
Abstract
Background
Sainfoin, alfalfa and clover are important forage crop plants in terms of their food value, efficiency and adaptation capabilities. Cymbalophora rivularis (Menetries, 1832) (Lepidoptera: Erebidae) causes significant damage to these plants. Chemicals are not recommended for pest control. Cultural pest control practices also remain insufficient. In this study, which was conducted to determine whether biological control can be applied in pest control, the pathogenic effects of Pseudomonas putida K-19B, Pantoea agglomerans RK-79 and RK-92, Bacillus megaterium TV-3D, P. fluorescens RK-1979, B. pumilus RK-1980 bacterial strains at a concentration of 1 × 108 CFU/ml and Beauveria bassiana (Bals.-Criv.) Vuill. (Cordycipitaceae) ET 10 fungal isolate at a concentration of 5.7 × 105 conidia/ml were tested under controlled conditions.
Results
According to the results of this study, at the end of 216 h, the tested biological control agent showed an efficacy ranged from 43.33 to 96.67% in the 1st and 2nd larval instars and 70 to 95% in the last instar larvae of C. rivularis. In the case of applications to 1st and 2nd larval instars, after 216 h, the efficacy of ET 10 fungal isolate was 96.67 and 83.33% of K-19B bacterial strain. In the last instar larvae, after 216 h, the efficacy of the strains TV-3D, K-19B and RK-1979 was 95, 90 and 90%, respectively. The efficacy of ET 10 and K-19B applications was found to be statistically not different from the efficacy of pathogenic application.
Conclusion
Environmentally friendly P. putida and B. megaterium bacterial strains and B. bassiana fungal isolate were determined and can be successfully used in the biological control of this pest.
Similar content being viewed by others


Background
In animal husbandry, farm animals need to be fed adequately with high-quality feeds so that they produce a high amount of quality products, including meat, milk, eggs, etc. (Demiroğlu Topçu and Özkan 2017). The development of the livestock sector and the achievement of productivity and profitability largely depend on the production of high-quality roughage. The amount of roughage needed cannot be met with the roughage obtained from pastures or by sowing forage crops, and the livestock has to be fed with poor-quality forages such as straw and grass. Therefore, it is necessary to increase the forage crop cultivation areas and ensure their sustainability in order to meet the roughage need of the existing livestock. Considering the nutritional value of the forage crops cultivated, especially legume forage plants such as Medicago sativa (L.) (Fabaceae), Onobrychis sativa (Lam.) (Fabaceae), Trifolium spp. (Fabaceae) and Vicia sativa (L.) (Fabaceae) are very important in animal nutrition. These plants are not only beneficial in terms of giving nitrogen to the soil, improving the physical and chemical properties of the soil, increasing the yield of the plants planted after them and using them in rotation but also play an important role in water and soil conservation and prevention of erosion.
As with all cultivated plants, there are pests, diseases and weeds that reduce the quality and yield of forage plants. Of these, Cymbalophora rivularis (Menetries, 1832) (Lepidoptera: Erebidae) larvae feeding on the leaves of newly emerging plants in spring causes significant losses in the sainfoin (Kılıç et al. 2013). As a cultural precaution in the control of the pest, when the nets that the larvae weave in the form of tents are seen, collecting and destroying them, together with the tents, are recommended. However, this form of control does not give successful results in large areas. Furthermore, although there are species belonging to Tachinidae (Diptera) and Ichneumonidae (Hymenoptera) families as natural enemies of this pest in Turkey, pesticides used in agricultural areas reduce the efficacy of these natural enemies.
In this study, insecticidal efficacy of entomopathogenic bacteria and fungi on larval instars of C. rivularis was tested under controlled conditions.
Methods
Pest, bioagent, bacterial strains and fungal isolate
Cymbalophora rivularis larvae were collected from forage plants, transferred to Atatürk University, Faculty of Agriculture, Department of Plant Protection, Pest Systematics laboratory, and stored in (30 × 45 × 30 cm) plastic containers at 25 ± 2 °C under 65% ± 5 RH and 16-h light/8-h dark conditions by providing daily fresh food and humidity check.
Bacterial strains and fungal isolate used in the study are presented in Table 1. These microorganisms have been reported to be effective on disease (Kotan et al. 2005) or pests (Tozlu et al. 2017) or to be highly beneficial in terms of plant growth in various plant groups (Ekinci et al., 2014). Bacterial strains were identified by microbial identification system (MIS), and it was determined that they give a positive reaction in terms of nitrogen fixation and phosphate dissolving properties. Bacterial biocontrol strains were kept in nutrient broth (NB; Difco) containing 15% glycerol at − 80 °C in the Culture Collection in Atatürk University Faculty of Agriculture Plant Protection Department Plant Clinical Laboratory. Fungal isolate ET 10 was identified as Beauveria bassiana according to ITS gene region and kept in tubes containing potato dextrose agar (PDA; Difco) medium at + 4 °C in Atatürk University Faculty of Agriculture Department of Plant Protection Mycology Laboratory.
