
Abstract
Background
The western conifer seed bug, Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae), is one of the major pest in seed orchards of various conifers, significantly affects pine seed production and causes economic damage. Biological control by natural enemies has been well studied, but its entomopathogens have been poorly studied. In this study, the efficacy of twelve indigenous entomopathogenic fungi belonging to the genera Beauveria, Metarhizium, and Isaria on L. occidentalis adults was investigated.
Results
All isolates were found to be pathogenic at a concentration of 107 conidia/ml where the virulence ranged from 16 to 90%. Moreover, the virulence of two Metarhizium flavoviride isolates (As2 and As18) reached 90% at 107 conidia/ml concentration. In addition, L. occidentalis treated with M. flavoviride As18 (LT50 = 2.53 days) died more rapidly than with M. flavoviride As2 (LT50 = 5.83 days) at the same treated concentration. For concentration-dependent virulence for isolate As18, five conidia concentrations: 103, 104, 105, 106, and 107 conidia/ml were used. Also, the median lethal (LC50) value of As18 isolate was estimated to be 7.61 × 103 conidia/ml after 10 days of treatment. Since L. occidentalis is known to accumulate during overwintering, conidial transmission among adults was investigated. Unfortunately, horizontal transmission between adults did not occur.
Conclusion
The isolate M. flavoviride As18, which had low LC50 and LT50 values, appeared to be a promising biological control agent against L. occidentalis. This isolate should be formulated as a myco-insecticide and tested under field conditions in further studies.
Similar content being viewed by others


Background
The western conifer seed bug, Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) is an invader pest of coniferous trees. It is native to North America (Allen 1969) and spread from the central to the eastern regions since the mid-nineteenth century. In Europe, it was reported for the first time in Italy (1999) and recorded all over Europe at the beginning of the twenty-first century. It also reached China (Zhu 2010), Russia (Gapon 2013), Japan (Ishikawa and Kikuhara 2009), Turkey (Fent and Kment 2011), Tunisia (Ben Jamâa et al. 2013).
The pest is a sucking insect that feeds on cones and seeds of at least 30 conifer species (Farinha et al. 2021). The reported damages caused by the insect include massive abortion of conelets and a high percentage of empty seeds in ripened cones, a phenomenon known as Dry Cone Syndrome (El Khoury et al. 2021). Increasing populations of the pest have led to a 95% decline in pine nut production in Mediterranean countries (Parlak 2017). In addition, L. occidentalis is a vector of Diplodia sapinea spores, the fungus responsible for Diplodia tip blight of pine trees.
Generally, broad-spectrum insecticides have been used to control the pest. However, their usage is limited due to their toxic effects on bees and beneficial insects. To overcome this limitation, the use of entomopathogenic fungi (EPF) is a good alternative tool (Sonmez et al. 2022). Many studies covered the insecticidal activity of EPF (Biryol et al. 2022). About the use of fungi for the control of L. occidentalis, a preliminary study by Rumine and Barzanti (2008) showed that isolates of Beauveria bassiana were the most pathogenic to adults of L. occidentalis. In another study, Barta (2010) evaluated the virulence of B. bassiana, Isaria fumosorosea, and Metarhizium anisopliae against this pest under laboratory conditions and reported that isolates of I. fumosorosea were the most virulent. However, studies on the control of L. occidentalis with EPF are limited; therefore, the objective of the present study was to test the efficacy of 12 indigenous EPF against L. occidentalis and to determine the concentration response and horizontal transmission ability of the most effective isolate.
Methods
Insects
Leptoglossus occidentalis adults were collected using landing nets in Bursa, Turkey. The collected individuals were placed in rearing cages (50 × 40 × 40 cm) with perforated lids. Insects were fed on fresh immature pine cones at 20 ± 2 °C, 70 ± 10% relative humidity (RH) and under a natural photoperiod until bioassays were performed (Barta 2010).
