
Abstract
Background
Amphibian parasites have been used as models for understanding important issues about host activity and sensitivity to environmental perturbations and amphibian decline. With the aim of comparing the parasitic fauna in Amietophrynus regularis from natural habitat and dumpsites within Lagos metropolis, a total of 172 toads were sampled to determine the occurrence of the gastrointestinal tract and blood parasites.
Methods
The liver, lungs, intestine and intestinal fluids, and blood were collected for parasitological determination. Histopathological analysis was carried out on the intestinal tracts.
Results
Helminths recovered from the gastrointestinal tract belonged to the family Nematoda, and they included Cosmocerca spp., Amplicaecum spp., Strongyloides spp., and oxyurides. Cosmocerca spp. was most abundant with a parasite load of 2304 and a prevalence of 70.93%. Protozoans recovered from the gastrointestinal fluids include Opalina spp. (7.5%) and Balantidium spp. (11.05%). The only protozoan recovered from blood was a species of Hepatozoon. Although parasite intensity was higher in larger females, comparison of parasites prevalence with morphometrics of Amietophrynus regularis was not significant at p = 0.05. There was also no significant difference (p = 0.05) between the parasite abundance and the location of collection. Histopathological findings revealed stunting of the villi with a high level of intestinal tissue alterations in infected Amietophrynus regularis from the natural habitat.
Conclusions
The high levels of parasitic infection together with the histopathological findings are indicative that the natural habitats of Amietophrynus regularis are being perturbed and this is of concern considering the prevailing issues of amphibian decline.
Similar content being viewed by others


Background
The common African toad (Amietophrynus regularis) is also known as the square-marked toad, or the African bouncing toad is a member of the Bufonidae. They are flat-bodied amphibians having warty skin and exhibiting sexual dimorphism with female toads being generally larger than males. Amietophrynus regularis (A. regularis) have been reported in various countries around the world. In Africa, Amietophrynus regularis is found in regions stretching from Senegal through West Africa to Central Africa and through North Africa to Egypt (Adejoju et al. 2014). Amietophrynus regularis occur in savannas, shrublands, grasslands, forests, rural gardens, rivers, freshwater lakes, and urban areas (Vasconcelos et al. 2010). They are used as a delicacy rich in protein in some parts of Africa, notably in Nigeria and Burkina Faso (Adediran et al. 2014). Also, adult Amietophrynus regularis feed on insects; hence, they are an important energy-efficient trophic link between invertebrates and other vertebrates (Akinsanya et al. 2020).
Amphibians are on an increasing biodiversity decline which is one of the most pressing environmental problems (Daszak, et al. 1999; Stuart et al. 2004; Collins 2010). The declining amphibian populations have attracted the attention of many scientists in the past two decades (Cunningham et al. 1996; Iyaji et al. 2015), and amphibian parasitism has been used as models for understanding important issues about host–parasite relationships and amphibian decline (Daszak et al. 2003). Some causes of amphibian population decline include: habitat loss or alteration (Bishop et al. 2012), introduced/exotic species that compete with, prey on, and parasitize native amphibians (Collins 2010), ultraviolet radiation, and chemical pollution (Blaustein et al. 2003), climate change and epidemic diseases (Alton and Franklin 2017; Akinsanya et al. 2020).
Amphibians have a rich parasite fauna, including viruses, protozoans, and helminths (Sulieman et al. 2015). Research on amphibian parasites particularly when anthropogenic pollution is to be monitored has increased especially because of their functional importance in animal communities and because certain groups of endoparasites are excellent at accumulating toxic metals and selected organic pollutants (Sures 2004; Sures et al. 2017). Helminths, in particular, can provide information about host activity, distribution, and sensitivity to environmental perturbations while also exhibiting direct vulnerability to such contaminants (Marcogliese and Cone 1997; Pietrock and Marcogliese 2003; Koprivnikar et al. 2012; Aisien et al. 2021).
Helminth parasites may result from factors such as polluted environments and poor food quality taken (Nworah and Olorunfemi 2011). The presence of gut helminth parasites causes damage to the intestinal mucosa, as they attack the sensitive epithelium, resulting in ulcers, severe enteritis, and extensive connective tissue proliferation which may lead to morbidity and eventual death. The presence of parasites in large numbers may also obstruct the digestive tracts (Iyaji et al. 2015).
