
Abstract
Background
Climate change is eroding forest resilience to disturbance directly through warming climate and indirectly through increasing disturbance activity. Forests characterized by stand-replacing fire regimes and dominated by serotinous species are at risk when the inter-fire period is insufficient for canopy seed bank development and climate conditions for recruitment in the post-fire growing season are unsuitable. Although both factors are critical to serotinous forest persistence, their relative importance for post-fire regeneration in serotinous forests remains poorly understood. To assess the relative effects of each factor, we established plots in severely burned knobcone pine (Pinus attenuata Lemmon) forests in Oregon and California, USA, representing a range of past fire intervals (6 to 31+ years). Specifically, we evaluated effects of fire interval and pre-fire canopy seed bank (proxies for seed supply) and post-fire climate on three metrics of post-fire tree regeneration (seedling density, probability of self-replacement, percent population recovery).
Results
Seed supply consistently had the strongest effect on post-fire regeneration. Between 6- and 31-year fire intervals, post-fire seedling density increased from 1000 to 100,000 seedlings ha−1, while probability of self-replacement increased from ~ 0 to ~ 100% and percent population recovery increased from 20 to 2000% of the pre-fire population, respectively. Similarly, increasing the canopy seed bank by two orders of magnitude increased seedling density and percent population recovery by two orders and one order of magnitude, respectively, and increased the probability of self-replacement by > 50%. Greater post-fire climatic moisture deficit exacerbated the effect of seed supply; an additional 4–6 years between fires was required under high moisture stress conditions to reach similar regeneration levels as under low moisture stress conditions.
Conclusion
The overriding effect of seed supply—strongly driven by pre-fire stand age—on post-fire regeneration suggests that altered fire frequency (an indirect effect of climate change) will have a profound impact on serotinous forests. Although direct effects of hot and dry climate are lower in magnitude, they can alter forest recovery where seed supply nears a threshold. These findings reveal how fire interval and climate combine to determine changes in forest cover in the future, informing management and vulnerability mapping.
Resumen
Antecedentes
El cambio climático está erosionando la resiliencia de los bosques a los disturbios de manera directa mediante el calentamiento global, e indirectamente a través del incremento de los disturbios. Los bosques caracterizados por regímenes de fuego que implican el reemplazo de rodales (stand-replacing fires) y dominados por especies serótinas, están en riesgo cuando el período entre fuegos es insuficiente como para el desarrollo de un banco de semillas aéreo (en el dosel) y las condiciones climáticas para el reclutamiento en la estación de crecimiento post-fuego no son las adecuadas. Aunque ambos factores son cruciales para la persistencia de bosques serótinos, su importancia relativa para la regeneración de especies forestales serótinas está muy pobremente comprendida. Para determinar los efectos relativos de cada factor, establecimos parcelas en bosques de pino nudoso (Pinus attenuatta Lemmon), severamente quemados en Oregón y California, EEUU, representando rangos de intervalos de fuego de 6 a 31 o más años. Específicamente, evaluamos los efectos de los intervalos de fuego y el banco de semillas serótinas en el dosel en el pre-fuego (como proxys en la provisión de semillas) y el clima post-fuego sobre tres variables en la regeneración post-fuego (densidad de plántulas, probabilidad de auto reemplazo de los rodales, y porcentaje de recuperación de la población).
Resultados
La provisión de semillas tuvo consistentemente el efecto más pronunciado en la regeneración post-fuego. Entre los 6 y 31 años de intervalo de fuego, la densidad de plántulas en los períodos post-fuego se incrementó de 1.000 a 100.000 por ha1, mientras que la probabilidad de auto reemplazo se incrementó de ~ 0 a ~ 100% y el porcentaje de recuperación de la población se incrementó del 20% al 2000% en relación a la población en el pre-fuego, respectivamente. De manera similar, el aumento del banco de semillas en los doseles incrementó la densidad de plántulas y el porcentaje de la recuperación por dos y un orden de magnitud, respectivamente, e incrementó la probabilidad del auto reemplazo por > 50%. El mayor déficit de humedad climática en el post-fuego exacerbó el efecto en la provisión de semillas; se necesitaron entre 4 a 6 años más entre fuegos bajo condiciones de alto estrés hídrico para alcanzar niveles de regeneración similares a cuando había condiciones bajas de estrés hídrico.
Conclusiones
El efecto primordial de la provisión de semillas (fuertemente condicionada por la edad de los rodales pre-fuego) sobre la regeneración en el post-fuego, sugiere que la alteración en la frecuencia de los fuegos (efecto indirecto del cambio climático) tendrá un profundo impacto en bosques serótinos. Aunque los efectos directos del clima más seco y cálido son de baja magnitud, pueden alterar la recuperación de los bosques donde la provisión de semilla es casi una limitante. Estos resultados revelan cómo el intervalo entre fuegos y el cambio climático se combinan para determinar cambios en la cobertura de bosques en el futuro, lo cual alerta a las acciones de manejo y la vulnerabilidad de los mapeos.
Similar content being viewed by others

Background
Evidence from ecosystems worldwide indicates that climate change is eroding forest resilience to disturbance (Trumbore et al. 2015)—defined as an ecosystem’s capacity to experience disturbance without transitioning to an alternative state (Gunderson 2000). Direct effects of warming climate are leading to increases in tree mortality and forest die-offs (Williams et al. 2013; Allen et al. 2015; Matusick et al. 2018). Indirect effects of climate change via altered disturbance regimes are also influencing forest resilience through decreased capacity to recover post-disturbance (Seidl et al. 2017). Increasing fire activity attributed to climate change is already apparent (Abatzoglou and Williams 2016), with evidence of an extended fire season (Jolly et al. 2015), increased total area burned (Boer et al. 2020; Abatzoglou et al. 2021; Collins et al. 2022), and proportion area burned at high severity (Parks and Abatzoglou 2020; Collins et al. 2021) in forests around the globe. Understanding how direct and indirect effects of climate change combine to affect forest resilience is critical to understanding when and where forests may be vulnerable to non-forest conversion (Coop et al. 2020; Falk et al. 2022).
Post-fire tree regeneration is a key indicator of resilience in fire-prone forests as it sets the template for the post-disturbance forest trajectory. Declines in post-fire regeneration have been observed in many forest ecosystems since the early 2000s (Stevens-Rumann et al. 2018) and have been mechanistically linked to both departures from historical fire regimes and changes in post-fire climate conditions. For species that depend on surviving trees to provide a seed source for post-fire regeneration, increasing fire severity and fire size can result in decreases in post-fire regeneration due to high mortality of seed trees within a fire perimeter and increased distance to an unburned edge (Gill et al. 2021). Such changes in the fire regime have led to decreases in post-fire regeneration in montane and subalpine forests in the Western US (Crotteau et al. 2013; Collins and Roller 2013; Kemp et al. 2016; Chambers et al. 2016; Busby et al. 2020; Boag et al. 2020). High moisture stress during the growing season also leads to poor post-fire regeneration outcomes (Harvey et al. 2016; Hansen and Turner 2019; Davis et al. 2019). Where uncharacteristically high fire severity is followed by extreme drought conditions, complete conifer regeneration failure and non-forest conversion has been observed and is likely to continue with warming climate (Guiterman et al. 2018).
Forests characterized by stand-replacing fire regimes and dominated by serotinous species (trees that release seeds from cones when heated by fire) are well-adapted to some projected changes in fire regimes, though potentially maladapted to others (Enright et al. 2015). Serotinous species are likely to respond favorably to increasing area burned at high severity, which is occurring in many systems (Parks and Abatzoglou 2020; Collins et al. 2022). Such changes to the fire regime favor serotinous species, as they do not depend on a live seed source for post-fire recruitment (Lamont et al. 1991), and can establish and expand where non-serotinous obligate seeding species require dispersal from live trees (Hansen et al. 2018; Gill et al. 2021). However, several dimensions of climate warming and altered fire regimes may erode resilience of serotinous species by altering key mechanisms underpinning successful post-fire regeneration.
One key mechanism of resilience to fire in forests dominated by serotinous species is a sufficient seed bank stored in the canopy of mature trees, producing the ability to recruit en masse from seed released within the first post-fire year. Serotinous coniferous forests often regenerate at densities exceeding 100,000 seedlings ha−1 following stand-replacing fire in North America (Harvey and Holzman 2014; Turner et al. 2016), but realization of this regeneration potential requires accumulation of a sufficient canopy seed bank at the time of fire. Climate change can alter fire regimes, indirectly eroding this resilience mechanism if fire occurs in young stands during the early years of reproductive maturity (Agne et al. 2022). If a short-interval reburn kills serotinous trees prior to the accumulation of a sufficient canopy seed bank, compound effects (sensu Paine et al. 1998) from two successive fires may convert forests to non-forest, or substantially shift tree species composition (Brown and Johnstone 2012).
