
Abstract
Background
Spodoptera litura (Lepidoptera: Noctuidae), commonly known as tobacco cutworm or cotton leafworm, is a polyphagous pest which causes considerable damage to cotton (Gossypium hirsutum) and other crops. Herbivore-induced defence response is activated in plants against chewing pests, in which plant secondary metabolites play an important role. Dhurrinase2 (SbDhr2), a cyanogenic β-glucosidase from Sorghum bicolor, is the key enzyme responsible for the hydrolysis of dhurrin (cyanogenic β-glucosidic substrate) to p-hydroxymandelonitrile. Hydroxynitrile lyase (MeHNL) from Mannihot esculanta catalyses the dissociation of cyanohydrins to hydrogen cyanide and corresponding carbonyl compound, both enzymes play a pivotal role in plant defence mechanism.
Results
SbDhr2 and MeHNL genes were expressed individually and co-expressed transiently in cotton leaves. We examined the feeding response of S. litura to leaves in the choice assay. The S. litura population used in this study showed better feeding deterrence to leaves co-expressing both genes compared with the expression of an individual gene.
Conclusion
Our results suggest that co-expression of SbDhr2 and MeHNL genes in cotton leaves demonstrate feeding deterrence to S. litura. Engineering cyanogenic pathway in aerial parts of cotton would be an additional defence strategy against generalist pests and can be enhanced against specialist pests.
Similar content being viewed by others


Background
Spodoptera litura (Lepidoptera: Noctuidae) commonly known as tobacco cutworm or cotton (Cheng et al. 2017) leafworm is a polyphagous pest, causes considerable damage to cotton (Gossypium hirsutum) and various other crops (Xue et al. 2010; Bragard et al. 2019). Failure to control S. litura and its resistance to various insecticides, lead to humongous economical loss (Ahmad and Gull 2017; Fand et al. 2015). Plant secondary metabolites, play a direct role in plant defence response (War et al. 2012; Schaller 2008) and in the adaptation of plants to abiotic/biotic stresses (Akula and Ravishankar 2011; Bartwal et al. 2013; Gleadow et al. 1998; Rosenthal and Berenbaum 1992). Engineering plant metabolic pathways would be a feasible alternative defence strategy against generalist insect pest. In two-component defence system, β-glucosidases and cyanogenic glucosides are separated by different subcellular compartments (Saunders and Conn 1978; Thayer and Conn 1981; Kesselmeier and Urban 1983; Poulton and Li 1994). In plant physiology, β-glucosidases play diverse roles (Morant et al. 2008), and more than 2 500 species of plants contain Cyanogenic glucoside (Panter 2018). Metabolic engineering of the whole cyanogenic pathway in different plants has been reported for insect herbivory deterrence (Franks et al. 2006; Blomstedt et al. 2016; Tattersall et al. 2001; Bak et al. 2000).
Transient gene expression in cotton using virus-induced gene silencing (VIGS) vector (Becker 2013) (TRV: Tobacco Rattle Virus) has already been performed (Li et al. 2018; Gao et al. 2011; Pang et al. 2013) for functional genomic studies. Expressing cyanogenic pathway enzymes in upland cotton can help to develop insect-pest resistant cotton varieties.
SbDhr2 (dhurrinase2/AF253508.1) from Sorghun bicolor and MeHNL (α-hydroxynitrile lyase/AY787210.1) from Mannihot esculanta are two genes of cyanogenic pathway. SbDhr2 is a monomeric unit (62 kDa) expressed in nodes and leaves. Due to broader substrate specificity SbDhr2 hydrolyses certain artificial substrates (4-Methylumbelliferyl β-D-galactopyranoside, 4-Nitrophenyl-β-D-glucopyranoside, triglochinin) in addition to its natural substrates, dhurrin and sambunigrin (Cicek and Esen 1998; Verdoucq et al. 2004). MeHNL monomer (29.3 kDa) is the key enzyme responsible for the release of hydrogen cyanide (HCN) from α -Hydroxynitrile glucosides (Dadashipour and Asano 2011), having broad substrate specificity (Chueskul and Chulavatnatol 1996; Yan et al. 2003; Cheng et al. 2001; Wajant and Pfizenmaier 1996; Lauble et al. 2002). This paper investigates feeding deterrence and settling preference choice assay with S. litura on cotton leaves, transiently expressing SbDhr2 and MeHNL genes, the former driven by phosphoenolpyruvate carboxylase (PEPC) and the latter by 2X CaMV 35S (duplicated CaMV 35S) promoter (Pauli et al. 2004).
Methods
Plant materials
Seeds of cotton (G. hirsutum) were sown in pots containing peat moss and kept at 23 °C, 200 μmol. m-2. S− 1 light, 65% relative humidity with 16 h/8 h day-night photoperiod in a growth room. After the emergence of four to five true leaves, cotyledons were used for infiltration.
Plasmid construction
SbDhr2 (Verdoucq et al. 2003) gene driven by PEPC promoter (S. vulgare Accession. No X63756.1) was digested with EcoRI and KpnI and ligated in TRV2 pYL156 (pTRV-RNA2) to get the recombinant plasmid pTD2 (Fig. 1a). MeHNL gene digested with EcoRI and SacI was ligated in TRV2 to get the second recombinant plasmid pTH2 (Fig. 1b). Verification of clones was done by restriction enzyme digestion.

Schematic representation of plasmid vectors maps employed in this study, a pTD2 (pYL156 (pTRV-RNA2) + SbDhr2) here dhurrinase2 gene is under control of PEPC promoter, b pTH2 (pYL156 (pTRV-RNA2) + MeHNL), α-Hydroxynitrile lyase gene is under control of 2X CaMV 35S promoter
Agrobacterium tumefaciens LBA4404
TRV1, TRV2, pTH2, pTD2, independent clones were transformed in A. tumefaciens strain LBA4404 on LB agar plate containing kanamycin (50 mg·L− 1), rifampicin (125 mg·L− 1) and streptomycin (50 mg·L− 1) using the freeze-thaw transformation method (Weigel and Glazebrook 2006). After growing at 28 °C for 2 days, polymerase chain reaction (PCR) was used to select positive transformants that would be used further in this study.