Anthropogenic activities both directly and indirectly affect the diversity and infection rates of parasites, which by implication impacts the diversity and health of amphibians (Marco and Ortiz-Santaliestra 2009). An insight into the degree of changes occasioned by environmental alterations on the amphibian faunal composition can be inferred by comparing them with those of relatively undisturbed habitat (Aisien et al. 2009).
With the aim of comparing the parasitic fauna in Amietophrynus regularis from both natural habitat and dumpsites within Lagos metropolis, the objective of this study is to investigate the occurrence of parasitic fauna in the gastrointestinal tract and blood of Amietophrynus regularis, evaluating the histopathological alterations on the small intestine of these Amietophrynus regularis obtained at the various habitats.
Methods
Description of the study area
Three locations were used for the study including Ikorodu, Ojota, and Badagry, all located within the Lagos metropolis in Southwestern Nigeria (Fig. 1). The study sites in these locations consisted of dumpsites and natural habitats. These locations were chosen because of the anthropogenic activities such as dumping of organic, inorganic, and toxic waste products within the vicinity and the presence of the natural habitat (less disturbed habitat) of the Amietophrynus regularis within the location. The first study station was Ikorodu dumpsite (06°35.8042'N, 03°34.8016'E) and natural habitat (06° 35′ 46.00″ N, 03° 34.5683′ E). The second was Badagry dumpsite (06° 25′ 42'' N, 02° 53′ 25″ E) and natural habitat (06°24′ 49″ N, 06° 53′ 52″ E). The third location was Ojota dumpsite (06° 35′ 40″ N, 03° 22′ 39″ E) and natural habitat (06° 34′ 47″ N, 03° 23′ 37″ E).

Map showing sampled stations
Sample collections
A total of 172 live Amietophrynus regularis (50 males, 122 females) were obtained randomly from the dumpsites and natural habitat of three different locations (Ikorodu, Ojota, and Badagry) from September 2019 to January 2021. They were collected with a sweep net or by hand. Samples were transported in well-aerated plastic containers to the Zoology department laboratory annex at the University of Lagos. On arrival to the laboratory, the morphometric features were obtained and the Amietophrynus regularis dissected. Selected organs (liver, lungs, and intestine) were collected for histopathology, and the blood smears for microscopy were carried out.
Morphometric assessment of the Amietophrynus regularis
Sex determination in the Amietophrynus regularis was conducted through physical observation of the throat area. The males are characterized by the dark or green throat, while the females are characterized by the white throat. Females also have coiled oviducts which are absent in males. A confirmatory determination was equally carried out on the reproductive system according to the description of Kobayashi et al. (2018).
The weight of each Amietophrynus regularis was determined to the nearest 0.01 g using a battery-powered digital Camry weighing balance (model EK-1A Series) before harvesting the organs. Using a vernier caliper, five measurements were taken to the nearest 0.1 cm. These included:
-
Snout—urostyle length (SVL): This is the distance between the anterior tip of the snout and the posterior tip of the urostyle.
-
Length of forelimb (LF): This is the distance between the posterior end of the humerus and the tip of the longest finger.
-
Length of hind limb (LH): This is the distance between the tibial head and the tip of the fourth toe (which is the longest toe).
-
Width across head (HW): This is the greatest width of the head at the level of the tympanum.
-
Tympanic diameter (TD): This is the longitudinal distance between the outer margins of the tympanic annulus.
Laboratory analysis
Using sterile blades, the intestine of the Amietophrynus regularis specimens was eviscerated and placed in saline solution. The intestines were then dissected to obtain the enteric parasites. The lungs were also washed in saline solution. For further histopathological analysis, the intestine was preserved in separate sampling bottles containing Bouin's fluid. Thick and thin blood films were made on a clean glass slide for microscopy on blood parasites.
The recovered parasites were counted and recorded and then fixed in 70% alcohol. The enteric parasites collected from the Amietophrynus regularis were nematodes identified as Cosmocerca sp. and Amplicaecum africanum. The lung nematodes recovered were identified as Rhabdias spp. The identification procedure was carried out at the pathology laboratory of the Department of Veterinary Pathology, University of Ibadan, Nigeria. Parasite samples were then labeled according to the location and sex of Amietophrynus regularis collected.