Another factor that determines resilience to fire for serotinous species is suitable climate for tree recruitment in the first post-fire year. Post-fire regeneration failure has been defined in a variety of ways; as regeneration density less than a set stocking density (Hansen et al. 2018; Littlefield 2019; Rodman et al. 2020), regeneration density less than pre-fire density (Stevens-Rumann et al. 2018) and even as complete recruitment failure (Coop et al. 2020; Baltzer et al. 2021). Regeneration failure is more likely with greater annual moisture deficit (i.e., direct effect of climate change; Stevens-Rumann et al. 2018), indicating that fire followed by drought can produce compound effects. These trends appear to be most pronounced in dry forests (Harvey et al. 2016; Davis et al. 2019; Rodman et al. 2019) and projections for increasingly warm and dry conditions suggest further declines in post-fire seedling recruitment through the twenty-first century (Rodman et al. 2020). Topographic complexity, variation in soils, and competition with shrub species also affect moisture availability and can lead to widely variable recruitment outcomes among microsites (Harvey et al. 2016; Busby et al. 2020; Hoecker et al. 2020). The risk of recruitment failure is exacerbated in serotinous conifers, as seed dispersal primarily occurs in a pulse following fire (Johnson and Fryer 1989; Keeley and Zedler 1998) and seedling establishment occurs in the first year post-fire with relatively little recruitment in subsequent years (Turner et al. 1999; Donato et al. 2009; Harvey and Holzman 2014). When harsh climate conditions (e.g., drought, heatwaves) co-occur with this brief recruitment window, seed available post-fire may fail to produce seedlings, leading to conversion to non-forest (Enright et al. 2014).
Although climate warming is likely to affect post-fire climate conditions and on-site seed availability, the relative importance of these factors in successful post-fire serotinous conifer recruitment has not been investigated within a single framework. Serotinous forests offer a powerful opportunity to ask such questions because seeds are contained on site and issues of distance to seed source are much less important. To understand the effect of short-interval severe reburns on serotinous forests, we examined forests dominated by serotinous knobcone pine (Pinus attenuata Lemmon) to ask: How are three measures of forest resilience to severe fire (seedling density, probability of self-replacement, and percent population recovery) affected by seed supply at the time of fire and post-fire recruitment conditions? We expected that increasing seed supply as functions of stand age and density (estimated using fire interval and pre-fire canopy seed bank size) would have strong positive effects on post-fire regeneration due to increased time to accumulate and protect seeds within cones (Fig. 1). We also expected that for a given seed supply, post-fire regeneration would decrease with warm and dry (relative to mean) post-fire climate. Further, at the local-scale, we expected decreased post-fire regeneration on xeric sites (warm and dry topoclimate), where soils are relatively shallow and poor, and with high shrub competition. Understanding how direct (post-fire climate) and indirect (on-site seed availability) effects of climate change influence post-fire recruitment individually and how they interact is critical for predicting how forests will respond to fire in the future.

Post-fire forest structure in knobcone pine (Pinus attenuata Lemmon) forests in Oregon and California, USA, following fire intervals of A 6 years, B 10 years, C 17 years, D 22 years, E 31 years, and F long unburned (> 34 year fire interval). Photos were taken 2 years post-fire during the summer of 2020. Photo credit for all panels: M. Agne
Methods
Study area
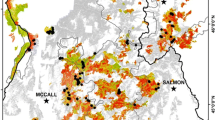
This study was conducted in knobcone pine stands in Oregon and California, USA, within the Klamath Mountains and Northern Coast Range which experienced stand-replacing fire in 2018 (Fig. 2). We selected three large fires that burned through knobcone pine forests (U.S. Geological Survey 1999a) during the summer of 2018 in which to establish field plots: the Carr Fire, Klondike Fire, and Mendocino Complex. The study area is characterized by a Mediterranean climate including a strong seasonality in precipitation, with > 90% falling between October and May (PRISM Climate Group 2019). Climate varies strongly among study fires, with the Klondike Fire in the northern Klamath Mountains representing the coolest and wettest area (average annual precipitation = 1897 mm and average January and July temperatures are 5.1 and 19.8 °C, respectively). The Carr Fire in the southern Klamath Mountains represents the warmest site (average January and July temperatures are 6.8 and 25.9 °C, respectively) with a moderate amount of precipitation (average annual precipitation = 1572 mm). The Mendocino Complex in the Northern Coast Range has a moderate temperature (average January and July temperatures are 5.8 and 23.2 °C, respectively) and represents the driest site (average annual precipitation = 1126 mm).

Study area map of sample fires in Oregon and California, USA. Top left: Continental United States with box around study region. Bottom left: Knobcone pine distribution in study region with the three 2018 sample fire perimeters. Scale bar refers to this panel. Right: 2018 sample fire perimeters with area reburned shaded by twice and thrice burned between 1984 and 2018. Twice and thrice burned plots were not treated separately in analyses due to similar pre-fire structure and post-fire response across plots with the same most recent fire interval (see Additional file 1: Fig. 1.1)
The study area encompasses low elevation to lower montane zones (320–1243 m above sea level [asl]). Stands are often characterized by steep slopes and soils are typically shallow and rocky, with serpentine substrate common for knobcone pine (Vogl et al. 1977). Serpentine soils are acidic with low nutrient concentrations, leading to low productivity and restriction of many plant species from such sites (Vogl 1973). Knobcone pine-dominated plant communities often occur as mono-dominant, dense forest stands for several decades following establishment but can also co-occur with Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) or exist at a low density interspersed with chaparral (Fry et al. 2012; Reilly et al. 2019). In this study, we focused on areas that were forested and dominated by knobcone pine (often monotypic stands) pre-fire and excluded areas where knobcone pine was a subordinate species to other conifers as it can be throughout its range. Understory plant communities vary across the study area with a high diversity of shrub and herbaceous species represented. Common co-occurring shrub species include Adenostoma fasciculatum Hook. & Arn., Arctostaphylos spp., Ceanothus spp., Heteromeles arbutifolia (Lindl.) M. Roem., and Toxicodendron diversilobum (Torr. & A. Gray) Greene. Arbutus menziesii Pursh, Chrysolepis chrysophylla (Douglas ex Hook.) Hjelmqvist, Notholithocarpus densiflorus (Hook. & Arn.) P.S. Manos, C.H. Cannon, & S.H. Oh, and Quercus spp. are also common, existing both in shrub and tree form. Herbaceous vegetation commonly includes Acmispon glaber (Vogel) Brouillet, Lupinus spp., and Mimulus spp.
Knobcone pine forests are typically characterized by a stand-replacing fire regime, which creates conditions suitable for regeneration, resulting in even-aged stands (Vogl 1973). However, non-stand-replacing fire can initiate secondary cohorts and uneven-aged stand structure in some cases (Fry et al. 2012), and cone opening in the absence of fire may produce inter-fire recruits. Knobcone pine is a relatively short-lived species, producing cones as early as 4–6 years of age (Keeley et al. 1999; Agne et al. 2022), with substantial stand-level decline by 60–75 years of age (Vogl 1973; Howard 1992), and individual trees rarely surviving to 100 years (Fry et al. 2012). Historical fire frequency for knobcone pine is poorly understood due to its patchy distribution and co-occurrence with plant communities that experienced widely differing fire regimes (Howard 1992). The historical mean fire interval for serotinous conifers (including knobcone pine) and chaparral as a vegetation community has been estimated at 30–90 years (van de Water and Safford 2011). This range likely overestimates the historical fire interval for knobcone pine specifically, given that stands begin to decline by 60 years (Howard 1992). Conversely, dry mixed-conifer forests dominated by ponderosa pine (Pinus ponderosa Lawson & C. Lawson) or Douglas-fir that overlap the range of knobcone pine were characterized by mean fire intervals 5–19 years (Taylor and Skinner 1998; Fry and Stephens 2006). Such shorter interval fires would tend to be non-stand replacing, not favor serotinous species, and likely underestimate the historical fire return interval for nearby knobcone pine populations. Therefore, we use the range of 30–60 years between stand-replacing fires as our estimate of historical fire frequency, although we recognize that fire activity, especially non-stand-replacing fire, may have been more frequent throughout the study area.
Site selection
To select sites, we used Monitoring Trends in Burn Severity data (MTBS; Eidenshink et al. 2007), to identify fire history after 1984 and prior to each 2018 fire to determine the fire intervals present within the range of knobcone pine for each of the sample fires. Collectively, seven known fire intervals, ranging from 6 to 31 years, overlapped the distribution of knobcone pine within the three 2018 fires. Additionally, knobcone pine forest with unknown prior fire history was present in each of the three 2018 sample fires (i.e., fire interval > 34 years). In each 2018 fire, we stratified our field sampling to establish at least 5 plots within each known fire interval, and at least 5 plots in areas unburned since 1984 (and no recorded fire history in the California Department of Forestry and Fire Protection’s Fire and Resource Assessment Program (FRAP) database [https://frap.fire.ca.gov/mapping/gis-data/]), hereafter, long-interval plots. Criteria for inclusion in our sample included pre-fire dominance or codominance of knobcone pine forest, burned at high severity (100% overstory tree mortality) in 2018, and burned at high severity in prior fire (as determined by stand structure—the presence of a single cohort of knobcone pine prior to 2018 fire). Plots with no available prior fire history were not required to meet the criterion of having burned at high severity in the prior fire as we lacked information on prior burn severity. Potential sample locations were selected in ArcGIS (Environmental Systems Research Institute [ESRI] 2018) using aerial imagery, species distribution maps, and burn severity maps prior to the field season. When pre-identified plot locations did not meet the criteria for sampling, we moved the plot location to the nearest area that met the criteria and plot center locations were randomized within the stand to avoid bias in placement. Plot replicates within fire intervals were established to cover the range of topography and pre-fire stand structure present. Plots were separated by at least 100 m and were located at least 50 m from roads and trails.