Agroinfiltration in cotton cotyledons
Five hundred microliter inoculum of freshly activated single colonies of A. tumefaciens carrying each binary vector of TRV1,TRV2, pTD2, pTH2 were transferred to flasks containing 50 mL of LB medium supplemented with kanamycin (50 mg·L− 1), rifampicin (125 mg·L− 1) and streptomycin (50 mg·L− 1), 10 mmol·L− 1 2-(4 morpholino)-ethane sulfonic acid (MES), 20 μmol·L− 1 acetosyringone (Gao et al. 2011; Pang et al. 2013), then were grown overnight at 28 °C, 160 r·min− 1. Cells were pelleted at 4 000 r·min− 1 for 5 min and resuspended in an infiltration buffer containing 10 mmol·L− 1 MgCl2, 10 mmol·L− 1 MES and 200 μmol·L− 1 acetosyringone (Pang et al. 2013). At 600 nm, O.D. value of the culture was adjusted to 0.9 and suspension was kept for shaking 3 ~ 4 h at 25 °C under 10 r·min− 1. Agrobacterial culture suspensions were prepared in the ratio of 1:1(Table 1). Then infiltrations were performed in 20 plants for each suspension, at the abaxial side of cotyledons with a needleless syringe (Gao et al. 2011; Senthil-Kumar and Mysore 2014) (Fig. 2).

Sites of Agrobacterium suspension infiltration in cotyledons underside
Gene detection in non-infilterated leaves using PCR
DNA was isolated from non-infiltrated leaves by the cetyl trimethyl ammonium bromide (CTAB) method (Healey et al. 2014). Then PCR analysis was performed using primers listed in Table 2 for SbDhr2 and MeHNL genes. PCR positive leaf samples were further analysed for protein expression.
Western blotting for confirmation of gene expression
Total protein was extracted from sorghum, cassava, control plants and PCR positive non-infiltrated cotton leaves (Fig. 3). For Western blotting (Trans Blot Turbo transfer system) 40 μg of total protein was transferred on Amersham Hybond- P 0.45 PVDF blotting membrane, as per manufacturer’s instructions. Amersham Hybond-P 0.45 PVDF, a 0.45 μm pore size polyvinylidene difluoride (PVDF) hydrophobic membrane, is used with standard colorimetric and chemiluminescent detection methods for proteins. Blots were probed with primary polyclonal antibodies raised in rabbit for SbDhr2 and MeHNL proteins, detected with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody, and chemiluminescence was performed using Pierce™ ECL Western blotting substrate as per manufacturer’s instruction. Membranes were exposed to X-ray film, then developed and fixed.

Representative SDS-PAGE (mass fraction 10%) analysis of total protein, a lane 1- S. bicolor, M-marker, lane 2- cotton, lane 3-TRV1: TRV2 infiltrated cotton, lanes 4 ~ 7 cotton samples positive for both pTD2 & pTH2 plasmids. b Lane 1- M. esculenta as positive, lane 2- cotton, lane 3- TRV1: TRV2 infiltrated cotton, lane 4- cotton samples positive for both pTD2 & pTH2 plasmids
S. litura herbivory deterrence assay
Three settling preference choice tests (Krothapalli et al. 2013) were conducted in petri-plates containing a control leaf and a leaf transiently singly expressing and co-expressing SbDhr2 and MeHNL genes on wet germination paper (Table 3). Plates were kept at room temperature at a relative humidity of 68% and 16 h/8 h day-night photoperiod. Five of the third instar larvae of S. litura were released in the centre of each plate after being starved for 3 ~ 4 h, and the settling preference of insects was measured after every 24 h period till the 8th day. Each choice assay was replicated four times. The number of insects on each leaf was used to measure the settling preference and t-test was performed with GraphPad prism-8 for insect count. Mean weight of the five larvae before feeding and post feeding for 2 days (48 h) and on the 8th day was recorded in all three set of tests.
Cyanogenic capacity (HCNc) in infiltrated leaves
Amount of hydrogen cyanide released per unit time is measured as Cyanogenic Capacity (HCNc) (Hay-Roe et al. 2011). In this experiment, we were interested in temporal comparisons of HCNc in three different experimental conditions. Leaves positive for Western blotting were crushed in 1.5 mL tubes for the qualitative test and Feigl-Anger cyanide test paper (Feigl and Anger 1966) was fixed inside the top portion of caps. Then changing in colour of test paper after every hour was monitored till 10 ~ 12 h ranked according to Hay-Roe et al. (2011).
Results
Validation of plasmid for transient expression
After verification of plasmids, pTH2 and pTD2 by restriction enzyme digestion, a 790 base pairs (bp) MeHNL gene fragment was released from pTH2 vector. A 2.3 kilobase (kb) SbDhr2 gene along with its PEPC promoter gene was released as a restriction digestion product from linearized pTD2 vector (Fig. 4). Colony PCR screening of A. tumefaciens (LBA4404) using gene specific primers confirmed the presence of pTH2 and pTD2 vectors (Fig. 5). Colony PCR gave amplification product of 770 bp for MeHNL gene (Fig. 5a) and a 2.3 kb amplicon of SbDhr2 gene along with its promoter (Fig. 5b).

