
Abstract
Background
Bats are important components of forested ecosystems and are found in forests worldwide. Consequently, they often interact with fire. Previous reviews of the effects of fire on bats have focused on prescribed fire effects, in part due to the limited number of studies on bat responses to wildfire. However, over the past several years, studies on bat responses to wildfire and prescribed fire have increased considerably. We reviewed this rapidly expanding body of literature to determine whether bats respond differently to prescribed fire and wildfire, and the important factors driving those differences. We also examined regional similarities and differences in bat response to prescribed fire and wildfire and identified areas in need of further research.
Results
Our review included 52 studies (29 prescribed fire, 23 wildfire) from North and South America, Europe, Australia, and Africa, although studies from Europe, South America, and Africa were limited. In general, we found that bats show positive or neutral responses to prescribed fire, whereas a greater proportion of negative responses were reported for wildfire. However, some of the negative responses to wildfire are short-lived or local, suggesting that bats may be resilient to the effects of fire. Factors such as fire severity, fire frequency, time since last burn, burn extent, season of burn, and pyrodiversity were all found to be important drivers of bats’ responses to both prescribed fire and wildfire.
Conclusions
The importance of the spatial and temporal aspects of fire suggests that these factors need to be considered when designing future studies and interpreting results. Pyrodiversity may be a particularly important concept to further our understanding of bats’ responses to fire. We found several gaps in our knowledge including lack of information on direct effects of fire (e.g., mortality), regional and taxonomic biases, effects of wildfire on roosting habitat, and the effects of climate change. Although current studies suggest that fire may be an important management tool for improving bat habitat, the threat of more frequent, extensive, and severe wildfires may put additional stress on some bat populations, particularly those being impacted by disease, habitat loss and fragmentation, and climate change.
Resumen
Antecedentes
Los murciélagos son importantes componentes de los ecosistemas boscosos y se encuentran en todo el mundo. En consecuencia, a menudo interactúan con el fuego. Revisiones previas de los efectos del fuego sobre los murciélagos se han enfocado en los efectos de quemas prescriptas, en parte debido al número limitado de estudios sobre las respuestas de los murciélagos a los incendios. Sin embargo, hace ya varios años que estudios de las respuestas de los murciélagos a los incendios y a las quemas prescriptas se han incrementado considerablemente. Revisamos este cuerpo de literatura que se expande rápidamente, para determinar si los murciélagos responden en forma diferente a las quemas prescriptas y a los incendios, y la importancia de los factores que producen esas diferencias. También examinamos similitudes regionales y diferencias en la respuesta de los murciélagos a las quemas prescriptas e incendios, e identificamos áreas que necesitan más investigaciones.
Resultados
Nuestra revisión incluyó 52 estudios (29 quemas prescriptas, 23 incendios) de Norte y Sudamérica, Europa, Australia y África, aunque los estudios de Europa, Sudamérica y África fueron limitados. En general, encontramos que los murciélagos muestran respuestas positivas o neutras a las quemas prescriptas, mientras que una mayor proporción de respuestas negativas fueron reportadas para incendios. No obstante, algunas de las respuestas negativas a los incendios son de corta duración o locales, sugiriendo que los murciélagos pueden ser resilientes a los efectos del fuego. Factores como la severidad del fuego, la frecuencia del fuego, tiempo desde la última quema, extensión de la quema, temporada de la quema, y piro-diversidad, se encontraron como importantes conductores de las respuestas de los murciélagos tanto para quemas prescriptas como para incendios.
Conclusiones
La importancia de los aspectos espaciales y temporales del fuego sugieren que estos factores necesitan ser considerados en el diseño de futuros estudios y en la interpretación de los resultados. La piro-diversidad puede ser un concepto particularmente importante para entender mejor las respuestas de los murciélagos al fuego. Encontramos varios huecos en nuestro conocimiento, incluyendo falta de información de los efectos directos del fuego (por ej. mortalidad), desvíos taxonómicos y regionales, efectos de los incendios en los hábitats de descanso, y los efectos del cambio climático. A pesar de que los estudios actuales sugieren que el fuego puede ser una herramienta importante para mejorar el hábitat de los murciélagos, la amenaza de fuegos más frecuentes, extensos y severos puede agregar un estrés adicional en algunas poblaciones de murciélagos, particularmente aquéllas que están siendo impactadas por enfermedades, pérdida de hábitat y fragmentación, y cambio climático.
Similar content being viewed by others

Introduction
Bats are a highly diverse group of mammals, varying broadly in morphology, life history, ecology, and conservation status (Willig et al. 2003; Frick et al. 2020). They are found on every continent except Antarctica and occupy a diversity of habitats ranging from deserts to rainforests and may be found from the equator to the Arctic as well as in remote, urban, and agricultural areas. Despite the diversity of habitats which bats utilize, most bat species rely on forests to some extent as forests provide critical resources for foraging, roosting, and drinking water (Law et al. 2016). Because forests throughout the world often experience fire, both natural and human-ignited, bats often interact with fire.
Fire may have direct and indirect effects on bats (Perry 2012). Direct effects include immediate mortality as well as carbon monoxide toxicity and tissue damage such as ear burns (Dickinson et al. 2010). Fires may also result in the need to abandon roosts (Moorman et al. 1999; Dickinson et al. 2009; Jorge et al. 2021), making bats more vulnerable to predation. Indirect effects result from changes in the habitat and include the creation or loss of suitable roost sites, changes in foraging habitat structure, and changes in the insect prey base (Perry 2012). Indirect effects may be positive, neutral, or negative. For example, use or activity of some bat species increases after prescribed fire or wildfire, while that of other species declines or does not change (e.g., Inkster-Draper et al. 2013; Austin et al. 2018a, 2018b; Law et al. 2018b; Ancillotto et al. 2020).
Due to the lack of research on bat responses to fire prior to the early 2000s, Carter et al. (2002) reviewed some of the potential effects of prescribed fire on bats in the eastern USA and developed several hypotheses about how fire may affect their roosting and foraging behavior. Perry (2012) provided a more thorough review of the effects of fire on bats in the eastern oak region of the USA, but most of the papers included in his review examined the effects of prescribed fire on bats. More recently, Loeb (2020) synthesized the literature on effects of silvicultural treatments including regeneration harvests, thinning, gap formation, plantation forestry, and fire on bat activity and habitat use in temperate zone forests. Like Perry’s (2012) review, most of the fire studies included in that review pertained to prescribed fire. The reviews by both Perry and Loeb concluded that although responses of bats to fire vary among species, oftentimes in relation to their ecological and morphological characteristics, in general prescribed fire tends to have either neutral or positive effects on bats. Due to the low number of wildfire studies included in the previous reviews, it is not clear whether bats respond to wildfire as they do to prescribed fire. Wildfires are often larger in extent than prescribed fires (e.g., 100,000s ha versus 100s ha) and more severe, causing extensive damage to forests and human communities (Hunter and Robles 2020). Thus, their effects on bats may differ from those of prescribed fire. Further, neither review examined the effects of fire parameters such as burn severity, time since last burn, burn extent, fire frequency, or pyrodiversity on bats.
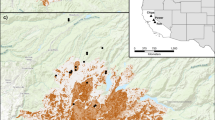
Wildfires have been increasing in frequency, severity, and extent throughout fire-prone regions of the world in recent decades, and there is broad scientific consensus that this is due to a warming and drying climate as well as other anthropogenic disturbances including fire suppression, increased human caused ignitions, removal of grazers, and the cessation of indigenous fire management (Jolly et al. 2015; Bowman et al. 2020). The use of prescribed fire has also been increasing, in part to reduce fuel loads to mitigate the effects of wildfires as well as to restore ecosystems. However, increasing periods of drought and monsoon may reduce the number of days suitable for prescribed fire, thereby leading to more intense wildfires (Chiodi et al. 2018). Megafires such as those that occurred in Australia in 2019–2020 and elsewhere in recent years can have devastating effects on wildlife (Ward et al. 2020). For example, Bosso et al. (2018) estimated that suitable habitat for the 12 bat species in Vesuvius National Park in southern Italy declined by approximately 40% for species that foraged in open areas but up to 60–80% for species that were dependent on interior forests after a severe wildfire burned roughly 40% (~ 3400 ha) of the park. Not only did the fire destroy habitat, but due to forest loss, it also increased fragmentation for species associated with closed forests.
Many bat species across the globe are endangered or threatened due to habitat loss and fragmentation, disease, wind energy development, urbanization, and climate change (Frick et al. 2020). Currently, 23% of the world’s bat species are considered to be globally threatened (listed as critically endangered, endangered, vulnerable, or near-threatened on the IUCN Red List) with 78% of species with either unknown or decreasing population trends (IUCN 2020). Thus, understanding the effects of fire on bats is critical for developing effective management guidelines and policies to prevent further endangerment and foster recovery. Over the past few years, several papers have been published on the response of bats to wildfires. Thus, we are better able to compare and contrast bats’ responses to prescribed fire and wildfire. Our objective was to review the literature on the responses of bats to prescribed fire and wildfire to address the following questions: (1) How do bats respond to the occurrence of prescribed fire and wildfire? (2) How do bats respond to various fire parameters including (a) burn extent (area burned); (b) fire frequency, fire return interval, and time since last burn defined as the number of fires in an area over a given time interval, the average number of years between fires, and months or years since the most recent fire, respectively (Sommers et al. 2011); (c) burn season (often described as growing season versus dormant season or wet versus dry season); (d) burn severity (a fire’s effect on the ecosystem, particularly the aboveground and belowground organic material) or intensity (energy released during a fire; Keeley 2009); and (e) pyrodiversity (spatial and temporal variability in post-fire landscape characteristics; Jones and Tingley In Press)? (3) Do bats respond differently to prescribed fire and wildfire? (4) What drives regional differences and similarities in bat response to prescribed fire and wildfire globally? (5) What are current information gaps and how can we best design research to address these gaps?
Review approach and available literature
We used the ISI Web of Science and included the Web of Science Core Collection, BIOSIS Citation Index, BIOSIS Previews, Current Contents Connect, and Zoological Record to search for papers on responses of bats to prescribed fire and wildfire through April 2021. Topic searches were [“bat OR chirop*” AND “fire”], [“bat OR chirop*” AND “burn”], [“bat OR chirop*” AND “wildfire”], and [“bat OR chirop*” AND “prescribed”]. We also examined the literature cited sections of these papers for additional references. For each paper included in the review, we determined the type of fire examined (prescribed fire or wildfire), the fire parameter(s) examined (e.g., burn versus no burn, time since last burn, severity), the response examined (foraging/commuting, roosting, physiology, demographic, or range distributions), and the response (positive, negative, or neutral) of each species or species group (either foraging guild or genus). Examples of positive responses included increased foraging or commuting activity or occupancy in burned compared to unburned sites, greater activity or occupancy in recently burned sites compared to sites burned at a greater time interval, and selection of roosting sites in or near burned areas compared to non-burned areas. Negative responses included lower activity or occupancy in burned areas compared to non-burned areas, lower activity or occupancy in more severely burned sites, and avoidance of burned sites for roosting. Although higher bat activity is often interpreted as a positive response to a treatment such as a fire, it is possible that high activity could be reflecting animals’ need to forage more in these areas to obtain the resources they need, and lower foraging rates could reflect high efficiency. Further, we recognize that other factors may also be contributing to responses of bats to fire such as logging history, other land uses, and invasive plants and have noted these where appropriate. We also determined the foraging guild (closed-space, edge-space, or open-space) of each species based on the literature. We defined closed-space foragers as species that forage primarily in interior forests; open-space foragers as those species that forage primarily in open habitats such as fields, forest gaps, or forests with relatively open canopies and little mid-story; and edge-space foragers as those species that can use both open and closed forests and often use the edges between (Aldridge and Rautenbach 1987; Fenton 1990). Open-space foragers are usually characterized by long, narrow wings and large body sizes, whereas closed-space foragers are usually small-bodied with short, broad wings which makes them more agile in closed spaces. Edge-space foragers tend to have intermediate characteristics. We limited our review to echolocating bats because of the strong relationships between echolocating bats and vegetation structure.