Field methods
During the summer of 2020, we established 71 field plots in which we collected pre-fire stand structure, tree regeneration, and understory vegetation data, using a plot design modified from established protocols (Harvey et al. 2016). To reconstruct pre-fire stand structure, we measured diameter at breast height (DBH, measured 1.37 m from the ground), assigned species to each standing and fallen tree and counted post-fire open and closed cones on each knobcone pine of any size killed by the most recent fire. We assumed that open cones contributed to seed release resulting in regeneration at the time of sampling, while closed cones did not. We also noted the presence of cones consumed by fire and assumed these cones were open prior to fire and did not contribute to post-fire seed release. Data were collected within a variable radius plot aimed at capturing approximately 50 pre-fire trees and with a maximum plot radius of 18 m. Tree regeneration was measured within four 2 × 16 m belt transects oriented to the clockwise side of the cardinal directions from plot center. The sampling area was decreased to four 1 × 16 m belt transects when seedling counts would have exceeded an estimated 400 using the default plot size and was increased to encompass the entire pre-fire stand plot when seedling counts were under 40 using the default plot size. Within the regeneration sampling area, we counted all tree seedlings by species, assigning them as live or dead. For knobcone pine only, we assigned seedlings as second year (established within the growing season following fire) if bud scars were present and first year (established within the second growing season following fire) if bud scars were absent and cotyledons were present. Understory vegetation cover was measured within eight 1 × 1 m subplots placed along each belt transect at the edge of the plot and half the distance between plot center and the edge of the plot. Within each subplot, we estimated (to the nearest 5%) understory vegetation cover by life form (shrub, graminoid, forb, tree). We collected a GPS point, elevation, slope, and aspect from each plot center.
Climate, topography, and soil variables
We obtained climate data for our plots from ClimateNA, a software package that locally downscales climate data to scale-free point estimates (Wang et al. 2016). We obtained 30-year normals (from 1981 to 2010) as well as data at a monthly resolution for the post-fire water year (October 2018 to September 2019) to assess the effect of post-fire climate on regeneration success. We considered two possible measures: absolute post-fire water year climatic moisture deficit (CMD) and post-fire relative CMD (the z-score of the post-fire water year CMD as compared with the 30-year normal; Table 1). Both measures can be used to reflect moisture stress as experienced by a plant, but a z-score rather than an absolute value is often used to understand ecological effects of post-fire climate, as deviation from the long-term average climate in a location may be more important than absolute climate (e.g., Harvey et al. 2016, Stevens-Rumann et al. 2018). However, we opted to use absolute post-fire water year CMD (hereafter, post-fire CMD) in our models as exploratory analyses indicated that the 2018–2019 water year was relatively cool and wet across the study region, resulting in negative z-scores across all study sites (Additional file 1: Fig. 1.2), reducing the utility of relative post-fire CMD to reflect moisture stress. Conversely, absolute post-fire CMD represented a broad gradient of moisture stress (Table 1), allowing interpretation of post-fire climate effects across the region.
We further characterized the post-fire environment using three topographic predictors to infer plot-level topo-climate: heat load index (HLI), topographic position index (TPI), and topographic wetness index (TWI) (Table 1). HLI represents the direct incident radiation at a site (McCune and Keon 2002), TPI represents a site’s topographic position relative to neighboring cells (De Reu et al. 2013), and TWI reflects the potential moisture balance at a site (Gessler et al. 1995). We used the Geomorphometry and Gradient Metric Toolbox (Evans et al. 2014) in ArcMap version 10.6.1 (ESRI 2018) to calculate each topographic predictor from 30-m digital elevation models (U.S. Geological Survey 1999b). Soil characteristics were characterized using data from the Probabilistic Remapping of SSURGO (POLARIS) database, a spatially contiguous soils dataset of ecologically relevant soils variables (Chaney et al. 2016). For each of our plots, we extracted depth to bedrock and soil clay content (Table 1) as two candidate predictors that can influence forest productivity and affect soil moisture and fertility (Romanyà and Vallejo 2004; Wall and Westman 2006). Although serpentine soils are common across the range of knobcone pine (Vogl et al. 1977), few plots occurred in areas mapped as serpentine (Natural Resources Conservation Service [NRCS] 2021) and serpentine was therefore not considered a factor in statistical models.
Statistical analyses
To test the effects of seed supply, post-fire climate and microsite characteristics on post-fire regeneration in knobcone pine forests, we fit generalized linear mixed models (Zuur et al. 2009) for three complementary response variables: total post-fire seedling density (knobcone pine seedlings ha−1), probability of self-replacement (a binary response where post-fire seedling density < pre-fire stand density = 0 and post-fire seedling density ≥ pre-fire stand density = 1), and percent population recovery (knobcone pine post-fire seedling density as a counted proportion of pre-fire knobcone pine density). We note that our measure of self-replacement compares post-fire seedling densities to mature pre-fire stand densities, a metric that has previously been used as an indicator of resilience of serotinous populations to short-interval fires (Enright et al. 2014; Turner et al. 2019). As seedling attrition is high within the first several years post-establishment for serotinous conifers (Harvey and Holzman 2014), this measure represents an overestimate of mature stand self-replacement. For each response variable, we fit two models with separate focal predictor variables: fire interval (years between stand-replacing fires) and pre-fire canopy seed bank density, defined as open cones on standing and fallen trees killed by the most recent fire (ln [cones ha−1]). We assessed these predictors separately as they are strongly correlated (Additional file 1: Fig. 1.3), but each may be mechanistically linked to regeneration success. Each of the six models included a random effect of sample fire and the following covariates: post-fire CMD, HLI, TPI, TWI, shrub cover and soil clay content or depth to bedrock (Table 1). Total post-fire seedling density models were fit to post-fire seedling counts using a negative binomial error structure and an offset term (log[plot areas]) to account for variable plot sizes (Zuur et al. 2009). Probability of self-replacement models were fit using a binomial error structure. Percent population recovery models were fit to post-fire seedling counts using a negative binomial error structure and an offset term (log[pre-fire tree count]) as appropriate for counted proportions with no upper bound (Mangiafico 2016).
Each generalized linear mixed model was fit using standardized predictor variables (mean-centered per two standard deviations) to compare relative effects of each predictor. A full model with all candidate predictors included was fit for each response variable, and model diagnostic tests were conducted. When the full model did not meet model assumptions, we fit a suite of new models using single term deletions (models were fit with each term dropped except fire interval or pre-fire canopy seed bank). Model diagnostics were conducted on each reduced model. If all reduced models failed to meet model assumptions, this process was repeated with additional single term deletions until a model was fitted that met assumptions. When two models with an equal number of predictors met assumptions, the model with the lowest Akaike Information Criterion adjusted for small sample sizes (AICc) (Akaike 1974) was selected for inference. We interpreted P ≤ 0.01, P ≤ 0.05, and P ≤ 0.10 as strong, moderate, and suggestive evidence of an effect, respectively (Ramsey and Schafer 2012). All analyses were conducted in R version 4.0.5 (R Core Team 2021). Models were fit with the glmmTMB package (Brooks et al. 2017) and model diagnostics were conducted with DHARMa (Hartig 2021). Visualizations of model effects were created with the packages broom (Robinson et al. 2021), ggeffects (Lüdecke 2018), ggplot (Wickham 2016), ggpubr (Kassambara 2020), and jtools (Long 2020).
Results
We counted a total of 15,111 seedlings, 99% of which were knobcone pine. At most plots, 100% of the conifer regeneration was knobcone pine, though plot-level values ranged from 51 to 100% (Table 2), with most of the remaining conifer regeneration composed of Douglas-fir. Where the pre-fire stand was composed of knobcone pine and Douglas-fir, knobcone pine generally became more dominant post-fire (Table 2; Additional file 1: Fig. 1.4). 96% of knobcone pine seedlings established in the first growing season post-fire and were therefore 2 years old at the time of sampling (Table 2). We counted 51,187 cones on 4336 dead knobcone pine trees to characterize the pre-fire canopy seed bank, and pre-fire stand structure varied widely among and within fire intervals (Table 2; Additional file 1: Fig. 1.5).
Post-fire seedling density
Post-fire seedling density ranged from 138 to 250,200 seedlings ha−1 across the study area; all plots had at least some knobcone pine regeneration (Table 2). For plots with previous fire intervals of known length (fire intervals up to 31 years), there was strong evidence of a positive effect of fire interval on post-fire seedling density (Table 3; Fig. 3A), with an estimated 19% increase in seedling density for each year increase in fire interval. Between fire intervals of 6 and 31 years, there was an estimated increase of two orders of magnitude, from ~ 1000 to ~ 100,000 seedlings ha−1, respectively (Fig. 4A). Data from long unburned plots (with unknown fire history) suggest that the relationship between fire interval and post-fire seedling density is non-linear, declining at some point after ~ 31 years (the longest known fire interval, Fig. 4A). Further, there was moderate evidence of a negative effect (approximately one third of the magnitude of that of fire interval) of increasing post-fire CMD on post-fire seedling density (Fig. 3A). Holding all other predictors constant, post-fire seedling density decreased by nearly an order of magnitude between areas of lowest moisture stress (post-fire CMD = 400 mm) to highest post-fire moisture stress (post-fire CMD = 700 mm; Fig. 5A). There was no evidence of an effect of heat load index, shrub cover, or soil clay content (Figs. 3A and 5B–D).