Agarose gel electrophoresis of restriction enzyme digestion of pTH2 by EcoRI and SacI, lane 1&3 are uncut pTH2 plasmids, lanes 2 & 4 pTH2 are digested, lanes 5 & 7 are uncut pTD2, lanes 6 & 8 pTD2 are digested with EcoRI and KpnI, M is a 2 000 bp DNA marker

Colony PCR for confirmation of genes in A. tumefaciens strain LBA4404, a lanes 1~10 pTH2 positive A. tumefaciens colonies, b lanes 1~10 pTD2 positive A. tumefaciens colonies, M: 2 000 bp marker, (+): positive control, B:blank
Screening of genes and expression in non-infiltrated cotton leaves
PCR analysis confirmed the presence of MeHNL (Fig. 6a) and SbDhr2 genes (Fig. 6b). Fifteen leaf samples were screened for presence of MeHNL and SbDhr2 genes, of which 13 samples were PCR positive for each gene. PCR results of negative control leaves confirmed the absence of either gene. Western blot analysis of total protein from five randomly selected PCR positive leaves confirmed the presence of MeHNL (29.3 kDa) (Fig. 7a) and SbDhr2 (62 kDa) (Fig. 7b) proteins bands. Un-infiltrated and empty vector infiltrated leaves of cotton served as negative control.

PCR analysis of transgene in non-agroinfiltrated leaf samples, DNA isolated from non-infiltrated true leaves. a Lane 1: cotton plant as negative control, lane 2: negative control (TRV1: TRV2 infiltrated), lanes 3 ~ 9: plants infiltrated only with pTH2, lanes 10 ~ 17: plants infiltrated with both pTH2 + pTD2 binary vector, M: 2000 bp marker, H+: positive control for MeHNL gene. b Lane 1: cotton plant as negative control, lane 2: negative control (TRV1: TRV2 infiltrated), lanes 3 ~ 9: plants infiltrated only with pTD2, lanes 10 ~ 17: plants infiltrated with both pTH2 + pTD2 binary vector, D+: positive control for SbDhr2, M: marker (Hind III λ-DNA marker mixed with 2000 bp marker)

Western Blot analysis of transiently expressed proteins. a Western blotting with polyclonal antibody against MeHNL protein, lane 1: total protein from M. esculanta as positive control, lane 2: cotton as negative control, lane 3: TRV1: TRV2 infiltrated cotton as negative control, lanes 4 ~ 7: cotton samples as positive (co-expressing MeHNL & SbDhr2 proteins) samples. b Western blotting with polyclonal antibody against SbDhr2 protein, lane 1: total protein from S. bicolor as positive control, lane 2: cotton as negative control, lane 3: TRV1 + TRV2 infiltrated cotton as negative control, lanes 4 ~ 7: cotton samples as positive (co-expressing MeHNL & SbDhr2 proteins) samples. Bands of our interest are highlighted; M: protein marker
Insect herbivory measurements
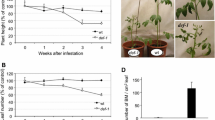
A two-tailed t-test (GraphPad prism-8) for the insect number on a leaf after 48 h indicated that a greater number of pests preferred settling on control leaf (Fig. 8a, c, e) as per observation. On the 8th day, settling preferences were recorded with no significant difference (P > 0.05) in plants singly expressing MeHNL and SbDhr2 genes with respect to the control leaves (Fig. 8b, d). Significant difference (P < 0.05) (Fig. 8f) was recorded in settling preference on the 8th day, where MeHNL and SbDhr2 genes were co-expressed compared with the control leaf. Settling preference results substantiate visual observation of damage caused by pests on control and independently expressed MeHNL and SbDhr2 proteins in leaves and was nearly homogeneous on the 8th day (Fig. 9a ~b) with no significant difference, whereas leaf samples co-expressing both SbDhr2 and MeHNL proteins demonstrated a better feeding deterrence till the 8th day (Fig. 9c ~d) as observed. The damage caused in co-expressed infiltrated leaf is far less compared with control and independently expressing proteins. Mean weight of five S. litura larvae before feeding was 0.020 mg, and the mean weight gained by larvae post feeding on all three sets of tests was recorded after 2 days (48 h) and on the 8th day (Table 4). There was no significant difference observed in weight gained by larvae (Fig. 10).

Insect (S. litura) settling preference on the control (TRV1: TRV2 infiltrated) leaf and infiltrated leaf, four replicates of each experiment with 5 insects in each plate were performed, to calculate two-way t-test for P –value. a there is a significant difference (P < 0.01) in 48 h, b no significant difference in settling preference (P > 0.14) of the 8th day, c there is a significant difference (P < 0.029) in 48 h, d no significant difference in settling preference (P > 0.099) of the 8th day, e there is a significant difference (P < 0.001) in 48 h, f there is a significant difference in settling preference (P < 0.001) on the 8th day

Damage caused till the 8th day of feeding, Con: control (TRV1: TRV2), a pTH2 infiltrated, b pTD2 infiltrated, c-i pTH2 + pTD2 co-infiltrated (top view), c-ii pTH2 + pTD2 co-infiltrated (underside view)

Mean weight of five larvae of S. litura (third instar) fed on three different feeding preference tests and weights recorded post feeding after 2 days and on the 8th day
Cyanogenic capacity (HCNc) in leaves
Cyanide released from transiently expressed positive leaves was measured per unit time (Ballhorn et al. 2010; Alonso-Amelot and Oliveros-Bastidas 2005) to determine cyanogenic capacity (HCNc). No remarkable difference in MeHNL infiltrated or SbDhr2 infiltrated leaves was observed, whereas light colour change was observed in leaves co-expressing both enzymes after 9 ~ 10 h (Table 5).
