
Abstract
Background
Entomopathogenic fungi (EPFs) have more important role in biological control of the important insect pests. In the present study 15 EPF isolates, selected from the EPF culture collection, were identified using molecular methods and their effectiveness against adults of Apple Blossom Beetle (ABB) [Tropinota (= Epicometis) hirta (Poda, 1761) (Coleoptera: Cetoniidae)] was investigated. To determine virulence of isolates against ABB adults, primarily a single-concentration trial was conducted at 1 × 107 conidia ml−1. Furthermore, concentration-mortality trials were conducted with some isolates that were determined to have a high effect, employing concentrations of 1 × 106, 1 × 108, and 1 × 109 conidia ml−1. For isolates tested in concentration-mortality trials, LT30, LT50 and LT90 values at a concentration of 1 × 108 conidia ml−1 were calculated using probit analysis.
Results
Molecular diagnostic tests revealed that the 13 isolates were Beauveria bassiana, 1 isolate was Clonostachys rosea, and 1 isolate was Purpureocillium lilacinum. According to the single- concentration trial data, some isolates with high efficacy were selected (GN16, GN22-1, GN29-1, HP3-1 and HP33-1) and concentration-mortality trials were established with these isolates using 1 × 106, 1 × 108 and 1 × 109 conidia ml−1 concentrations. According to the results of the concentration-mortality trials, all isolates used in 1 × 108 and 1 × 109 conidia ml−1 concentrations started to show a significant effect from the 11th day, and at the end of the 17th day, the effect in all concentrations was over 90% (except 1 × 106 conidia ml−1 concentration of GN22-1).
Conclusions
According to the data obtained, all isolates used in concentration-mortality trials were effective against ABB adults. With these isolates, more detailed studies should be conducted under orchard conditions.
Similar content being viewed by others


Background
There are many factors that limit fruit production. Among them, diseases, pests and weeds have an important place. When these factors are not managed properly, significant economic losses occur. Apple Blossom Beetle (ABB) Tropinota (= Epicometis) hirta (Poda, 1761) (Coleoptera: Cetoniidae), is one of the fruit pests (Yaşar and Dahham 2019). It is an important species that spreads to across the Palaearctic region and to almost all regions of Turkey (Aydın 2011). In the spring, ABB adults cause significant damage by feeding on different parts of the flowers of the plants, especially fruit trees, strawberries and roses (Aslan and Aslan 2015). They live on leaves during non-flowering periods (Atanasova et al. 2017). Kutinkova and Andreev (2004) reported that the ABB is one of the most important pests of cherry trees in Bulgaria that causes up to 70% damage by feeding on the flowers of young cherry trees.
Chemical control against ABB adults is not recommended due to the presence of pollinators and beneficial insects in the orchards during the flowering period of fruit trees. As a result, eco-friendly control strategies are needed to limit the damage caused by ABB while also protecting the non-target species (Yaşar and Dahham 2019). Biological control with entomopathogenic microorganisms is one of the most effective non-chemical pest control approaches (Mantzoukas and Eliopoulos 2020). EPFs are common in terrestrial environments and serve a crucial role in insect population control (Khan et al. 2012). Approximately 750 EPF species have been found to infect a wide variety of insects and mites (Inglis et al. 2001). Strains of EPFs are concentrated in Hypocreales (various genera), Onygenales (Ascosphaera genus), Entomophthorales, and Neozygitales (Entomophthoromycota) (Mora et al. 2017). These biocontrol agents are the most important of all microbial biocontrol agents because they can be mass-produced easily, quickly, and cheaply, and they can be utilized with the same technological methods as traditional contact insecticides. Many commercial mycoinsecticides are being developed from EPFs however most of these products are based in only four genera of entomopathogenic fungi: Beauveria, Metarhizium, Lecanicillium and Isaria (Vega et al. 2009).
In this study, EPF isolates selected from the culture collection in Mycology Laboratory (Tokat Gaziosmanpaşa University, Faculty of Agriculture, Plant Protection Department) were identified using morphological and molecular methods and their virulence against adults of ABB was investigated under laboratory conditions.