Estimated effect sizes of predictor variables in A fire interval models and B canopy seed bank models on post-fire seedling density, probability of self-replacement, and percent population recovery. Dots represent medians, horizontal lines represent 95% confidence intervals, and both are shaded by model (response variable). The effects for each predictor are per two standard deviations

Partial effect plots of each measure of seed supply on each measure of post-fire regeneration. Panels show the partial effects of fire interval (A) and pre-fire canopy seed bank (B) on post-fire seedling density, fire interval (C) and pre-fire canopy seed bank (D) on probability of knobcone pine self-replacement, and fire interval (E) and pre-fire canopy seed bank (F) on percent population recovery, with the dashed line representing the line of self-replacement. The solid line represents the median estimate with all other covariates held at their means and the shaded region represents the 95% confidence interval. For panels A, C, and E, each point represents a plot; black points represent plots with known fire intervals used to fit the model (n = 52), gray points with unknown “long” fire intervals were not used to fit the model but are presented for reference (n = 19). For panels B, D, and F, points are shaded by fire interval and each point represents a plot (n = 71)

Change in fire interval and A–D post-fire seedling density, E–H probability of knobcone pine self-replacement, and I–L percent population recovery across gradients of covariates in each model. Solid lines represent median estimates; shaded areas represent 95% confidence intervals. Predictions consider the effect of each covariate individually, holding all other covariates at their means. Each column has a single legend except for the right column, where each panel has its own legend. Dashed lines are the line of self-replacement for panels I–L
When using canopy seed bank rather than fire interval as the predictor representing seed source, there was strong evidence of a positive effect of pre-fire canopy seed bank on post-fire seedling density (Table 3; Fig. 3B). Holding all covariates at their means, pre-fire canopy seed banks of 692, 10,900, and 47,600 cones ha−1 (representing the 10th, 50th, and 90th percentiles of canopy seed bank size, respectively) resulted in an estimated 1000, 10,000, and 35,000 knobcone pine seedlings ha−1, respectively (Fig. 4B). There was strong evidence of a positive effect of depth to bedrock (Fig. 3B), although the magnitude of the effect was approximately one fourth that of pre-fire canopy seed bank. There was an estimated four-fold increase in post-fire seedling density at plots with the deepest (~ 120 cm) versus shallowest (~ 70 cm) depth to bedrock in the study area (Fig. 6B). There was no evidence of an effect of post-fire CMD, TPI, or shrub cover in the fitted model (Figs. 3B and 6A, C, and D).

Change in pre-fire canopy seed bank and A–D post-fire seedling density, E–H probability of knobcone pine self-replacement, and I–M percent population recovery across gradients of covariates in each model. Solid lines represent median estimates; shaded areas represent 95% confidence intervals. Predictions consider the effect of each covariate individually, holding all other covariates at their means. Legends apply to all panels in each column
Probability of self-replacement
Knobcone pine self-replacement (where seedling density two years post-fire reached or exceeded pre-fire density) occurred at 68% of plots. There was strong evidence of a positive effect of fire interval on probability of self-replacement for fire intervals up to 31 years and the magnitude of this effect size was at least four times greater than that of all other predictors in the model (Table 3; Fig. 3A). Holding all covariates at their means, probability of self-replacement was estimated at < 5% for intervals < 11 years, 50% for a 16-year interval, and > 95% for intervals > 20 years (Fig. 4B). Data from long unburned plots suggest that this trend continues beyond the 31-year chronosequence (Fig. 4E). There was moderate evidence for an effect of post-fire CMD on probability of self-replacement, with an additional 4 years needed to accumulate seed to reach a given probability of self-replacement for areas of high compared to low post-fire moisture stress (Fig. 5E). There was suggestive evidence of a negative effect of mean shrub cover on probability of self-replacement—although the magnitude of the effect size was similar to that of post-fire CMD, there was considerable uncertainty around this relationship (Figs. 3A and 5F). There was no effect of TPI or soil clay content on probability of self-replacement (Figs. 3A and 5G, H). Using canopy seed bank rather than fire interval as the predictor for seed source, there was strong evidence of a positive effect of pre-fire canopy seed bank on probability of self-replacement that overrode all other predictor variables in the model (Table 3; Fig. 3B). Probability of self-replacement was estimated at 38%, 83%, and 94% for pre-fire canopy seed banks of 692, 10,900, and 47,600 cones ha−1, respectively (Fig. 4D). There was no evidence of an effect of any covariates considered in this model (Figs. 3B and 6E–H).
Percent population recovery
For fire intervals up to 31 years, the fitted model showed strong evidence of a positive effect of fire interval on post-fire knobcone pine density percent of pre-fire knobcone pine density (hereafter, percent population recovery; Table 3; Fig. 3A). Percent population recovery was estimated at 50% following a 10-year fire interval, and between 13-year to 31-year fire intervals increased from 100 to 2000% (i.e., a 20-fold increase in population density post-fire; Fig. 4E). Long-interval plots encompassed a broad range of percent population recovery values, representing threefold to 764-fold increases (median = 51-fold increase) from pre-fire to post-fire, suggesting that this metric of regeneration success may continue to increase over time beyond the bounds of the investigated fire intervals (Fig. 4E). Post-fire CMD had a strong negative effect on percent population recovery, though the magnitude of effect was approximately 30% of that of fire interval (Fig. 3A). Compared with areas of low post-fire CMD, an estimated additional 6 years was needed to reach the same percent population recovery for areas with high post-fire CMD (Fig. 5I). Conversely, HLI had a moderate positive effect (approximately 20% of the magnitude of the effect size for fire interval, Fig. 3a). Holding all remaining covariates at their means, areas with a relatively low heat load (HLI = 0.7) required an additional 4 years to reach a given value of percent population recovery compared with areas with a relatively high heat load (HLI = 0.9, Fig. 5L). There was no evidence of effects of shrub cover or soil clay content on percent population recovery (Figs. 3A and 5J, K).
Using pre-fire canopy seed bank rather than fire interval as the predictor representing seed source, there was strong evidence of a positive effect of pre-fire canopy seed bank on percent population recovery (Table 3; Fig. 3B). Percent population recovery was estimated at 100% and 1000% for pre-fire canopy seed banks of 300 and 6000 cones ha−1, respectively, holding all covariates at their means (Fig. 4F). There was strong evidence of a positive effect of depth to bedrock, with a magnitude ~ 70% of the effect of pre-fire canopy seed bank (Fig. 3B). For sites with similar pre-fire canopy seed banks, there was an estimated fourfold increase in percent population recovery at plots with the deepest (~ 120 cm) versus shallowest (~ 70 cm) depth to bedrock in the study area (Fig. 6J). Further, there was suggestive evidence of a negative effect of TWI (Fig. 6M) but overlap of the 95% confidence interval with zero suggests considerable uncertainty regarding the directionality of this effect (Fig. 3B). There was no evidence of an effect of post-fire CMD, shrub cover, or TPI (Figs. 3B and 6I, K, and L).
Discussion
By simultaneously testing the individual effects of seed supply and post-fire growing conditions on serotinous forest resilience to fire, our study highlights several key insights with implications for fire-prone ecosystems. First, seed supply at the time of fire was by far the most important factor driving each measure of post-fire forest recovery, suggesting that fire interval—an important dimension of changing fire regimes in many forests—potentially outweighs the direct effects of variation in regional climate on resilience. Second, the direct effects of first year post-fire climate conditions on post-fire forest recovery, while minor compared to the effects of seed source, can combine with seed supply to cause compounding effects near thresholds of seed availability when fire intervals are short (e.g., < 15 years). Our findings show that while post-fire tree regeneration varies widely with fire interval length, knobcone pine can persist following reburns occurring on intervals as short as six years, although the effects of additional warming and changes to the fire regime are uncertain. Finally, our findings are relevant for fire and vegetation management in serotinous forests and can help inform management goals of expanding or reducing the extent of serotinous forests.
The overriding effect of seed supply (3–4x that of other predictors) on all measures of post-fire forest recovery suggests that altered disturbance regimes will have important ecological impacts on serotinous forests. Each measure of seed supply—pre-fire canopy seed bank and fire interval—showed similar effects on post-fire regeneration outcomes. This indicates that although canopy seed banks for a given stand age can vary with stand structure (Enright and Lamont 1989; Gill et al. 2021; Agne et al. 2022), fire interval is an effective proxy when direct quantification of the pre-fire canopy seed bank is infeasible. A strong effect of fire interval on post-fire regeneration has been noted in many systems dominated by serotinous species, with 2x to several orders of magnitude greater post-fire seedling density following long versus short-interval fires (Espelta et al. 2008; Brown and Johnstone 2012; Enright et al. 2014; Turner et al. 2019; Whitman et al. 2019), as we found here. However, whereas short-interval severe reburns can lead to complete recruitment failure or conversion to non-forest where the historical fire regime was characterized by low severity fires or longer fire intervals (Coppoletta et al. 2016; Stevens-Rumann and Morgan 2016), we saw no evidence of complete recruitment failure. Even following the shortest fire interval (6 years), we found knobcone pine seedling densities that meet or exceed typical post-fire seedling densities for many non-serotinous conifer species in montane and subalpine forests in western North America (Welch et al. 2016; Harvey et al. 2016; Chambers et al. 2016). This indicates high resilience of serotinous species to increases in short-interval fire (Hansen et al. 2018) where trees have reached reproductive maturity. Post-fire regeneration at long fire intervals declined slightly, driven by a decline in the pre-fire canopy seed bank. However, post-fire seedlings per cone were stable (Additional File 1 - Fig. 1.6) and overall densities remained high; we found no evidence of senescence risk (sensu Enright et al. 2015), despite evidence that fire exclusion has led to loss of knobcone pine in parts of its range (Reilly et al. 2019). Overall, the effect of fire frequency on seed supply drives variation in post-fire regeneration, although knobcone pine appears to be resilient across a broad range of pre-fire seed supply.