Discussion
Sources of HCN in cotton plants and its detoxification pathways
Cyanogenic glycoside (CNglcs), also known as specialized secondary metabolites, is derived from amino acids, L-tryosine, L-valine, L-leucine, L-isolucine, L-phenylalanine with oximes and cyanohydrins as important intermediates. Enzyme CYC79 family of cytochrome P450 (Andersen et al. 2000; Bak et al. 2006; Jørgensen et al. 2011; Morant et al. 2003) is responsible for the synthesis of majority of oximes with E or Z configuration. The derivatives of oximes play diverse roles in plant defence, growth regulation and communication. Oximes in plants exist in E or Z configurations, with E-oxime having broader biological activities (Sørensen et al. 2018). Cyanohydrins (α-hydroxynitrile) in plants are converted from E-oximes by the action of CYC71 or CYC736 (Jørgensen et al. 2011; Takos et al. 2011) family of enzymes. UDP-glucosyl transferase catalyses the last step in the conversion of a cyanohydrin to a cyanogenic glucoside, dhurrin (UGT85B1), linamarin (UGT85K), prunasin (UGT85A19), lotaustralin (UGT85K). The degradation of cyanogenic glucoside into HCN and aglycones is a two-step process (Jørgensen et al. 2011; Gleadow and Moller 2014) catalysed by cyanogenic β-glucosidases (EC: 3.2.1.21) (Morant et al. 2008; Cressey and Reeve 2019; Esen 1993; Vetter 2017) and α-hydroxynitrile lyases [dhurrin (EC: 4.1.2.11), lotaustralin (EC: 4.1.2.46), linamarin (EC: 4.1.2.46 & 4.1.2.47) and prunasin (EC: 4.1.2.10)] (Dadashipour and Asano 2011; Asano et al. 2005; Kassim and Rumbold 2014).
Tiny amount of hydrogen cyanide is produced by all plants as a product or a co-product of a biosynthesis pathway. There are four reactions/ metabolic pathways that would liberate hydrogen cyanide in cotton on the basis of enzyme predictions (http://ptools.cottongen.org), i.e., ethylene biosynthesis I (plants) pathway (Xu and Zhang 2015), linustatin bioactivation (Schmidt et al. 2018; Jørgensen et al. 2005), neolinustatin bioactivation (Forslund et al. 2004; Lai et al. 2015) and vicianin bioactivation pathway (Mizutani et al. 2007).
HCN in plants is detoxified by two pathways. In the first pathway, HCN is converted to 3-cyano-L-alanine (Machingura et al. 2016), and is further metabolized to L-asparagine and L-aspartate (Asparagine pathway); in the second pathway, thiosulfate sulfurtransferase (rhodanese) (Nakajima 2015; Steiner et al. 2018) catalyses the conversion of thiosulfate and cyanide to thiocyanate and sulfite.
Heterologous expression of SbDhr2 and MeHNL in aerial parts of cotton
We have successfully demonstrated that transient co-expression of SbDhr2 and MeHNL could help to deter S. litura from feeding on cotton leaves. Higher expression of SbDhr2 compared with MeHNL gene was observed in Western blotting, which can be attributed to the choice of promoters, the use of PEPC (Matsuoka et al. 1994) and 2X CaMV 35S (Samac et al. 2004; Christensen et al. 1992; Weeks et al. 1993). According to hydrogen cyanide release detection by using Fiegl-Anger test paper, no colour change was observed in control and leaf tissue independently infiltrated with pTD2 or pTH2, whereas light colour change was observed after more than 9 h in leaf tissue samples co-infiltrated with pTD2 & pTH2 construct.
Bioinformatic analysis
Cyanogenesis in green tissue of cotton (Radin 1972), presence of (R)-mandelonitrile lyase-like enzyme (UniProtKB - A0A1U8PEZ9) E.C: 4.1.2.10 and β-glucosidae (UniProtKB - Q7XAS3) E.C: 3.2.1.21 in G. hirsutum have been reported, the former having 74% amino acid sequence similarity with M. esculanta (R)-mandelonitrile lyase and the latter having 76.1% identity with S. bicolor beta-glucosidase. Phenylalanine N-monooxygenase (E.C: 1.14.14.40) catalysing the conversion of L-phenylalanine to phenylacetaldoxime has been reported in G. hirsutum. Genome sequencing of G. hirsutum reported cyanohydrin beta-glucosyltransferase / uridine diphosphoglucose: aldehydecyanohydrin beta-glucosyltransferase (E.C: 2.4.1.85) gene sequence, the gene involved in the conversion of a cyanohydrin to cyanogenic glucoside. In our previous study (Mahajan et al. 2015), we have explored the possibilities of engineering SbDhr2 enzyme to broaden its substrate specificity, and extensive work on hydroxynitrile lyase regarding improving its substrate specificity and activity (Dadashipour and Asano 2011; Yan et al. 2003; Cheng et al. 2001; Dadashipour et al. 2011; Semba et al. 2008; Semba et al. 2010; Bühler et al. 2003; Lauble et al. 2001) have been published, which can aid in developing herbivory resistant variety of cotton (cyanogenic).
Lessons learned from transient expression
These findings along with the previous work (Pant et al. 2016) indicate that α-hydroxynitrile glucoside exists in G. hirsutum. There are possibilities that cyanide detoxification route/pathway (Gleadow and Moller 2014; Machingura et al. 2016; Ting and Zschoche 1970; Zagrobelny et al. 2004; Miller and Conn 1980; Sun et al. 2018; Nielsen et al. 2016; Pičmanová et al. 2015) is more active in cotton. The key to herbivore deterrence is the rapid release of HCN (Krothapalli et al. 2013) and HCNp (Miller and Conn 1980; Bokanga et al. 1994) against pests (Howe and Jander 2008; Gleadow et al. 2002). Engineering metabolic pathways in acyanogenic /cyanogenic plants are all about trails and errors (Morant et al. 2007).
Conclusion
This study was conducted to investigate whether transient expression of cyanogenic pathway enzymes in aerial parts of cotton protects plants against herbivory by S. litura. The results presented here clearly support the finding that transient co-expression of cyanoamino acid metabolism pathway enzymes can deter S. litura from feeding on cotton leaves. It has also demonstrated that strong green tissue-specific promoter of enzyme/transgene expression is a prerequisite for enhancing HCNp in cotton. These findings extrapolate novel opportunities for metabolic engineering of cyanogenesis in G. hirsutum, for which detailed knowledge of metabolic cross-talk, cyanogenic glucoside synthesis, transport, regulation and degradation is a prerequisite. Engineering cyanogenesis in cotton can be envisioned as an additional pest control strategy.
Availability of data and materials
All data generated or analysed in this study are included in the published article.