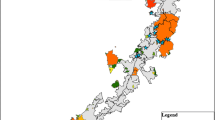
We found 49 published papers and three theses that addressed the effects of fire on bats, 29 studies on prescribed fire, and 23 studies on wildfire. We were not able to find any studies that addressed the effects of fire on bats prior to 2005, but the number of studies on both prescribed fire and wildfire has increased since then, particularly the number of wildfire studies in 2019, 2020, and early 2021 (Fig. 1A). Eleven studies were from Australia, one study was from Africa, two studies were from Europe, two studies were from Brazil, and 36 were from North America (one from Canada, the rest from the USA). No studies from western North America examined prescribed fire, whereas only one study from eastern North America examined wildfire (Fig. 1B and 2A). Four of the 29 prescribed fire studies and two of the 23 wildfire studies were Before-After-Control-Impact designs, while the remainder of the studies were either space-for-time (e.g., retrospective, post-treatment versus control experiments), or foraging or roost habitat selection studies conducted with radiotelemetry. Thirty-eight of the studies (23 prescribed fire, 15 wildfire) examined foraging responses of bats, and the majority of these (30) used acoustic detectors while five studies used radiotelemetry and three used mist-net captures. Eleven studies examined roost responses (including two that examined both foraging and roosting responses), only three of which examined roosting responses to wildfire. One study examined physiological responses of bats to wildfire, one study examined demographic responses to wildfire in addition to examining foraging responses, and two studies used species distribution models of database occurrences or range maps along with remotely sensed burn layers to make inferences about associations between bat distributions and pyrodiversity across Africa and southeastern Australia.

Temporal (A) and spatial (B; continental scale) distribution of studies that examined prescribed fire and wildfire on bats, from the first identified study in 2005 through April 2021. The data for 2020 includes five papers that were published in 2021 or are in press

A Global distribution of studies that examined relationships between bats and fire, and B inset of North American studies showing distribution of wildfire (triangle) and prescribed fire (star) studies. The underlying map shows fire potential, measured by the linear trend in the number of days annually that exceeded the 95th percentile of the Fire Weather Index over the years 1979–2019. These data were reproduced with permission from Bowman et al. (2020). Both panels are shown in the WGS84 coordinate reference system
Bat responses to burned versus unburned sites
Most bat species respond in either a positive or neutral way to prescribed fire (Additional file 1). However, some edge- and edge/open-space foraging bats in two studies in northern Australia (Inkster-Draper et al. 2013; Broken-Brow et al. 2020) show negative responses to prescribed fire although in one study, this only occurred after high-intensity fires (Additional file 1). Reduction in vegetation clutter or the creation of small gaps due to prescribed fire most likely contributes to the increase in bat use in response to prescribed fire compared to control sites (e.g., Smith and Gehrt 2010; Cox et al. 2016; Griffitts 2016; Silvis et al. 2016; Burns et al. 2019; Blanco and Garrie 2020; Smith et al. 2020). Foraging efficiency is higher in more open habitats, particularly for edge- and open-space foragers (Aldridge and Rautenbach 1987; Fenton 1990). However, in some studies, the prescribed fires may not have been intense enough to reduce clutter, resulting in neither positive nor negative responses (Austin et al. 2018a). An increase in the insect prey base after prescribed fire may also contribute to positive responses to prescribed fire in some areas (Lacki et al. 2009) but not in others (Cox et al. 2016). The relationship between insects and fire is complex and depends on the local insect community as well as many other factors including time of year that sampling occurs, time since the burn, and burn severity (Perry 2012).
Positive responses to prescribed fire are particularly evident in terms of roost responses. In six of the eight studies that examined roost responses of bats to prescribed fire, bats were more likely to select roosts in or near prescribed burn sites than unburned sites, and in the other two studies, there was a neutral response (e.g., Johnson et al. 2009, 2010; Thalken and Lacki 2018; see Additional file 1). For example, Myotis sodalis (Miller and Allen) in northern Arkansas select mature forest stands that had been burned at least once for roosting and avoid unburned forests (Perry et al. 2016). Large canopy gaps or low clutter resulting from prescribed fire may increase solar radiation on roost trees, resulting in lower energetic costs for thermoregulation (Boyles and Aubrey 2006).
Compared to prescribed fires, a greater number of bat species show negative responses to wildfire (Additional file 2). Some of the species that show negative responses are closed-space foragers (e.g., Nyctophilus spp., Rhinolophus ferrumequinum [Schreber], Plecotus austriacus [J. B. Fischer]) but some are edge-space (Vespadelus darlingtonia [G. M. Allen], V. vulturnus [O. Thomas], Hypsugo savii [Bonaparte]) or open-space foragers (Saccolaimus flaviventris [Peters]). However, even though the number of bat passes by R. ferrumequinum and P. austriacus significantly declined 1 year after a wildfire in Southern Italy, they either recovered to pre-burn conditions or showed positive responses 2 years after the fire compared to pre-fire activity (Ancillotto et al. 2020). Similarly, the proportion of reproductive female H. savii in this same study declined in the year following the fire but returned to pre-fire levels 2 years after the fire. This suggests that time since last burn is an important variable to consider in studies of responses of bats to fire as discussed below.
Two studies compared roosting behavior of bats in areas that experienced wildfire to non-burned areas. Myotis evotis (H. Allen), a closed-space forager, roosts farther from burned areas than randomly located roosts in southwestern Colorado (Snider et al. 2013). Insect abundance was also higher in unburned areas, suggesting that bats may have selected roosts in unburned areas to decrease foraging costs (Snider et al. 2013). In contrast, M. evotis and M. lucifugus (Le Conte) in Montana roost almost exclusively (91% of roost days) within mixed severity burn sites in Montana 1–2 years post-fire (Schwab 2006). In further contrast with the Snider et al. (2013) study, insect abundance was almost twice as high in burned areas compared to unburned areas. The differences between the two studies may have been due to the environmental conditions and insect communities experienced by bats in the two study areas. The southwestern Colorado study area was in the semi-arid Upper Sonoran Desert, and the authors speculated that bats selected unburned areas because they represented less harsh conditions (higher humidity, less variable and extreme temperatures) compared to burned areas. In contrast, the Montana study was conducted within two river drainages, and thus, the environmental conditions may have been less harsh and more favorable for bat roosting and foraging. Further, the responses of the insect communities to the wildfires may have varied between the two sites due to fire severity, seasonal timing, previous management, and the insect community (Swengel 2001).
Effects of fire parameters on bats—burn extent
Austin et al. (2018b, 2020) found no significant effects of the extent of either prescribed fires or wildfires on bat habitat use in the central Appalachian Mountains of Virginia (Additional files 3 and 4). Prescribed fires ranged from 1 to 8 years old but the areas covered by the prescribed burns were not reported. In contrast, the wildfires ranged from 3 to 85 years old and wildfire burn extents ranged from 20 to 9322 ha The authors suggested that the time since fire for many sites was so long that the structural changes to the forest were no longer biologically relevant (Austin et al. 2020). In contrast, Law et al. (2020) found a positive relationship between burn extent and activity of several edge- and open-space foragers for wildfire burns < 5 years old as well as burns that were 5–15 years old. Only one species, Chalinolobus picatus (Gould), an edge-space forager, showed a negative response to the extent of recent burns (< 5 years); however, this species did not have a negative response to the extent of older burns. Further, the presence of C. picatus at the landscape scale was relatively stable over the 5 years (Law et al. 2020) and its distribution across the Murray Mallee Region of southeastern Australia is not affected by burn extent (Senior et al. 2021), suggesting that the negative effects of recent wildfire extent are local for this species. However, the distributions of several other species of bats in the Murray Mallee Region are negatively associated with burn extent (Additional file 4). Blakey et al. (2021) related bat species richness in mixed-conifer forests of the Sierra Nevada, California, to extent of areas burned by wildfires in the 30 years prior to the study, measured as proportion of circular buffers with 1–10-km radii, and found that bat species richness was positively associated with burned area extent across all buffer sizes. They concluded that bat communities of the region were resilient to wildfire and potentially adapted to the heterogeneous forest structures that likely occurred prior to the era of fire suppression.
Effects of fire parameters on bats—fire frequency, time since last burn, and fire return interval
Temporal aspects of fire such as fire frequency, time since last burn, and fire return interval are important drivers of bat responses to both prescribed fire and wildfire (Fig. 3). Three studies examined the effects of prescribed fire frequency and two studies examined the effects of wildfire frequency on bat habitat use (Additional files 3 and 4). In southeastern Australia, the effect of prescribed fire frequency varies with logging history (Law et al. 2019). Bat activity in sites that have been previously logged, and thus represent thick regrowth, is higher where prescribed fires are more frequent (burned every 2 years compared to every 4 years). In contrast, in unlogged sites activity is higher in those sites burned every 4 years compared to every 2 years. Higher frequency burns in the logged sites are more effective at reducing stem density than routine burning (every 4 years). Nonetheless, the effect of fire frequency is small, and logging history and its effect on forest structure have a greater effect on bat activity and species richness than fire frequency (Law et al. 2019).

Examples of studies from various regions and fire types (wildfire and prescribed fire) that have directly examined the spatial (top) and temporal (bottom) components of the relationship between bats and fire
In Florida, the effects of prescribed fire frequency on Eumops floridanus (G. M. Allen) vary with season of burn and, consequently, fire intensity (Braun de Torrez et al. 2018b). Overall activity and foraging activity are higher in sites burned every 3–5 years (moderate frequencies) compared to sites burned at low frequencies (> 5 years) during the early wet season (April–June), but for sites burned during the cool, dry season (January–March), activity is higher in sites burned at low frequencies than at moderate frequencies. As discussed below, the most effective burn schedules for managing bat foraging habitat in an area may be those that best mimic the historical fire regime, which in south Florida is the early wet season based on burn seasons prior to European colonization (Braun de Torrez et al. 2018b). Starbuck et al. (2015) also found a positive relationship between evening bat (Nycticeius humeralis) (Rafinesque) habitat use and frequency of prescribed fires but found neutral effects for all other species analyzed (Additional file 3). Evening bats are often associated with savannas (grass, forb, and shrub dominated ecosystems with widely spaced trees) and woodlands (open forests with > 50% canopy but little mid-story and a ground layer dominated by forbs and woody species; Dey and Kabrick 2015), both of which can be maintained with frequent low-intensity fires (Greenberg and Collins In Press).
Responses to wildfire frequency appear to vary with species. Milne et al. (2005) found that a community of bats made up primarily of edge-space foragers in northern Australia was positively associated with wildfire frequency, whereas other bat communities were neither positively or negatively associated with fire frequency. In contrast, P. austriacus, a clutter-tolerant bat, avoids areas that have burned more than three times in the past 30 years, though these areas make up most of the landscape in northeast Iberia (López-Baucells et al. 2021).
Bats in the eastern USA respond in either a neutral way to time since last prescribed fire (Austin et al. 2018b) or are positively associated with more recent burns (Armitage and Ober 2012; Burns et al. 2019). For example, activity of open-space foragers and two edge-space foragers (Lasiurus borealis [Muller] and L. seminolus [Rhoads]) is higher in recently burned sites compared to sites burned > 8 years before sampling in Florida (Armitage and Ober 2012). Perimyotis subflavus (F. Cuvier), an edge-space forager, is also more likely to occupy more recently burned stands in the Cumberland Plateau (Burns et al. 2019). Because vegetation often regrows after burns and becomes dense after several years, bats may prefer more recently burned stands due to lower clutter in those stands.
