish or destroy the environment that promotes healthy and sustainable life, the Indigenous people developed and maintained ways that promoted life for human and non-human alike. Fire has always been part of the natural environment (Anderson 2005) as an agent of cleansing and rebirth. With natural fire (non-human ignition) and cultural burning, the habitat of the Mountain Maidu was continually being stimulated for the enhancement of life.
Through constant promotion of fire, forest floor fuels and shrubs were never allowed to accumulate to dangerous levels, eliminating catastrophic wildfire and threats to California black oaks. Through constant promotion of fire, plants necessary for food (e.g., California black oaks), medicine, and basketry regrew strong and healthy, which led to the Mountain Maidu being excellent basket weavers from the collection of post-fire shoots that were of high quality. Through constant promotion of fire, herbivores always had plenty of new growth and animals that relied on the herbivores were able to successfully hunt their prey.
Natural fire generally occurred through summer and early fall thunderstorm lightning strikes, something that continues today. These natural fires in combination with Indigenous fire eliminated the possibility of all-consuming wildfire, regardless of weather or drought conditions. When the lightning strikes did not occur at a “good frequency,” the Mountain Maidu intentionally set fire on the land. The old people would say there was always smoke in the air somewhere from landscape fires.
Local stand structure and fire history
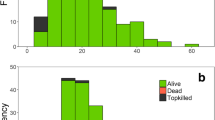
The mixed conifer forests in our focal study site located east of the 2009 Silver Fire (Fig. 2) had very high tree densities (1121 trees ha−1 including all trees > 2.5 cm DBH) (Table 1) before this wildfire burned. When the minimum DBH of trees included in the inventory was increased to > 30.5 cm DBH, this resulted in a tree density of 44 trees ha−1. Species composition was dominated by white fir, followed by Douglas- fir, and even lesser proportions of sugar pine (Table 1). No conifer trees survived the 2021 Dixie Fire, i.e., 100% mortality recorded in 2022. California black oak was quite common in the 2-ha area surrounding the eight inventory plots, with 214 trees included in the census (Fig. 2). Stem diameters varied from 4 to 18 cm DBH but most of the oaks were overtopped by conifer trees (Fig. 1). As part of the 2022 census of California black oaks, we found that 100% of the oaks had been top-killed by the 2021 Dixie Fire and 61% those oaks had sprouted from the base. This high resprouting ability agrees with other studies on California black oak (Nemens et al. 2018).
Fire history recorded in pine stump
Thirty-three fires were recorded in the ponderosa pine stump. Years when fire occurred include 1518, 1525, 1529, 1532, 1541, 1546, 1557, 1567, 1579, 1590, 1592, 1599, 1602, 1611, 1618, 1621, 1629, 1636, 1639, 1643, 1651, 1667, 1693, 1708, 1713, 1717, 1723, 1729, 1739, 1743, 1772, 1783, and 1794. The mean fire return interval was 8.6 years from 1518 to 1794; the fire interval range was 2–43 years with 23 intervals of 10 years or less, 11 intervals of 5 years or less, and one interval of 2 years (Fig. 1). Fire seasonality was 42% dormant, 21% latewood, 6% late earlywood, and 31% undetermined with the latewood fires corresponding to approximately September from previous work on fire scar phenology (Stephens et al. 2018). The last fire recorded approximately 5 km south of our ponderosa pine fire history sample was in 1893 (Stephens 2021, unpublished data).
Regional historical and contemporary forest inventory data
Average conifer tree density and average DBH for historical (1924; QQ sections) forest conditions were 53 trees ha−1 and 66 cm, while contemporary (2011–2018 FIA) forest conditions had an average of 151 trees ha−1 and 64 cm DBH when applying the common filter of only including live conifers >30.5 cm. The average relative stand density index for contemporary forests is 159% higher (~70% of maximum SDI) than what existed historically (~27% of maximum SDI) for conifers >30.5 cm DBH. A majority (82%) of QQ sections fell below the 35% benchmark (Fig. 5) indicating that these forests were not experiencing full conifer competition historically. This is in contrast to contemporary forest conditions, which showed that 73% of FIA plots were either experiencing full site occupancy (35–65% of maximum SDI [17% of plots]) or were within the zone of imminent mortality (≥60% of maximum SDI [56% of plots]) (Fig. 5). The distribution of relative SDI was also substantially different between historical and contemporary forest conditions (Fig. 5). The variability of relative SDI for the historical dataset was much lower (coefficient of variation of 34%) than contemporary conditions (coefficient of variation of 59%).