Microscopy examination of blood and gastrointestinal samples
Blood specimens were obtained from clipped toes or the heart. Body fluid from the body cavity was also examined for the presence of parasites. The blood and body fluid specimens were first transferred to heparinized tubes. Thick and thin blood smears were prepared and left to air dry, and the thin film was fixed using absolute methanol. A working solution of Giemsa stain was prepared using the rapid method at 10% for 10 min after which it was flooded using buffered water at 7.0pH. The films were left to air dry and viewed using × 100 Objective with immersion oil. The direct examination method was used for the gastrointestinal samples. Intestinal samples were emulsified into normal saline and two drops were placed on the slide and a cover slip was placed on top. It was viewed using × 40 Objective.
Histopathology assessment
The intestinal tissues were placed in bottles containing Bouin’s fluid for 6 h, after which it was decanted and 10% buffered formalin was added to preserve the tissue. The tissues were routinely dehydrated in an ascending series of alcohol at 30 min intervals; they were then embedded in molten paraffin wax and allowed to solidify. The blocked tissues were sectioned at 4–5 microns processed and stained with hematoxylin and eosin (H&E) stains. The stained tissues were washed off in tap water, and the overstained ones were destained in 1% alcohol (Akinsanya et al. 2018). The tissues were mounted using DPX mountant and dried. Coverslips were then mounted over the sections and examined using a binocular dissecting microscope (American Optical Corporation, Model 570). The photomicrographs were taken with the aid of a Camera (INFINITY, 3-3URC 4.54 × 4.54 μm) in the pathology laboratory of the Department of Veterinary Pathology, the University of Ibadan, Nigeria.
Quality control and assurance
Glassware and dissecting kit used in this study were washed with detergent soaked in acetone and rinsed with tap water. They were then sealed with aluminum foil and autoclaved at 120 °C for 2 h. After drying and cooling, they were stored in a clean environment to prevent any accumulation of dust or other contaminants pending the next use. A single surgical blade was used per tissue sample, and thereafter, it was discarded safely. To avoid hand contamination of samples, sterile latex gloves and nose masks were used throughout the experimental session. For quality assurance analyte grade saline water was subjected to microbial and contamination analysis before use. All readings were taken in triplicate to minimize error.
Statistical analysis
The parasitological matrices: prevalence (%), mean intensity, and mean abundance, were analyzed and calculated as follows:
The relationship between the prevalence of parasite infection and host factors such as the habitat of the host, sex, weight, and length was examined from data pooled from the three sampled locations within Lagos using Student’s t test analysis at a 5% level of significance. All statistical analyses were done using SPSS version 17 for Windows.
Results
A total number of 172 specimens of Amietophrynus regularis were randomly obtained for the study. Ninety-three Amietophrynus regularis were collected from dumpsites, while 79 were collected from the natural habitat. The standard body length and weight of Amietophrynus regularis individuals were determined and categorized into different body lengths and weight cohorts according to the sex and sampling sites. The numbers of female and male Amietophrynus regularis examined at dumpsites were n = 69 and n = 24, respectively, and at the natural habitats, females were n = 53 and males n = 26. Female Amietophrynus regularis from both the dumpsites and natural habitats had a higher mean weight and body length than the males. The highest mean weight and body length were recorded from females from dumpsites (Table 1).
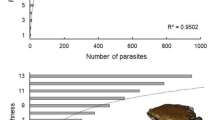
The prevalence of parasites in Amietophrynus regularis was generally > 50% (Additional file 1: Fig. SM1). From the three sampling locations, Ojota showed the highest prevalence of 90.19% followed by Badagry, 88.89%, and Ikorodu, 67.24%. The females of Amietophrynus regularis from Ojota dumpsite recorded the highest parasite load (923) with 100% prevalence. This was followed by female Amietophrynus regularis from Badagry natural habitat with a parasite load of 480 and a prevalence of 85.71%. The highest parasite load from the male counterparts was recorded in the Badagry natural habitat (221 with a prevalence of 100%. There was no significant difference (p = 0.145) between the sexes and parasite prevalence. Also, total parasite prevalence pooled from the three locations was higher at the natural habitat (86.08%) than the dumpsite (78.49%), but there was no significant difference (p = 0.698) in the parasite prevalence. The parasite abundance was also compared for the dumpsite and natural habitat between each location—Ikorodu, Badagry, and Ojota, and there was no significant difference as p = 0.606, p = 0.157, p = 0.135, respectively.