Methods
Insect sources
ABB adults were collected from apple orchards in the research area of Tokat Gaziosmanpaşa University, Faculty of Agriculture (N40°20′01′′ E36°28′27′′, 622 m), in Tokat-Central-Büyükyıldız (N40°20′11′′ E36°23′27′′, 606 m) and in Tokat-Central-Akyamaç (N40°20′47′′ E36°29′53′′, 615 m). The beetles were kept in plastic containers, covered with fine mesh for aeration until used in the bioassay.
Fungal isolates
Fifteen EPF isolates were chosen for the current investigation from the fungal culture collection of the Tokat Gaziosmanpasa University, Faculty of Agriculture, Department of Plant Protection in Tokat, Turkey. These isolates were initially isolated from Hypera postica (Gyllenhal 1813) (Coleoptera, Curculionidae) and Gonioctena fornicata (Brüggemann 1873) (Coleoptera, Chrysomelidae) adults gathered from alfalfa fields in Tokat Province, Turkey (Baysal 2017).
Morphological characterization
Fifteen isolates were preliminarily identified using morphological characteristics such as appearance of the fungal infection, colony shape, spore size and spores shape (Humber 1997).
Molecular characterization
DNA isolation and PCR studies
DNA isolation of the EPF isolates was performed according to the "Genomic DNA Purification Kit Thermo Fisher Scientific" method. ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) as the forward primer and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) as reverse primers were used for ITS PCR-amplification (White et al. 1990). For PCR, 1 µl template DNA, 2.5 µl 10X Taq Buffer, 0.2 µl dNTP (25 mM), 0.5 µl ITS5 primer (100 pmol), 0.5 µl ITS4 primer (100 pmol), 1.5 µl MgCl2 (25 mM), 0.25 µl Taq polymerase enzyme (Thermo) and 1 µl of Dimethylsulfoxside (DMSO) were mixed in PCR tubes and made up to 25 µl with distilled water and placed in the termocycler (Techne Prime). Thirty-five cycles were performed in thermocycler following the denaturation at 95 °C for 5 min, 95 °C for 1 min, 55 °C for 55 s, 72 °C for 2 min, with a final extension at 72 °C for 10 min (Sevim et al. 2014). The PCR products obtained as a result of PCR were subjected to electrophoresis at 100 V for 1 h in an agarose gel containing 10 mg/ml ethidium bromide, prepared at a rate of 1.2% and visualized under the imaging device (Sevim et al. 2014).
Phylogenetic analysis
For phylogenetic studies, the products obtained at the end of PCR were sent to Atlas Biotechnology (Ankara-Turkey) for the sequencing. The data obtained at the end of the sequencing were analysed with the MEGAX (Kumar et al. 2018) computer program. The data were then compared with reference isolates registered in the National Center for Biotechnology Information (NCBI) gene bank, and molecular identification of the isolates was made.
Inoculum preparation from entomopathogenic fungal isolates
Fungal isolates were cultured on Potato Dextrose Agar (PDA) in Petri dishes. A small part of mycelia was taken from each fungal isolates and inoculated in of PDA and incubated at 25 ± 2 °C with a 16/8 (L/D) photoperiod. At the end of the 21-day incubation period, spores were harvested with 10 ml of sterilized water containing 0.02% Tween 80. The conidial suspensions were filtered through 3 layers of sterile muslin to remove particles and then conidial concentration adjusted to 1 × 106, 1 × 107, 1 × 108 and 1 × 109 conidia ml−1 (Saruhan et al. 2017).
Bioassays
To determine virulence of isolates against ABB adults, primarily a single- concentration trial was conducted at 1 × 107 conidia ml−1. To test the effect of each of the isolates, ABB adults were immersed into 1 × 107 conidia ml−1 suspension of each isolate for 10 s. and transferred into a Petri dish (5 adults per plate) containing fresh apple leaf and flower. The control group was treated with sterile water including 0.02% Tween 80. Mortality of the adults was assessed on the 3rd, 5th, 7th, 9th, 13th, 15th and 17th days of incubation periods. Furthermore, concentration-mortality trials were conducted similar to single-concentration trials. These tests were performed with some isolates that were determined to have a high effect, employing concentrations of 1 × 106, 1 × 108, and 1 × 109 conidia ml−1. A completely randomized block design with 5 replications was used for the experiments and replicated 2 times.