Violin plots of regional historical (1924, left) and contemporary (2011–2018, right) distributions of relative stand density index (% of maximum SDI) including all trees ≥30.5 cm DBH. The gradient of gray shaded boxes indicates SDI benchmarks for free competition (in white; ≤25%), partial competition (25–35%), full site occupancy (35–60%), and imminent mortality (≥60%). The solid black line indicates 100% of the maximum SDI (902 trees ha−1; 365 trees ac−1)
Although we did not include California black oak in our estimates of relative SDI for contemporary conditions, we found that the highest percentage of FIA plots containing black oak (15%) were in plots not experiencing full conifer competition (<35% of maximum SDI). These plots of low conifer competition also had California black oak with the highest average DBH (15 cm), average density (361 trees ha−1) (Fig. 6), and average density (30 trees ha−1) of large (29 cm DBH; 75th percentile) California black oak. Conversely, the highest percentage of FIA plots without any black oak (46%) were in plots with high conifer competition (≥60% of maximum SDI; zone of imminent mortality), which also had California black oak with the lowest average DBH (5 cm), average density (11 trees ha−1), and average density (4 trees ha−1) of large California black oak relative to the other SDI benchmarks.

Average California black oak diameter (DBH [cm]; top panel) and density (trees ha−1; bottom panel) for FIA plots used to quantify contemporary forest conditions. FIA plots are grouped based on the relative SDI benchmark (% of maximum SDI) determined by conifer competition
Discussion
Native American land stewardship practices have increasingly come to the attention of natural resource management agencies such as the US National Park Service, California State Parks, US Bureau of Land Management, California Department of Forestry and Fire Protection (CAL FIRE), US Forest Service, and private land owners (Cuthrell et al. 2012). Indigenous practitioners champion holistic views on the benefits of burning rather than the more narrow objectives of fuel reduction and wildfire mitigation (Long et al. 2021). Tribes would like to burn more widely and frequently to promote resilience to wildfire and drought, conserve biocultural diversity, maintain spiritual values, and provide material goods (Long et al. 2021). Purcell and Stephens (2005) suggest Indigenous fires were likely of lower intensity due to their frequency, but intensity likely increased as shifts in fire management that included fire suppression and exclusion increased fuel loads. The second author of this paper would also like to point out our purposeful avoidance of the term “traditional ecological knowledge” in this paper, which reflects the views of the Mountain Maidu. The relationship between Native people and the natural environment is difficult to translate in English terms; a true understanding of this relationship cannot be fully defined, but must be experienced.
The forests sampled in the early 1920s throughout the broader study region had very low tree densities and were dominated by large ponderosa and sugar pine trees (Table 1). Such forest conditions would produce average canopy covers in the range of 20–30% (Collins et al. 2011) which would have allowed California black oak to thrive since it is shade intolerant. While the 1920s forest inventory did not include California black oak because it was not a commercial species, the open forest structure would have provided excellent conditions for California black oak establishment and growth. This is evident in the contemporary forests experiencing low conifer competition (<35% of maximum SDI), which had the highest densities of large California black oak (Fig. 6). Removal of Indigenous burning and lighting fires allowed heavy conifer recruitment (Knapp et al. 2013), the beginning of which can be seen in the photo that accompanied the 1920s inventory (Fig. 3). The impact of conifer recruitment in limiting California black oak establishment and growth is evident in the contemporary forests experiencing the highest levels of conifer competition (>60% of maximum SDI), which had 87% lower densities of large California black oak relative to forests with low conifer competition.
The fire history sample included in this work is illuminating. Although it is only a single sample, it has great temporal depth, spanning a long period that was devoid of the impacts of colonization (1518–1794). The last fire recorded (1794) was decades before the first non-Indigenous person came through the northern Sierra Nevada. The number of very short fire intervals recorded (11 intervals ≤5 years) is difficult to explain by lightning alone, pointing to the management of this area for Indigenous purposes. The number of California black oaks in this area in 2022 is substantial with an average density of 107 ha−1 even after > 100 years of fire suppression and exclusion. The high number of oak trees along with the fire scar record provides substantial evidence that this area was once an Indigenous area managed for California black oak.