Entomopathogenic fungi
The fungi used in the study were obtained from the microbiology laboratory of the Department of Biology, Karadeniz Technical University. Twelve EPFs belonging to the genera Beauveria, Metarhizium and Isaria were used in the study (Table 1). A loopful of spore suspension from glycerol stock culture was inoculated onto Potato Dextrose Agar (PDA) plates and incubated for 3 days at 28 ± 2 °C in the dark. Then, a single colony was transferred to a fresh PDA plate and incubated for 2 weeks until the plates were fully grown (Biryol et al. 2021a).
Preparation of conidia suspensions
Conidial suspensions were prepared from 2-week-old cultures. Fungal spores were collected from Petri dishes using a sterile scalpel and added to 10 ml of sterile distilled water containing 0.01% (v/v) Tween 80. The conidial suspensions were filtered through several layers of sterile muslin to remove fungal debris after the suspension was homogenized with a vortex for 2 min. Filtrates containing conidia were examined under a light microscope, using a hemocytometer, and conidia in the suspensions were quantified by direct counting. The concentration was adjusted to 107 conidia/ml for use in the bioassay (Biryol et al. 2021a).
Screening experiments
The efficacy of the fungi was tested on adults of L. occidentalis. After applying 5 ml (107 conidia/ml) of the fungal suspensions to the adults with a hand-operated sprayer, the insects were placed in a rearing cage (50 × 40 × 40 cm) containing fresh pine shoots. Thirty adults were used for each treatment and repeated three times. Control adults were treated with 0.01% aqueous Tween80. Bioassays were performed under laboratory conditions at 20 ± 2 °C, 70 ± 10% RH and at a natural photoperiod. Dead insects were transferred in a humidity chamber after surface sterilization with 70% alcohol to stimulate fungal sporulation. Mortality and mycosis were recorded daily for 10 days.
Concentration response experiment
For concentration-dependent virulence tests, the M. flavoviride As18 isolate with a mortality rate of 90% and a relatively short LT50 was used at five conidia concentrations: 103, 104, 105, 106, and 107 conidia/ml. L. occidentalis adults were sprayed using the same procedure as described above. Control adults were treated with an aqueous solution of 0.01% Tween 80. Three replicates of 30 larvae were used for each concentration. Treated adults were maintained as previously described, and each insect was monitored daily for 10 days after treatment.
Horizontal transmission experiment
Conidial transmission of As18 isolate among L. occidentalis was performed on overwintering adults under laboratory conditions. The experiment was performed as previously described by Biryol et al. (2021a) with a concentration of 1 × 107 conidia/ml. Horizontal transmission was studied with fungus-treated and untreated adults at four vector ratios of 0, 25, 50, 75, and 100% in groups of 40 insects with three replicates. After fungus-treated individuals were air-dried at room temperature, both treated and untreated individuals were released into the rearing cage. After 10 days, mortality and mycosis were assessed as described above.
Data analysis
Mortality and mycosis rates were corrected for control mortality using the Abbott formula (Abbott 1925). Data were analyzed using ANOVA and then the least significant difference (LSD) test to compare test isolates with each other and with the control group (p < 0.01). The lethal time value required to kill 50% (LT50) of L. occidentalis exposed to the fungal isolates and the median lethal concentration (LC50) of the most toxic isolate were estimated with 95% confidence interval by probit analysis. Kaplan–Meier survival analysis was also performed, followed by a log-rank test for multiple comparisons to compare different survival curves. The SPSS Statistics 25 software package was used for the analyses (SPSS Inc. Chicago, IL, USA).
Results
Screening and concentration response experiments
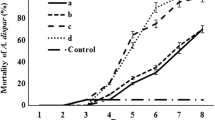
Screening tests were performed on L. occidentalis adults with 12 EPF whose insecticidal activity on various insects had been determined in previous studies. In screening tests, which were carried out at the concentration of 107 conidia/ml, all isolates were found to be pathogenic. However, a highly significant difference in mortality of L. occidentalis was observed among isolates (F = 92.2; df = 11 p < 0.01). Mortality rates ranged from 16 to 90%. Moreover, two of the M. flavoviride (As2 and As18) isolates caused 90% mortality (Fig. 1). In addition, L. occidentalis treated with M. flavoviride As18 died more rapidly than with M. flavoviride As2 at a conidial concentration of 107 conidia/ml. LT50 values were 5.83 and 2.53 days for isolates As2 and As18, respectively, at same concentration (Table 2). On the other hand, the lowest mortality rate was observed in the insects treated with B. bassiana KTU-24 (16%) and I. fumosorosea KTU-1 (26%). Mortality rate in the control group was less than 5%. Dead insects showed symptoms of M. flavoviride infection after incubation in the humid chamber (Fig. 2).