Time since wildfire also appears to be an important factor in assessing wildfire effects on bats. For example, activity of two closed-space foragers in southern Italy decreased in burned areas the year after a wildfire but recovered to pre-burn levels 2 years after the fire (Ancillotto et al. 2020). Demographic responses of H. savii followed the same pattern. Similarly, activity of many species of Australian bats returns to pre-burn levels 4 years after a wildfire after showing significant decreases 1-year post-fire (Law et al. 2018b). In the Pantanal forest of Brazil, burned forests are used by sanguivorous, carnivorous, frugivorous, and gleaning insectivorous bats 3 months after a wildfire but primarily by frugivorous bats only 3 months later (6 months post-fire) (Santos et al. 2021). The authors suggested that reduction in vegetation clutter immediately after fire facilitates hunting and gleaning activities by predators for a short period, after which regeneration of fruiting plants provides resources for frugivorous bats.
While the above studies examined bats’ responses in a relatively short period of time after wildfires (3 months to 4 years), Blakey et al. (2019) examined bats’ responses to burns where time since last burn was 3–107 years. They found that in the montane forests of California, occupancy of M. lucifugus declined as time since last fire increased, while occupancy of Eptesicus fuscus (Palisot de Beauvois) increased up to about 30 years post-fire, leveled off at its highest point from about 30–60 years post-wildfire, and then decreased at about 70 years post-fire. Fire return interval had no effect on occupancy in the same study. However, other factors such as insect outbreaks, succession, and land use change could also have influenced occupancy across the landscape.
Nyctophilus geoffroyi (Leach), a closed-space forager in Australia, exhibits contrasting responses to time since last burn at the local (Doty et al. 2016) and range-wide scale within southeastern Australia (Senior et al. 2021). Torpor use is significantly lower 4 months after a wildfire compared to 2 years post-fire, which the authors attribute to an increase in vegetation on the forest floor over time. Nyctophilus geoffroyi hawks insects in mid-air but also captures prey from the forest floor. Thus, a reduction in ground vegetation may facilitate prey capture on the forest floor, thus reducing the need to use torpor to compensate for energy deficits. In the drier and more sparsely vegetated Mallee region, occurrence of N. geoffroyi is associated with areas that had not burned for up to 100 years at the landscape scale (100,000 km2) likely due to slow regeneration times of suitable roosts.
Several studies have also examined the short-term (hours to days) effects of fire on bat habitat use. Activity of E. floridanus increases significantly during the 24 days after prescribed fires in Florida compared to the 12 days before the fires and peaks 9 days after the fire (Braun de Torrez et al. 2018a). While structural changes to the habitat may have been responsible for this immediate effect, the authors speculated that bats were most likely attracted to burned areas because of an increase in prey availability or accessibility. In contrast, bat species richness and diversity increase gradually one night to 7 months after a wildfire in a gallery forest of the Brazilian Cerrado most likely in response to recovery of the forest canopy and a gradual increase in available resources (de Oliveira and Aguiar 2015). In tropical northern Australian forests, bat activity doubled at burned sites 2–39 days after prescribed fire, coupled with a community shift towards edge- and open-adapted species (Inkster-Draper et al. 2013). Immediate responses of roosting bats are of particular interest because bats are most vulnerable while they are in their roosts. In Kentucky, two M. septentrionalis (Trouessart) left their roosts within 10 min of a spring prescribed fire being ignited during the late afternoon approximately 20 m from their roosts (Dickinson et al. 2009). Similarly, one L. seminolus and one L. borealis flushed approximately 10 min after a prescribed burn was ignited near their roost (also during the late afternoon) in Florida (Jorge et al. 2021). Both the L. seminolus and L. borealis flew to roost trees in nearby mesic forests. In contrast, five other bats in the Florida study did not evacuate their roosts but presumably survived after ignition of a prescribed fire near their roosts, but these bats roosted in taller trees and the fires that were set near their roosts were much lower intensity with lower mean flame heights.
Only one study examined the effects of prescribed fire return interval. During winter, L. seminolus in northern Florida select day roosts in areas with longer mean fire return intervals particularly in mesic sites (Jorge et al. 2021). Since prescribed fires are usually conducted in winter in this area, the use of areas with longer fire return intervals may reduce the need to evacuate roosts during burning.
Effects of fire parameters on bats—burn season
Prescribed fires in the southeastern USA are usually conducted during the non-growing season, although growing season burns are becoming more common in the Coastal Plain (Knapp et al. 2009) and other regions such as the eastern broadleaf and Appalachian forests (Arthur et al. In Press). In the western USA, prescribed fires are often conducted during spring or early summer. Spring and fall are also typical times for prescribed fires in southern Australia (McCaw 2013), while prescribed burns are mostly conducted in the early or late dry season in northern Australia (Broken-Brow et al. 2020). Prescribed fire seasons often do not coincide with the historical fire season due to operational considerations as well as concerns about the direct and indirect effects on wildlife (Knapp et al. 2009). However, E. floridanus respond most favorably to prescribed fires conducted during the historical season which is April to June (early wet season) (Braun de Torrez et al. 2018b). In the Cumberland Plateau of Tennessee, foraging activity of several species is greater in sites that are burned in the spring compared to the fall but amount of thinning is also a factor (Cox et al. 2016). Seasonal differences in bat responses to fire are likely due to timing of fire in relation to bat and prey life cycles as well as intensity of fires in various seasons. For example, in northern Australia, early dry season fires are usually low severity, whereas late dry season fires are usually high severity (Russell-Smith and Edwards 2006; Broken-Brow et al. 2020).
Effects of fire parameters on bats—burn severity or intensity
Two studies examined the effects of prescribed fire severity on bats of the Cumberland Plateau of Kentucky and Tennessee with contrasting results. Lacki et al. (2017) found no effect of burn severity on bat activity in Mammoth Cave National Park, whereas occupancy of medium-severity burn sites by P. subflavus and Myotis spp. is significantly lower than occupancy of low-severity burn sites in Big South Fork National River and Recreation Area (Burns et al. 2019). Burns et al. (2019) suggested that medium-severity fires may reduce clutter below a threshold that is preferred by some closed-space foragers. Differences between the studies may be due to differences in measures of use (activity versus occupancy) or because Lacki et al. (2017) grouped all bats with echolocation frequencies > 34 kHz, thus obscuring species-specific responses. For example, Myotis spp. and P. subflavus, whose echolocation calls are ~ 40 kHz, would have been included in the > 34 kHz group. Because populations of Myotis spp. and P. subflavus have declined considerably in this area due to white-nose syndrome (Thalken et al. 2018), the > 34 kHz group was probably made up primarily of L. borealis and N. humeralis, both edge-space foragers who may be less likely to be negatively impacted by higher severity fires.
In tropical northern Australia, two prescribed burn intensities are commonly used by land managers: low-intensity burns, which are generally applied in the early dry season, and high-intensity burns, which are generally applied in the early wet season for weed control (Broken-Brow et al. 2020). In a BACI experiment, Broken-Brow et al. (2020) found that 2–4 weeks post-fire, edge/open-space foragers respond negatively to high-intensity prescribed fires but respond positively to low-intensity fires (Additional file 3). In contrast, open-space foragers respond positively to high-intensity fires but have neutral responses to low-intensity fires.
The effects of wildfire burn severity on bats in the western USA vary considerably across studies. Based on a trait analysis, Blakey et al. (2019) concluded that bats with traits consistent with clutter tolerance in mixed-conifer forests of California are associated with less frequent fires and less severe burns, whereas bats with traits adapted to open areas are positively associated with frequent, high-severity burns. However, in another study of the effects of wildfire severity on bats in mixed-conifer forests of California, two closed-space foraging species or groups (M. thysanodes [G. S. Miller] and Myotis with echolocation calls in the 40 kHz range) were positively associated with higher severity burns (Buchalski et al. 2013). Similarly, Steel et al. (2019) found positive associations between six bat species (closed-space, open-space, and edge-space foragers) and burn severity in mixed-conifer forests across four national forests in California. In southeastern Arizona, open-space foragers show a moderate positive response to burn severity, whereas closed-space foraging bats with high-frequency calls show a negative response to burn severity (Saunders 2015; Starbuck et al. 2020). However, Starbuck et al. (2020) found that landscape variables such as elevation and water density are better predictors of bat habitat use than burn severity. Saunders (2015) also examined roost selection in this landscape and found no effect of burn severity on selection of roosts. Variations in bat responses among these studies may be related to differences in times since fires or differences in landscape composition and environmental conditions. For example, in the arid southwest USA, water availability is particularly important and may override burn factors (Starbuck et al. 2020).
Effects of fire parameters on bats—pyrodiversity
In the same way that heterogeneous habitats provide a breadth of niches for species with different habitat needs, pyrodiversity is hypothesized to lead to higher biodiversity by providing a range of habitat structures and resources resulting from spatio-temporal variation in fire characteristics (Jones and Tingley In Press). Because bat responses to burned landscapes are variable among species based on ecological and morphological differences, it follows that bat diversity is likely to be positively associated with pyrodiversity in fire-prone regions (Blakey et al. 2019; Burns et al. 2019; Broken-Brow et al. 2020). This is supported by studies in Africa where bat species richness is positively associated with pyrodiversity in wet regions (Beale et al. 2018) and in the western USA where two studies in the Sierra Nevada found positive relationships between bat richness and pyrodiversity across scales from 250 m to 10 km (Steel et al. 2019; Blakey et al. 2021). However, when considered at the species level, bats have variable responses to pyrodiversity, with only three species showing a positive association with pyrodiversity in a western USA assemblage of 17 species (Steel et al. 2019) and 8 out of 9 species in semi-arid Australian woodlands being negatively associated with pyrodiversity (Senior et al. 2021). The overwhelmingly negative relationship between bat species and pyrodiversity in the Australian landscape (measured by diversity in post-fire age classes) is likely because bat roosting habitat is limited in this environment and takes > 40 years to regenerate after severe wildfires (Senior et al. 2021). In other landscapes such as the Sierra Nevada of California, pyrodiversity results in a heterogenous landscape consisting of stands of differing structure and suitable roosting, and foraging habitat is provided for a wide range of species with varying life histories and morphologies (Steel et al. 2019; Blakey et al. 2021).
Bat responses to fire—prescribed fire versus wildfire
Most studies included in this review indicated that prescribed fire had neutral or positive effects on bats, whereas a greater proportion of negative responses were reported for wildfire. However, where bat responses to wildfire were studied over time, they appeared to be largely resilient to the effects of wildfire, often recovering from any negative effects within a few years. Further, this review suggests that the characteristics of fires and their interactions with environmental factors may be more important in driving bat responses than the distinction between prescribed fire and wildfire.
The scales of the fires and the designs employed in studies that examined them relative to the home range of the bats also need to be considered when comparing prescribed fire and wildfire (Fig. 3). For example, many of the studies that have tested the effects of prescribed fire have been conducted at the stand scale (10–20 ha, Loeb and Waldrop 2008; Cox et al. 2016; Smith et al. 2020), whereas those examining wildfire are often conducted at the landscape scale (8000–> 463,000 ha, Jemison et al. 2012; Steel et al. 2019; Ancillotto et al. 2020; Austin et al. 2020; Law et al. 2020; Blakey et al. 2021). Although some bats have relatively restricted foraging ranges (~ 0.5 km from the roost, Waldien and Hayes 2001), others such as Brazilian free-tailed bats (Tadarida brasiliensis [I. Geoffrey Saint-Hilaire]) or spotted bats (Euderma maculatum [J. A. Allen]) can fly 30–56 km per night (Best and Geluso 2003; Chambers et al. 2011). The vagility of bats suggests that we should not make direct inferences or comparisons between the results of stand and landscape-scale studies. For example, while C. picatus activity is negatively related to the extent of recent burns at the stand scale, its presence across the landscape is relatively stable over time suggesting that the effects of fire are localized (Law et al. 2020). Nonetheless, at the community scale, relationships between bat richness and wildfire extent, configuration, and pyrodiversity are relatively consistent across scales from ~ 3–30,000 ha (Blakey et al. 2021).