It is worth noting that reconstructed fire frequency from a single fire-scarred cross-section may not fully capture fire occurrence at our focal site. The ability of a tree to record a fire depends on factors such as tree species and size, topographic position, and if the tree has been previously scarred (Collins and Stephens 2007; Farris et al. 2010). As fire frequency increases, the amount of time between fires decreases and this will impact fuel loads at the base of a tree that can burn and create a new scar. One study in Jeffrey pine-mixed conifer forests in Yosemite and Sequoia-Kings Canyon National Parks found that when a previous fire burned 1–10 years before the most recent event, less than 5% of the trees with an existing fire scar recorded the most recent fire (Stephens et al. 2010). This result highlights challenges of recording frequent burning because insufficient fuels may accumulate between fires to scar a tree when burned. A simulation study of fire scarring recommended including all fire dates when interested in Indigenous burning, rather than the common practice of filtering out individual small fires in most fire history studies (Roos et al. 2019). Even though the fire history sample in this work recorded intervals as short as 2 years with many fires with intervals < 10 years, it is probable that even more frequent Indigenous burning to manage California black oaks was done at this site. Sampling more fire history samples in and around our focal study site could provide more robust information but would not overcome the fact that fire scars are hard to create when fire return intervals are < 10 years. Our fire history sample provides a remarkable record that was likely produced by Indigenous burning on this site for > 250 years.
The fire seasonality record in the fire history sample warrants further discussion. The position of fire scars was 21% latewood and 42% dormant with a few late earlywood scars (6%). The latewood and late earlywood scars were likely from fires in September with fewer fires in August (Stephens et al. 2018), and this corresponds to cultural burning periods to facilitate resource collection and reduced insects that infest acorns (Lewis 1985; Anderson 2005). However, dormant season fires were the majority of our record and they could have occurred anytime the tree was not actively growing in circumference (from approximately October through April). Some of these dormant season fires could have occurred in winter between storms when sunny conditions could have dried out the surface litter; this is particularly possible when the overstory was pine dominated with its much more flammable and faster drying litter (York et al. 2022) which were the historic conditions in our greater study area (Table 1). The deeply lobed and highly flammable leaves of California black oak that would have been historically present in the litter layer would have also increased flammability during sunny, winter periods (Engber and Varner 2012). Winter fires could have been safely ignited to ensure fuel loads in our study area and surrounding areas were kept low to maintain these critical cultural resources when wildfires occurred (Bill Tripp, personnel communication 9/2022, Skinner 1995).
The conditions of this California black oak area before the 2021 Dixie Fire were substantially altered by fire exclusion and suppression along with logging. Current tree density of > 1000 trees ha−1 (trees > 2.5 cm DBH) with most of them being shade tolerant (Table 1) document a very hazardous forest condition. The more regional 1920s inventory that surrounded or focal study area (Fig. 3) documents a forest with tree densities of 53 ha−1 (> 30.5 cm DBH) that is similar to contemporary conditions (44 trees ha−1 > 30.5 cm DBH). While not recorded in this study, there are many large stumps in our study area where trees were harvested and likely transported to the now closed Meadow Valley sawmill that was only 4 km away. Unfortunately, the 1920s inventory did not measure trees < 30.5 cm DBH but other historical studies in the Sierra Nevada using similar protocols documented low tree densities below this DBH threshold (Stephens et al. 2015; Collins et al. 2015). We can confidently conclude that our study area has changed drastically without Mountain Maidu stewardship.
The competitive environment of the forests surrounding our focal study site has changed drastically over the last century. A majority (82%) of the forests sampled in 1924 had relative densities below the 35% benchmark (Table 1, Fig. 5) indicating that these forests were not experiencing full competition historically. This is a stark contrast to contemporary forest conditions which showed that 73% of plots were either experiencing full site occupancy (35–60% relative density) or were within the zone of imminent mortality (relative density ≥60%) which agrees with a larger-scale assessment of Sierra Nevada forests (North et al. 2022). This result highlights the vulnerability of current mixed conifer forests in the Sierra Nevada to widespread mortality even without experiencing wildfire (Restaino et al. 2019). The only remedy to this problem is to dramatically increase forest restoration efforts (Hessburg et al. 2021; Stephens et al. 2020, 2021b) including more cultural burning and stewardship by the Mountain Maidu.
In contrast, if we rely on wildfire as the de facto treatment, the likelihood of forest loss and the conversion to an alternate vegetation condition will only increase (Coop et al. 2020). That is not to say that all forests will be lost, as some areas that wildfires burn at low and moderate severity can benefit forests (Steel et al. 2021). The problem is that we cede any control over which specific areas and resources will be retained under a wildfire-dominated management regime.