Preparation of bacterial strains and fungal isolate
The tested bacterial strains were cultured in four phases in nutrient agar (NA; Difco) medium at 27 °C for 24 h in order to obtain fresh cultures by checking the purity of the colonies. A single bacterial colony taken from these cultures in sterile loops was inoculated into Erlenmeyer flasks with 300 ml of NB medium and incubated for 24 h at 250 rpm and 27 °C in a horizontal shaker incubator. The bacterial density of the resulting aqueous culture was adjusted to 1 × 108 CFU/ml by BIOLOG turbidimeter and transferred to sterile spray vials.
Conidia production was achieved by incubation of B. bassiana ET 10 isolate in Sabouraud dextrose agar (SDA; Difco) medium at 25 °C, 80% RH for 2–3 weeks. Then, a stock suspension was prepared by washing the surface of the culture into bottles containing sterile water with 0.2 ml/l Tween 80 solution (Quesada-Moraga et al. 2006). Using a hemocytometer, the concentration of conidia suspension to be applied was found to be 5.7 × 105conidia/ml.
Virulence test in controlling conditions
Fresh sainfoin leaves were placed in polyethylene plastic boxes (19 × 12.5 × 7 cm) laid with blotting paper on the base, and 1st and 2nd and last instars mature 10 larvae of the pest were placed on them. Then, the bacterial and fungal suspensions were sprayed onto the leaves. The entomopathogen-applied plastic boxes were kept under controlled conditions (25 ± 2 °C, 65–70% RH, 16:8 (light/dark)). The number of dead larvae was recorded regularly every 24 h. The final evaluation of the trial was carried out, and the mortality rates were determined at 216 h. By isolating the larvae determined to be infected according to Koch’s postulates, entomopathogenic bacterial strains and fungal isolates were obtained again. In the study, sterile NB medium was used as negative control, and Coption 65 EC (650 g/l malathion) suitable for pests with biting chewing mouth structure was used as a positive control since there is no recommended chemical for control of this pest. The experiment was carried out in three repetitions for each application on the same day.
Data analysis
The number of dead insects obtained in the trial was converted to mortality rate (below formula), and analysis of variance was applied and the differences between the means were compared to the LSMeans Student’s t test at a significance level of P < 0.01. Data analyses were performed using statistical software package JMP IN (SAS Institute, Cary, NC, 0.0% PC version).
Results
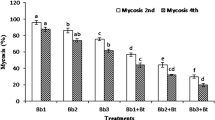
According to the results obtained, all bacterial strains and fungal isolates were found to be effective on the 1st, 2nd and last larval instars of the pest at varying rates. In the study, the highest mortality rate at 100% in the 1st and 2nd larval instars at 24 h was obtained for pathogenic application (Table 2). Following this application, the earliest mortality was counted 48 h in larvae to which ET 10 (3.33%) fungal isolate was applied, followed by bacterial strains TV-3D (3.33%) with 72 h, K-19B (66.67%), RK-1979 (20%), RK-92 (6.67%) and RK-1980 (6.67%) with 120 h (Table 2). At the end of 216 h, the highest mortality rates were identified in the larvae to which ET 10 fungal isolate (96.67%) and K-19B bacterial strain (83.33%) were applied (Table 2).
When each application was evaluated within itself, it was observed that all larvae died within 24 h of pathogenic application. In the larvae to which ET 10 fungal isolate was applied, mortality started early, but the mortality rate increased (60%) after 120 h and reached above 90% at 144 h, and a mortality rate of 96.67% occurred at 216 h (Fig. 1). In the K-19B bacterial strain, on the other hand, the first mortalities were observed at 120 h (66.67%), and 83.33% mortality was determined after 216 h (Fig. 1). While 20% mortality was detected in the RK-1979 bacterial strain in 120 h, the mortality rate increased gradually and reached 83.33% at the end of 216 h (Fig. 1). Although mortality started in 72 h in TV-3D bacterial strain, it was determined that the mortality rate caused by this bacterial strain was 76.67% in the evaluations made at the end of 216 h, and this strain was in 5th place in terms of mortality rate among the applications (Fig. 1). While 6.67% mortality was observed in RK-92 and RK-1980 bacterial strains at the end of 120 h, this rate was 76.67% in RK-92 and 70% in RK-1980 at the end of 216 h. (Fig. 1). The lowest results among the applications were obtained for the RK-79 bacterial strain. The larvae to which this strain was applied began to die at the end of 68 h (26.67%), and a mortality rate of 43.33% was reached at the end of 216 h (Fig. 1). In the control to which NB was applied, the mortality rate was 6.67% at 144 h and 10% at 216 h (Fig. 1).