Post-fire climate had a strong mediating effect on post-fire regeneration near thresholds of seed supply, suggesting that it may play an increasingly important role in the future as warm post-fire climate occurs more frequently. Increases in post-fire CMD had a significant negative effect across post-fire regeneration response variables, consistent with findings across North American coniferous forests (Harvey et al. 2016; Kemp et al. 2019; Davis et al. 2019; Rodman et al. 2019). Further, the high proportion of seedlings measured as recruited in the first year post-fire (~ 95%) supports the importance of suitable climate conditions in the first post-fire growing season, because the vast majority of seed of serotinous species is released immediately post-fire (Vogl 1973; Enright and Lamont 1989; Johnson and Fryer 1989; Greene et al. 1999; Nathan and Ne’eman 2004). However, the lack of complete recruitment failure at any site suggests knobcone pine forests are more resilient to harsh post-fire conditions than many forests in which incidence of zero conifer regeneration post-fire has increased in recent decades (Stevens-Rumann et al. 2018). The relative magnitude of the effect of post-fire climate suggests that this factor is unlikely to change regeneration outcomes with respect to self-replacement at very short or long fire intervals, where seed supply is limiting or available in a large amount. Therefore, post-fire climate is most likely to have an ecologically meaningful effect on stands near the self-replacement threshold in cases where seed supply is intermediate, such as stands that burn on ~ 13–18-year fire intervals. Our study examined climate conditions across a broad gradient of moisture stress in a single, relatively wet, post-fire year (Table 1); additional work is needed to understand the effect of post-fire drought conditions on post-fire regeneration. Post-fire climate conditions may increasingly constrain regeneration as short-interval fires continue to occur in stands with intermediate seed supply. Future work could be undertaken to integrate seed supply and projected climate in vulnerability mapping to anticipate where post-fire regeneration may decline or fail.
The effects of local abiotic and biotic drivers on post-fire regeneration outcomes were generally minor compared with seed supply and post-fire climate. Several studies have shown the importance of topography (Hansen and Turner 2019; Littlefield 2019), which can compound with post-fire climate and a shortened fire interval to create a harsh microclimate for seedlings (Hoecker et al. 2020). However, we found a negligible effect of topography on post-fire regeneration, due to low variation in the variables assessed (Table 1), associated with the topo-edaphic restriction of knobcone pine (Vogl 1973). While knobcone pine can establish well in shallow, nutrient-poor soils (Vogl et al. 1977), seedling growth rates are greater in deep versus shallow soils (Vogl 1973). Our finding of greater post-fire regeneration for a given seed supply with increased depth to bedrock suggests that such areas may be further buffered against low regeneration should a short-interval severe fire occur, while areas with shallow soils are more vulnerable to decreases in forest cover. Shrub cover often has a strong negative effect on early post-fire conifer regeneration (Collins and Roller 2013; Welch et al. 2016), due to strong competition for light and soil moisture in the first several years post-fire (Knapp et al. 2012). Contrary to expectation, given the shade intolerance and poor competitive ability of knobcone pine (Howard 1992), we found limited evidence of an effect of shrub cover on post-fire regeneration. Early post-fire establishment and the relatively fast growth rate of knobcone pine may mask competitive effects, making them difficult to detect until several years post-fire, as has occurred in similar serotinous pine forests (Harvey and Holzman 2014). Overall, site conditions had minor effects on post-fire regeneration, but could play an important role in localized population persistence in some cases.
Although our study identifies apparent thresholds of fire frequency and post-fire climate that could lead to loss of knobcone pine resilience, this species is well-suited to current climate and fire regimes. Short-interval reburns capable of decreasing knobcone pine density can occur, but fire intervals < 10 years represented 4% of reburned area between 1985 and 2015 (Reilly et al. 2019) and 8.5% of reburned area in 2018 (M.C. Agne, unpublished data). Further, many systems dominated by serotinous species follow multiple pathways of stand structural development (Harvey and Holzman 2014; Turner et al. 2016). Oscillation between low- and high-density stand structures over multiple fire cycles is typical for serotinous populations (Esler and Cowling 1990), and decreased population density following a single short-interval fire may not represent resilience loss over a longer temporal scale. Despite clear potential for knobcone pine forests to burn on relatively short intervals, the likelihood of short-interval reburns that would threaten resilience on a broad spatial scale is currently low. Rather, the potential for local knobcone pine densification following severe reburns is high, especially when co-occurring with non-serotinous conifers (Buma et al. 2013). Our study supports this point, as knobcone pine increased in dominance post-fire where it co-occurred with Douglas-fir (Additional file 1: Fig. 1.4). Some knobcone pine range expansion has already occurred (Reilly et al. 2019) and future fire conditions may facilitate further range expansion or changes in forest composition, especially where knobcone pine is currently a poor competitor.
Our findings have implications for better understanding the historical fire regime of knobcone pine. First, the sparse regeneration following fire intervals of 6–10 years suggests that fire occurrence at this frequency or greater was unlikely to support knobcone pine and are indicative of a contemporary departure from the historical range of variability. Second, that regeneration density was at self-replacement following fire intervals of ~ 13–18 years suggests that such fire intervals could have included the historical minimum fire interval for knobcone pine forests, and that previous estimates of the historical minimum interval (30 years; van de Water and Safford 2011) may be too high. However, rapid and intense seedling attrition beyond 2 years post-fire is common in serotinous forests (e.g., Harvey and Holzman 2014) and can vary widely with post-fire climate, local topography, and soil conditions (Hansen and Turner 2019; Hoecker et al. 2020; Carroll et al. 2021). Given that these losses are not yet accounted for in our data, densities could later fall below self-replacement at ~ 13–18-year fire intervals, as has been suggested in previous work (Enright et al. 2014). Post-fire seedling attrition rates are expected to increase with climate warming, raising the possibility that fire intervals within this range could erode resilience. Future tracking of long-term population dynamics between fire intervals is an important area of research to better understand the historical range of fire return intervals for knobcone pine and how this relates to warming climate and increasing fire potential.
Our study has implications for the management of knobcone pine, and serotinous forests more broadly, as fire regimes change. Fires occurring on at least 15-year intervals (or several years longer where post-fire moisture stress is high) should foster self-replacement or potential expansion of knobcone pine. Occurrence of fire on shorter intervals can achieve the aim of density reduction, although there are likely to be challenges with implementation of prescribed burning in this forest type (Fry et al. 2012). Severe reburns within 4–5 years of a previous high severity fire, prior to cone development and maturation (Keeley et al. 1999; Agne et al. 2022), could lead to local eradication. However, fuel limitation lasting at least 5 years after severe fire has been observed in lower montane forests in Oregon and California (Donato et al. 2013; Coppoletta et al. 2016), suggesting that reburns are unlikely to recur on this interval. Recurrent short-interval severe reburns may eventually remove knobcone pine from a site (Keeley et al. 1999), but it is unclear how such an increase in fire activity would affect fuel loads and subsequent reburn potential in the long term (Hurteau et al. 2019; Abatzoglou et al. 2021). Further, the potential for increases in non-native species may increase with increased fire activity (Smith et al. 2019), and transition to non-forest is likely if fire is sufficiently frequent to eradicate knobcone pine from a forest stand.
Our work has implications for resilience to increasing fire frequency in serotinous forests across the world. Seed supply is likely the primary constraint on post-fire regeneration in other forests historically dominated by very high densities of serotinous species following severe fire. We expect that where historical fire intervals were relatively short and seed supply begins to accumulate soon after fire, as with knobcone pine, Aleppo pine (Pinus halepensis), and bishop pine (Pinus muricata) (Keeley et al. 1999; Tapias et al. 2001; Agne et al. 2022), serotinous conifer species should be highly resilient to severe short-interval fires, despite potential for localized population decreases. However, where historical fire intervals were longer, as with lodgepole pine (Pinus contorta) and black spruce (Picea mariana), increases in severe fire frequency can lead to significant decreases in the serotinous population, and altered stand structure and species composition (Brown and Johnstone 2012; Turner et al. 2019; Whitman et al. 2019). Where persistent declines in moisture availability have already occurred, as in southwest Australia, serotinous species are threatened by direct depletion of seed supply by climate warming (Enright et al. 2014, 2015). For species such as Banskia hookeriana, for which the time to optimal canopy seed bank storage for population self-replacement was similar to the historical fire interval, decreases in fire interval and seed supply for a given stand age threaten the species across its range (Enright et al. 1996). Understanding drivers of demographic rates for individual species in the context of current and expected future fire activity is key to anticipating effects of both direct climate warming and altered fire regimes on serotinous species persistence.