Conclusions
The current western US landscape represents the historical legacy of one land paradigm superimposed on another, the colonial blanketing of the Indigenous, and nowhere is this more apparent than in the attitudes toward fire (Kimmerer and Lake 2001). Fire removal in the western US landscape is linked with the history of federal Indian policy that removed Tribal people and therefore Indigenous land management (Kimmerer and Lake 2001). The most powerful tool for landscape manipulation was fire for many tribes (Anderson 2005). Altered approaches to land management and the removal of frequent cultural burns placed California black oaks at risk from encroachment, forest densification, and destructive wildfires (Gorman et al. 2022).
The revitalization of cultural burning is a priority for many Indigenous people (Adlam et al. 2022). Ecosystems dependent on cultural fire regimes have deteriorated over the last century (Long et al. 2017) and active stewardship is needed to reverse this trend. Unlike western prescribed burning, cultural burning is rarely primarily focused on hazard and fuel reduction (Marks-Block and Tripp 2021). Indigenous people describe using fire to increase the quality and quantity of plant resources, to increase wildlife populations, to steward the land, and to maintain their cultural identity (Hankins 2015; Adlam et al. 2022).
Current conditions of the mixed conifer forests examined in this study highlight their vulnerability to widespread mortality. This studies local fire history and tree inventory information provides strong evidence for past cultural burning that likely created an area dominated by California black oak. The 2021 Dixie Fire killed all of the conifer trees in our study area but 61% of the oaks sprouted from the base after being top-killed. This provides an opportunity for California black oaks to regain dominance in our focal study site or in other areas that have similar post-wildfire outcomes. However, the removal of the numerous conifer snags created by the 2021 wildfire, coupled with competing vegetation control, will likely be necessary to reduce susceptibility to subsequent wildfire (Lydersen et al. 2019). It would take active stewardship to produce an area dominated by oaks again but it is possible and could be led by the Mountain Maidu with the assistance of the US Forest Service which owns the land.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- km:
-
Kilometer
- km2 :
-
Square kilometers
- US:
-
United States
- n.d.:
-
No date
- N:
-
North
- W:
-
West
- mm:
-
Millimeter
- NE:
-
Northeast
- T:
-
Township
- R:
-
Range
- E:
-
East
- ha:
-
Hectare
- HFQLG:
-
Quincy Library Group
- m:
-
Meter
- DBH:
-
Diameter at breast height
- cm:
-
Centimeter
- m2 :
-
Square meters
- GPS:
-
Global positioning system
- EE:
-
Early earlywood
- ME:
-
Middle earlywood
- LE:
-
Late earlywood
- LW:
-
Latewood
- D:
-
Dormant
- U:
-
Undetermined
- PLSS:
-
Public Land Survey System
- QQs:
-
Quarter-quarter sections
- FIA:
-
Forest Inventory and Analysis
- °C:
-
Degree Celsius
- n:
-
Number
- in:
-
Inch
- SDI:
-
Stand density index
- THP:
-
Trees per hectare
- D:
-
Average DBH (cm) of plot
- %:
-
Percent
- ac:
-
Acre
- CAL FIRE:
-
California Department of Forestry and Fire Protection
- UCB:
-
University of California, Berkeley
- ESRI:
-
Environmental Systems Research Institute
References
Adlam, C., D. Almendariz, R.W. Goode, D.J. Martinez, and D.J. and B.R. Middleton. 2022. Keepers of the flame: Supporting the revitalization of Indigenous cultural burning. Society and Natural Resources 35: 575–590.
Ahlstrand, G.M., 1980. Fire history of a mixed conifer forest in Guadalupe Mountains National Park. In Proceedings of the fire history workshop (Vol. 2044, pp. 4-7). USDA Forest Service, Rocky Mountain Research Station.
Anderson, M.K. 2005. Tending the wild: Native American knowledge and the management of California’s natural resources, 151–288. Berkeley: University of California Press.
Ansley, J.A.S., and J.J. Battles. 1998. Forest composition, structure, and change in an old-growth mixed conifer forest in the northern Sierra Nevada. Journal of the Torrey Botanical Society: 125: 297–308.
Bean, L.J., and H.W. Lawton. 1993. Some explanations for the rise of cultural complexity in Native California with comments on proto-agriculture and agriculture. In Before the wilderness: Environmental management by native Californians, ed. T.C. Blackburn and K. Anderson, vol. 40, 27–54. Ballena Press, Menlo Park. Anthropol. Pap.
Bernal, A.A., S.L. Stephens, B.M. Collins, and J.J. Battles. 2022. Biomass stocks in California’s fire-prone forests: Mismatch in ecology and policy. Environmental Research Letters 17: 044047.