Statistical analysis
Data was analysed with analysis of variance (ANOVA) and the means were compared to Tukey's multiple comparison test by using the MINITAB Release 16 packet program. To determine the statistical interactions between treatments, MINITAB Release 16 was used with a general linear model. For isolates tested in concentration-mortality trials, LT30, LT50 and LT90 values at a concentration of 1 × 108 conidia ml−1 were calculated using probit analysis.
Results
Molecular identification of the fungi
The 15 isolates used in the study were identified by molecular methods, and 13 isolates were Beauveria bassiana, 1 isolate was Clonostachys rosae and 1 isolate was Purpureocillium lilacinum (Table 1; Fig. 1).

Phylogenetic tree constructed based on ITS region of the obtained isolates and references using the Neighbor-Joining method. The isolates obtained in this study are shown in blue. Obtained Beauveria bassiana, Purpureocillium lilacinum and Clonostachys rosea isolates are indicated using a black-filled circle, black-filled square, black-filled triangle respectively
Virulence of the fungi
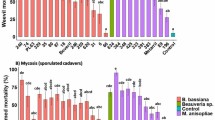
According to the results of the single- concentration trial, all isolates, except HP36-3(1), HP5-2, HP22, HP14 and HP19, had an effect of more than 60% against ABB at the end of the 17th day. Concentration-mortality trials were carried out using concentrations of 1 × 106, 1 × 108 and 1 × 109 conidia ml−1 with some of the effective isolates (GN29-1, HP3-1, GN16, HP33-1, and GN22-1 isolates of B. bassiana) (Table 2).
Based on the results of concentration-death tests, the mortality rates caused by EPF isolates varied depending to period, isolates, and concentration. EPFs concentration, exposure time, and mortality all had a positive relationship. The GN16 isolate started to show a remarkable effect (68.5%) at a concentration of 1 × 109 conidia ml−1 after 7 days post-inoculation. This effect reached 99% after 11 days (F: 133.94; DF: 8.81; P < 0.05). At the end of the incubation period, the effectiveness was found above 90% at all concentrations (F: 265.18; DF: 3.36; P < 0.05) (Table 3). GN22-1 isolate reached 50% effect only at the end of the 9th day at a concentration of 1 × 109 conidia ml−1. It was able to show 100% efficiency after 17 days exposure at same concentration (F: 103.74; DF: 8.81; P < 0.05) (Table 4). Similar to the GN16 isolate, GN29-1 caused a mortality rate of over 60% at a concentration of 1 × 109 conidia ml−1 after seven days post-inoculation and this effect was 100% at the end of the 11th day (F: 157.38; DF: 8.81; P < 0.05) (Table 5). HP3-1 isolate caused more than 50% mortality from the end of the 9th day at 1 × 108 and 109 conidia ml−1 concentrations (F: 209.61; DF: 3.36; P < 0.05). At the end of the 17th day exposure, all concentrations, including the lowest concentration, produced an effect of almost 100% (F: 622.47; DF: 3.36; P < 0.05) (Table 6). In HP33-1 isolate, 58% mortality was determined at a concentration of 1 × 109 conidia ml−1 at the end of 7th day exposure time, this rate reached 100% at the end of the 13th day (F: 208.19; DF: 8.81; P < 0.05). Similar to the HP3-1 isolate, after seventeen days post-inoculation, all concentrations, caused an effect of almost 100% (F: 622.47; DF: 3.36; P < 0.05) (Table 7).
A complete ANOVA analysis was performed to determine the effects of isolate, concentration, and time for ABB. Isolate*concentration, isolate*time, concentration*time, isolate* concentration*time interactions were statistically significant (Table 8).
When the values were examined, it was seen that GN29-1 isolate caused the fastest effect in all LT rates among the isolates applied against ABB (LT30: 5.897, LT50: 6.817, LT90: 9.715). HP33-1 isolate is the second isolate that causes the fastest death in respect of LT30 (6.722) and LT50 (8.060) rates. HP3-1 isolate was found to be the second isolate causing the fastest mortality in terms of LT90 (12.333) rate. The slowest effect in all LT values was determined in GN22-1 isolate (LT30: 7.651, LT50: 9.416, LT90: 15.638) (Table 9).