Conclusion
Seed supply was the key constraint on post-fire regeneration by all metrics, suggesting that effects of climate change on serotinous forests are likely to occur primarily through indirect effects on changes to fire regimes. The effect of post-fire climate was small in magnitude relative to that of seed supply, but climate may affect post-fire resilience around thresholds of seed supply. While knobcone pine failed to regenerate at pre-fire densities at the shortest fire intervals, this species is generally resilient to its current fire regime and climate. Conversely, serotinous species with longer historical fire return intervals and time to maturity may face greater risk of post-fire resilience loss. Changes in post-fire regeneration have implications for post-fire forest structure and reproductive capacity as well as future fire potential, critical to understanding serotinous forest resilience to changing conditions over multiple fire intervals.
Availability of data and materials
The datasets used during the current study are available from the corresponding author upon reasonable request.
Abbreviations
- AICc:
-
Akaike Information Criterion adjusted for small sample sizes
- asl:
-
Above sea level
- CMD:
-
Climatic moisture deficit
- DBH:
-
Diameter at breast height
- HLI:
-
Heat load index
- MFRI:
-
Mean fire return interval
- MTBS:
-
Monitoring Trends in Burn Severity
- TPI:
-
Topographic position index
- TWI :
-
Topographic wetness index
References
Abatzoglou, J.T., and A.P. Williams. 2016. Impact of anthropogenic climate change on wildfire across western US forests. Proc Natl Acad Sci U S A 113: 11770–11775. https://doi.org/10.1073/pnas.1607171113.
Abatzoglou, J. T., D. S. Battisti, A. P. Williams, W. D. Hansen, B. J. Harvey, and C. A. Kolden. 2021. Projected increases in western US forest fire despite growing fuel constraints. Commun Earth Environ 2: 227. https://doi.org/10.1038/s43247-021-00299-0.
Agne, M. C., J. B. Fontaine, N. J. Enright, S. M. Bisbing, and B. J. Harvey. 2022. Demographic processes underpinning post-fire resilience in California closed-cone pine forests: The importance of fire interval, stand structure, and climate. Plant Ecol 223: 751-767. https://doi.org/10.1007/s11258-022-01228-7.
Akaike, H. 1974. A new look at the statistical model identification. IEEE Trans Auto Control 19: 716–723. https://doi.org/10.1109/TAC.1974.1100705.
Allen, C. D., D. D. Breshears, and N. G. McDowell. 2015. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6: art129. https://doi.org/10.1890/ES15-00203.1.
Baltzer, J.L., N.J. Day, X.J. Walker, D. Greene, M.C. Mack, H.D. Alexander, D. Arseneault, J. Barnes, Y. Bergeron, Y. Boucher, L. Bourgeau-Chavez, C.D. Brown, S. Carrière, B.K. Howard, S. Gauthier, M.-A. Parisien, K.A. Reid, B.M. Rogers, C. Roland, L. Sirois, S. Stehn, D.K. Thompson, M.R. Turetsky, S. Veraverbeke, E. Whitman, J. Yang, and J.F. Johnstone. 2021. Increasing fire and the decline of fire adapted black spruce in the boreal forest. Proc Natl Acad Sci U S A 118: e2024872118. https://doi.org/10.1073/pnas.2024872118.
Boag, A.E., M.J. Ducey, M.W. Palace, and J. Hartter. 2020. Topography and fire legacies drive variable post-fire juvenile conifer regeneration in eastern Oregon, USA. Forest Ecol Manage 474: 118312. https://doi.org/10.1016/j.foreco.2020.118312.
Boer, M. M., V. Resco de Dios, and R. A. Bradstock. 2020. Unprecedented burn area of Australian mega forest fires. Nat Clim Chang 10: 170–172. https://doi.org/10.1038/s41558-020-0710-7.
Brooks, M. E., K. Kristensen, K. J. van Benthem, A. Magnusson, C. W. Berg, A. Nielsen, H. J. Skaug, M. Maechler, and B. M. Bolker. 2017. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9: 378–400.
Brown, C.D., and J.F. Johnstone. 2012. Once burned, twice shy: Repeat fires reduce seed availability and alter substrate constraints on Picea mariana regeneration. Forest Ecol Manage 266: 34–41. https://doi.org/10.1016/j.foreco.2011.11.006.
Buma, B., C. D. Brown, D. C. Donato, J. B. Fontaine, and J. F. Johnstone. 2013. The impacts of changing disturbance regimes on serotinous plant populations and communities. BioScience 63: 866–876. https://doi.org/10.1525/bio.2013.63.11.5.
Busby, S. U., K. B. Moffett, and A. Holz. 2020. High-severity and short-interval wildfires limit forest recovery in the Central Cascade Range. Ecosphere 11: e03247. https://doi.org/10.1002/ecs2.3247.
Carroll, C. J. W., A. K. Knapp, and P. H. Martin. 2021. Higher temperatures increase growth rates of Rocky Mountain montane tree seedlings. Ecosphere 12: e03414. https://doi.org/10.1002/ecs2.3414.
Chambers, M.E., P.J. Fornwalt, S.L. Malone, and M.A. Battaglia. 2016. Patterns of conifer regeneration following high severity wildfire in ponderosa pine – dominated forests of the Colorado Front Range. Forest Ecol Manage 378: 57–67. https://doi.org/10.1016/j.foreco.2016.07.001.
Chaney, N. W., E. F. Wood, A. B. McBratney, J. W. Hempel, T. W. Nauman, C. W. Brungard, and N. P. Odgers. 2016. POLARIS: A 30-meter probabilistic soil series map of the contiguous United States. Geoderma 274: 54–67. https://doi.org/10.1016/j.geoderma.2016.03.025.
Collins, B.M., and G.B. Roller. 2013. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landscape Ecol 28: 1801–1813. https://doi.org/10.1007/s10980-013-9923-8.
Collins, L., R.A. Bradstock, H. Clarke, M.F. Clarke, R.H. Nolan, and T.D. Penman. 2021. The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire. Environ Res Lett 16: 044029. https://doi.org/10.1088/1748-9326/abeb9e.
Collins, L., H. Clarke, M.F. Clarke, S.C. McColl Gausden, R.H. Nolan, T. Penman, and R. Bradstock. 2022. Warmer and drier conditions have increased the potential for large and severe fire seasons across south-eastern Australia. Global Ecol Biogeography. https://doi.org/10.1111/geb.13514.
Coop, J. D., S. A. Parks, C. S. Stevens-Rumann, S. D. Crausbay, P. E. Higuera, M. D. Hurteau, A. Tepley, E. Whitman, T. Assal, B. M. Collins, K. T. Davis, S. Dobrowski, D. A. Falk, P. J. Fornwalt, P. Z. Fulé, B. J. Harvey, V. R. Kane, C. E. Littlefield, E. Q. Margolis, M. North, M.-A. Parisien, S. Prichard, and K. C. Rodman. 2020. Wildfire-driven forest conversion in western North American landscapes. Bioscience 70: 659–673. https://doi.org/10.1093/biosci/biaa061.
Coppoletta, M., K.E. Merriam, and B.M. Collins. 2016. Post-fire vegetation and fuel development influences fire severity patterns in reburns. Ecol Appl 26: 686–699. https://doi.org/10.1890/15-0225.
Crotteau, J.S., J. Morgan Varner, and M.W. Ritchie. 2013. Post-fire regeneration across a fire severity gradient in the southern Cascades. Forest Ecol Manage 287: 103–112. https://doi.org/10.1016/j.foreco.2012.09.022.
Davis, K.T., S.Z. Dobrowski, P.E. Higuera, Z.A. Holden, T.T. Veblen, M.T. Rother, S.A. Parks, A. Sala, and M.P. Maneta. 2019. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc Natl Acad Sci U S A 116: 6193–6198. https://doi.org/10.1073/pnas.1815107116.
De Reu, J., J. Bourgeois, M. Bats, A. Zwertvaegher, V. Gelorini, P. De Smedt, W. Chu, M. Antrop, P. De Maeyer, P. Finke, M. Van Meirvenne, J. Verniers, and P. Crombé. 2013. Application of the topographic position index to heterogeneous landscapes. Geomorphology 186: 39–49. https://doi.org/10.1016/j.geomorph.2012.12.015.
Donato, D.C., J.B. Fontaine, W.D. Robinson, J.B. Kauffman, and B.E. Law. 2009. Vegetation response to a short interval between high-severity wildfires in a mixed-evergreen forest. J Ecol 97: 142–154. https://doi.org/10.1111/j.1365-2745.2008.01456.x.
Donato, D. C., J. B. Fontaine, J. B. Kauffman, W. D. Robinson, and B. E. Law. 2013. Fuel mass and forest structure following stand-replacement fire and post-fire logging in a mixed-evergreen forest. Int J Wildland Fire 22: 652. https://doi.org/10.1071/WF12109.
Eidenshink, J., B. Schwind, K. Brewer, Z.-L. Zhu, B. Quayle, and S. Howard. 2007. A project for monitoring trends in burn severity. Fire Ecol 3: 3–21. https://doi.org/10.4996/fireecology.0301003.
Enright, N. J., and B. B. Lamont. 1989. Seed banks, fire season, safe sites and seedling recruitment in five co-occurring Banksia species. J Ecol 77: 1111–1122. https://doi.org/10.2307/2260826.
Enright, N.J., B.B. Lamont, and R. Marsula. 1996. Canopy seed bank dynamics and optimum fire regime for the highly serotinous shrub, Banksia hookeriana. J Ecol 84: 9. https://doi.org/10.2307/2261695.