It is evident from our review that bat responses to both prescribed fire and wildfire are complex and vary with a number of factors including burn extent, timing (season and time since fire), severity, and pyrodiversity and environmental factors such as forest type, structure, and health as well as topography, ecoregion, and climate, as well as the ecology of the bats themselves. Further, many of the fire parameters are not independent. For example, in a managed forest of the Sierra Nevada, California, burned area extent is highly correlated (|R| > 0.7) with mean burned patch area, edge density, and pyrodiversity across scales from ~ 300 to 30,000 ha (Blakey et al. 2021).
Understanding the spatial, temporal, and taxonomic dimensions of relationships between bats and fire is likely to help unravel the differences between prescribed and wildfire, as well as regional differences across environments, fire regimes, and bat communities. Studies that deal directly with spatial and or temporal aspects of the bat-fire relationship are valuable for making these links (Fig. 3). For example, at smaller spatial scales (~ 1 ha), bats are likely to respond to fine-scale features such as basal area (Blanco and Garrie 2020), but at larger spatial scales (80,000 ha), bats are more likely to respond to factors such as configuration of burned areas (Blakey et al. 2021). Furthermore, the scale at which bats respond most strongly to fire varies among species, likely due to varying levels of mobility. For example, closed-space bats with high-frequency calls are negatively associated with burn severity at fine (90-m radius) spatial scales, while edge- and open-space bats with low-frequency calls are positively associated with burn severity at coarser spatial scales (5.76-km radius) (Starbuck et al. 2020) (Fig. 3).
Knowledge gaps
Bat mortality from fire events
Although it has been suggested that bats may suffer direct mortality during fires resulting from being burned, smoke inhalation, heat exposure (Dickinson et al. 2010; O'Shea et al. 2016), or as a result of predation when evacuating roosts (Hovick et al. 2017), few data are available on the magnitude of this mortality. Based on laboratory experiments of Australian and U.S. bat species, bats may take > 20 min to respond to smoke and sound cues of fire when in torpor, with longer times to arousal at lower ambient temperatures (Scesny 2006; Layne 2009; Doty et al. 2018; Geiser et al. 2018). Nonetheless, bats in both Kentucky and Florida evacuated their roosts within approximately 10 min after prescribed fires were set nearby (Dickinson et al. 2009; Jorge et al. 2021). However, the temperatures under which these field experiments took place were warmer than the laboratory experiments. These few examples suggest that bats may experience significant mortality from high-intensity fast-moving fires if they are not able to arouse in time to escape. Because bats are most likely to be in torpor during morning (e.g., Klug and Barclay 2013), it has been suggested that prescribed fires be set later in the day when they are more likely to respond to audio and olfactory cues (Layne 2009; Doty et al. 2018). Because controlling the timing of wildfires is not possible, risks of mortality from wildfires may be greater than those from prescribed fire.
Regional variability in the relationships between bats and fire
Our literature search did not reveal any studies investigating relationships between bats and fire within Asia or the Indo-Pacific, and only two empirical studies in the Neotropics, two empirical studies in Europe, and one study in Africa (Fig. 2A). Africa and Asia contain vast areas that experience frequent fires (Archibald et al. 2013) and increasing fire seasons are predicted with global climate change (Jolly et al. 2015). Additionally, fire has been identified as a risk factor to the speciose bat communities of both regions (Suyanto and Struebig 2007; Edirisinghe et al. 2018; McCleery et al. 2018). The only study that has investigated the relationship between bats and fire in Africa was based on species’ range maps rather than direct observations (Beale et al. 2018). It is of considerable importance to global bat conservation to support local efforts to study relationships in these understudied regions, as they constitute a large proportion of global bat diversity.
The Neotropics contain the highest number of mammalian species in the world, including a highly rich bat fauna (Burgin et al. 2018). Fire is important in many neotropical ecosystems but is also an increasing threat in many regions due to increasing incidents of drought (Aragão et al. 2018). Further, 63% of the Neotropics consist of fire-sensitive ecosystems (defined as ecosystems where most of the organisms have not evolved in the presence of fire; Shlisky et al. 2007). Thus, understanding the response of bats to fire is critical to conservation of this highly diverse fauna. To date, only two studies have investigated the relationship between bats and fire in the Neotropics (de Oliveira and Aguiar 2015; Santos et al. 2021) with somewhat conflicting results. It is not clear why so few studies have examined the effects of fire on bats in the Neotropics as many studies have examined the effects of logging and deforestation on neotropical bats (Meyer et al. 2016). However, due to increasing droughts accompanying climate change, fires are likely to play a greater role in the ecology and conservation of neotropical bats and the need for more research on its effects is needed.
In the Mediterranean region of Europe, wildfires burn frequently and are a key driver of the rapid conversion of forest to shrubland (San-Miguel-Ayanz et al. 2012). Additionally, megafires are becoming more frequent in the region due to climate change and land use changes such as movement of people into fire-prone areas, introduction of fire-prone invasive species, and establishment of plantations of highly flammable tree species (Moreira et al. 2020). Model-based assessments in the region indicate that such fires can significantly reduce and fragment the habitat of entire bat communities (Bosso et al. 2018). The two recent empirical studies in the region have indicated that high-frequency fires can restrict movements of a clutter-tolerant bat species (López-Baucells et al. 2021) and high-severity wildfire can lead to reduced reproduction and lower body condition in the short term (Ancillotto et al. 2020).
In Australia, there are roughly equal numbers of studies investigating prescribed and wildfire (Fig. 1B). Australia is the most fire-prone of all continents (Murphy et al. 2019), and though certain ecosystems are adapted to fire, Australian wildfire events have been increasing in size and severity, most notably the 2019–2020 fire season which burned 5.8 million ha of temperate broadleaf forests (Boer et al. 2020). Australia also has a long history of prescribed burning, which has been practiced by Indigenous Australians for many generations to increase landscape productivity, to assist hunting, and for cultural reasons (e.g., Yibarbuk et al. 2001). Australian studies concerning bats and fire range from the temperate southeast to the southwest, which is characterized by a Mediterranean climate, to the tropical north. Studies across regions using prescribed fire mostly show positive responses by bats with mostly positive to neutral responses to wildfire. However, only one study examined the influence of fire on bat roosting habitat (Burgar et al. 2015), and more research is needed to understand the trade-offs between roost creation and destruction as a result of fire (Parnaby et al. 2010). No studies have examined the influence of fire on the two bat species of New Zealand, which, compared to Australia, has historically less frequent fires, a shorter history of fire management, and fewer fire-adapted plant communities (Perry et al. 2014; Baillie and Bayne 2019).
Although the greatest number of studies on bat responses to fire have been conducted in North America (Figs. 1B and 2A), there are large regional differences in our knowledge of bat responses to prescribed fire versus wildfire. For example, to date, no studies have been published on bat responses to prescribed fires in western North America, and although there are many published studies on responses of bats to prescribed fire in eastern North America, there is only one study of responses to wildfire. This dichotomy is in part due to the relative lack of prescribed fire being conducted in the western USA compared to the eastern USA (Kolden 2019). However, due to differences in climate, topography, forest types, and fire seasons, responses of bats to both prescribed fire and wildfire may vary across geographical and ecological regions. Prescribed fires are usually conducted during winter, spring, and fall in eastern North America and during spring and fall in western North America (Ryan et al. 2013). These time periods may or may not align with the historic fire season which vary by region. Further, wildfire seasons differ between the east and west. While winter is the season with the greatest probability of wildfire in eastern North America, the greatest risk of wildfire in western North America is during late spring, summer, and fall (Short 2017; Fig. 4). As bats, insects, and flora are at different stages of their life cycles when both prescribed fires and wildfire occur in the various geographical regions of North America, the responses of bats are likely to be different based on their phenology. Thus, predictions based on results of studies conducted in one region may not apply in other regions. Further, extrapolating from North America to other parts of the world could also be problematic.

Understudied bat taxa
The majority of research concerning bats and fire has involved two widespread families of bats, the Vespertilionidae and, to a lesser degree, the Molossidae. As noted above, very little research has been conducted in the Neotropics, and therefore, the highly diverse Phyllostomidae family has been understudied. Phyllostomid bats play key roles in forest regeneration due to the high diversity of seed dispersers (frugivores) and pollinators (nectarivores) in this group. Therefore, they are likely to contribute to post-fire vegetation recovery in the region (de Oliveira and Aguiar 2015). While our review focused on echolocating bats, Old World fruit bats are another understudied group of bats who are impacted by fire. Fire can trigger dispersal or fragmentation of fruit bat colonies in the short or long term via modification and destruction of colonial roosting sites (Jenkins et al. 2007; Baranowski et al. 2021). Furthermore, fire can greatly impact availability of seasonal nectar and fruit resources (Law et al. 2000). For example, during the 2019–2020 Australian bushfires, 33.7% of grey-headed flying fox (Pteropus poliocephalus [Temminck]) habitat area burned, including areas of critical winter foraging habitat (Baranowski et al. 2021).
Effects of fire on roosts and roosting habitat
Roosts are critical for bat survival and reproduction, and more than one-half of all bats use trees or other plant structures as roosts (Kunz and Lumsden 2003). Fire has the potential to significantly affect the availability and quality of roosts and roosting habitat and, consequently, bat populations. However, fewer than one-quarter of the studies we found examined the roosting responses of bats to fire, and only three examined the roosting responses of bats to wildfire or its severity (Schwab 2006; Snider et al. 2013; Saunders 2015). Snags are important roost structures for a number of bat species, and both wildfires and prescribed fires can affect the availability of snags (Horton and Mannan 1988; Stephens and Moghaddas 2005; Holden et al. 2006; Bagne et al. 2008; Parnaby et al. 2010). Little information is available on the effects of either prescribed fire or wildfire on characteristics of snags that determine their suitability as bat roosts such as the presence and amount of bark, snag height, and placement on the landscape. However, a recent study on the effects of prescribed fire frequency found that the density of snags suitable for bat roosting (defined as pine snags ≥ 15 cm diameter at breast height) is not affected by fire frequency (Baldwin 2019). Understanding bats’ roost use and selection after wildfire and how those habitats relate to foraging habitat is critical to fully understanding the short- and long-term effects of wildfire on bats.
The influence of climate change on relationships between bats and fire
Changing climates along with changes in land use history and management practices are significantly influencing the frequency, severity, and extent of wildfires across the globe (Westerling et al. 2006; Dennison et al. 2014; Jolly et al. 2015; Abram et al. 2021). Warming climates and increased probability of drought may also decrease the ability to conduct prescribed fire (Kupfer et al. 2020), further increasing the probability of wildfires (Fernandes and Botelho 2003; Kalies and Yocom Kent 2016). It is unclear how the interaction between increased fire frequency, fire severity, and fire extent will affect bats at local and landscape scales. For example, more than 23% of the temperate forest of southeast Australia (~ 6.5 million ha) burned during the 2019–2020 megafires (Abram et al. 2021), and fires of this severity and extent may have larger impacts on bats and their habitats, including mortality, than the wildfires reviewed here. Since frequent and high-severity fires favor bats with traits associated with open and edge habitats, current wildfire trends in fire-prone forests could lead to bat community shifts, with reductions in clutter-tolerant species (Blakey et al. 2019). Further, because wildfire seasons are beginning earlier and lasting longer, there is greater probability that fires will occur during bats’ reproductive period. Other effects of climate change such as drought may interact with increasing wildfire risk to exacerbate negative effects on bats (Piccioli Cappelli et al. 2021). Drought significantly impacts reproductive success of bats in arid regions (Adams 2010) and in many areas of the world, and increased fire potential (Fig. 2) may coincide with increasing drought (Dai 2012), putting further strain on bat populations.