Screening test of entomopathogenic fungi on adults of the pest under laboratory condition at the concentration of 107 conidia/ml. Different case letters represent statistically significant differences among mortalities according to the least significant difference (LSD) multiple comparison test (p < 0.01). Mortality indicates the mean of three replications. Bars show standard error

Mycosis of the pest exposed to isolate of Metarhizium flavoviride As18. The white conidia were initially observed on the insect cadaver 6 days after treatment in the moist chamber, then green conidia were observed 10 days after treatment
Metarhizium flavoviride As18 isolate with 90% mortality rate and a relatively short LT50 was selected for the concentration–response experiment. Survival analysis showed that there was a significant difference among the concentrations (log-rank, p < 0.01). However, there was no statistical difference between the concentrations of 104 and 105 conidia/ml (log-rank, p > 0.01) (Fig. 3). The LC50 value of M. flavoviride As18 for L. occidentalis was estimated to be 7.61 × 103 conidia/ml using probit analysis.

Kaplan–Meier survival diagram for adult of Leptoglossus occidentalis exposed to isolate Metarhizium flavoviride As18 at five different concentrations. Holm-Sidak estimates of survival data show significant differences between treatment groups represented by lowercase letters
Horizontal transmission experiments
Conidial transmission from treated to untreated adult bugs over a 10-day period was assessed and As18 isolate showed no ability to disseminate. None of the insects were died in the non-treated control groups during experiments. Mortality rate was 22.7% at vector ratio 25%. A similar trend was observed at vector ratios 50 and 75% with 43.8 and 70% mortality, respectively. When 100% of adult beetles were treated with 107 conidia/ml concentration of As18, 95.5% mortality was observed (Table 3).
Discussion
Entomopathogenic fungi (EPF) have long been associated with different families in heteropteran species. In the present study, 12 isolates of three EPFs (B. bassiana, M. flavoviride and M. brunneum) were examined under laboratory conditions for their virulence to adult of L. occidentalis. Isolates of all three hypocrealean fungi were pathogenic to adults of L. occidentalis, although their virulence varied greatly among isolates. Isolates of M. flavoviride (As2 and As18) were more virulent than isolates of the other two fungal species. On the other hand, a preliminary study by Rumine and Barzanti (2008) showed that isolates of B. bassiana were most pathogenic to adults of L. occidentalis. In another study, Barta (2010) evaluated the virulence of B. bassiana, I. fumosorosea, and M. anisopliae to this pest under laboratory conditions and reported that isolates of I. fumosorosea were more virulent. Previous studies have shown that the difference in virulence of EPF is related to a higher germination rate, hyphal growth rate and production of cuticle-degrading enzymes because infection can potentially occur much more quickly (Zhang et al. 2011). In the present study, although M. flavoviride As2 and As18 isolates caused 90% mortality at 107 conidia/ml concentration in screening experiment, the LT50 values were 5.83 and 2.53 days at 107 conidia/ml concentration, respectively. Therefore, the concentration–response experiment was conducted with As18 isolate, which had the lowest LT50 value at the concentration of 107 conidia/ml. The LC50 value was estimated as 7.6 × 103 conidia/ml after 10 days of treatment for adults of L. occidentalis. Barta (2010) isolated two EPF, I. fumosorosea (AMSAS 06) and B. bassiana (AMSAS 03), from dead L. occidentalis and determined LC50 values as 0.86 × 105 and 10.3 × 105 conidia/ml, respectively. Also, they reported that soil originated B. bassiana (SUA a38) isolate was less virulent and LC50 value was 8.46 × 106 conidia/ml. Generally, isolates obtained from naturally infected insects are more virulent than those isolated from different habitats (Biryol et al. 2020). As in this study, EPF isolated from a particular host or soil have shown to be virulent to other hosts. Biryol et al. (2021b) reported that soil originated M. anisopliae (KTU-51) isolate showed 100% mortality on Orosanga japonica Melichar (Hemiptera: Ricaniidae).