Parasites were recovered from different microhabitats of the gastrointestinal tract (stomach, small intestine, large intestine, and rectum) and the lungs.
Prevalence of gastrointestinal nematodes infection in Amietophrynus regularis
Out of the 172 Amietophrynus regularis examined, 141 Amietophrynus regularis were infected with nematodes with a prevalence of 81.98%. Both sexes were infected with a total worm load of 2,760, and female Amietophrynus regularis were observed to have a higher worm load (2,157) than males (603) (Table 2).
Nematodes recovered from the gastrointestinal tract include Cosmocerca sp., Amplicaecum africanum, and Rhabdias spp. The most abundant parasite was Cosmocerca sp. with 70.93% prevalence, and a total of 2,304 recovered from Amietophrynus regularis present in the 3 locations. Forty-five Amietophrynus regularis out of 54 sampled for Rhabdias spp. were infected with a prevalence of 83.33%. Amplicaecum africanum recorded the lowest prevalence (8.14%). Parasites microscopically detected in the gastrointestinal (GI) fluids include the larval nematode: oxyurides and Strongyloides sp. with a prevalence of 15.69% and 8.72%, respectively (Table 3).
Prevalence of protozoan infection in Amietophrynus regularis
A total number of 188 Opalina sp. and 336 Balantidium sp. with a prevalence of 7.5% and 11.05%, respectively, were detected in the gastrointestinal fluid. Microscopy examination of blood revealed the presence of Hepatozoon sp., an apicomplexan blood parasite genus of both warm-blooded and cold-blooded vertebrates (Fig. 2).

Microscopy examination of the blood and GI fluids of Amietophrynus regularis showing A Hepatozoon sp., B Strongyloides sp., C Balantidium sp., D oxyurides, E Opalina sp. (arrow), (F) Balantidium and oxyurides coinfection (same as with C). A at ×1000 magnification and B–F at ×400 magnifications
Parasite prevalence and amphibian morphometrics
The percentage parasite prevalence, parasite abundance, and parasite mean intensity of different weight cohorts (1–19, 20–39, 40–59, 60–79 g) and standard body length (3.0–3.9, 4.0–4.9, 5.0–5.9, 6.0–6.9, 7.0–7.9, 8.0–8.9 cm) of male and female Amietophrynus regularis from all stations were obtained for determination of their effects on the rate of parasitic infection (Additional file 2: Fig. SM2-3).
Prevalence above 50% was recorded in Amietophrynus regularis with body length that fell in general between 4.0–4.9 cm and 5.0–5.9 cm from both dumpsite and natural habitat of the three sampling locations. Female Amietophrynus regularis from Badagry natural habitat ranging from 3.0–7.9 all had a prevalence of 100% with the least being 80%. Female Amietophrynus regularis recovered from the Ojota dumpsite with body length ranging from 5.0–5.9 had the highest parasite load recorded in the study (Additional file 2: Fig. SM2).
Amietophrynus regularis used in the study mostly had weight ranges that fell between 1 and 39 g. The least recorded fell between 40 and 79 g. Amietophrynus regularis with weight ranging between 1 and 19 g recorded the highest prevalence in all the sampling locations with the exception of males and females from natural habitat. The only Amietophrynus regularis with a weight range of 60–79 g were sampled at Ikorodu dumpsite and had 100% prevalence (Additional file 2: Fig. SM3).
The morphometric values of Amietophrynus regularis pulled from each collection site showed a weak correlation with parasite abundance (Table 4) with weight at r = 0.090, head length (HDL) at r = 0.091, snout-to-vent length (SVL) at r = 0.143, forelimb (LF) at r = 0.095, hind limb (HL) at r = 0.065, tympanic diameter (TD) at r = 0.049.