A decrease in the number of suitable days for prescribed fires due to increasing temperatures and drought during prescribed fire seasons could have significant impacts on maintenance and restoration of habitat for some species. For example, the endangered Indiana bat (M. sodalis) relies on yellow pines (Pinus subgenus Diploxylon) as their primary roosts in the Southern Appalachian Mountains (Britzke et al. 2003; O'Keefe and Loeb 2017), which are dependent on fire for their persistence (Lafon et al. 2007). The inability to conduct prescribed fires could prevent restoration of critical habitat for this species. Because prescribed fire appears to improve foraging habitat for many species, decreasing the amount of prescribed fire may decrease the amount of suitable foraging habitat for some species. While thinning may be a suitable fire surrogate for improving bat foraging habitat structure (Humes et al. 1999; Blakey et al. 2016; Gonsalves et al. 2018a, 2018b), potential benefits to roosting structures from thinning are likely to take decades to develop and effects of thinning on bat roosting may be negative in the short term (Loeb 2020).
Conclusions and management implications
This review highlighted the complex nature of the relationships between bats and fire whether those fires are prescribed or wildland. Because bats appear to show positive or neutral responses to prescribed fire, prescribed fire may be an important management tool for bat habitat management as well as restoration of habitat for other species, control of invasive species, and reduction in the intensity and extent of wildfires. In contrast, negative responses are more prevalent in response to wildfire. However, some of these negative responses are short-lived perhaps due to bats’ vagility and factors such as fire timing, extent, and severity. Thus, we suggest that focusing on characteristics of fires (e.g., burn extent, severity, frequency) across landscapes over space and time may be more informative than focusing on the presence of fire and its type.
While some general patterns emerged from our review, many questions remain unanswered. This is in part due to the necessity of taking advantage of opportunities as they arise (e.g., wildfires), the lack of truly replicated studies, and the wide variation in ecosystems and study approaches that are represented in the studies included in this review. Further, most studies of bats are conducted over relatively short time scales, substituting space for time (Law and Blakey 2021). As several studies in this review pointed out, responses of bats to fire may change over time. Thus, future studies need to be conducted over long enough time periods to detect these changes. Many of the studies reviewed here relied on the use of acoustic detectors to measure responses of bats to fire. If the probability of detecting bats with detectors is related to treatments (e.g., burn versus no burn, burn severity), then the data may be biased (e.g., Burns et al. 2019). Other biases and caveats described by Loeb (2020) for studies on bats’ responses to silvicultural treatments such as the assumption that greater activity means better habitat and restriction of studies to the summer months apply to studies of bats’ responses to fire. Further, acoustic studies only measure habitat use. As with silvicultural treatments, we know little about the physiological and demographic responses of bats to either prescribed fire or wildfire (Loeb 2020). Demographic responses including population genetic effects are particularly important for determining population-level effects and understanding the long-term and large-scale impacts of fire on the status and management of bats (e.g., Brown et al. 2009). Thus, we suggest that future studies strive to address specific hypotheses about population-level responses of bats to fire as has been done for other silvicultural treatments (Law et al. 2018a).
Our review also highlights the importance of burn parameters in assessing fire’s impact on bats. Many of these parameters are interrelated and must be addressed in tandem. One way to do this is to further explore the concept of pyrodiversity and its utility for understanding bats’ response to fire at a range of spatial and temporal scales. While questions remain about how to manage for or even quantify pyrodiversity (Jones and Tingley In Press), a range of long-used (indigenous fire management) and emerging (pyrosilviculture) approaches may be used to restore and maintain pyrodiversity in landscapes where it is beneficial to bats and other biodiversity (Kelley et al. 2020; North et al. In Press). Taking an adaptative management approach to both prescribed fire and managed wildfire could provide an avenue to further our understanding of how the different properties and configurations of fire influence biodiversity, with attention to spatial and temporal scales (Driscoll et al. 2010; Van Wilgen et al. 2014; Kelley et al. 2020).
Availability of data and materials
No data were presented in this paper. All papers included in the review are listed in the Additional Files.
References
Abram, N.J., B.J. Henley, A.S. Gupta, T.J.R. Lippmann, H. Clarke, A.J. Dowdy, J.J. Sharples, R.H. Nolan, T. Zhang, M.J. Wooster, J.B. Wurtzel, K.J. Meissner, A.J. Pitman, A.M. Ukkola, B.P. Murphy, N.J. Tapper, and M.M. Boer. 2021. Connections of climate change and variability to large and extreme forest fires in southeast Australia. Communications Earth and Environment 2 (1): 8. https://doi.org/10.1038/s43247-020-00065-8.
Adams, R.A. 2010. Bat reproduction declines when conditions mimic climate change projections for western North America. Ecology 91 (8): 2437–2445. https://doi.org/10.1890/09-0091.1.
Aldridge, H.D.J.N., and I.L. Rautenbach. 1987. Morphology, echolocation and resource partitioning in insectivorous bats. Journal of Animal Ecology 56 (3): 763–778. https://doi.org/10.2307/4947.
Ancillotto, L., L. Bosso, P. Conti, and D. Russo. 2020. Resilient responses by bats to a severe wildfire: conservation implications. Animal Conservation https://doi.org/10.1111/acv.12653 24 (3): 470–481.
Aragão, L.E.O.C., L.O. Anderson, M.G. Fonseca, T.M. Rosan, L.B. Vedovato, F.H. Wagner, C.V.J. Silva, C.H.L. Silva Junior, E. Arai, A.P. Aguiar, J. Barlow, E. Berenguer, M.N. Deeter, L.G. Domingues, L. Gatti, M. Gloor, Y. Malhi, J.A. Marengo, J.B. Miller, O.L. Phillips, and S. Saatchi. 2018. 21st Century drought-related fires counteract the decline of Amazon deforestation carbon emissions. Nature Communications 9 (1): 1–12. https://doi.org/10.1038/s41467-017-02771-y.
Archibald, S., C.E.R. Lehmann, J.L. Gómez-Dans, and R.A. Bradstock. 2013. Defining pyromes and global syndromes of fire regimes. Proceedings National Academy of Science 110 (16): 6442–6447. https://doi.org/10.1073/pnas.1211466110.
Armitage, D.W., and H.K. Ober. 2012. The effects of prescribed fire on bat communities in the longleaf pine sandhills ecosystem. Journal of Mammalogy 93 (1): 102–114. https://doi.org/10.1644/11-MAMM-A-169.1.
Arthur, M.A., J.M. Varner, C.W. Lafon, H.D. Alexander, D.C. Dey, C.A. Harper, S.P. Horn, T.F. Hutchinson, T.L. Keyser, M.A. Lashley, C.E. Moorman, and C.J. Schweitzer. 2021. Fire ecology and management in eastern broadleaf and Appalchian forests. In Fire ecology and management across US forests: Past, present, and future, ed. C.H. Greenberg and B.S. Collins, 105–147. Switzerland: Springer.
Austin, L.V., A. Silvis, W.M. Ford, M. Mthersbaugh, and K.E. Powers. 2018a. Bat activity following restoration prescribed burning in the central Appalachian upland and riparian habitats. Natural Areas Journal 38 (2): 183–195. https://doi.org/10.3375/043.038.0208.
Austin, L.V., A. Silvis, W.M. Ford, and K.E. Powers. 2020. Effects of historic wildfire and prescribed fire on site occupancy of bats in Shenandoah National Park, Virginia, USA. Journal of Forest Research 31 (4): 1255–1270. https://doi.org/10.1007/s11676-019-00923-y.
Austin, L.V., A. Silvis, M.S. Muthersbaugh, K.E. Powers, and W.M. Ford. 2018b. Bat activity following repeated prescribed fire in the central Appalachians, USA. Fire Ecology 14: 10 https://doi.org/10.1186/s42408-018-0009-5.
Bagne, K.E., K.L. Purcell, and J.T. Rotenberry. 2008. Prescribed fire, snag population dynamics, and avian nest site selection. Forest Ecology and Management 255 (1): 99–105. https://doi.org/10.1016/j.foreco.2007.08.024.
Baillie, B.R., and K.M. Bayne. 2019. The historical use of fire as a land management tool in New Zealand and the challenges for its continued use. Landscape Ecology 34 (10): 2229–2244. https://doi.org/10.1007/s10980-019-00906-8.
Baldwin, M.E. 2019. Prescribed fire effects on snags in the southern Appalachian Mountains, 53. M. S. Thesis, University of West Georgia, Carollton, GA.
Baranowski, K., C.L. Faust, P. Eby, and N. Bharti. 2021. Quantifying the impacts of Australian bushfires on native forests and gray-headed flying foxes. Global Ecology and Conservation 27: e01566 https://doi.org/10.1016/j.gecco.2021.e01566.
Beale, C.M., C.J. Courtney Mustaphi, T.A. Morrison, S. Archibald, T.M. Anderson, A.P. Dobson, J.E. Donaldson, G.P. Hempson, J. Probert, and C.L. Parr. 2018. Pyrodiversity interacts with rainfall to increase bird and mammal richness in African savannas. Ecology Letters 21 (4): 557–567. https://doi.org/10.1111/ele.12921.
Best, T.L., and K.N. Geluso. 2003. Summer foraging range of Mexican free-tailed bats (Tadarida brasiliensis mexicana) from Carlsbad Cavern. New Mexico. Southwestern Naturalist 48 (4): 590–596. https://doi.org/10.1894/0038-49092003)048<0590:SFROMF>2.0.CO;2.
Blakey, R.V., B.S. Law, R.T. Kingsford, J. Stoklosa, P. Tap, and K. Williamson. 2016. Bat communities respond positively to large-scale thinning of forest regrowth. Journal of Applied Ecology 53 (6): 1694–1703. https://doi.org/10.1111/1365-2664.12691.
Blakey, R.V., E.B. Webb, D.C. Kesler, R.B. Siegel, D. Corcoran, J.S. Cole, and M. Johnson. 2021. Extent, configuration and diversity of burned and forested areas predict bat richness in a fire-maintained forest. Landscape Ecology 36 (4): 1101–1115. https://doi.org/10.1007/s10980-021-01204-y.
Blakey, R.V., E.B. Webb, D.C. Kesler, R.B. Siegel, D. Corcoran, and M. Johnson. 2019. Bats in a changing landscape: Linking occupancy and traits of a diverse montane bat community to fire regime. Ecology and Evolution 9 (9): 5324–5337. https://doi.org/10.1002/ece3.5121.
Blanco, C.M., and J. Garrie. 2020. Species specific effects of prescribed burns on bat occupancy in northwest Arkansas. Forest Ecology and Management 460: 117890 https://doi.org/10.1016/j.foreco.2020.117890.
Boer, M.M., V. Resco de Dios, and R.A. Bradstock. 2020. Unprecedented burn area of Australian mega forest fires. Nature Climate Change 10 (3): 171–172. https://doi.org/10.1038/s41558-020-0716-1.
Bosso, L., L. Ancillotto, S. Smeraldo, S. D'Arco, A. Migliozzi, P. Conti, and D. Russo. 2018. Loss of potential bat habitat following a severe wildfire: a model-based rapid assessment. International Journal of Wildland Fire 27 (11): 756–769. https://doi.org/10.1071/WF18072.