Change history
13 August 2020
An amendment to this paper has been published and can be accessed via the original article.
Abbreviations
- HCN:
-
Hydrogen cyanide
- MeHNL :
-
Mannihot esculanta α-hydroxynitrile lyase
- PEPC:
-
Phosphoenolpyruvate carboxylase
- SbDhr2:
-
Sorghum bicolor dhurrinase2
- TRV:
-
Tobacco rattle virus
- VIGS:
-
Virus-induced gene silencing
References
Ahmad M, Gull S. Susceptibility of armyworm Spodoptera litura (Lepidoptera: Noctuidae) to novel insecticides in Pakistan. Can Entomol. 2017;149(5):649–61. https://doi.org/10.4039/tce.2017.29.
Akula R, Ravishankar GA. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal Behav. 2011;6(11):1720–31. https://doi.org/10.4161/psb.6.11.17613.
Alonso-Amelot ME, Oliveros-Bastidas A. Kinetics of the natural evolution of hydrogen cyanide in plants in neotropical Pteridium arachnoideum and its ecological significance. J Chem Ecol. 2005;31(2):315–31. https://doi.org/10.1007/s10886-005-1343-z.
Andersen MD, Busk PK, Svendsen I, Møller BL. Cytochromes p-450 from cassava (Manihot esculenta Crantz) catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin. Cloning, functional expression in Pichia pastoris, and substrate specificity of the isolated recombinant enzymes. J Biol Chem. 2000;275(3):1966–75. https://doi.org/10.1074/jbc.275.3.1966.
Asano Y, Tamura K, Doi N, et al. Screening for new hydroxynitrilases from plants. Biosci Biotechnol Biochem. 2005;69(12):2349–57. https://doi.org/10.1271/bbb.69.2349.
Bak S, Olsen CE, Halkier BA, Møller BL. Transgenic tobacco and arabidopsis plants expressing the two multifunctional Sorghum cytochrome p450 enzymes, cyp79a1 and cyp71e1, are cyanogenic and accumulate metabolites derived from intermediates in dhurrin biosynthesis. Plant Physiol. 2000;123(4):1437–48. https://doi.org/10.1104/pp.123.4.1437.
Bak S, Paquette SM, Morant M, et al. Cyanogenic glycosides: a case study for evolution and application of cytochromes p450. Phytochem Rev. 2006;5(2–3):309–29. https://doi.org/10.1007/s11101-006-9033-1.
Ballhorn DJ, Kautz S, Lieberel R. Comparing responses of generalist and specialist herbivores to various cyanogenic plant features. Entomologia Experimentalis et Applicata. 2010;134(3):245–59. https://doi.org/10.1111/j.1570-7458.2009.00961.x.
Bartwal A, Mall R, Lohani P, et al. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J Plant Growth Regul. 2013;32(1):216–32. https://doi.org/10.1007/s00344-012-9272-x.
Becker A. Virus-induced gene silencing: methods and protocols. Totowa: Humana Press; 2013. https://doi.org/10.1007/978-1-62703-278-0.
Blomstedt CK, O'Donnell NH, Bjarnholt N, et al. Metabolic consequences of knocking out UGT85B1, the gene encoding the glucosyltransferase required for synthesis of dhurrin in Sorghum bicolor (L. Moench). Plant Cell Physiol. 2016;57(2):373–86. https://doi.org/10.1093/pcp/pcv153.
Bokanga M, Ekanayake IJ, Dixon AGO, Porto MCM. Genotype-environment interactions for cyanogenic potential in cassava. Acta Hortic. 1994;375:131–40. https://doi.org/10.17660/actahortic.1994.375.11.
Bragard C, Dehnen-Schmutz K, Di Serio F, et al. Pest categorisation of Spodoptera litura. EFSA J. 2019;17(7). https://doi.org/10.2903/j.efsa.2019.5765.
Bühler H, Effenberger F, Förster S, et al. Substrate specificity of mutants of the hydroxynitrile lyase from Manihot esculenta. Chembiochem. 2003;4(2–3):211–6. https://doi.org/10.1002/cbic.200390033.
Cheng T, Wu J, Wu Y, et al. Genomic adaptation to polyphagy and insecticides in a major east Asian noctuid pest. Nat Ecol Evol. 2017;1(11):1747–56. https://doi.org/10.1038/s41559-017-0314-4.
Cheng SH, Yan GH, Wu J, Sun WR. Cloning, expression and preliminary application of a alpha-hydroxynitrile lyase from cassave. Sheng Wu Gong Cheng Xue Bao. 2001;17(1):78–83. (in Chinese). https://www.ncbi.nlm.nih.gov/pubmed/11330194.
Christensen AH, Sharrock RA, Quail RH. Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol Biol. 1992;18(4):675–89. https://doi.org/10.1007/bf00020010.
Chueskul S, Chulavatnatol M. Properties of alpha-hydroxynitrile lyase from the petiole of cassava (Manihot esculenta Crantz). Arch Biochem Biophys. 1996;334(2):401–5. https://doi.org/10.1006/abbi.1996.0471.
Cicek M, Esen A. Structure and expression of a dhurrinase (beta-glucosidase) from sorghum. Plant Physiol. 1998;116(4):1469–78. https://doi.org/10.1104/pp.116.4.1469.
Cressey P, Reeve J. Metabolism of cyanogenic glycosides: a review. Food Chem Toxicol. 2019;125:225–32. https://doi.org/10.1016/j.fct.2019.01.002.
Dadashipour M, Asano Y. Hydroxynitrile lyases: insights into biochemistry, discovery, and engineering. ACS Catal. 2011;1(9):1121–49. https://doi.org/10.1021/cs200325q.
Dadashipour, Fukuta Y, Asano Y. Comparative expression of wild-type and highly soluble mutant his103leu of hydroxynitrile lyase from Manihot esculenta in prokaryotic and eukaryotic expression systems. Protein Expr Purif. 2011;77(1):92–7. https://doi.org/10.1016/j.pep.2010.12.010.