Since L. occidentalis is known to accumulate for overwintering in many different places, e.g., under loose bark, in holes of dead logs, or in bird nests, conidial transmission among adults may be possible during this time. The tested isolate As18 had not transmitted horizontally within a time. This is the first report of horizontal transmission of EPF among the L. occidentalis population. A similar study to ours, found that nymph mortality of Cimex lectularius L. (Hemiptera: Cimicidae) induced by horizontal transfer was low using B. bassiana and did not reach more than 11%, even the ratio of 67%. Horizontal transmission of conidia in aggregations may have been limited by an increase in anti-fungal secretions of insects (Ulrich et al. 2015). In addition, some studies have emphasized the importance of mating activity for horizontal transmission (Srei et al. 2020). Because our study was conducted on overwintering adults, mating did not occur as well horizontal transmission.
Conclusions
The virulence of twelve EPF belonging to the genera Beauveria, Metarhizium and Isaria was evaluated on the adults of L. occidentalis. M. flavoviride As18 was determined to be the most virulent and fast killing isolate for microbial control of L. occidentalis. This is the first study to investigate the efficacy of M. flavoviride on L. occidentalis adults. Based on the results of the present study, M. flavoviride isolate As18 could prove to be a suitable microbial control agent that could be used as part of an IPM strategy of L. occidentalis. Further studies should be conducted to validate these results under field conditions.
Availability of data and materials
All data generated and analyzed during this study are indicated in the manuscript.
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J Econ Entomol 18(2):265–267
Allen RC (1969) revision of the genus Leptoglossus Guerin (Hemiptera: Coreidae). Entomol Am 45:35–140
Barta M (2010) Preliminary evaluation of insect-pathogenic Hypocreales against Leptoglossus occidentalis (Heteroptera: Coreidae) in laboratory conditions. Folia Oecol 37(2):137
Ben Jamâa ML, Mejri M, Naves P, Sousa E (2013) Detection of Leptoglossus occidentalis Heidemann, 1910 (Heteroptera: Coreidae) in Tunisia. Afr Entomol 21(1):165–167
Biryol S, Efe D, Eski A, Demirbağ Z, Demir I (2020) Fungal pathogens of Amphimallon solstitiale Linnaeus, 1758 (Coleoptera: Scarabaeidae). Turk J Entomol 44(3):375–384
Biryol S, Araz N, Eski A, Aktürk R, Aksu Y, Çelik Göktürk B, Demir I (2021a) Biodiversity and pathogenicity of entomopathogenic fungi associated with the lesser spruce sawfly Pristiphora abietina. Entomol Exp Appl 169(5):414–423
Biryol S, Güney E, Eski A, Bayramoğlu Z, Sezen K, Demirbag Z, Demir İ (2021b) Development of mycoinsecticide formulations with Beauveria bassiana and Metarhizium brunneum for the control of Orosanga japonica (Hemiptera: Ricaniidae). Ann Appl Biol 179(3):319–330
Biryol S, Demirbağ Z, Erdoğan P, Demir İ (2022) Development of Beauveria bassiana (Ascomycota: Hypocreales) as a mycoinsecticide to control green peach aphid, Myzus persicae (Homoptera: Aphididae) and investigation of its biocontrol potential. J Asia Pac Entomol 25:101878
El Khoury Y, Noujeim E, Bubici G, Tarasco E, Al Khoury C, Nemer N (2021) Potential factors behind the decline of Pinus pinea nut production in Mediterranean pine forests. Forests 12:1167
Farinha AO, Carvalho C, Correia AC, Branco M (2021) Impact assessment of Leptoglossus occidentalis in Pinus pinea: Integrating population density and seed loss. For Ecol Manag 496:119422
Fent M, Kment P (2011) First record of the invasive western conifer seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Turkey. North West J Zool 7(1):72–80