Bowman, D.M.J.S., C.A. Kolden, J.T. Abatzoglou, F.H. Johnston, G.R. van der Werf, and M. Flannigan. 2020. Vegetation fires in the Anthropocene. Nature Reviews Earth and Environmment 1 (10): 500–5115. https://doi.org/10.1038/s43017-020-0085-3.
Boyles, J.G., and D.P. Aubrey. 2006. Managing forests with prescribed fire: Implications for a cavity-dwelling bat species. Forest Ecology and Management 222 (1-3): 108–115. https://doi.org/10.1016/j.foreco.2005.09.024.
Braun de Torrez, E., H.K. Ober, and R.A. McCleery. 2018a. Activity of an endangered bat increases immediately following prescribed fire. Journal of Wildlife Management 82 (6): 1115–1123. https://doi.org/10.1002/jwmg.21481.
Braun de Torrez, E., H.K. Ober, and R.A. McCleery. 2018b. Restoring historical fire regimes increases activity of endangered bats. Fire Ecology 14 (2): 9. https://doi.org/10.1186/s42408-018-0006-8.
Britzke, E.R., M.J. Harvey, and S.C. Loeb. 2003. Indiana bat, Myotis sodalis, maternity roosts in the southern United States. Southeastern Naturalist 2: 235–242 https://doi.org/10.1656/1528-7092(2003)002[0235:IBMSMR]2.0.CO;2.
Broken-Brow, J., A.T. Hitch, K.N. Armstrong, and L.K.-P. Leung. 2020. Effect of fire on insectivorous bat activity in northern Australia: does fire intensity matter on a local scale? Australian Journal of Zoology 67 (6): 260–268. https://doi.org/10.1071/ZO20030.
Brown, S., M. Clarke, and R. Clarke. 2009. Fire is a key element in the landscape-scale habitat requirements and global population status of a threatened bird: The Mallee Emu-wren (Stipiturus mallee). Biological Conservation 142 (2): 432–445. https://doi.org/10.1016/j.biocon.2008.11.005.
Buchalski, M.R., J.B. Fontaine, P.A. Heady III, J.P. Hayes, and W.F. Frick. 2013. Bat response to differing fire severity in mixed-conifer forest California, USA. PLoS One 8 (3): e57884. https://doi.org/10.1371/journal.pone.0057884.
Burgar, J.M., M.D. Craig, and V.L. Stokes. 2015. The importance of mature forest as bat roosting habitat within a production landscape. Forest Ecology and Management 356: 112–123 https://doi.org/10.1016/j.foreco.2015.07.027.
Burgin, C.J., J.P. Colella, P.L. Kahn, and N.S. Upham. 2018. How many species of mammals are there? Journal of Mammalogy 99 (1): 1–14. https://doi.org/10.1093/jmammal/gyx147.
Burns, L.K.L., S.C. Loeb, and W.C. Bridges Jr. 2019. Effects of fire and its severity on occupancy of bats in mixed pine-oak forests. Forest Ecology and Management 446: 151–163 https://doi.org/10.1016/j.foreco.2019.05.024.
Carter, T.C., W.M. Ford, and M.A. Menzel. 2002. Fire and bats in the Southeast and mid-Atlantic: more questions than answers? In The Role of Fire in Nongame Wildlife Management and Community Restoration: Traditional Uses and New Directions, Proceedings of a Special Workshop, 139–143. US Department of Agriculture, Forest Service, Northeastern Research Station. General Technical Report NE-128.
Chambers, C.L., M.J. Herder, K. Yasuda, D.G. Mikesic, S.M. Dewhurst, W.M. Masters, and D. Vleck. 2011. Roosts and home ranges of spotted bats (Euderma maculatum) in northern Arizona. Canadian Journal of Zoology 89 (12): 1256–1267. https://doi.org/e10.1139/Z11-106.
Chiodi, A.M., N.S. Larkin, and J.M. Varner. 2018. An analysis of Southeastern US prescribed burn weather windows: seasonal variability and El Niño associations. International Journal of Wildland Fire 27 (3): 176–189. https://doi.org/10.1071/WF17132.
Cox, M.R., E.V. Willcox, P.D. Keyser, and A.L. Vander Yacht. 2016. Bat response to prescribed fire and overstory thinning in hardwood forest on the Cumberland Plateau, Tennessee. Forest Ecology and Management 359: 221–231 https://doi.org/10.1016/j.foreco.2015.09.048.
Dai, A. 2012. Increasing drought under global warming in observations and models. Nature Climate Change 3 (1): 52–58. https://doi.org/10.1038/NCLIMATE1633.
de Oliveira, H.F., and L.M.S. Aguiar. 2015. The response of bats (Mammalia: Chiroptera) to an incidental fire on a gallery forest at a Neotropical savanna. Biota Neotropica 15 (4): e–0091. https://doi.org/10.1590/1676-0611-BN-2015-0091.
Dennison, P.E., S.C. Brewe, J.D. Arnold, and M.A. Moritz. 2014. Large wildfire trends in the western United States, 1984–2011. Geophyscial Research Letters 41 (8): 2928–2933. https://doi.org/10.1002/2014GL059576.
Dey, D.C., and J.M. Kabrick. 2015. Restoration of midwestern oak woodlands and savannas. In Restoration of boreal and temperate forests, ed. J.A. Stanturf, 2nd ed., 401–428. Boca Raton: CRC Press https://doi.org/10.1201/b18809.
Dickinson, M.B., M.J. Lacki, and D.R. Cox. 2009. Fire and the endangered Indiana bat. In Fire in eastern oak forests, ed. T.F. Hutchinson, 51–75. Carbondale: Southern Illinois University.
Dickinson, M.B., J.C. Norris, A.S. Bova, R.L. Kremens, V. Young, and M.J. Lacki. 2010. Effects of wildland fire smoke on a tree-roosting bat: integrating a plume model, field measurements, and mammalian dose-response relationships. Canadian Journal of Forest Research 40 (11): 2187–2203. https://doi.org/10.1139/X10-148.
Doty, A.C., S.E. Currie, C. Stawski, and F. Geiser. 2018. Can bats sense smoke during deep torpor? Physiology and Behavior 185: 31–38 https://doi.org/10.1016/j.physbeh.2017.12.019.
Doty, A.C., C. Stawski, B.S. Law, and F. Geiser. 2016. Post-wildfire physiological ecology of an Australian microbat. Journal of Comparative Physiology B 186 (7): 937–946. https://doi.org/10.1007/s00360-016-1003-3.
Driscoll, D.A., D.B. Lindenmayer, A.F. Bennett, M. Bode, R.A. Bradstock, G.J. Cary, M.F. Clarke, N. Dexter, R. Fensham, G. Friend, M. Gill, S. James, G. Kay, D.A. Keith, C. MacGregor, J. Russell-Smith, D. Salt, J.E.M. Watson, R.J. Williams, and A. York. 2010. Fire management for biodiversity conservation: Key research questions and our capacity to answer them. Biological Conservation 143 (9): 1928–1939. https://doi.org/10.1016/j.biocon.2010.05.026.
Edirisinghe, G., T. Surasinghe, D. Gabadage, M. Botejue, K. Perera, M. Madawala, D. Weerakoon, and S. Karunarathna. 2018. Chiropteran diversity in the peripheral areas of the Maduru-Oya National Park in Sri Lanka: Insights for conservation and management. Zookeys 784 (784): 139–162. https://doi.org/10.3897/zookeys.784.25562.
Fenton, M.B. 1990. The foraging behaviour and ecology of animal-eating bats. Canadian Journal of Zoology 68 (3): 411–422. https://doi.org/10.1139/z90-061.
Fernandes, P.M., and H.S. Botelho. 2003. A review of prescribed burning effectiveness in fire hazard reduction. International Journal of Wildland Fire 12 (2): 117–128. https://doi.org/10.1071/WF02042.
Frick, W.F., T. Kingston, and J. Flanders. 2020. A review of the major threats and challenges to global bat conservation. Annals of New York Academy of Science 1469 (1): 5–25. https://doi.org/10.1111/nyas.14045.
Geiser, F., C. Stawski, A.C. Doty, C.E. Cooper, and J. Nowak. 2018. A burning question: what are the risks and benefits of mammalian torpor during and after fires? Conservation Physiology 6 (1): coy057 https://doi.org/10.1093/conphys/coy057.
Gonsalves, L., B. Law, and R. Blakey. 2018a. Experimental evaluation of the initial effects of large-scale thinning on structure and biodiversity of river red gum (Eucalyptus camaldulensis) forests. Wildlife Research 45 (5): 397–410. https://doi.org/10.1071/WR17168.
Gonsalves, L., B. Law, T. Brassil, C. Waters, I. Toole, and P. Tap. 2018b. Ecological outcomes for multiple taxa from silvicultural thinning of regrowth forest. Forest Ecology and Management 425: 177–188 https://doi.org/10.1016/j.foreco.2018.05.026.
Greenberg, C.H., and B.S. Collins. 2021. Fire ecology and management across US forests: Past, present, and future. 502. Switzerland: Springer.
Griffitts, R. 2016. Assessing the effects of prescribed fire on foraging bats at Mammoth Cave National Park after the arrival of white-nose syndrome. M.S. thesis, Richmond: University of Eastern Kentucky.
Holden, Z.A., P. Morgan, M.G. Rollins, and R.G. Wright. 2006. Ponderosa pine snag densities following multiple fires in the Gila Wilderness, New Mexico. Forest Ecology and Management 221 (1-3): 140–146 https://doi.org/10.1016/j.foreco.2005.09.014.
Horton, S.P., and R.W. Mannan. 1988. Effects of prescribed fire on snags and cavity-nesting birds in southeastern Arizona pine forests. Wildlife Society Bulletin 16: 37–44.
Hovick, T.J., D.A. McGranahan, R.D. Elmore, J.R. Weir, and S.D. Fuhlendorf. 2017. Pyric-carnivory: Raptor use of prescribed fires. Ecology and Evolution 7 (21): 9144–9150. https://doi.org/10.1002/ece3.3401.
Humes, M.L., J.P. Hayes, and M.W. Collopy. 1999. Bat activity in thinned, unthinned, and old-growth forests in western Oregon. Journal of Wildlife Management 63 (2): 553–561. https://doi.org/10.2307/3802642.
Hunter, M.E., and M.D. Robles. 2020. Tamm review: The effects of prescribed fire on wildfire regimes and impacts: A framework for comparison. Forest Ecology and Management 475: 118435 https://doi.org/10.1016/j.foreco.2020.118435.
Inkster-Draper, T.E., M. Sheaves, C.N. Johnson, and S.K.A. Robson. 2013. Prescribed fire in eucalypt woodlands: immediate effects on a microbat community of northern Australia. Wildlife Research 40 (1): 70–76. https://doi.org/10.1071/WR12133.
IUCN. 2020. The IUCN red list of threatened species. www.iucnredlist.org. Accessed 29 Dec 2020.
Jemison, M., L. Lumsden, J. Nelson, M. Scroggie, and R. Chick. 2012. Assessing the impact of the 2009 Kilmore East-Murrindindi Complex fire on microbats. Heidelberg: Department of Sustainability and Environment.
Jenkins, R.K.B., D. Andriafidison, H.J. Razafimanahaka, A. Rabearivelo, N. Razafindrakoto, Z. Ratsimandresy, R.H. Andrianandrasana, E. Razafimahatratra, and P.A. Racey. 2007. Not rare, but threatened: the endemic Madagascar flying fox Pteropus rufus in a fragmented landscape. Oryx 41 (2): 263–271 https://doi.org/10.1017/S0030605307001883.