Esen A. ß-glucosidases. Washington, D.C.: American Chemical Society; 1993. https://doi.org/10.1021/bk-1993-0533.
Fand BB, Sul NT, Bal SK, Minhas PS. Temperature impacts the development and survival of common cutworm (Spodoptera litura): simulation and visualization of potential population growth in India under warmer temperatures through life cycle modelling and spatial mapping. PLoS One. 2015;10(4):e0124682. https://doi.org/10.1371/journal.pone.0124682.
Feigl F, Anger V. Replacement of benzidine by copper ethylacetoacetate and tetra base as spot-test reagent for hydrogen cyanide and cyanogen. Analyst. 1966;91(1081):282–4. https://doi.org/10.1039/an9669100282.
Forslund K, Morant M, Jørgensen B, et al. Biosynthesis of the nitrile glucosides rhodiocyanoside a and d and the cyanogenic glucosides lotaustralin and linamarin in Lotus japonicus. Plant Physiol. 2004;135(1):71–84. https://doi.org/10.1104/pp.103.038059.
Franks TK, Powell KS, Choimes S, et al. Consequences of transferring three sorghum genes for secondary metabolite (cyanogenic glucoside) biosynthesis to grapevine hairy roots. Transgenic Res. 2006;15(2):181–95. https://doi.org/10.1007/s11248-005-3737-7.
Gao X, Britt RC Jr, Shan L, He P. Agrobacterium-mediated virus-induced gene silencing assay in cotton. J Vis Exp. 2011;(54):e2938. https://doi.org/10.3791/2938.
Gleadow RM, Møller BL. Cyanogenic glycosides: synthesis, physiology, and phenotypic plasticity. Annu Rev Plant Biol. 2014;65:155–85. https://doi.org/10.1146/annurev-arplant-050213-040027.
Gleadow RM, Foley WJ, Woodrow IE. Enhanced CO2 alters the relationship between photosynthesis and defence in cyanogenic Eucalyptus cladocalyx F. Muell. Plant Cell Environ. 1998;21(1):12–22. https://doi.org/10.1046/j.1365-3040.1998.00258.x.
Gleadow RM, Woodrow IE. Mini-review: constraints on effectiveness of cyanogenic glycosides in herbivore defense. J Chem Ecol. 2002;28(7):1301–13. https://doi.org/10.1023/A:1016298100201.
Hay-Roe MM, Meagher RL, Nagoshi RN. Effects of cyanogenic plants on fitness in two host strains of the fall armyworm (Spodoptera frugiperda). J Chem Ecol. 2011;37(12):1314–22. https://doi.org/10.1007/s10886-011-0049-7.
Healey A, Furtado A, Cooper T, Henry RJ. Protocol: a simple method for extracting next-generation sequencing quality genomic DNA from recalcitrant plant species. Plant Methods. 2014;10:21. https://doi.org/10.1186/1746-4811-10-21.
Howe GA, Jander G. Plant immunity to insect herbivores. Annu Rev Plant Biol. 2008;59:41–66. https://doi.org/10.1146/annurev.arplant.59.032607.092825.
Jørgensen K, Bak S, Busk PK, et al. Cassava plants with a depleted cyanogenic glucoside content in leaves and tubers. Distribution of cyanogenic glucosides, their site of synthesis and transport, and blockage of the biosynthesis by RNA interference technology. Plant Physiol. 2005;139(1):363–74. https://doi.org/10.1104/pp.105.065904.
Jørgensen K, Morant AV, Morant M, et al. Biosynthesis of the cyanogenic glucosides linamarin and lotaustralin in cassava: isolation, biochemical characterization, and expression pattern of cyp71e7, the oxime-metabolizing cytochrome p450 enzyme. Plant Physiol. 2011;155(1):282–92. https://doi.org/10.1104/pp.110.164053.
Kassim MA, Rumbold K. HCN production and hydroxynitrile lyase: a natural activity in plants and a renewed biotechnological interest. Biotechnol Lett. 2014;36(2):223–8. https://doi.org/10.1007/s10529-013-1353-9.
Kesselmeier J, Urban B. Subcellular localization of saponins in green and etiolated leaves and green protoplasts of oat (Avena sativa L.). Protoplasma. 1983;114(1–2):133–40. https://doi.org/10.1007/BF01279877.
Krothapalli K, Buescher EM, Li X, et al. Forward genetics by genome sequencing reveals that rapid cyanide release deters insect herbivory of Sorghum bicolor. Genetics. 2013;195(2):309–18. https://doi.org/10.1534/genetics.113.149567.
Lai D, Pičmanová M, Hachem MA, et al. Lotus japonicus flowers are defended by a cyanogenic beta-glucosidase with highly restricted expression to essential reproductive organs. Plant Mol Biol. 2015;89(1–2):21–34. https://doi.org/10.1007/s11103-015-0348-4.
Lauble H, Miehlich B, Förster S, et al. Mechanistic aspects of cyanogenesis from active-site mutant ser80ala of hydroxynitrile lyase from Manihot esculenta in complex with acetone cyanohydrin. Protein Sci. 2001;10(5):1015–22. https://doi.org/10.1110/ps.01301.
Lauble H, Miehlich B, Förster S, et al. Structure determinants of substrate specificity of hydroxynitrile lyase from Manihot esculenta. Protein Sci. 2002;11(1):65–71. https://doi.org/10.1110/ps.33702.
Li H, Li K, Guo Y, et al. A transient transformation system for gene characterization in upland cotton (Gossypium hirsutum). Plant Methods. 2018;14:50. https://doi.org/10.1186/s13007-018-0319-2.
Machingura M, Salomon E, Jez JM, Ebbs SD. The beta-cyanoalanine synthase pathway: beyond cyanide detoxification. Plant Cell Environ. 2016;39(10):2329–41. https://doi.org/10.1111/pce.12755.
Mahajan C, Patel K, Khan BM, Rawal SS. In silico ligand binding studies of cyanogenic beta-glucosidase, dhurrinase-2 from Sorghum bicolor. J Mol Model. 2015;21(7):184. https://doi.org/10.1007/s00894-015-2730-1.