Histopathological analysis
Histopathological assessment of intestinal tissues of Amietophrynus regularis from the dumpsites and pooled from the natural habitat three sampling locations showed normal tissue conditions of the villi structure, normal mucosa, submucosa, and muscularis, and well-preserved crypt-villous architecture in healthy Amietophrynus regularis, and this was common in tissues from the dumpsite. On the other hand, intestinal tissues of Amietophrynus regularis from the natural habitat exhibited mild to severe stunting of the villi and vascular congestion, lymphoid follicles within the villous structure, fibrosis of the villous, and detritus of the lumen, and this was common in tissues (Fig. 3).


Photomicrograph of intestinal tissues. With few exceptions of stunting of villi (A thin arrow), photomicrographs of intestinal tissues of both infected and uninfected Amietophrynus regularis from dumpsites show normal villi structure, normal mucosa, submucosa, and muscularis with well-preserved crypt-villous architecture (B–D). Comparatively, uninfected Amietophrynus regularis from natural habitat also showed disseminated moderate stunting of villi (E, F thin arrows). Tissues of infected Amietophrynus regularis from the natural habitat showed the most of villi structure alterations which included the presence of lymphoid follicles (G thin arrow), mild vascular congestion (H thin arrow), chronic inflammatory cells (I thicker arrow), and severe fibrosis of villous structure (J thicker arrows). The lighter arrows indicate stunting of the villi structure. Parasite presence was observed in the lumen (K thin arrow) and focal area of fibrosis of submucosa (L thin arrow)
Discussion
Amietophrynus regularis collected from the natural habitat and dumpsites of the three sampling areas in Lagos metropolis were found with a large number of parasites recovered from the gastrointestinal tract and lungs, and within the blood. The overall parasite infection of 81.98% in this study was compared favorably with the 94.17% recorded in 120 of the same host species examined in Kogi state (Iyaji et al. 2015), and also with 99% recorded in 162 samples examined in Ile-Ife (Ayodele and Akinpelu 2004). The predominant helminth group recovered was the nematode parasites, and the infection rate was confirmed with other findings in Kogi, Ile-Ife, Awka, and southwestern Nigeria (Iyaji et al. 2015; Ayodele and Akinpelu 2004; Nworah and Olorunfemi 2011; Aisien et al. 2001).
Cosmocerca spp., Rhabdias spp., oxyurides, and Amplicaecum africanum were among the predominant groups. Cosmocerca spp. was the most commonly encountered, with a prevalence of 70.93%. They naturally infect Amietophrynus regularis, and other studies have equally confirmed their high prevalence (Aisien et al. 2001; Hamann and González 2013; Iyaji et al. 2015). Furthermore, Cosmocerca spp. which inhabits moist soil and dry areas where these Amietophrynus regularis spend most of their time have a direct terrestrial life cycle in which the larvae penetrate the skin of the host before migrating to the large intestine (Saad et al. 2009). According to previous studies (Aisien et al. 2001), the prevailing ecological conditions of Lagos metropolis also account for the prevalence of Cosmocerca spp. and other nematodes encountered in the study.
Protozoan parasites detected included Hepatozoon spp. from the blood; Opalina sp. and Balantidium spp. from the intestinal fluids. Hepatozoon spp. detected in the blood of the Amietophrynus regularis has been reported in few studies outside Nigeria (Desser 2001; Sailasuta et al. 2011; Netherlands et al. 2014), but to the best of our knowledge, this is the first report of Hepatozoon spp. in Amietophrynus regularis in Nigeria. Sailasuta et al. (2011) described the heteroxenous life cycle of Hepatozoon spp. as having both sexual reproduction and sporogony occurring in an arthropod definitive host. Transmission occurs when the vertebrate host ingests such an arthropod, infected with mature oocysts, often containing thousands of sporozoites. These infective stages then migrate to the visceral organs, primarily to the liver, and undergo merogony. Meronts are released into the bloodstream where they form gamonts in the erythrocytes (Fig. 2).
The weak correlation between the morphometrics of the Amietophrynus regularis and parasite abundance shows that parasites infected Amietophrynus regularis irrespective of their size and weight, although other studies (Hamann et al. 2013; Nworah and Olorunfemi 2011; Iyaji et al. 2015) have affirmed positive correlation between Amietophrynus regularis size and increased parasitism. This could be as a result of some extrinsic factors such as the anthropogenic disturbances (e.g., environments perturbed by intensive agricultural activity), which could influence parasitic infection of the host (Hamann et al. 2006). At the same time, intrinsic factors such as the reproductive cycle of the host, feeding preferences, range of prey species, host body size, and the life cycle of the parasites are key factors in determining parasite abundance and species richness (Hamann et al. 2013).