Cocking, M.I., J.M. Varner, and R.L. Sherriff. 2012. California black oak responses to fire severity and native conifer encroachment in the Klamath Mountains. Forest Ecology and Management 270: 25–34.
Collins, B.M., A. Bernal, R.A. York, J.T. Stevens, A. Juska, and S.L. Stephens. 2021. Mixed-conifer forest reference conditions for privately owned timberland in the southern Cascade Range. Ecological Applications 31: 1–14.
Collins, B.M., R.G. Everett, and S.L. Stephens. 2011. Impacts of fire exclusion and recent managed fire on forest structure in old growth Sierra Nevada mixed-conifer forests. Ecosphere 2 (4): art 51.
Collins, B.M., D.L. Fry, J.M. Lydersen, R. Everett, and S.L. Stephens. 2017. Impacts of different land management histories on forest change. Ecological Applications 27: 2475–2486.
Collins, B.M., J.M. Lydersen, R.G. Everett, D.L. Fry, and S.L. Stephens. 2015. Novel characterization of landscape-level variability in historical vegetation structure. Ecological Applications 25: 1167–1174.
Collins, B.M., and S.L. Stephens. 2007. Fire scarring patterns in Sierra Nevada wilderness areas burned by multiple wildland fire use fires. Fire Ecology 3: 53–67.
Comer, P., D. Faber-Langendoen, R. Evans, S. Gawler, C. Josse, G. Kittel, S. Menard, M. Pyne, M. Reid, K. Schulz, K. Snow, and J. Teague. 2003. Ecological systems of the United States: A working classification of U.S. terrestrial systems. Nature Serve: Ecological Systems of the United States: A Working Classification of U.S. Terrestrial Systems, 75.
Coop, J.D., S.A. Parks, C.S. Stevens-Rumann, S.D. Crausbay, P.E. Higuera, M.D. Hurteau, A. Tepley, E. Whitman, T. Assal, B.M. Collins, K.T. Davis, S. Dobrowski, D.A. Falk, P.J. Fornwalt, P.Z. Fulé, B.J. Harvey, V.R. Kane, C.E. Littlefield, E.Q. Margolis, M. North, M.-A. Parisien, S. Prichard, and K.C. Rodman. 2020. Wildfire-driven forest conversion in Western North American landscapes. Bioscience 70: 659–673.
Cuthrell, R.Q., C. Striplen, M.G. Hylkema, K.G. Lightfoot, T.L. Jones, and J.E. Perry. 2012. In A land of fire: Anthropogenic burning on the central coast of California. Contemporary issues in California archaeology, ed. T.L. Jones and J.E. Perry, 153–172. Walnut Creek: Left Coast Press.
Dieterich, J.H., 1980. The composite fire interval—a tool for more accurate interpretation of fire history. In Proceedings of the fire history workshop (pp. 8-14). Fort Collins, Colorado, USA: Rocky Mountain Forest and Range Experiment Station.
Dieterich, J.H., and T.W. Swetnam. 1984. Dendrochronology of a fire-scarred ponderosa pine. Forest Science 30: 238–247.
Echo-Hawk, R.C. 2000. Ancient history in the New World: Integrating oral traditions and the archaeological record in deep time. American Antiquity 65 (2): 267–290.
Engber, E.A., and J.M. Varner. 2012. Patterns of flammability of the California oaks: The role of leaf traits. Canadian Journal of Forest Research 42: 1965–1975.
Evett, R., E. Franco-Vizcaino, and S.L. Stephens. 2007. Comparing modern and past fire regimes to assess changes in prehistoric lightning and anthropogenic ignitions in a Jeffrey pine-mixed conifer forest in the Sierra San Pedro Martir, Mexico. Canadian Journal of Forest Research 37: 318–330.
Farris, C.A., C.H. Baisan, D.A. Falk, S.R. Yool, and T.W. Swetnam. 2010. Spatial and temporal corroboration of a fire-scar-based fire history in a frequently burned ponderosa pine forest. Ecological Applications 20: 1598–1614.
Ferguson, T.J., S.B. Koyiyumptewa, and M.P. Hopkins. 2015. Co-creation of knowledge by the Hopi Tribe and archaeologists. Advances in Archaeological Practice 3 (3): 249–262.
Finney, M.A., and R.E. Martin. 1992. Short fire intervals recorded by redwoods at Annadel State Park, California. Madrono 39: 251–262.