Matsuoka M, Kyozuka J, Shimamoto K, Kano-Murakami Y. The promoters of two carboxylases in a C4 plant (maize) direct cell-specific, light-regulated expression in a C3 plant (rice). Plant J. 1994;6(3):311–9. https://doi.org/10.1046/j.1365-313x.1994.06030311.x.
Miller JM, Conn EE. Metabolism of hydrogen cyanide by higher plants. Plant Physiol. 1980;65(6):1199–202. https://doi.org/10.1104/pp.65.6.1199.
Mizutani M, Saino H, Mizutani M, et al. Vicianin hydrolase is a novel cyanogenic beta-glycosidase specific to beta-vicianoside (6-o-alpha-l-arabinopyranosyl-beta-d-glucopyranoside) in seeds of Vicia angustifolia. Plant Cell Physiol. 2007;48(7):938–47. https://doi.org/10.1093/pcp/pcm065.
Morant AV, Jørgensen K, Jørgensen C, et al. Beta-glucosidases as detonators of plant chemical defense. Phytochemistry. 2008;69(9):1795–813. https://doi.org/10.1016/j.phytochem.2008.03.006.
Morant M, Bak S, Møller BL, et al. Plant cytochromes p450: tools for pharmacology, plant protection and phytoremediation. Curr Opin Biotechnol. 2003;14(2):151–62. https://doi.org/10.1016/s0958-1669(03)00024-7.
Morant VA, Jørgensen K, Jørgensen B, et al. Lessons learned from metabolic engineering of cyanogenic glucosides. Metabolomics. 2007;3(3):383–98. https://doi.org/10.1007/s11306-007-0079-x.
Nakajima T. Roles of sulfur metabolism and rhodanese in detoxification and anti-oxidative stress functions in the liver: responses to radiation exposure. Med Sci Monit. 2015;21:1721–5. https://doi.org/10.12659/msm.893234.
Nielsen LJ, Stuart P, Pičmanová M, et al. Dhurrin metabolism in the developing grain of Sorghum bicolor (L.) Moench investigated by metabolite profiling and novel clustering analyses of time-resolved transcriptomic data. BMC Genomics. 2016;17(1):1021. https://doi.org/10.1186/s12864-016-3360-4.
Pang J, Zhu Y, Li Q, et al. Development of agrobacterium-mediated virus-induced gene silencing and performance evaluation of four marker genes in Gossypium barbadense. PLoS One. 2013;8(9):e73211. https://doi.org/10.1371/journal.pone.0073211.
Pant SR, McNeece BT, Sharma K, et al. The heterologous expression of a Glycine max homolog of nonexpressor of pr1 (npr1) and α-hydroxynitrile glucosidase suppresses parasitism by the root pathogen Meloidogyne incognita in Gossypium hirsutum. J Plant Interact. 2016;11(1):41–52. https://doi.org/10.1080/17429145.2016.1163423.
Panter KE. Chapter 64 - cyanogenic glycoside–containing plants. In: Gupta RC, editor. Veterinary toxicology. 3rd ed. Amsterdam: Academic Press; 2018. p. 935–40. https://doi.org/10.1016/B978-0-12-811410-0.00064-7.
Pauli S, Rothnie HM, Chen G, et al. The cauliflower mosaic virus 35S promoter extends into the transcribed region. J Virol. 2004;78(22):12120–8. https://doi.org/10.1128/jvi.78.22.12120-12128.2004.
Pičmanová M, Neilson EH, Motawia MS, et al. A recycling pathway for cyanogenic glycosides evidenced by the comparative metabolic profiling in three cyanogenic plant species. Biochem J. 2015;469(3):375–89. https://doi.org/10.1042/bj20150390.
Poulton JE, Li CP. Tissue level compartmentation of (r)-amygdalin and amygdalin hydrolase prevents large-scale cyanogenesis in undamaged Prunus seeds. Plant Physiol. 1994;104(1):29–35. https://doi.org/10.1104/pp.104.1.29.
Radin JW. Cyanogenesis in green tissues of cotton plants. Crop Sci. 1972;12(5):609–11. https://doi.org/10.2135/cropsci1972.0011183x001200050017x.
Rosenthal GA, Berenbaum MR. Herbivores. Their interactions with secondary plant metabolites, the chemical participants, vol. 1. 2nd ed. In: Rosenthal GA, Berenbaum MR, editors. The chemical participants. Hoboken: Wiley; 1992. https://doi.org/10.1111/j.1365-3032.1992.tb01027.x.
Samac DA, Tesfaye M, Dornbusch M, et al. A comparison of constitutive promoters for expression of transgenes in alfalfa (Medicago sativa). Transgenic Res. 2004;13(4):349–61. https://doi.org/10.1023/B:TRAG.0000040022.84253.12.
Saunders JA, Conn EE. Presence of the cyanogenic glucoside dhurrin in isolated vacuoles from Sorghum. Plant Physiol. 1978;61(2):154–7. https://doi.org/10.1104/pp.61.2.154.
Schaller A. Induced plant resistance to herbivory. Dordrecht: Springer Netherlands; 2008. https://doi.org/10.1007/978-1-4020-8182-8.
Schmidt FB, Cho SK, Olsen CE, et al. Diurnal regulation of cyanogenic glucoside biosynthesis and endogenous turnover in cassava. Plant Direct. 2018;2(2):e00038. https://doi.org/10.1002/pld3.38.
Semba H, Dobashi Y, Matsui T. Expression of hydroxynitrile lyase from Manihot esculenta in yeast and its application in (s)-mandelonitrile production using an immobilized enzyme reactor. Biosci Biotechnol Biochem. 2008;72(6):1457–63. https://doi.org/10.1271/bbb.70765.
Semba H, Ichige E, Imanaka T, et al. Efficient production of active form recombinant cassava hydroxynitrile lyase using Escherichia coli in low-temperature culture. Methods Mol Biol. 2010;643:133–44. https://doi.org/10.1007/978-1-60761-723-5_10.