Notwithstanding, the larger female Amietophrynus regularis from both natural habitats and dumpsites in this study were observed to be more parasitized than their male counterparts, although not statistically significant. This is yet indicative that the size ranges of the host favor higher parasitism and species richness as there are greater surface areas of the host vulnerable to parasite colonization. More so, as observed by Hamann and Gonzalez (2013), the differential behavior of female Amietophrynus regularis, that is different times in arriving at the source of larval infections, may equally explain the increased levels of parasitic infection.
There was no significant difference in parasite prevalence in relation to the habitats of collection, although there was an observed higher prevalence (86.08%) at the natural habitat than at the dumpsites (78.49%). Studies have shown that parasite transmission and abundance can be positively or negatively affected by their host environment (McKenzie 2007; Widmann 2013). Investigations into the degree of these changes occasioned by the host environment have been reported in a study that compared the amphibian parasitic faunal composition of degraded rainforest habitat and those of pristine or relatively undisturbed habitat in southwestern Nigeria. There was the presence of four Ptychadena species that were absent from the altered environment (Aisien et al. 2009). Thus, the insignificant difference in parasite prevalence from both the natural habitat and dumpsite as recorded in this study is a pointer to the possibility that these natural habitats where these Amietophrynus regularis were collected from are becoming degraded and disturbed just as with the dumpsites.
The comparative histopathology of the infected and uninfected intestinal tissues of the Amietophrynus regularis from dumpsites and natural habitats showed that there were more tissues from the dumpsites that showed normal villi structure, normal mucosa, submucosa, and muscularis than tissues of Amietophrynus regularis recovered from natural habitats. Tissues of infected Amietophrynus regularis from the natural habitat showed most of the alterations in villi structure. A possible explanation for these alterations could be attributed to the higher prevalence of parasites (86.08%) observed in Amietophrynus regularis from the natural habitat. Lymphoid follicles observed in infected tissues from natural habitats were indicative that immune cells had been secreted to fight against parasites causing the lymphoid follicles to be aggregated within the villous structure. This equally hampers the activities of the villi. Consequently, the mild vascular congestion also observed in the infected tissues from natural habitat generally results in altered permeability of the gastrointestinal tract, which can lead to the accumulation of endotoxins such as lipopolysaccharides and bacteria. Also, the mild–severe fibrosis and severe stunting of villous structure could be a result of injury to the villous due to the presence of the parasites or their waste product of metabolite. The fact that these alterations were predominantly reported from Amietophrynus regularis found in the natural habitat further demonstrates that the natural habitat of this Amietophrynus regularis has been disturbed by potentially several extrinsic factors other than the observed parasite prevalence. A recent study by Akinsanya et al. (2020) implicates trace metal accumulation in the intestine with marked alterations from Amietophrynus regularis recovered from both dumpsites and natural habitats.