Johnson, J.B., J.W. Edwards, W.M. Ford, and J.E. Gates. 2009. Roost tree selection by northern myotis (Myotis septentrionalis) maternity colonies following prescribed fire in a Central Appalachian Mountains hardwood forest. Forest Ecology and Management 258 (3): 233–242. https://doi.org/10.1016/j.foreco.2009.04.008.
Johnson, J.B., W.M. Ford, J.L. Rodrigue, J.W. Edwards, and C.M. Johnson. 2010. Roost selection by male Indiana myotis following forest fires in Central Appalachian Hardwoods forests. Journal of Fish and Wildlife Management 1 (2): 111–121. https://doi.org/10.3996/042010-JFWM-007.
Jolly, W.M., M.A. Cochrane, P.H. Freeborn, Z.A. Holden, T.J. Brown, G.J. Williamson, and D.M.J.S. Bowman. 2015. Climate-induced variations in global wildfire danger from 1979 to 2013. Nature Communications 6 (1): 7537. https://doi.org/10.1038/ncomms8537.
Jones, G.M., and M.W. Tingley. In Press. Pyrodiversity and biodiversity: A history, synthesis, and outlook. Diversity and Distributions. https://doi.org/10.1111/ddi.13280.
Jorge, M.H., W.M. Ford, S.E. Sweeten, S.R. Freeze, M.C. True, M.J.St. Germaine, H. Taylor, K.M. Gorman, E.P. Garrison, and M.J. Cherry. 2021. Winter roost selection of Lasiurine tree bats in a pyric landscape. PLoS One 16 (2): e0245695. https://doi.org/10.1371/journal.pone.0245695.
Kalies, E.L., and L.L. Yocom Kent. 2016. Tamm Review: Are fuel treatments effective at achieving ecological and social objectives? A systematic review. Forest Ecology and Management 375: 84–95 https://doi.org/10.1016/j.foreco.2016.05.021.
Keeley, J.E. 2009. Fire intensity, fire severity and burn severity: a brief review and suggested usage. International Journal of Wildland Fire 18 (1): 116–126. https://doi.org/10.1071/WF07049.
Kelley, L.T., K.M. Giljohann, A. Duane, N. Aquilué, S. Archibald, E. Batllori, A.F. Bennett, S.T. Buckland, Q. Canelles, M.F. Clarke, M.-J. Fortin, V. Hermoso, S. Herrando, R.E. Keane, F.K. Lake, M.A. McCarthy, A. Morán-Ordóñez, C.L. Parr, J.G. Pausas, T.D. Penman, A. Regos, L. Rumpff, J.L. Santos, A.L. Smith, A.D. Syphard, M.W. Tingley, and L. Brotons. 2020. Fire and biodiversity in the Anthropocene. Science 370: eabb0355 https://doi.org/10.1126/science.abb0355.
Klug, B.J., and R.M.R. Barclay. 2013. Thermoregulation during reproduction in the solitary, foliage-roosting hoary bat (Lasiurus cinereus). Journal of Mammalogy 94 (2): 477–487 https://doi.org/10.1644/12-MAMM-A-178.1.
Knapp, E.E., B.L. Estes, and C.N. Skinner. 2009. Ecological effects of prescribed fire season: A literature review and synthesis for managers, vol. PSW-GTR-224, 80. Albany: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station.
Kolden, C.A. 2019. We’re not doing enough prescribed fire in the Western United States to mitigate wildfire risk. Fire 2 (2): 30. https://doi.org/10.3390/fire2020030.
Kunz, T.H., and L.F. Lumsden. 2003. Ecology of cavity and foliage roosting bats. In Bat ecology, ed. T.H. Kunz and M.B. Fenton, 3–89. Chicago: The University of Chicago Press.
Kupfer, J.A., A.J. Terando, P. Gao, C. Teske, and J.K. Hiers. 2020. Climate change projected to reduce prescribed burning opportunities in the south-eastern United States. International Journal of Wildland Fire 29 (9): 764–778. https://doi.org/10.1071/WF19198.
Lacki, M.J., D.R. Cox, L.E. Dodd, and M.B. Dickinson. 2009. Response of northern bats (Myotis septentrionalis) to presribed fires in eastern Kentucky forests. Journal of Mammalogy 90 (5): 1165–1175. https://doi.org/10.1644/08-MAMM-A-349.1.
Lacki, M.J., L.E. Dodd, N.S. Skowronski, M.B. Dickinson, and L.K. Rieske. 2017. Relationships among burn severity, forest canopy structure and bat activity from spring burns in oak–hickory forests. International Journal of Wildland Fire 26 (11): 963–972. https://doi.org/10.1071/WF16159.
Lafon, C.W., J.D. Waldron, D.M. Cairns, M.D. Tchakerian, R.N. Coulson, and K.D. Klepzig. 2007. Modeling the effects of fire on the long-term dynamics and restoration of the yellow pine and oak forests in the southern Appalachian Mountains. Restoration Ecology 15 (3): 400–411. https://doi.org/10.1111/j.1526-100X.2007.00236.x.
Law, B., A. Doty, M. Chidel, and T. Brassil. 2018b. Bat activity before and after a severe wildfire in Pilliga forests: Resilience influenced by fire extent and landscape mobility? Austral Ecology 43 (6): 706–718. https://doi.org/10.1111/aec.12617.
Law, B., L. Gonsalves, A. McConville, and P. Tap. 2020. Landscape monitoring reveals initial trends in occupancy and activity of bats in multiple-use forests. Austral Ecology 46: 261–276 https://doi.org/10.1111/aec.12976.
Law, B., A. Kathuria, M. Chidel, and T. Brassil. 2019. Long-term effects of repeated fuel-reduction burning and logging on bats in south-eastern Australia. Austral Ecology 44 (6): 1013–1024. https://doi.org/10.1111/aec.12768.
Law, B., C. Mackowski, L. Schoer, and T. Tweedie. 2000. Flowering phenology of myrtaceous trees and their relation to climatic, environmental and disturbance variables in northern New South Wales. Austral Ecology 25 (2): 160–178. https://doi.org/10.1046/j.1442-9993.2000.01009.x.
Law, B., K.J. Park, and M.J. Lacki. 2016. Insectivorous bats and silviculture: balancing timber production and bat conservation. In Bats in the Anthropocene: conservation of bats in a changing world, ed. C.C. Voight and T.T. Kingston, 105–150. Heidleberg: Springer.
Law, B.S., and R.V. Blakey. 2021. Bats in temperate forests: Where are the trends in bat populations. In 50 Years of bat research: Foundations and new frontiers, ed. B.K. Lim, M.B. Fenton, R.M. Brigham, S. Mistry, A. Kurta, E.H. Gillam, A. Russell, and J. Ortega, 93–104. Cham: Springer. https://doi.org/10.1007/978-3-030-54727-1_6.
Law, B.S., M. Chidel, and P.R. Law. 2018a. Forest bat population dynamics over 14 years at a climate refuge: Effects of timber harvesting and weather extremes. PLoS One 13 (2): e0191471. https://doi.org/10.1371/journal.pone.0191471.
Layne, J.T. 2009. Eastern red bat (Lasiurus borealis) response to fire stimulus during torpor. Springfield: M.S. thesis, Missouri State University.
Loeb, S.C. 2020. Qualitative synthesis of temperate bat responses to silvicultural treatments—where do we go from here? Journal of Mammalogy 101 (6): 1513–1525. https://doi.org/10.1093/jmammal/gyaa089.
Loeb, S.C., and T.A. Waldrop. 2008. Bat activity in relation to fire and fire surrogate treatments in southern pine stands. Forest Ecology and Management 255 (8-9): 3185–3192. https://doi.org/10.1016/j.foreco.2007.10.060.
López-Baucells, A., C. Flaquer, M. Mas, P. Pons, and X. Puig-Montserrat. 2021. Recurring fires in Mediterranean habitats and their impact on bats. Biodiversity and Conservation 30 (2): 385–402. https://doi.org/10.1007/s10531-020-02095-2.
McCaw, W.L. 2013. Managing forest fuels using prescribed fire – A perspective from southern Australia. Forest Ecology and Management 294: 217–224 https://doi.org/10.1016/j.foreco.2012.09.012.
McCleery, R., A. Monadjem, B. Baiser, R. Fletcher, K. Vickers, and L. Kruger. 2018. Animal diversity declines with broad-scale homogenization of canopy cover in African savannas. Biological Conservation 226: 54–62 https://doi.org/10.1016/j.biocon.2018.07.020.
Meyer, C.F.J., M.J. Struebig, and M.R. Willig. 2016. Responses of tropical bats to habitat fragmentation, logging, and deforestation. In Bats in the Anthropocene: conservation of bats in a changing world, ed. C.C. Voight and T. Kingston, 63–103. Heidleberg: Springer https://doi.org/10.1007/978-3-319-25220-9.
Milne, D.J., M. Armstrong, A. Fisher, T. Flores, and C.R. Pavey. 2005. Structure and environmental relationships of insectivorous bat assemblages in tropical Australian savannas. Austral Ecology 30 (8): 906–919. https://doi.org/10.1111/j.1442-9993.2005.01535.x.
Moorman, C.E., K.R. Russell, M.A. Menzel, S.M. Lohr, J.E. Ellenberger, and D.H. Van Lear. 1999. Bats roosting in deciduous leaf litter. Bat Research News 40: 74–75.
Moreira, F., D. Ascoli, H. Safford, M.A. Adams, J.M. Moreno, J.M.C. Pereira, F.X. Catry, J. Armesto, W. Bond, M.E. González, T. Curt, N. Koutsias, L. McCaw, O. Price, J.G. Pausas, E. Rigolot, S. Stephens, C. Tavsanoglu, V.R. Vallejo, B.W. Van Wilgen, G. Xanthopoulos, and P.M. Fernandes. 2020. Wildfire management in Mediterranean-type regions: Paradigm change needed. Environmental Research Letters 15 (1): 011001. https://doi.org/10.1088/1748-9326/ab541e.
Murphy, B.P., L.D. Prior, M.A. Cochrane, G.J. Williamson, and D.M.J.S. Bowman. 2019. Biomass consumption by surface fires across Earth’s most fire prone continent. Global Change Biology 25 (1): 254–268. https://doi.org/10.1111/gcb.14460.
National Interagency Fire Center. 2021. National significant wildfire potential outlooks, https://www.predictiveservices.nifc.gov/outlooks/outlooks.htm. Accessed 6 Mar 2021.
North, M.P., R.A. York, B.M. Collins, M.D. Hurteau, G.M. Jones, E.E. Knapp, L. Kobziar, H. McCann, M.D. Meyer, S.L. Stephens, R.E. Tompkins, and C.L. Tubbesing. In Press. Pyrosilviculture needed for landscape resilience of dry western United States forests. Journal of Forestry https://doi.org/10.1093/jofore/fvab026.
O'Keefe, J.M., and S.C. Loeb. 2017. Indiana bats roost in ephemeral, fire-dependent pine snags in the southern Appalachian Mountains, USA. Forest Ecology and Management 391: 264–274 https://doi.org/10.1016/j.foreco.2017.01.036.
O'Shea, T.J., P.M. Cryan, D.T.S. Hayman, R.K. Plowright, and D.G. Streicker. 2016. Multiple mortality events in bats: a global review. Mammal Review 46 (3): 175–190. https://doi.org/10.1111/mam.12064.
Parnaby, H., D. Lunney, I. Shannon, and M. Fleming. 2010. Collapse rates of hollow-bearing trees following low intensity prescription burns in the Pilliga forests, New South Wales. Pacific Conservation Biology 16 (3): 209–220. https://doi.org/10.1071/PC100209.
Perry, G.L.W., J.M. Wilmshurst, and M.S. McGlone. 2014. Ecology and long-term history of fire in New Zealand. New Zealand Journal of Ecology 38: 157–176.