Gapon DA (2013) First records of the western conifer seed bug Leptoglossus occidentalis Heid (Heteroptera, Coreidae) from Russia and Ukraine, regularities in its distribution and possibilities of its range expansion in the Palaearctic region. Entomol Rev 93(2):174–181
Ishikawa T, Kikuhara Y (2009) Leptoglossus occidentalis Heidemann (Hemiptera: Coreidae), a presumable recent invader to Japan. Jpn J Entomol 12(3):115–116
Khun KK, Ash GJ, Stevens MM, Huwer RK, Wilson BA (2021) Transmission of Metarhizium anisopliae and Beauveria bassiana to adults of Kuschelorhynchus macadamiae (Coleoptera: Curculionidae) from infected adults and conidiated cadavers. Sci Rep 11(1):1–12
Mbata GN, Shapiro-Ilan D (2013) The potential for controlling Pangaeus bilineatus (Heteroptera: Cydnidae) using a combination of entomopathogens and an insecticide. J Econ Entomol 106(5):2072–2076
Parlak S (2017) An invasive species: Leptoglossus occidentalis (Heidemann) how does it affect forestry activities? Kastamonu Univ J Fac 17(3):531–542
Rumine P, Barzanti GP (2008) Controllo microbiologico della “cimice delle conifer”: prove preliminari di laboratorio. ATTI Gior Fitopatolog 1:307–308
Sevim A, Demir I, Demirbağ Z (2010a) Molecular characterization and virulence of Beauveria spp. from the Pine processionary moth, Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae). Mycopathologia 170:269–277
Sevim A, Demir I, Höfte M, Humber RA, Demirbağ Z (2010b) Isolation and characterization of entomopathogenic fungi from hazelnut-growing region of Turkey. Biocontrol 55:279–297
Sevim A, Sevim E, Demir İ, Demirbağ Z (2014) Molecular characterization and pathogenicity of Beauveria bassiana isolated from Rhynchites bacchus L. (Coleoptera: Rhynchitidae). Nevsehir J Sci Tech 3(2):33–47
Sonmez E, Uzunoğlu H, Eski A, Demirbağ Z, Demir İ (2022) Stability of Metarhizium anisopliae (Hypocreales: Clavicitaceae) isolates during repeated in vitro subculture and evaluation of an oil-in-water mycoinsecticide. Can Entomol 154(e26):1–16
Srei N, Lavallée R, Guertin C (2020) Horizontal transmission of the entomopathogenic fungal isolate INRS-242 of Beauveria bassiana (Hypocreales: Cordycipitaceae) in emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae). J Econ Entomol 113(1):543–545
Ulrich KR, Feldlaufer MF, Kramer M, St Leger RJ (2015) Inhibition of the entomopathogenic fungus Metarhizium anisopliae sensu lato in vitro by the bed bug defensive secretions (E)-2-hexenal and (E)-2-octenal. Biocontrol 60(4):517–526
Yucel B, Gozuacik C, Gencer D, Demir I, Demirbag Z (2018) Determination of fungal pathogens of Hypera postica (Gyllenhall) (Coleoptera: Curculionidae): isolation, characterization, and susceptibility. Egypt J Biol Pest Control 28(1):1–8
Zhang S, Xia YX, Kim B, Keyhani NO (2011) Two hydrophobins are involved in fungal spore coat rodlet layer assembly and each play distinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol Microbiol 80(3):811–826
Zhu WB (2010) Exotic coreid bugs introduced into China, In: Proceedings of the 4th meeting of the International Heteropterist's Society (Tianjin: Nankai University)
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
ID and SB designed the study. AE and OA performed the study. ID and AE took part in writing original draft. SB and OA involved in review and editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Eski, A., Biryol, S., Acici, O. et al. Biocontrol of the western conifer seed bug, Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) using indigenous entomopathogenic fungi. Egypt J Biol Pest Control 32, 140 (2022). https://doi.org/10.1186/s41938-022-00641-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00641-4