Senthil-Kumar M, Mysore KS. Tobacco rattle virus-based virus-induced gene silencing in Nicotiana benthamiana. Nat Protoc. 2014;9(7):1549–62. https://doi.org/10.1038/nprot.2014.092.
Sørensen M, Neilson EHJ, Møller BL. Oximes: unrecognized chameleons in general and specialized plant metabolism. Mol Plant. 2018;11(1):95–117. https://doi.org/10.1016/j.molp.2017.12.014.
Steiner A-M, Busching C, Vogel H, Wittstock U. Molecular identification and characterization of rhodaneses from the insect herbivore Pieris rapae. Sci Rep. 2018;8(1):10819. https://doi.org/10.1038/s41598-018-29148-5.
Sun Z, Zhang K, Chen C, et al. Biosynthesis and regulation of cyanogenic glycoside production in forage plants. Appl Microbiol Biotechnol. 2018;102(1):9–16. https://doi.org/10.1007/s00253-017-8559-z.
Takos AM, Knudsen C, Lai D, et al. Genomic clustering of cyanogenic glucoside biosynthetic genes aids their identification in Lotus japonicus and suggests the repeated evolution of this chemical defence pathway. Plant J. 2011;68(2):273–86. https://doi.org/10.1111/j.1365-313x.2011.04685.x.
Tattersall DB, Bak S, Jones PR, et al. Resistance to an herbivore through engineered cyanogenic glucoside synthesis. Science. 2001;293(5536):1826–8. https://doi.org/10.1126/science.1062249.
Thayer SS, Conn EE. Subcellular localization of dhurrin beta-glucosidase and hydroxynitrile lyase in the mesophyll cells of sorghum leaf blades. Plant Physiol. 1981;67(4):617–22. https://doi.org/10.1104/pp.67.4.617.
Ting IP, Zschoche WC. Asparagine biosynthesis by cotton roots. Carbon dioxide fixation and cyanide incorporation. Plant Physiol. 1970;45(4):429–34. https://doi.org/10.1104/pp.45.4.429.
Verdoucq L, Czjzek M, Moriniere J, et al. Mutational and structural analysis of aglycone specificity in maize and sorghum beta-glucosidases. J Biol Chem. 2003;278(27):25055–62. https://doi.org/10.1074/jbc.m301978200.
Verdoucq L, Morinière J, Bevan D, et al. Structural determinants of substrate specificity in family 1 beta-glucosidases: novel insights from the crystal structure of sorghum dhurrinase-1, a plant beta-glucosidase with strict specificity, in complex with its natural substrate. J Biol Chem. 2004;279(30):31796–803. https://doi.org/10.1074/jbc.m402918200.
Vetter J. Plant cyanogenic glycosides. In: Carlini CR, Ligabue-Braun R, editors. Plant toxins. Dordrecht: Springer Netherlands; 2017. p. 287–317. https://doi.org/10.1007/978-94-007-6464-4_19.
Wajant H, Pfizenmaier K. Identification of potential active-site residues in the hydroxynitrile lyase from Manihot esculenta by site-directed mutagenesis. J Biol Chem. 1996;271(42):25830–4. https://doi.org/10.1074/jbc.271.42.25830.
War AR, Paulraj MG, Ahmad T, et al. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012;7(10):1306–20. https://doi.org/10.4161/psb.21663.
Weeks JT, Anderson OD, Blechl AE. Rapid production of multiple independent lines of fertile transgenic wheat (Triticum aestivum). Plant Physiol. 1993;102(4):1077–84. https://doi.org/10.1104/pp.102.4.1077.
Weigel D, Glazebrook J. Transformation of agrobacterium using the freeze-thaw method. In: Weigel D, Glazebrook J, editors. Cold Spring Harbor protocols (2006/01/01 ed., Vol. 2006). Cold Spring Harbor: Cold Spring Harbor Laboratory; 2006. https://doi.org/10.1101/pdb.prot4666.
Xu J, Zhang S. Ethylene biosynthesis and regulation in plants. In: Wen C-K, editor. Ethylene in plants. Dordrecht: Springer Netherlands; 2015. pp. 1–25. https://doi.org/10.1007/978-94-017-9484-8_1.
Xue M, Pang YH, Wang HT, et al. Effects of four host plants on biology and food utilization of the cutworm, Spodoptera litura. J Insect Sci. 2010;10(1):1–14. https://doi.org/10.1673/031.010.2201.
Yan G, Cheng S, Zhao G, et al. A single residual replacement improves the folding and stability of recombinant cassava hydroxynitrile lyase in E. coil. Biotechnol Lett. 2003;25(13):1041–7. https://doi.org/10.1023/A:1024182228057.
Zagrobelny M, Bak S, Rasmussen AV, et al. Cyanogenic glucosides and plant-insect interactions. Phytochemistry. 2004;65(3):293–306. https://doi.org/10.1016/j.phytochem.2003.10.016.
Acknowledgements
We thank Dr. Farooqui, Mazhar, the principal of Maulana Azad College of Arts, Science and Commerce, for his critical comments and technical help.
Funding
This work was funded by the Dept. of Botany, Maulana Azad College of Arts, Science and Commerce, Aurangabad, Maharashtra, India and Government Institute of Science, Aurangabad, Maharashtra, India.
Author information
Authors and Affiliations
Contributions
Mahajan C conducted this study for his PhD research, designed, investigated and collected the data. Naser R as supervisor and Gupta S as co-supervisor guided and provided technical expertise in this study for PhD studies of Mahajan C. All authors reviewed, read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
MAHAJAN, C., NASER, R. & GUPTA, S. Transient expression of SbDhr2 and MeHNL in Gossypium hirsutum for herbivore deterrence assay with Spodoptera litura. J Cotton Res 3, 2 (2020). https://doi.org/10.1186/s42397-020-0044-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42397-020-0044-z