
Abstract
Background
The flower thrips, Frankliniella schultzei Trybom (Thysanoptera: Thripidae) is a notorious pest that attacks the plants of economic importance. The anthocorid bug Blaptostethus pallescens Poppius (Heteroptera: Anthocoridae) is a predator of thrips in field crops and greenhouses. Another biocontrol agent, the entomopathogenic fungi (EPF) Lecanicillium lecanii (Zimm.) (Hypocreales: Cordycipitaceae), has been effectively used in management of various insect pests. With the aim to develop an effective combination of biocontrol agents like microbial and insect predator for the management of the serious pest F. schultzei, the present studies were carried out on development, predation rate and prey preferences of the predator B. pallescens provided with the EPF (ICAR-NBAIR Vl-8)-treated thrips under laboratory conditions.
Results
The predator, B. pallescens, could complete its life cycle on treated thrips. The nymphal duration of bugs fed on treated thrips was higher (25.25 ± 0.13 days). The Vl-8 strain did not show any negative effect on hatchability of eggs (83% hatchability in treatment; 86% in control). However, the longevity was comparatively less in bugs fed on treated thrips than that of control group The fecundity of the treated group was found to be reduced to one half of the control group. The rate of daily feeding potential of bugs on treated thrips was less (7.29 ± 0.15) than the untreated thrips (12.54 ± 0.1) throughout their lifetime. Moreover, when the F1 generation from both parents line fed on treated and untreated thrips allowed to feed on Corcyra cephalonica eggs, it did not show any difference in terms of nymphal duration which revealed that the fungi did not affect the F1 generation. When choice was given between treated and untreated thrips to different instars of this predatory bug, all the instars including adult significantly preferred the untreated thrips. However, nymphs and adult bugs were found to encounter both the treated and untreated thrips. No mortality was observed in any stage of the predator.
Conclusion
This study shows that the effect of L. lecanii (ICAR-NBAIR Vl-8) on B. pallescens is not harmful. However, further field studies are required to evaluate their combined effect against this pest.
Similar content being viewed by others

Background
The flower thrips, Frankliniella schultzei (Trybom), is a polyphagous insect pest and has ability to feed on alternative food source. It is a severe pest of cucumber, bean, snap bean, pepper, corns (Pereira et al. 2017). It is also called as common blossom thrips and distributed in the subtropical as well as the tropical regions around the world. In addition to being a serious pest that causes major losses through feeding damage, it also acts as a vector transmitting Tospovirus (Tamíris et al. 2020).
Minute pirate bugs as well as entomopathogenic fungi (EPF) could be alternatives to chemicals to control thrips. The anthocorid predatory bug, Blaptostethus pallescens Poppius (Heteroptera: Anthocoridae), has been found feeding on thrips, lepidopteran insects and mites (Ballal et al. 2018). This bug is suitable for laboratory rearing conditions, and its biological characteristics have already been studied (Ballal et al. 2003). B. pallescens has been found preying upon several pests like eggs of lepidopteran pests like spider mites, woolly aphid infesting cotton and young caterpillars (Ballal et al. 2018). EPFs have got numerous benefits in terms of various insect pest management, and it is safer to environment and could be a replacement for chemical pesticides (Bamisile et al. 2021).
To manage serious pests like thrips, it is desirable to use more than one management practices in a compatible manner. Integration strategies that include insect natural enemies and microbials for the management of pests have potential against insect pests. But then, it is very much warranted to test lethal and sublethal effects of such microbial agents over the natural predators (Croft 1990) to understand compatibility of both the bioagents for better management of insect pest (Najme et al. 2011). These microbials could affect biology, life table parameters, predatory potential of both parental as well as F1 generation. Thus, the present study aimed to study the effect of L. lecanii (strain NBAIR Vl-8) on biology, predation rate and prey preference of the predatory bug B. pallescens when fed on F. schultzei. This study will enhance our understanding and provide information on combined application of these two bioagents to manage the thrips.