Conclusions
This study has shown that there is a prevailing high level of parasitic infection in Amietophrynus regularis irrespective of the habitats (natural habitat or dumpsites). With the ongoing declines in amphibian populations in the environment, this is of concern. The histopathological finding on the effects of the parasites on the intestinal tissues further buttresses this concern. Furthermore, ecotoxicological factors such as heavy metals and pesticide accumulation in the intestine affect parasite abundance which could also contribute to histopathological alterations. Further studies are recommended to check these correlations. Cosmocerca spp. being the most abundant nematode in this study may have beneficial roles such as depuration benefit to the host. The net effect of parasitological harm based on its prevalence should also be looked into as they have life cycle stages that involve tissue penetration which could lead to secondary infections. A further study on this parasite is recommended. The Amietophrynus regularis used in this study shared common parasitic fauna as with other studies except for the protozoan—Hepatozoon, which is the first identification in Amietophrynus regularis from Nigeria. The finding of Hepatozoon is novel, and authors recommend further investigation in future with a combined morphological and molecular approach.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Adediran OA, Adebiyi AI, Uwalaka EC (2014) Detection of Kalicephalus spp., Ascaridoid spp and Strongyloides spp in Bufo regularis (Reuss) in Ibadan, southwest Nigeria. Adv Environ Biol 8(16):204–206
Aisien SO, Ugbo AD, Ilavbare AN, Ogunbor O (2001) Endoparasites of amphibians from south-western Nigeria. Acta Parasitol 46(4):299–305
Aisien MSO, Ogoannah SO, Imasuen AA (2009) Helminth parasites of amphibians from a rainforest reserve in southwestern Nigeria. Afr Zool 44(1):1–7
Aisien MS, Edo-Taiwo O, Imasuen AA (2021) Ecological scenarios and parasite diversity in anurans of West Africa: a review. Diversity 13(6):223
Akinsanya B, Adebusayo SA, Allison T, Ukwa UD (2018) Bioaccumulation of Polycyclic Aromatic Hydrocarbons, Histopathological Alterations and Parasite-Fauna in Benthopelagic Host from Snake Island, Lagos, Nigeria. J Basic Appl Zool 79:40. https://doi.org/10.1186/s41936-018-0046-2
Akinsanya B, Isibor PO, Onadeko B, Tinuade AA (2020) Impacts of trace metals on African common toad, Amietophrynus regularis (Reuss, 1833) and depuration effects of the Amietophrynus regularis’s enteric parasite, Amplicaecum africanum (Taylor, 1924) sampled within Lagos metropolis, Nigeria. Heliyon 6(3):e03570
Alton LA, Franklin CE (2017) Drivers of amphibian declines: effects of ultraviolet radiation and interactions with other environmental factors. Clim Change Responses 4(1):1–26
Ayodele HA, Akinpelu AI (2004) The helminth parasitofauna of Bufo regularis (Reuss) in Ile-Ife. IFE J Sci 6(2):101–104
Bishop PJ, Angulo A, Lewis, JP, Moore RD, Rabb GB, & Moreno JG (2012) The Amphibian Extinction Crisis-what will it take to put the action into the Amphibian Conservation Action Plan?. SAPI EN. S. Surveys and Perspectives Integrating Environment and Society, (5.2).
Blaustein AR, Romansic JM, Kiesecker JM, Hatch AC (2003) Ultraviolet radiation, toxic chemicals and amphibian population decline. Divers Distrib 9(2):123–140
Collins JP (2010) Amphibian decline and extinction: what we know and what we need to learn. Dis Aquat Org 92(2–3):93–99
Cunningham AA, Langton TES, Bennett PM, Lewin JF, Drury SEN, Gough RE, Macgregor SK (1996) Pathological and microbiological findings from incidents of unusual mortality of the common frog (Rana tempraria). Philos Trans R Soc Lond B Biol Sci 351:1539–1557
Daszak P, Berger L, Cunningham AA, Hyatt AD, Green DE, Speare R (1999) Emerging infectious diseases and amphibian population declines. Emerg Infect Dis 5(6):735–748
Daszak P, Cunningham AA, Hyatt AD (2003) Infectious disease and amphibian population decline. Divers Distrib 9(2):141–150
Desser SS (2001) The blood parasites of anurans from Costa Rica with reflections on the taxonomy of their trypanosomes. J Parasitol 87(1):152–160
Hamann MI, Kehr AI, González CE (2006) Species affinity and infracommunity ordination of helminths of Leptodactylus chaquensis (Anura: Leptodactylidae) in two contrasting environments from northeastern Argentina. J Parasitol 92(6):1171–1179. https://doi.org/10.1645/GE-862R1.1