Perry, R.W. 2012. A review of fire effects on bats and bat habitats in the eastern oak region. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, vol GTR-NRS-P-102, 170–191. Springfield: Northern Research Station.
Perry, R.W., S.C. Brandebura, and T.S. Risch. 2016. Selection of tree roosts by male Indiana bats during the autumn swarm in the Ozark Highlands, USA. Wildlife Society Bulletin 40 (1): 78–87. https://doi.org/10.1002/wsb.624.
Piccioli Cappelli, M., R.V. Blakey, D. Taylor, J. Flanders, T. Badeen, S. Butts, W.F. Frick, and H. Rebelo. 2021. Limited refugia and high velocity range-shifts predicted for bat communities in drought-risk areas of the Northern Hemisphere. Global Ecology and Conservation 28: 301608 https://doi.org/10.1016/j.gecco.2021.e01608.
Russell-Smith, J., and A.C. Edwards. 2006. Seasonality and fire severity in savanna landscapes of monsoonal northern Australia. International Journal of Wildland Fire 15 (4): 541–550. https://doi.org/10.1071/WF05111.
Ryan, K.C., E.E. Knapp, and J.M. Varner. 2013. Prescribed fire in North American forests and woodlands: history, current practice, and challenges. Frontiers in Ecology and the Environment 11 (s1): e15–e24. https://doi.org/10.1890/120329.
San-Miguel-Ayanz, J., M. Rodrigues, S.S. de Oliveira, C.K. Pacheco, F. Moreira, B. Duguy, and A. Camia. 2012. Land cover change and fire regime in the European Mediterranean Region. In Post-fire management and restoration of Southern European Forests, ed. F. Moreira, M. Arianoutsou, P. Corona, and J. De Las Heras, 21–43. Switzerland: Springer. https://doi.org/10.1007/978-94-007-2208-8_2.
Santos, C.F., R.C. Teixeira, J. Raizer, and E. Fischer. 2021. Post-fire phyllostomid assemblages in forest patches of the Pantanal wetland. Mammalia 85 (2): 155–158. https://orcid.org/0000-0001-8722-9876. https://doi.org/10.1515/mammalia-2020-0037.
Saunders, E.H. 2015. Bat assemblage and selection of maternity roosts in a post-wildfire landscape. M.S. thesis, Flagstaff: Northern Arizona University.
Scesny, A.A. 2006. Detection of fire by eastern red bats (Lasiurus borealis): Arousal from torpor. M.S. thesis, Springfield: Missouri State University.
Schwab, N.A. 2006. Roost-site selection and potential prey sources after wildland firefor two insectivorous bat species (Myotis evotis and Myotis lucifugus) in mid-elevation forests of western Montana. Missoula: M.S. thesis, Arlington: University of Montana.
Senior, K.L., K.M. Giljohann, M.A. McCarthy, F.W. Rainsford, and L.T. Kelly. 2021. Predicting mammal responses to pyrodiversity: From microbats to macropods. Biological Conservation 256: 109031 https://doi.org/10.1016/j.biocon.2021.109031.
Shlisky, A., J. Waugh, P. Gonzalez, M. Gonzalez, M. Manta, H. Santoso, E. Alvarado, A. Ainuddin, D.A. Rodríguez-trejo, R. Swaty, D. Schmidt, M. Kaufmann, R. Myers, A. Alencar, F. Kearns, D. Johnson, J. Smith, and D. Zollner. 2007. Fire, ecosystems and people: threats and strategies for global biodiversity conservation, 17. The Nature Conservancy, Global Fire Initiative, Technical Report.
Short, K.C. 2017. Spatial wildfire occurrence data for the United States, 1992-2015 [FPA_FOD_20170508]. Fort Collins: USDA Forest Service Research Data Archive.
Silvis, A., S.D. Gehrt, and R.A. Williams. 2016. Effects of shelterwood harvest and prescribed fire in upland Appalachian hardwood forests on bat activity. Forest Ecology and Management 360: 205–212 https://doi.org/10.1016/j.foreco.2015.10.010.
Smith, C.K., A.J. Turner, J.K. Hiers, J. Garai, W.N. Wilson, and A.N. Nunley. 2020. The effects of oak (Quercus) restoration on forest trajectory and small mammal use in the southern Cumberland Plateau, USA. Fire Ecology 16 (1): 20. https://doi.org/10.1186/s42408-020-00080-2.
Smith, D.A., and S.D. Gehrt. 2010. Bat response to woodland restoration within urban forest fragments. Restoration Ecology 18 (6): 914–923. https://doi.org/10.1111/j.1526-100X.2009.00538.x.
Snider, E.A., P.M. Cryan, and K.R. Wilson. 2013. Roost selection by western long-eared myotis (Myotis evotis) in burned and unburned pinon–juniper woodlands of southwestern Colorado. Journal of Mammalogy 94 (3): 640–649. https://doi.org/10.1644/11-MAMM-A-153.1.
Sommers, W.T., S.G. Coloff, and S.G. Conard. 2011. Synthesis of knowledge: Fire history and climate change, 190. Boise: Joint Fire Science Program.
Starbuck, C.A., S.K. Amelon, and F.R. Thompson III. 2015. Relationships between bat cccupancy and habitat and landscape structure along a savanna, woodland, forest gradient in the Missouri Ozarks. Wildlife Society Bulletin 39 (1): 20–30. https://doi.org/10.1002/wsb.512.
Starbuck, C.A., E.S. Considine, and C.L. Chambers. 2020. Water and elevation are more important than burn severity in predicting bat activity at multiple scales in a post-wildfire landscape. PLoS One 15 (4): e0231170. https://doi.org/10.1371/journal.pone.0231170.
Steel, Z.L., B. Campos, W.F. Frick, R. Burnett, and H.D. Safford. 2019. The effects of wildfire severity and pyrodiversity on bat occupancy and diversity in fire-suppressed forests. Scientific Reports 9 (1): 16300. https://doi.org/10.1038/s41598-019-52875-2.
Stephens, S.L., and J.J. Moghaddas. 2005. Fuel treatment effects on snags and coarse woody debris in a Sierra Nevada mixed conifer forest. Forest Ecology and Management 214 (1-3): 53–64. https://doi.org/10.1016/j.foreco.2005.03.055.
Suyanto, A., and M.J. Struebig. 2007. Bats of the Sangkulirang limestone karst formations, East Kalimantan - A priority region for Bornean bat conservation. Acta Chiropterologica 9 (1): 67–95. https://doi.org/10.3161/1733-5329(2007)9[67:BOTSLK]2.0.CO;2.
Swengel, A.B. 2001. A literature review of insect responses to fire, compared to other conservation managements of open habitat. Biodiversity and Conservation 10 (7): 1141–1169. https://doi.org/10.1023/A:1016683807033.
Thalken, M.M., and M.J. Lacki. 2018. Tree roosts of northern long-eared bats following white-nose syndrome. Journal of Wildlife Management 82 (3): 629–638. https://doi.org/10.1002/jwmg.21411.
Thalken, M.M., M.J. Lacki, and J.S. Johnson. 2018. Shifts in assemblage of foraging fats at Mammoth Cave National Park following arrival of white-nose syndrome. Northeastern Naturalist 25 (2): 202–214. https://doi.org/10.1656/045.025.0203.
Van Wilgen, B.W., N. Govender, I.P.J. Smit, and S. MacFadyen. 2014. The ongoing development of a pragmatic and adaptive fire management policy in a large African savanna protected area. Journal of Environmental Management 132: 358–368 https://doi.org/10.1016/j.jenvman.2013.11.003.
Waldien, D.L., and J.P. Hayes. 2001. Activity areas of female long-eared myotis in coniferous forests in western Oregon. Northwest Science 75: 307–314.
Ward, M., A.I.T. Tulloch, J.Q. Radford, B.A. Williams, A.E. Reside, S.L. Macdonald, H.J. Mayfield, M. Maron, H.P. Possingham, S.J. Vine, J.L. O’Connor, E.J. Massingham, A.C. Greenville, J.C.Z. Woinarski, S.T. Garnett, M. Lintermans, B.C. Scheele, J. Carwardine, D.G. Nimmo, D.B. Lindenmayer, R.M. Kooyman, J.S. Simmonds, L.J. Sonter, and J.E.M. Watson. 2020. Impact of 2019–2020 mega-fires on Australian fauna habitat. Nature Ecology and Evolution 4 (10): 1321–1326. https://doi.org/10.1038/s41559-020-1251-1.
Westerling, A.L., H.G. Hidalgo, D.R. Cayan, and T.W. Swetnam. 2006. Warming and earlier spring increase Western U.S. forest wildfire activity. Science 313 (5789): 940–943. https://doi.org/10.1126/science.1128834.
Willig, M.R., B.D. Patterson, and R.D. Stevens. 2003. Patterns of range size, richness, and body size in the Chiroptera. In Bat ecology, ed. T.H. Kunz and M.B. Fenton, 580–621. Chicago: The University of Chicago Press.
Yibarbuk, D., Whitehead, P. J., Russell-Smith, J., Jackson, D., Godjuwa, C., Fisher, A., Cooke, P., Choquenot, D., Bowman D. M. J. S. 2001. Fire ecology and Aboriginal land management in central Arnhem Land northern Australia: a tradition of ecosystem management. Journal of Biogeography 28(3): 325-343 https://doi.org/10.1046/j.1365-2699.2001.00555.x.
Acknowledgements
This manuscript was conceptualized after discussions during the Wildfire and Prescribed Fire Effects on Wildlife in US Forests Symposium at the 2020 Wildlife Society Annual Conference. We thank David M. J. S. Bowman and T. Abatzolglou for providing the data and allowing reproduction of Fig. 2. and three anonymous reviewers for helpful comments on previous drafts. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Funding
S. C. Loeb was supported by the U.S. Forest Service, Southern Research Station and R.V. Blakey was supported by the La Kretz Center for California Conservation Science at the University of California, Los Angeles.
Author information
Authors and Affiliations
Contributions
The paper was conceived and framed by S.C. Loeb. Both authors conducted separate literature searches and combined results. S.C. Loeb and R.V. Blakey each wrote sections of the manuscript and jointly edited and approved it.
Author information
S. C. Loeb is a Research Ecologist. R. V. Blakey is a Postdoctoral Fellow.
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Responses bats to the presence of prescribed fire in Australian and North American forests. Responses were categorized as positive, neutral, or negative based on relative activity, occupancy, or selection of sites in burned areas. Guilds for individual species were assigned based on the literature (Closed- [C], Edge- [E], and Open-Space [O] foragers).
Additional file 2.
Responses of bats to the presence of wildfire in Australian, European, and North American forests. Responses were categorized as positive, neutral, or negative based on relative activity, occupancy, or selection of sites in burned areas. Guilds for individual species were assigned based on the literature (Closed- [C], Edge- [E], and Open-Space [O] foragers).
Additional file 3.
Responses of bats to various prescribed fire parameters in Australian and North American forests. Responses were categorized as positive, neutral, or negative based on relative activity, occupancy, or selection of sites in burned areas. Guilds for individual species were assigned based on the literature. Guilds for individual species were assigned based on the literature (Closed- [C], Edge- [E], and Open-Space [O] foragers).
Additional file 4.
Responses of bats to various wildfire parameters in Australian, Africa, Brazilian, European, and North American forests. Responses were categorized as positive, neutral, or negative based on relative activity, occupancy, or selection of sites in burned areas. Guilds for individual species were assigned based on the literature. Guilds for individual species were assigned based on the literature (Closed- [C], Edge- [E], and Open-Space [O] foragers).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Loeb, S.C., Blakey, R.V. Bats and fire: a global review. fire ecol 17, 29 (2021). https://doi.org/10.1186/s42408-021-00109-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-021-00109-0