Methods
Entomopathogenic fungi culture
The EPF, L. lecanii Vl-8 strain, was grown and maintained in Sabouraud Dextrose Yeast agar (SDYA) Medium at 25 ± 1 °C for 8–10 days. After the growth of pure culture in the plates, the fungal spore culture was inoculated into Sabouraud Dextrose Broth media (SDYB) liquid media. It was then placed in shaker at 25 ± 1 °C and 150 rpm for 8 days. The 8-day-old culture was then mixed with the pre-autoclaved talc in the ratio 1:2 (500 ml fungi grown broth: 1 kg talc). Then, the mixed talc was covered with a black cloth and kept for drying for 3 days under aseptic conditions. Quality analysis of EPF was done by checking the spore count through hemocytometer and serial dilution method. Briefly, 1 g of talc was mixed to 9 ml of water and was vortexed and 1 ml from it was transferred to another vial to make it 10–2. The same method was repeated up to 10–12 dilution. Further 1 ml taken from 10–6, 10–8, 10–10, 10–12 dilutions and transferred to a Petri plate containing SDYA medium. The colony count was observed in 10–8 plates to confirm that the talc contains 10–8 spores. After the spore count was quantified, the suspensions were finally prepared by serial dilution in distilled water by adding 0.1% of Tween-80 and stored in the refrigerator at 4 °C until further use.
Insect culture
Frankliniella schultzei
Adults of F. schultzei were reared on chemical free beans in a plastic container (1 L capacity) with a lid. The lid of the container was cut open and stuck with a fine mesh for aeration. Daily bean pods were collected from the adult container and kept for hatching to get desirable stage.
Blaptostethus pallescens
Adult of B. pallescens was obtained from the stock culture reared at ICAR-NBAIR laboratory, Bengaluru, and subcultured in plastic pearl pet containers of 500 ml capacity. Corcyra cephalonica eggs ad libitum were given as food, and 4–5 pieces of bean pods each of size 5–6 cm were provided as ovipositional substrate in each container which can hold up to 20 adult B. pallescens. Collection of bean pods was done on daily basis to collect the eggs of B. pallescens and kept for hatching. The newly hatched nymphs were reared in plastic containers using the same procedure as followed for adults. Corcyra eggs were replenished on daily basis as food. A small cotton piece soaked in water was introduced into the containers to maintain the moisture.
Biology and feeding potential on Vl-8 treated and untreated thrips
Laboratory studies were carried out at 26 ± 2 °C and 65 ± 2% R.H. Second instar larvae of thrips were taken and sprayed with 1 × 108 spores/ml of Vl-8 fungal strain such that the entire body surface is covered. After 24 h. of spraying, the treated thrips were then counted and provided as feed for the predator. The predator B. pallescens was acclimatized to laboratory conditions for 2 generations on untreated F. schultzei thrips before commencing the experiment. Biology was also studied on untreated thrips.
Thirty newly hatched nymphs of the predator were taken for the experiment. Each nymph was placed individually in plastic containers (90 mm diameter) with tissue paper in the bottom and Vl-8-treated thrips as feed. Two bean pods were introduced in each plastic container for feeding for thrips. Ten number of treated or untreated second instar thrips larvae were provided per nymph initially up to first two instars of predatory bug and were increased to about 30 for late instars and adults because of increase in requirement by the advance stages of predatory bug. Observations on the number of thrips consumed and the time of molting were recorded on daily basis in both the treatments. Thrips were replenished on daily basis to all the containers. The experimental setup was maintained till emergence of adults. The nymphal duration for each individual was recorded. After the adult emergence, the sex ratios were recorded. Further, the adults were left for mating in order to observe the fecundity and per cent hatching. Longevity of male and female predator was also recorded. Morphometrics of each nymphal instars and adults were recorded using LEICA M205C microscope.
F1 generation
The eggs laid by the adult bugs reared on Vl-8 treated or untreated thrips were collected and kept for hatching. Twenty nymphs were taken and reared on UV-treated C. cephalonica eggs to check if there was any adverse or sublethal effect of Vl-8 on nymphal duration of F1.
Prey preference of different instars of predatory bugs
Second, third, fourth and fifth nymphal instars of B. pallescens along with female adults were taken and provided with Vl-8 treated and untreated second instar thrips larvae at the same time. Fifteen treated and fifteen untreated thrips were provided for second and third instar, and the number of thrips was increased to 20 each for fourth instar to adult stage. Experiment setup of each bug was maintained in a large Petri plate and kept for 24 h. The thrips consumed by the bugs were recorded.
Adult preference at different time intervals of thrips infection
Three-day-old adult bugs were offered 20 untreated and 20 Vl-8 treated second instar thrips larvae at different intervals, i.e., 24, 48, 72, 96 h. post-inoculation, and were checked for their preference. The thrips consumed by the bugs were recorded after 24 h.
Statistical analyses
Prey preference of different instars of B. pallescens was analyzed using an independent t test comparing the means of Vl-8 treated and untreated thrips. Manly’s preference index for prey type was calculated using the formula given below:
where β1 is the prey preference toward prey type 1, e1 is the remaining prey after the experiment, and A1 is the number of prey provided initially. The value near to 0.5 reveals that there is no preference toward provided prey. If the value is nearing to 1, it shows the preference toward prey type 1 and value nearing to 0 shows the preference toward prey type 2 (Varshney and Ballal 2018). In this study, untreated thrips were chosen as prey type 1. One-sample t test was used to compare the Manly’s preference index for untreated thrips setting a value 0.5 to test the null hypothesis that predator prefers prey at random.