Discussion
The results showed that all B. bassiana EPF isolates used in concentration-mortality trials were significantly effective at 1 × 108 and 1 × 109 conidia ml−1 concentrations from the 11th day. This effect increased above 80% in all isolates by the end of the 13th day. 17 days after post-inoculation, it was observed that the effect exceeded 90%, including the lowest concentration (except 1 × 106 conidia ml−1 concentration of GN22-1). Only one investigation had reported the susceptibility of ABB to EPFs. In that study, similar to the present, it was reported that the effectiveness of B. bassiana and Fusarium sp. against ABB adults at 1 × 106, 1 × 107 and 1 × 108 conidia ml−1 concentrations was found as 70, 90 and 90% for B. bassiana and 50, 70 and 75% for Fusarium sp., respectively on the 14th day (Atmaca et al. 2018).
According to the results of the single concentration trial, the effects of HP36-3(1), HP5-2, HP22, HP14 and HP19 isolates against ABB at the end of the 17th day did not exceed 40%. The virulence of isolates obtained from the same fungal species may differ from each other depending on some factors such as toxins produced by pathogens (Wang 2021).
Since native isolates can be ecologically compatible with the pest species, their use against the target pest always has advantages in comparison to exotic isolates (Sutanto et al. 2021). All of the isolates used in concentration-mortality studies were native isolates and showed a significant effect against ABB. In addition, ABB spends its larval and pupal stages in the soil and overwinters in the soil as adults (Kara 1995). Many hypomycetous fungal entomopathogens are considered soil-borne microorganisms and have significant potential in the control of pests in the soil (Keller and Zimmermann 1989).
Conclusions
All of the isolates (GN16, GN22-1, GN29-1, HP3-1 and HP33-1) used in concentration-mortality studies showed significant activity against ABB adults. The present study showed that these EPFs can be used as a bio-control agent against adults of ABB. Further studies should be conducted under orchard conditions.
Availability of data and materials
The dataset(s) supporting the conclusions of this article is (are) included within the article (and its additional file(s)).
Abbreviations
- ABB:
-
Apple Blossom Beetle
- EPFs:
-
Entomopathogenetic fungi
- PDA:
-
Potato Dextrose Agar
- LT:
-
Lethal time
References
Aslan ÖM, Aslan MM (2015) The activity of catching Epicometis hirta (Poda, 1761) (Coleoptera: Scarabaeidae) with different trap types on almond trees in Kahramanmaraş province. KSÜ Doğa Bil Derg 18(4):6–12. https://doi.org/10.18016/ksujns.83030
Atanasova D, Ganchev D, Nenov N (2017) In vitro screening for insecticidal activity of natural plant protection products against Tropinota (Epicometis) hirta (Poda)(Coleoptera; Cetoniidae). Agric Sci 9(21):47–51. https://doi.org/10.22620/agrisci.2017.21.007
Atmaca S, Pekbey G, Canhilal R (2018) Virulence of some entomopathogenic fungi against adult of Blossom Beetle, Epicometis hirta (Poda) (Coleoptera: Scarabaeidae). Iğdır Univ J Inst Sci Tech 8(2):37–42
Aydın G (2011) Plant phenology-related shifts in color preferences of Epicometis (Tropinota) hirta (Coleoptera: Scarabaeidae: Cetoniinae) adults-key to effective population monitoring and suppression. Fla Entomol 94(4):832–838
Baysal E (2017) Bazı Entomopatojen Fungus İzolatlarının, Yonca Yaprak Böceği [Gonioctena fornicata (Coleoptera: Chrysomelidae)] ve Yonca Hortumlu Böceği [Hypera postica) (Coleoptera: Curculionidae)]’na Karşı Etkinliklerinin Araştırılması. M.Sc. Thesis, Tokat Gaziosmanpaşa Üniversity, Tokat, Turkey
Humber RA (1997) Fungi-identification. In: Lacey LA (ed) Manual of techniques in insect pathology. Academic Press, London, pp 153–185
Inglis GD, Goettel MS, Butt TM, Strasser H (2001) Use of hyphomycetous fungi for managing insect pests. In: Butt TM, Jackson C, Magan N (eds) Fungi as biocontrol agents. CABI International, Wallingford, pp 23–69