Flint, L.E., A.L. Flint, J.H. Thorne, and R. Boynton. 2013. Fine-scale hydrologic modeling for regional landscape applications: The California Basin Characterization Model development and performance. Ecological Processes 2: 1–21.
Flint, L.E., A.L. Flint, J.H. Thorne, and R. Boynton. 2014. California BCM (Basin Characterization Model) downscaled climate and hydrology – 30-year summaries. California Climate Commons. http://climate.calcommons.org/dataset/2014-CA-BCM.
Garibaldi, A., and N. Turner. 2004. Cultural keystone species: Implications for ecological conservation and restoration. Ecology and Society 9 (3): 1.
Haag, A.M. 2007. The Indian boarding school era and its continuing impact on tribal families and the provision of government services. Tulsa Law Review 43: 149–168.
Hankins, D.L., 2015. Restoring indigenous prescribed fires to California oak woodlands. Gen. Tech. Rep. PSW-GTR-251. Berkeley: US Department of Agriculture, Forest Service, Pacific Southwest Research Station: 123-129, 251, 123-129.
Hessburg, P.F., S.J. Prichard, R.K. Hagmann, N.A. Povak, F.K. Lake, and F. K. 2021. Wildfire and climate change adaptation of western North American forests: A case for intentional management. Ecological Application 31 (8): e02432.
Kelsey, H. 1973. The California Indian treaty myth. Southern California Quarterly 55 (3): 225–238.
Kimmerer, R.W., and F.K. Lake. 2001. The role of indigenous burning in land management. Journal of Forestry 99: 36–41.
Knapp, E.E., C.N. Skinner, M.P. North, and B.L. Estes. 2013. Long-term overstory and understory change following logging and fire exclusion in a Sierra Nevada mixed-conifer forest. Forest Ecology and Management 310: 903–914.
LANDFIRE. 2022a. LANDFIRE, Elevation, LF 2.0.0, U.S. Department of the Interior, Geological Survey. http://LANDFIRE.cr.usgs.gov/viewer
LANDFIRE. 2022b. LANDFIRE, Biophysical settings, LF 2.0.0, U.S. Department of the Interior, Geological Survey. http://LANDFIRE.cr.usgs.gov/viewer
LANDFIRE. 2022c. LANDFIRE, Slope, LF 2.0.0, U.S. Department of the Interior, Geological Survey. http://LANDFIRE.cr.usgs.gov/viewer
Leiberg, J. B. 1902. Forest conditions in the northern Sierra Nevada. Professional Paper No. 8, Department of Interior, U.S. Geological Survey.
Lewis, H.T., 1985. Why Indians burned: Specific versus general reasons. In Lotan, JE; Kilgore, BM; Fischer, WC; Mutch, RW, tech. cords. Proceedings of the symposium and workshop on wilderness fire. Gen. Tech. Rep. Report INT-182. Ogden: US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station (pp. 75-80).
Long, J.N. 1985. A practical approach to density management. The Forestry Chronicle 61: 23–27.
Long, J.N., and J.D. Shaw. 2005. A density management diagram for even-aged ponderosa pine stands. Western Journal of Applied Forestry 20: 1–11.
Long, J.W., M.K. Anderson, L. Quinn-Davidson, R.W. Goode, F.K. Lake, and C.N. Skinner. 2016. Restoring California black oak ecosystems to promote tribal values and wildlife. General Technical Report PSW GTR-252. Albany: U.S. Forest Service, Pacific Southwest Research Station.
Long, J.W., R.W. Goode, R.J. Gutteriez, J.J. Lackey, and M.K. Anderson. 2017. Managing California Black Oak for tribal ecocultural restoration. Journal of Forestry 115 (5): 426–434.
Long, J.W., F.K. Lake, and R.W. Goode. 2021. The importance of Indigenous cultural burning in forested regions of the Pacific West, USA. Forest Ecology and Management 500: 119597.
Lydersen, J.M., B.M. Collins, M. Coppoletta, M.R. Jaffe, H. Northrop, and S.L. Stephens. 2019. Fuel dynamics and reburn severity following high severity fire in a Sierra Nevada mixed-conifer forest. Fire Ecology 15: 43.
Marks-Block, T., and W. Tripp. 2021. Facilitating prescribed fire in Northern California through Indigenous Governance and interagency partnerships. Fire 4 (3): 37.
McDonald, P.M., R.M. Burns, and B.H. Honkala. 1990. Quercus kelloggii Newb., California black oak. Silvics of North America. Volume 2, Hardwoods. Agriculture Handbook 654: 661–671.