Results
Biology on Vl-8 treated and untreated thrips
Blaptostethus pallescens was able to complete life cycle feeding on Vl-8-treated thrips. All the morphometric data on mean body length, width, labium, hind femur and tibia of adult B. pallescens when fed on treated thrips did not differ than the adults fed on untreated thrips. Hence, we did not show data in this paper.
The comparative data on biological parameters of B. pallescens fed on Vl-8 treated and untreated thrips are given in Table 1. The data revealed that the nymphal duration was higher (25.25 ± 0.13 days) for B. pallescens reared on Vl-8-treated thrips than the untreated ones (19.56 ± 0.29 days). The longevity of both male and females of the bugs reared on Vl-8-treated thrips were reduced (male: 13.5 ± 2.22; female: 14.2 ± 2.20) than the bugs reared on untreated thrips (male: 38.67 ± 1.86; female: 40.25 ± 0.85). The bugs reared on untreated thrips were able to survive for around 24 days more than the bugs fed on treated thrips. This certainly showed the reduction in the predation capacity of the bugs when reared on treated thrips. The fecundity and nymphs per female of insects reared on treated thrips were reduced to one half of the control insects.
However, non-significant difference was observed in percent hatchability of the bugs fed on treated thrips (83%) and bugs fed on untreated thrips (86%). The sex ratio revealed that the female emergence was higher in both the treatments.
Feeding potential
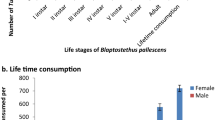
Feeding potential data of the B. pallescens revealed non-significant difference in per day and total feeding for the 1st instar of predatory bug when allowed to feed on Vl-8 treated and untreated thrips. A difference in feeding potential was observed from the second instar to fifth instar resulting in difference in total nymphal feeding potential being 103.5 ± 1.42 in nymphal duration of 25 days and 124.33 ± 1.94 in 19 days for Vl-8 treated and untreated thrips, respectively. The average daily feeding rate of the adult bug on treated thrips was 7.29 ± 0.15 which was significantly lower than the control (12.54 ± 0.1). The total adult feeding on treated thrips was 51 ± 1.03 for one week and on untreated thrips, it was 100.33 ± 0.91 (Fig. 1).

Feeding potential of different instars of Blaptostethus pallescens fed on Lecanicillium lecanii (Vl-8) treated and untreated Frankliniella schultzei
F1 generation
The nymphal duration of F1 generation from both treatments where B. pallescens was reared on Vl-8 treated thrips and untreated thrips was observed. There was non-significant difference in nymphal durations in both the treatments. Nymphs from both treatments were able to complete nymphal duration in 18 days (Table 2).
Prey preference of different instars
Data in Table 3 show the prey preference data of B. pallescens on Vl-8 treated thrips and untreated thrips recorded after 24 h. of inoculation. The predator was found to encounter both groups, but it preferred the untreated thrips. In choice test, second (t = 3.130, df = 4.927, p = 0.026), third (t = − 5.892, df = 5.880, p = 0.001), fourth (t = − 3.667, df = 5.146, p = 0.014), fifth (t = − 12.247, df = 6.00, p < 0.0001) instar nymphs and adult female (t = − 7.155, df = 5.534, p < 0.001) preferred untreated thrips. It was further confirmed by Manley’s preference index β 1 for each nymphal instar and adult B. pallescens (Table 3).
Prey preference at different time interval of infection
Comparative data of prey preference of adult B. pallescens on Vl-8-treated thrips at different post-inoculation period (24–96 h) and untreated thrips showed that at each post-inoculation period, significant preference was observed for untreated thrips (24 h: t = 5.73 P < 0.0001; 48 h: t = 9.86 P < 0.0001; 72 h: t = 11.77 P < 0.0001; 96 h: t = 19.5 P < 0.0001). As the time of post-inoculation increased, adult predator consumed more number of untreated thrips. Very low feeding was observed on treated thrips after 48 h of post-inoculation.