Hamann M, Kehr A, González C (2013) Helminth communities in the burrowing Amietophrynus regularis, Rhinella fernandezae, from Northeastern Argentina. Biologia 68(6):1155–1162
Iyaji FO, Medayedupin IT, Echi PC, Falola OO, Omowaye OS (2015) Gastrointestinal helminth parasites of Amietophyrnus regularis, Bufonidae (African common Amietophrynus regularis) in Anyigba, Kogi State, Nigeria. Animal Res Int 12(2):2231–2240
Kobayashi K, Kitano T, Iwao Y, Kondo M (2018) Reproductive and developmental strategies: the continuity of life. Springer Nature, Berlin
Koprivnikar J, Marcogliese DJ, Rohr JR, Orlofske SA, Raffel TR, Johnson PTJ (2012) Macroparasite infections of amphibians: what can they tell us? EcoHealth 9(3):342–360. https://doi.org/10.1007/s10393-012-0785-3
Marco A, Ortiz-Santaliestra M (2009) Pollution: impact of reactive nitrogen on amphibians. Amphibian Biol 8:3145–3185
Marcogliese DJ, Cone DK (1997) Parasite communities as indicators of ecosystem stress. Parasitologia 39:227–232
McKenzie VJ (2007) Human land use and patterns of parasitism in tropical amphibian hosts. Biol Cons 137(1):102–116
Netherlands EC, Cook CA, Smit NJ (2014) Hepatozoon species (Adeleorina: Hepatozoidae) of African bufonids, with morphological description and molecular diagnosis of Hepatozoon ixoxo sp. Nov. parasitising three Amietophrynus species (Anura: Bufonidae). Parasites Vectors 7(1):1–12
Nworah DC, Olorunfemi OJ (2011) The helminth parasitofauna of Bufo regularis (Reuss) in Awka, Anambra State, Nigeria. Int J Parasitol Res 3(2):26–30
Pietrock M, Marcogliese DJ (2003) Free-living endohelminth stages: at the mercy of environmental conditions. Trends Parasitol 19:293–299
Saad AI, Khalifa R, Moustafa N (2009) Studies on the life cycle and identity of Paracosmocerca macronata (Nematoda: Cosmocercidae) in amphibians under experimental conditions. World J Zool 4(1):29–36
Sailasuta A, Satetasit J, Chutmongkonkul M (2011) Pathological study of blood parasites in rice field frogs, Hoplobatrachus rugulosus (Wiegmann, 1834). Vet Med Int 2011.
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fischman DL, Waller RW (2004) Status and trends of amphibian decline and extinctions worldwide. Science 306:1783–1786
Sulieman Y, Afifi A, Awad HA, Pengsakul T (2015) Helminth parasites of the subdesert Amietophrynus regularis, Amietophrynus (bufo) xeros (anura: bufonidae). Int J Res Granthaalayah 3(10):75–83
Sures B (2004) Environmental Parasitology: relevancy of parasites in monitoring environmental pollution. Trends Parasitol 20(4):170–177
Sures B, Nachev M, Selbach C, Marcogliese DJ (2017) Parasite responses to pollution: what we know and where we go in ‘Environmental Parasitology.’ Parasit Vectors 10(1):1–19
Vasconcelos R, Froufe E, Brito JC, Carranza S, Harris DJ (2010) Phylogeography of the African common Amietophrynus regularis, Amietophrynus regularis, based on mitochondrial DNA sequences: inferences regarding the Cape Verde population and biogeographical patterns. Afr Zool 45(2):291–298
Widmann M (2013) Impact of large-scale environmental features changes on host-parasite interaction in marine and freshwater ecosystems. BioSciences Master Rev 1:1–9
Acknowledgements
We appreciate the diligence and relentless efforts of Dr. Isibor Patrick Omoregie who help us through the experimentations, ensuring accuracy in data analysis and interpretation.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
A.B and O.M.O conceived and designed the experiments. O.M.O and U.T performed the experiments on the toads. O.M.O analyzed and interpreted the data. O.M.O and U.T contributed reagents, materials, analysis tools, and ideas. O.M.O wrote the Manuscript. All authors read and approved the final manuscripts.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval for this research was given by the Health Research Ethics committee of the College of Medicine of the University of Lagos. CMULHREC No: CMUL/ACUREC/03/20/729.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
. Indices of sex-based parasitic infection in Amietophrynus regularis.
Additional file 2
. Prevalence of Gastrointestinal Helminths Infections in Amietophrynus regularis.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Okeagu, O.M., Akinsanya, B. & Uzoka, T. A comparative study of parasitic fauna in Amietophrynus regularis (Reuss, 1833) from natural habitats and dumpsites within Lagos Metropolis, Southwest Nigeria. Bull Natl Res Cent 46, 105 (2022). https://doi.org/10.1186/s42269-022-00797-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-022-00797-x