Enright, N.J., J.B. Fontaine, B.B. Lamont, B.P. Miller, and V.C. Westcott. 2014. Resistance and resilience to changing climate and fire regime depend on plant functional traits. J Ecol 102: 1572–1581. https://doi.org/10.1111/1365-2745.12306.
Enright, N.J., J.B. Fontaine, D.M. Bowman, R.A. Bradstock, and R.J. Williams. 2015. Interval squeeze: altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front Ecol Environ 13: 265–272. https://doi.org/10.1890/140231.
Esler, K.J., and R.M. Cowling. 1990. Effects of density on the reproductive output of Protea lepidocarpodendron. South+A1443 Afr J Botany 56: 29–33. https://doi.org/10.1016/S0254-6299(16)31107-3.
Espelta, J. M., I. Verkaik, M. Eugenio, and F. Lloret. 2008. Recurrent wildfires constrain long-term reproduction ability in Pinus halepensis Mill. Int J Wildland Fire 17: 579. https://doi.org/10.1071/WF07078.
ESRI [Environmental Systems Research Institute]. 2018. ArcGIS Release 10.6.1. Redlands: ESRI.
Evans, J. S., J. Oakleaf, S. A. Cushman, and D. Theobald. 2014. An ArcGIS toolbox for surface gradient and geomorphometric modeling, version 2.0–0. http://evansmurphy.wix.com/evansspatial. Accessed 21 Jan 2021.
Falk, D.A., P.J. van Mantgem, J.E. Keeley, R.M. Gregg, C.H. Guiterman, A.J. Tepley, J.N. Young, and D. Marshall. 2022. Mechanisms of forest resilience. Forest Ecol Manage 512: 120129. https://doi.org/10.1016/j.foreco.2022.120129.
Fry, D.L., and S.L. Stephens. 2006. Influence of humans and climate on the fire history of a ponderosa pine-mixed conifer forest in the southeastern Klamath Mountains, California. Forest Ecol Manage 223: 428–438. https://doi.org/10.1016/j.foreco.2005.12.021.
Fry, D. L., J. Dawson, and S. L. Stephens. 2012. Age and structure of mature knobcone pine forests in the northern California Coast Range, USA. Fire Ecol 8: 49–62. https://doi.org/10.4996/fireecology.0801049.
Gessler, P. E., I. D. Moore, N. J. McKenzie, and P. J. Ryan. 1995. Soil-landscape modelling and spatial prediction of soil attributes. Int J Geogr Inf Syst 9: 421–432. https://doi.org/10.1080/02693799508902047.
Gill, N. S., T. J. Hoecker, and M. G. Turner. 2021. The propagule doesn’t fall far from the tree, especially after short-interval, high-severity fire. Ecology 102: e03194. https://doi.org/10.1002/ecy.3194.
Greene, D.F., J.C. Zasada, L. Sirois, D. Kneeshaw, H. Morin, and I. Charron. 1999. A review of the regeneration dynamics of North American boreal forest tree species. Can J Forest Res 29: 824–839.
Guiterman, C. H., E. Q. Margolis, C. D. Allen, D. A. Falk, and T. W. Swetnam. 2018. Long-term persistence and fire resilience of oak shrubfields in dry conifer forests of northern New Mexico. Ecosystems 21: 943–959. https://doi.org/10.1007/s10021-017-0192-2.
Gunderson, L.H. 2000. Ecological resilience—in theory and application. Ann Rev Ecol Syst 31: 425–439. https://doi.org/10.1146/annurev.ecolsys.31.1.425.
Hansen, W.D., and M.G. Turner. 2019. Origins of abrupt change? Postfire subalpine conifer regeneration declines nonlinearly with warming and drying. Ecol Monographs 89: e01340. https://doi.org/10.1002/ecm.1340.
Hansen, W. D., K. H. Braziunas, W. Rammer, R. Seidl, and M. G. Turner. 2018. It takes a few to tango: Changing climate and fire regimes can cause regeneration failure of two subalpine conifers. Ecology 99: 966–977. https://doi.org/10.1002/ecy.2181.
Hartig, F. 2021. DHARMa: Residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.4.0. https://CRAN.R-project.org/package=DHARMa. Accessed 11 Nov 2021.
Harvey, B.J., and B.A. Holzman. 2014. Divergent successional pathways of stand development following fire in a California closed-cone pine forest. J Vegetation Sci 25: 88–99. https://doi.org/10.1111/jvs.12073.
Harvey, B. J., D. C. Donato, and M. G. Turner. 2016. High and dry: Post-fire tree seedling establishment in subalpine forests decreases with post-fire drought and large stand-replacing burn patches: Drought and post-fire tree seedlings. Global Ecol Biogeogr 25: 655–669. https://doi.org/10.1111/geb.12443.
Hoecker, T.J., W.D. Hansen, and M.G. Turner. 2020. Topographic position amplifies consequences of short-interval stand-replacing fires on postfire tree establishment in subalpine conifer forests. Forest Ecol Manage 478: 118523. https://doi.org/10.1016/j.foreco.2020.118523.
Howard, J. L. 1992. Pinus attenuata. Fire Sciences Laboratory: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Hurteau, M.D., S. Liang, A.L. Westerling, and C. Wiedinmyer. 2019. Vegetation-fire feedback reduces projected area burned under climate change. Sci Rep 9: 2838. https://doi.org/10.1038/s41598-019-39284-1.
Johnson, E. A., and G. I. Fryer. 1989. Population dynamics in lodgepole pine-Engelmann spruce forests. Ecology 70: 1335–1345. https://doi.org/10.2307/1938193.
Jolly, W.M., M.A. Cochrane, P.H. Freeborn, Z.A. Holden, T.J. Brown, G.J. Williamson, and D.M.J.S. Bowman. 2015. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat Commun 6: 7537. https://doi.org/10.1038/ncomms8537.
Kassambara, A. 2020. ggpubr: “ggplot2” Based Publication Ready Plots. R package version 0.4.0. https://CRAN.R-project.org/package=ggpubr. Accessed 2 Feb 2021.
Keeley, J. E., and P. Zedler. 1998. Life history evolution in pines. In Ecology and biogeography of Pinus, ed. D. M. Richardson, 219–242. Cambridge: Cambridge University Press.
Keeley, J. E., G. Ne’eman, and C. J. Fotheringham. 1999. Immaturity risk in a fire-dependent pine. J Mediterr Ecol 1: 41–48.
Kemp, K.B., P.E. Higuera, and P. Morgan. 2016. Fire legacies impact conifer regeneration across environmental gradients in the U.S. northern Rockies. Landscape Ecol 31: 619–636. https://doi.org/10.1007/s10980-015-0268-3.
Kemp, K. B., P. E. Higuera, P. Morgan, and J. T. Abatzoglou. 2019. Climate will increasingly determine post-fire tree regeneration success in low-elevation forests. Northern Rockies USA Ecosphere 10: e02568. https://doi.org/10.1002/ecs2.2568.
Knapp, E. E., C. P. Weatherspoon, and C. N. Skinner. 2012. Shrub seed banks in mixed conifer forests of northern California and the role of Fire in regulating abundance. Fire Ecol 8: 32–48. https://doi.org/10.4996/fireecology.0801032.
Lamont, B.B., D.C.L. Maitre, R.M. Cowling, and N.J. Enright. 1991. Canopy seed storage in woody plants. Botanical Rev 57: 277–317.
Littlefield, C. E. 2019. Topography and post-fire climatic conditions shape spatio-temporal patterns of conifer establishment and growth. Fire Ecol 15: 34. https://doi.org/10.1186/s42408-019-0047-7.
Long, J. A. 2020. jtools: Analysis and presentation of social scientific data. R package version 2.1.0. https://cran.r-project.org/package=jtools. Accessed 1 Apr 2021.
Lüdecke, D. 2018. ggeffects: tidy data frames of marginal effects from regression models. J Open Source Softw 3: 772. https://doi.org/10.21105/joss.00772.
Mangiafico, S. S. 2016. Summary and analysis of extension program evaluation in R, version 1.19.10. https://rcompanion.org/handbook/. Accessed 5 Apr 2021.
Matusick, G., K.X. Ruthrof, J. Kala, N.C. Brouwers, D.D. Breshears, and G.E.S.J. Hardy. 2018. Chronic historical drought legacy exacerbates tree mortality and crown dieback during acute heatwave-compounded drought. Environ Res Lett 13: 095002. https://doi.org/10.1088/1748-9326/aad8cb.
McCune, B., and D. Keon. 2002. Equations for potential annual direct incident radiation and heat load. J Vegetation Sci 13: 603–606.
Nathan, R., and G. Ne’eman. 2004. Spatiotemporal dynamics of recruitment in Aleppo pine (Pinus halepensis Miller). Plant Ecol 171: 123–137.
Natural Resources Conservation Service [NRCS]. 2021. Soil Survey Geographic (SSURGO) Database. United States Department of Agriculture. https://data.nal.usda.gov/dataset/soil-survey-geographic-database-ssurgo. Accessed 8 Jan 2021.
Paine, R. T., M. J. Tegner, and E. A. Johnson. 1998. Compounded perturbations yield ecological surprises. Ecosystems 1: 535–545. https://doi.org/10.1007/s100219900049.