Discussion
The present study revealed that there was no mortality in any stage of B. pallescens when fed on Vl-8-treated thrips and predatory bugs were successfully abled to complete their life cycle on treated thrips. This finding is similar to Broza et al. (2001) who found that collembolans were not susceptible to Metarhizium anisopliae, Verticillium lecanii, Beauveria bassiana, Hirsutella spp., and its endotoxins. Longevity of the B. pallescens bugs was reduced when fed on treated prey compared to the control group. Similar observations were recorded by Liu et al. (2019) who found the reduction of longevity and predation rate of predatory mite Amblydromalus limonicus Garman when treated with EPF Beauveria bassiana (Balsamo). Feeding potential data of B. pallescens on Vl-8-treated thrips revealed that there was a reduction in their feeding potential than the control group. This finding is corroborated with the study of Pourian et al (2011) who observed reduction in feeding, searching and predation of Orius albidipennis (Reuter) when fed with Metarhizium anisopliae treated prey. Trizelia et al. (2017) found the reduction in the predation ability of Menochilus sexmaculatus when EPF-sprayed aphids were provided. Such reduction in the feeding potential of B. pallescens might be the cause of elongated nymphal duration in the bugs fed on infected prey due to lack of enough nutritional requirement for growth. Reduced feeding on infected prey might hamper pest control and needs field study. However, it is not uncommon in field situation that general predator like B. pallescens can switch the prey or might prefer uninfected prey.
The present study clearly showed that there was no influence of the EPF (Vl-8) on the percent egg hatching of the bugs, but reduction in fecundity was observed. Sayed et al. (2021) also found that the egg hatching of the coccinellids, Coccinella undecimpunctata and Hippodamia variegate, was not affected by Beauveria bassiana. Similar to present study, fecundity of Cryptolaemus montrouzieri Mulsant was affected by both B. bassiana and M. anisopliae (Mohammed 2019). On the contrary, Nielsen et al. (2005) reported non-significant difference in the fecundity of Spalangia cameroni Perkins, a pupal parasitoid when infected with Metarhizium anisopliae. The nymphal duration of F1 generation of B. pallescens obtained from the parents fed on Vl-8-treated thrips was similar to that of F1 generation obtained from the control parents fed on untreated thrips. Similarly, Liu et al. (2019) observed no influence of B. bassiana on the fecundity of F1 generation when compared to the uninfected control group. It is therefore clearly evident that the trans-generational effect of EPF was not carried forward to the progeny of infected parents (Midthassel et al. 2016). When choice was given between Vl-8 treated and untreated thrips to the different instars of B. pallescens, they significantly preferred the untreated prey. Similar results were documented in the study of Meyling and Pell (2006) where they found that the males and females of Anthocoris nemorum L detected and avoided the contact with B. bassiana inoculated leaf. In present study, the adult bugs preferred the untreated thrips to that of Vl-8-treated thrips in all tests. Almost nil feeding was observed when fed on treated thrips at 96 h. post-inoculation. This may be due to the bug’s ability to detect infected prey particularly when mycelia growth develops on prey and when it comes in contact to them. Similar observations were made for Dicyphus hesperus bug which did not prefer whitefly nymphs treated with Isaria fumosorosea five days prior (Alma et al. 2010).
Conclusion
The study showed no mortality in any of the stage of predator when Vl-8-treated thrips were provided for feeding. However, fecundity and longevity were affected. But absolutely no adverse effect was observed on developmental period and feeding potential of nymphs in F1 generation which shows that trans-generational effect of this fungi (Vl-8) was not carried over to F1 generation. Furthermore, B. pallescens always preferred untreated thrips over Vl-8-treated thrips. In field, there is less chance of B. pallescens encountering treated thrips/prey. Greenhouse and field experiments need to be conducted to ascertain the compatibility between B. pallescens and L. lecanii (Vl-8) and their role in management of thrips.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the first author/corresponding author on reasonable request.