Meigs, P. 1935. The Dominican missions frontier of Lower California. University of California Publication in Geography 7: 1–232.
Moghaddas, J.J., B.M. Collins, K. Menning, E.E.Y. Moghaddas, and S.L. Stephens. 2010. Fuel treatment effects on modeled landscape level fire behavior in the northern Sierra Nevada. Canadian Journal of Forest Research 40: 1751–1765.
Nemens, D.G., J.M. Varner, K.R. Kidd, and B. Wing. 2018. Do repeated wildfires promote restoration of oak woodlands in mixed-conifer landscapes? Forest Ecology and Management 427: 143–151.
North, M.P., R.E. Tompkins, A.A. Bernal, B.M. Collins, S.L. Stephens, and R.A. York. 2022. Operational resilience in western US frequent-fire forests. Forest Ecology and Management 507: 120004.
North, M.P., R.A. York, B.M. Collins, M.D. Hurteau, G.M. Jones, , E.E. Knapp, L.N. Kobziar, H. McCann, M.D. Meyer, S.L. Stephens, R.E. Tompkins, and C.L. Tubbesing, 2021. Pyrosilviculture needed for landscape resilience of dry Western U.S. forests. Journal of Forestry 119(5): 520-544..
O’Gorman, C., L. Bentley, C. McKay, M. Purser, and K. Everly. 2022. Examining abiotic and biotic factors influencing specimen black oaks (Quercus kelloggii) in northern California to reimplement traditional ecological knowledge and promote ecosystem resilience post-wildfire. Ecology and Society 27 (2).
Parks, S. A., Holsinger L. M, Miller C. , and Nelson C. R.. 2015. Wildland fire as a self-regulating mechanism: the role of previous burns and weather in limiting fire progression. Ecological Applications 25:1478-1492
Plumas County Museum. n.d. Maidu History. https://www.plumasmuseum.org/maiduhist.html#:~:text=The%20Maidu%20also%20endured%20numerous,was%20estimated%20at%20about%204%2C000. Accessed 7 Jan 2022.
Purcell, K.L., and S.L. Stephens. 2005. Changing fire regimes and the avifauna of California oak woodlands. Studies in Avian Biology 30: 33–45.
Restaino, C., D.J.N. Young, B. Estes, S. Gross, A. Wuenschel, M. Meyer, and H. Safford. 2019. Forest structure and climate mediate drought-induced tree mortality in forests of the Sierra Nevada, USA. Ecological Applications 29: e01902.
Roos, C.I., G.J. Williamson, and D.M.J.S. Bowman. 2019. Is anthropogenic pyrodiversity invisible in paleofire records? Fire 2: 42.
Safford, H.D., and J.T. Stevens. 2017. Natural range of variation for yellow pine and mixed-conifer forests in the Sierra Nevada, southern Cascades, and Modoc and Inyo National Forests, California, USA. Gen. Tech. Rep. PSW-GTR-256. Vol. 229, 256. Albany: US Department of Agriculture, Forest Service, Pacific Southwest Research Station.
Skinner, C.N. 1995. Using prescribed fire to improve wildlife habitat near Shasta Lake. Redding: Unpublished report U.S. Department of Agriculture Forest Service, Pacific Southwest Research Station.
Skinner, C.N., C.S. Abbott, D.L. Fry, S.L. Stephens, A.H. Taylor, and V. Trouet. 2009. Human and climatic influences on fire occurrence in California’s North Coast Range, USA. Fire Ecology 5 (3): 76–99.
Steel, Z.L., D.E. Foster, M. Coppoletta, J.M. Lydersen, S.L. Stephens, A. Paudel, S.H. Markwith, K. Merriam, and B.M. Collins. 2021. Ecological resilience and vegetation transition in the face of two successive large wildfires. Journal of Ecology 109: 3340–3355.
Steen-Adams, M.M., S. Charnley, R.J. McLain, M.D.O. Adams, and K.L. Wendel. 2019. Traditional knowledge of fire use by the Confederated Tribes of Warm Springs in the eastside Cascades of Oregon. Forest Ecology and Management 450: 117405.
Stephens, S.L., M.A. Battaglia, D.J. Churchill, B.M. Collins, M. Coppoletta, C.M. Hoffman, J.M. Lydersen, M.P. North, R.A. Parsons, S.M. Ritter, and J.T. Stevens. 2021b. Forest restoration and fuels reduction: Convergent or divergent? Bioscience 71 (1): 85–101.