Parks, S. A., and J. T. Abatzoglou. 2020. Warmer and drier fire seasons contribute to increases in area burned at high severity in western US forests from 1985 to 2017. Geophysical Reseach Letters 47:e2020GL089858. https://doi.org/10.1029/2020GL089858.
PRISM Climate Group. 2019. Oregon State University. http://prism.oregonstate.edu. Accessed 12 Dec 2019.
Ramsey, F., and D. Schafer. 2012. The statistical sleuth: A course in methods of data analysis, 3rd Edition. Boston: Cengage Learning.
R Core Team. 2021. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Reilly, M. J., V. J. Monleon, E. S. Jules, and R. J. Butz. 2019. Range-wide population structure and dynamics of a serotinous conifer, knobcone pine (Pinus attenuata L.), under an anthropogenically-altered disturbance regime. Forest Ecol Manag 441: 182–191. https://doi.org/10.1016/j.foreco.2019.03.017.
Robinson, D., A. Hayes, and S. Couch. 2021. broom: Convert statistical objects into tidy tibbles. R package version 0.7.6. https://CRAN.R-project.org/package=broom.
Rodman, K.C., T.T. Veblen, M.A. Battaglia, M.E. Chambers, P.J. Fornwalt, Z.A. Holden, T.E. Kolb, J.R. Ouzts, and M.T. Rother. 2020. A changing climate is snuffing out post-fire recovery in montane forests. Global Ecol Biogeography 29: 2039–2051. https://doi.org/10.1111/geb.13174.
Rodman, K. C., T. T. Veblen, T. B. Chapman, M. T. Rother, A. P. Wion, and M. D. Redmond. 2020. Limitations to recovery following wildfire in dry forests of southern Colorado and northern New Mexico, USA. Ecol Appl. 30: e02001. https://doi.org/10.1002/eap.2001.
Romanyà, J., and V. R. Vallejo. 2004. Productivity of Pinus radiata plantations in Spain in response to climate and soil. Forest Ecol Manag 195: 177–189. https://doi.org/10.1016/j.foreco.2004.02.045.
Seidl, R., D. Thom, M. Kautz, D. Martin-Benito, M. Peltoniemi, G. Vacchiano, J. Wild, D. Ascoli, M. Petr, J. Honkaniemi, M.J. Lexer, V. Trotsiuk, P. Mairota, M. Svoboda, M. Fabrika, T.A. Nagel, and C.P.O. Reyer. 2017. Forest disturbances under climate change. Nat Clim Change 7: 395–402. https://doi.org/10.1038/nclimate3303.
Smith, A. G., B. A. Newingham, A. T. Hudak, and B. C. Bright. 2019. Got shrubs? Precipitation mediates long-term shrub and introduced grass dynamics in chaparral communities after fire. Fire Ecol 15: 12. https://doi.org/10.1186/s42408-019-0031-2.
Stevens-Rumann, C., and P. Morgan. 2016. Repeated wildfires alter forest recovery of mixed-conifer ecosystems. Ecol Appl 26: 1842–1853. https://doi.org/10.1890/15-1521.1.
Stevens-Rumann, C.S., K.B. Kemp, P.E. Higuera, B.J. Harvey, M.T. Rother, D.C. Donato, P. Morgan, and T.T. Veblen. 2018. Evidence for declining forest resilience to wildfires under climate change. Ecol Lett 21: 243–252. https://doi.org/10.1111/ele.12889.
Tapias, R., L. Gil, P. Fuentes-Utrilla, and J. A. Pardos. 2001. Canopy seed banks in Mediterranean pines of south-eastern Spain: a comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J Ecol 89: 629–638. https://doi.org/10.1046/j.1365-2745.2001.00575.x.
Taylor, A.H., and C.N. Skinner. 1998. Fire history and landscape dynamics in a late-successional reserve, Klamath Mountains, California, USA. Forest Ecol Manage 111: 285–301. https://doi.org/10.1016/S0378-1127(98)00342-9.
Trumbore, S., P. Brando, and H. Hartmann. 2015. Forest health and global change. Science 349: 814–818. https://doi.org/10.1126/science.aac6759.
Turner, M. G., W. H. Romme, and R. H. Gardner. 1999. Prefire heterogeneity, fire severity, and early postfire plant reestablishment in subalpine forests of Yellowstone National Park, Wyoming. Int J Wildland Fire 9: 21-36. https://doi.org/10.1071/WF99003.
Turner, M. G., T. G. Whitby, D. B. Tinker, and W. H. Romme. 2016. Twenty-four years after the Yellowstone Fires: Are postfire lodgepole pine stands converging in structure and function? Ecology 97: 1260–1273. https://doi.org/10.1890/15-1585.1.
Turner, M.G., K.H. Braziunas, W.D. Hansen, and B.J. Harvey. 2019. Short-interval severe fire erodes the resilience of subalpine lodgepole pine forests. Proc Natl Acad Sci U S A 116: 11319–11328. https://doi.org/10.1073/pnas.1902841116.
U.S. Geological Survey. 1999a. Digital representation of “Atlas of United States Trees” by Little, E. L. Jr. http://climchange.cr.usgs.gov/data/atlas/little/. Accessed 24 Jan 2018.
U.S. Geological Survey. 1999b. National elevation dataset. https://apps.nationalmap.gov/3depdem/ Accessed 1 Feb 2021.
van de Water, K. M., and H. D. Safford. 2011. A summary of fire frequency estimates for California vegetation before Euro-American settlement. Fire Ecol 7: 26–58. https://doi.org/10.4996/fireecology.0703026.
Vogl, R.J. 1973. Ecology of knobcone pine in the Santa Ana Mountains, California. Ecol Monographs 43: 125–143. https://doi.org/10.2307/1942191.
Vogl, R. J., W. P. Armstrong, K. L. White, and K. L. Cole. 1977. The closed-cone pines and cypress. In Terrestrial Vegetation of California, eds. Michael G. Barbour, and Jack Major, 295–358. New York: John Wiley and Sons.
Wall, A., and C.J. Westman. 2006. Site classification of afforested arable land based on soil properties for forest production. Can J Forest Res 36: 1451–1460. https://doi.org/10.1139/x06-031.
Wang, T., A. Hamann, D. Spittlehouse, and C. Carroll. 2016. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS One 11: e0156720. https://doi.org/10.1371/journal.pone.0156720.
Welch, K. R., H. D. Safford, and T. P. Young. 2016. Predicting conifer establishment post wildfire in mixed conifer forests of the North American Mediterranean-climate zone. Ecosphere 7: e01609. https://doi.org/10.1002/ecs2.1609.
Whitman, E., M.-A. Parisien, D.K. Thompson, and M.D. Flannigan. 2019. Short-interval wildfire and drought overwhelm boreal forest resilience. Sci Rep 9: 18796. https://doi.org/10.1038/s41598-019-55036-7.
Wickham, H. 2016. ggplot2: Elegant graphics for data analysis. New York: Springer-Verlag.
Williams, A.P., C.D. Allen, A.K. Macalady, D. Griffin, C.A. Woodhouse, D.M. Meko, T.W. Swetnam, S.A. Rauscher, R. Seager, H.D. Grissino-Mayer, J.S. Dean, E.R. Cook, C. Gangodagamage, M. Cai, and N.G. McDowell. 2013. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Change 3: 292–297. https://doi.org/10.1038/nclimate1693.
Zuur, A. F., E. N. Ieno, N. J. Walker, A. A. Saveliev, and G. M. Smith. 2009. Mixed effects models and extensions in ecology and R. New York, New York, USA: Springer.
Acknowledgements
We thank D. Donato, S. Drury, E. Engber, J. Gibson, M. Johnson, and K. Kopper for supporting this research. We benefited from insightful discussions and feedback on this project from M. Buonanduci and K. Rodman. We thank F. Nelson and K. Stork for invaluable field support. We acknowledge three anonymous reviewers whose comments improved this manuscript. Funding for this project was provided by the Australian Research Council award DP170101288 to NJE and JBF and a Murdoch University Strategic PhD Scholarship to MCA, the Achievement Rewards for College Scientists (ARCS) Foundation, the Graduate Research Innovation Award from the Joint Fire Science Program (award #19-1-01-16) to MCA and BJH, and the University of Washington. This research was funded by a U.S. Geological Survey Northwest Climate Adaptation Science Center award G17AC000218 to MCA and BJH. BJH acknowledges support from the Jack Corkery and George Corkery Jr. Endowed Professorship in Forest Sciences.
Funding
This research was unded by the Australian Research Council award DP170101288 to NJE and JBF and a Murdoch University Strategic PhD Scholarship to MCA, a U.S. Geological Survey Northwest Climate Adaptation Science Center award G17AC000218 to BJH and MCA, the Achievement Rewards for College Scientists (ARCS) Foundation to MCA, the Graduate Research Innovation Award from the Joint Fire Science Program (award #19-1-01-16) to BJH and MCA, and the University of Washington.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection and analysis were performed by MA. The first draft of the manuscript was written by MA and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Extended results from our study in post-fire knobcone pine forests in Oregon and California, USA.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Agne, M.C., Fontaine, J.B., Enright, N.J. et al. Fire interval and post-fire climate effects on serotinous forest resilience. fire ecol 18, 22 (2022). https://doi.org/10.1186/s42408-022-00145-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-022-00145-4