Abbreviations
- EPF:
-
Entomopathogenic fungi
- ICAR:
-
Indian Council of Agricultural Research
- NBAIR:
-
National Bureau of Agricultural Insect Resources, Bengaluru
References
Alma CR, Gillespie DR, Roitberg BD, Goettel MS (2010) ‘Threat of infection and threat-avoidance behavior in the predator Dicyphus hesperus feeding on whitefly nymphs infected with an Entomopathogen. J Insect Behav 23:90–99
Ballal CR, Singh SP, Poorani J, Gupta T (2003) Biology and rearing requirements of an anthocorid predator, Blaptostethus pallescens Poppius (Heteroptera: Anthocoridae). J Biol Control 17(1):29–33
Ballal CR, Akbar S, Yamada K, Wachkoo A, Varshney R (2018) Annotated catalogue of the flower bugs from India (Heteroptera: Anthocoridae; Lasiochilidae). Acta Entomol Musei Nationalis Pragae 58:207–226
Bamisile BS, Akutse KS, Siddiqui A, Junaid Ali A, Yijuan X (2021) Model application of entomopathogenic fungi as alternatives to chemical pesticides: prospects, challenges, and insights for next-generation sustainable agriculture. J Front Plant Sci. https://doi.org/10.3389/fpls.2021.741804
Broza M, Pereira RM, Stimac JL (2001) The non-susceptibility of soil Collembola to insect pathogens and their potential as scavengers of microbial pesticides. Pedobiologia 45:523
Croft AB (1990) Arthropod biological control agents and pesticides. Wiley, New York, p 723
Liu J, Zhang Z-Q, Beggs JR, Wei X-Y (2019) Influence of pathogenic fungi on the life history and predation rate of mites attacking a psyllid pest. J Ecotoxicol Environ Saf 183:109585
Meyling NV, Pell JK (2006) Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol Entomol 31(2):162–171. https://doi.org/10.1111/j.0307-6946.2006.00781.x
Midthassel A, Leather SR, Wright DJ, Baxter IH (2016) Compatibility of Amblyseius swirskii with Beauveria bassiana: two potentially complimentary biocontrol agents. Biocontrol 61:437–447
Mohamed GS (2019) The virulence of the entomopathogenic fungi on the predatory species, Cryptolaemus montrouzieri Mulsant (Coleoptera: Coccinellidae) under laboratory conditions. Egypt J Biol Pest Control 29:42
Najme A, Kamal A, Imani S, Takalloozadeh H (2011) Effects of some pesticides on biological parameters and predation of the predaceous plant bug Deraeocoris lutescens (Hemiptera: Miridae). Adv Environ Biol 5(10):3219
Nielsen C, Skovgård H, Steenberg T (2005) Effect of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) on survival and reproduction of the filth fly parasitoid, Spalangia cameroni (Hymenoptera: Pteromalidae). Environ Entomol 34(1):133–139. https://doi.org/10.1603/0046-225X-34.1.133
Pereira PS, Sarmento RA, Galdino TV, Lima CH, Dos Santos FA, Silva J, dos Santos GR, Picanço MC (2017) Economic injury levels and sequential sampling plans for Frankliniella schultzei in watermelon crops. Pest Manage Sci 73:1438–1445
Pourian HR, Talaei-Hassanloui R, Kosari AA, Ashouri A (2011) Effects of Metarhizium anisopliae on searching, feeding and predation by Orius albidipennis (Hem., Anthocoridae) on Thrips tabaci (Thy., Thripidae) larvae. Biocontrol Sci Technol 21:15–21
Sayed S, Elarrnaouty SA, AlOtaibi S, Salah M (2021) Pathogenicity and side effect of indigenous Beauveria bassiana on Coccinella undecimpunctata and Hippodamia variegata (Coleoptera: Coccinellidae). Insects 12(1):42
Tamíris AA, Daniela TP, Rodrigo RS, Marcelo CP, Cristina SB, Thomas EH, William DH (2020) Development and validation of sampling plans for Frankliniella schultzei on tomato. Crop Prot 134:105163
Trizelia T, Busniah M, Permadi A (2017) Pathogenicity of entomopathogenic fungus Metarhizium spp. against predators Menochilus sexmaculatus Fabricius (Coleoptera: Coccinellidae). Asian J Agric 1:1–5
Varshney R, Ballal CR (2018) Intraguild predation on Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) by the generalist predator Geocoris ochropterus Fieber (Hemiptera: Geocoridae). Egypt J Biol Pest Control. 28:5
Acknowledgements
The authors are grateful to Indian Council of Agricultural Research, New Delhi, India, and Director, ICAR−NBAIR, Bengaluru, India, for providing research facilities and encouragement. We are also thankful to Mrs. Usha Ravikumar for technical assistance.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
RV conceived the study and planned experiment. KS performed experiments and analyzed the data under the guidance of RV. AK formulated the entomopathogenic fungi. KS wrote the manuscript. RV and KR reviewed manuscript. All the authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This manuscript does not contain any studies with human participants or animals performed by any of the authors.
Consent for publication
Not applicable.
Competing interests
The authors have not found any potential conflicts of interest, and all ethical aspects are considered.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sundaravalli, K., Varshney, R., Kandan, A. et al. Effect of the entomopathogenic fungus, Lecanicillium lecanii, on the biology and predation rate of the anthocorid predatory bug, Blaptostethus pallescens, feeding on the flower thrips, Frankliniella schultzei Trybom (Thysanoptera: Thripidae). Egypt J Biol Pest Control 32, 142 (2022). https://doi.org/10.1186/s41938-022-00634-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00634-3