Kara K (1995) Tropinota (Epicometis) hirta (Poda) (Coleoptera: Scarebeidae)’nın Tokat ve çevresindeki konukçuları, yayılışı, zarar düzeyi, bazı biyolojik özellikleri üzerine araştırmalar. Gaziosmanpaşa Üniversitesi Ziraat Fakültesi Dergisi 12:15–26
Keller S, Zimmermann G (1989) Mycopathogens of soil insects. In: Wilding N, Collins NM, Hammond PM, Webber JF (eds) Insect–fungus interactions. Academic Press, London, pp 239–270
Khan S, Guo L, Maimaiti Y, Mijit M, Qiu D (2012) Entomopathogenic fungi as microbial biocontrol agent. Mol Plant Breed 3(7):63–79. https://doi.org/10.5376/mpb.2012.03.0007
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35(6):1547–1549. https://doi.org/10.1093/molbev/msy096
Kutinkova H, Andreev R (2004) Integrated pest management in sweetcherry (Prunus avium L.) orchards in Bulgaria. J Fruit Ornam Plant Res 12:41–47
Mantzoukas S, Eliopoulos PA (2020) Endophytic entomopathogenic fungi: a valuable biological control tool against plant pests. Appl Sci 10(360):1–13. https://doi.org/10.3390/app10010360
Mora MAE, Castilho AMC, Fraga ME (2017) Classification and infection mechanism of entomopathogenic fungi. Arq Inst Biol 84:1–10. https://doi.org/10.1590/1808-1657000552015
Saruhan I, Toksöz Ş, Erper I (2017) Evaluation of some entomopathogenic fungi against the fall webworm (Hyphantria cunea Durry, Lepidoptera: Arctidae). Selcuk J Agr Food Sci 31:76–81. https://doi.org/10.15316/SJAFS.2017.22
Sevim A, Sevim E, Demir İ, Demirbağ Z (2014) Molecular characterization and pathogenicity of Beauveria bassiana isolated from Rhynchites bacchus L. (Coleoptera: Rhynchitidae). Nevşehir Bilim Ve Teknoloji Dergisi 3(2):33–47. https://doi.org/10.17100/nevbiltek.210929
Sutanto KD, Husain M, Rasool KG, Al-Qahtani WH, Aldawood AS (2021) Pathogenicity of local and exotic entomopathogenic fungi isolates against different life stages of red palm weevil (Rhynchophorus ferrugineus). PLoS ONE 16(7):1–12. https://doi.org/10.1371/journal.pone.0255029
Vega FE, Goettel MS, Blackwell M, Chandler D, Jackson MA, Keller S, Koike M, Maniania NK, Monzón A, Ownley BH, Pell JK, Rangel DEN, Roy HE (2009) Fungal entomopathogens: new insights on their ecology. Fungal Ecol 2:149–159. https://doi.org/10.1016/j.funeco.2009.05.001
Wang H, Peng H, Li W, Cheng P, Gong M (2021) The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front Microbiol 12(705343):1–11. https://doi.org/10.3389/fmicb.2021.705343
Yaşar B, Dahham OAD (2019) The effects of traps hanged on different apple varieties on the capture of Tropinota hirta (Poda, 1761) (Coleoptera: Cetoniidae). Turk J Agric Res 6(1):57–64. https://doi.org/10.19159/tutad.458712
Acknowledgements
The authors thank to Tokat Gaziosmanpaşa University Scientific Research Projects Coordination Unit for financial support, and to Dr. Şerife TOPKAYA (Department of Plant Protection, Faculty of Agriculture, Tokat Gaziosmanpasa University, Tokat, Turkey) for her help in molecular characterization studies and to Dr. Mustafa ALKAN for his help in statistical analysis (Directorate of Plant Protection Central Research Institute, Ankara, Turkey).
Funding
The study was supported by Tokat Gaziosmanpaşa University Scientific Research Projects Coordination Unit (Project Number: 2019/106).
Author information
Authors and Affiliations
Contributions
SU, TA and YY conceived and designed the research. SU conducted the experiments. TA analyzed the data and wrote the manuscript. YY corrected and revised the manuscript, corrected language mistakes and translation, and corrected references. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Uçar, S., Atay, T. & Yanar, Y. Identification of some native entomopathogenic fungal species and their pathogenicity against Apple Blossom Beetle, Tropinota (Epicometis) hirta (Poda, 1761) (Coleoptera: Cetoniidae) adults. Egypt J Biol Pest Control 32, 95 (2022). https://doi.org/10.1186/s41938-022-00594-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00594-8