Stephens, S.L., and D.L. Fry. 2005. Fire history in coast redwood stands in the Northeastern Santa Cruz Mountains, California. Fire Ecology 1 (1): 2–19.
Stephens, S.L., D.L. Fry, B.M. Collins, C.N. Skinner, E. Franco-Vizcaíno, and T.J. Freed. 2010. Fire-scar formation in Jeffrey pine–mixed conifer forests in the Sierra San Pedro Mártir, Mexico. Canadian Journal of Forest Research 40 (8): 497–1505.
Stephens, S.L., J.M. Lydersen, B.M. Collins, D.L. Fry, M.D. Meyer, and M.D. 2015. Historical and current landscape-scale ponderosa pine and mixed-conifer forest structure in the Southern Sierra Nevada. Ecosphere 6 (5): art 79.
Stephens, S.L., L. Maier, L. Gonen, J.D. York, B.M. Collins, and D.L. Fry. 2018. Variation in fire scar phenology from mixed conifer trees in the Sierra Nevada. Canadian Journal of Forest Research 48 (1): 101–104.
Stephens, S.L., S. Thompson, G. Boisramé, B.M. Collins, L.C. Ponisio, E. Rakhmatulina, Z.L. Steel, J.T. Stevens, J.W. van Wagtendonk, and K. Wilkin. 2021a. Fire, water, and biodiversity in the Sierra Nevada: A possible triple win. Environmental Research Communications 3 (8): 081004.
Stephens, S.L., A.L. Westerling, M.D. Hurteau, M.Z. Peery, C.A. Schultz, and S. Thompson. 2020. Fire and climate change: Conserving seasonally dry forests is still possible. Frontiers in Ecology and the Environment 18: 354–360.
Stokes, M.A., and T.L. Smiley. 1977. An introduction to tree-ring dating. Chicago: University of Chicago Press.
Van Wagtendonk, J.W., and D.R. Cayan. 2008. Temporal and spatial distribution of lightning strikes in California in relation to large-scale weather patterns. Fire Ecology 4: 34–56.
Wensel, L.C., and C.M. Olson. 1995. Tree volume equations for major California conifers. Hilgardia 62: 1–94.
Whittaker, R.H. 1960. Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs 30: 279–338.
Woudenberg, S.W., B.L. Conkling, B.M. O’Connell, E.B. LaPoint, J.A. Turner, and K.L. Waddell. 2010. The Forest Inventory and Analysis database: Database description and users manual version 4.0 for Phase 2. General Technical Report RMRS-GTR-245. Ft. Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
York, R.A., J.I. Levine, D.E. Foster, S.L. Stephens, and B.M. Collins. 2022. Silviculture can facilitate repeat prescribed burn programs with long-term strategies. California Agriculture 75: 104–111.
Zhang, J., K.A. Finley, N.G. Johnson, and M.W. Ritchie. 2019. Lowering stand density enhances resiliency of ponderosa pine forests to disturbances and climate change. Forest Science 65: 496–507.
Zhang, J., W.W. Oliver, and R.F. Powers. 2013. Reevaluating the self-thinning boundary line for ponderosa pine (Pinus ponderosa) forests. Canadian Journal of Forest Research 43: 963–971.
Acknowledgements
We thank Celeste Abbott for crossdating our fire history sample, Bob Carlson for preparing it, and Eric Knapp for supporting this study. Natalie MacMillan and Brandon Pundamiera helped Scott Stephens collect the scarred stump and Mary Gross assisted with field measurements. We thank Jonathan Long and Peter Nelson for supplying background information and UCB summer camp students for collecting data from the Meadow Valley plots over the years.
Funding
None.
Author information
Authors and Affiliations
Contributions
SS: conceptualization, methodology, investigation, writing—original draft, writing—review and editing, and supervision. LH: methodology, writing—original draft, and writing—review and editing. CS: conceptualization, investigation, methodology, formal analysis, and writing—review and editing. AB: data curation, formal analysis, methodology, writing—original draft, and writing—review and editing. BC: methodology, formal analysis, investigation, methodology, writing—original draft, and writing—review and editing. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: “__Formatting error in abstract has been corrected_”.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Stephens, S.L., Hall, L., Stephens, C.W. et al. Degradation and restoration of Indigenous California black oak (Quercus kelloggii) stands in the northern Sierra Nevada. fire ecol 19, 12 (2023). https://doi.org/10.1186/s42408-023-00172-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-023-00172-9