Virulence effect of the applications on 1st and 2nd larval instars of Cymbalophora rivularis
A mortality rate of 100% was determined in the last instar mature larvae in the application of pathogens in 24 h, while mortality (10.00%) occurred in the larvae to which RK-1979 bacterial strain was applied as from the 72 h after the pathogen application, followed by ET 10 fungal isolate (25.00%) during the evaluation made at 120 h (Table 3). The highest mortality rate was identified for TV-3D bacterial strain (95.00%) at the end of 216 h in the applications made to the last instar larvae, and this strain was followed by K-19B and RK-1979 bacterial strains with a mortality rate of 90.00% (Table 3).
As a result of the study, all last instar larvae died within 24 h of insecticide application (Fig. 2). In the larvae to which RK-1979 bacterial strain was applied, mortality began at the end of 72 h, but it was determined that the mortality rate did not change in the evaluations made at 120 and 144 h, the mortality rate increased gradually as from 168 h, and the mortality rate was 90% at 216 h (Fig. 2). In ET 10 fungal isolate, on the other hand, mortality began as from the 72 h, and the mortality rate was the same (40%) at 144 and 168 h and reached 85% at the end of 216 h (Fig. 2). In the larvae to which K-19B bacterial strain was applied, mortality began at 120 h with a rate of 6.67% and the mortality rate increased to 90% at 216 h (Fig. 2). In RK-79, RK-92 and TV-3D bacterial strains, the mortality rate was 25% at 144 h, which then increased to 85, 85 and 90% at 216 h, respectively (Fig. 2). In the RK-1980 bacterial strain, the mortality rate was 5% at 144 h and increased to 70% at 216 h. There was no larval mortality in the controls to which NB was applied (Fig. 2). Some examples of the virulence effects of bacterial strains and fungal isolate used in the study on the pest are shown in Fig. 3.

Virulence effect of the applications on the last instar larvae of Cymbalophora rivularis

Virulence effect of bacterial strains and fungal isolate
Discussion
Entomopathogens are widely used in the biological control of pests that cause yield and quality loss in many crops and is among the most effective factors regulating pest populations (Kumar et al. 2021). Among these entomopathogens, bacteria are among the most commonly used biocontrol agents against plant pathogens and pests. Among them, Bacillus thuringiensis, B. brevis, B. cereus, B. circulans, B. megaterium, B. pumilus and B. subtilis are known to be used for biotechnological and industrial applications (Tozlu et al. 2019). Among these species, B. megaterium produces the enzyme amylase, which is effective in carbohydrate digestion and therefore important with this feature (Prasanna et al. 2014), and B. pumilus, with effective results in this study, has been shown to be antifungal, antibacterial (Lehman et al. 2001) and pathogenic (Tozlu et al. 2019) effects and these effects are due to chitinase activity (Rishad et al. 2017), while P. fluorescens enzymatically hydrolyzes chitin in the exoskeleton of insects, the enzyme affects the digestion of the insect and directly affects the growth and development of the insect. It has been reported that it causes death by preventing the disease (Suganthi et al. 2017).
Besides bacteria, fungi are also important in biological control (Zhang et al. 2021). Entomopathogen fungi are also used as an alternative to chemical insecticides in the biological control of pests (Sharma and Sharma 2021). The use of these organisms has numerous advantages such as protecting beneficial organisms, being safe for humans, and not having harmful effects on the environment (Mantzoukas and Eliopoulos 2020). In particular, it has been noted that some species of Beauveria infect and kill insects in the soil, interact with plant roots for their growth and survival, and mostly depend on insects for carbon (Inglis et al. 2001). It has been noted that since this fungus directly infects the insect's cuticle, it does not have to be eaten by the host and this feature makes entomopathogenic fungi important in the control of pests (Goettel et al. 2005).
Conclusions
Among the safe methods to control the pests, entomopathogens are considered as the leading alternative. As a result of this study, it was determined for the first time that entomopathogenic bacteria (P. putida, B. megaterium and P. fluorescens) and fungi (B. bassiana) can be used successfully as an alternative to chemicals in the control of C. rivularis. Further field studies should be tested.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- − :
-
Negative
- +:
-
Positive
- °C:
-
Degrees Celsius
- B. bassiana :
-
Beauveria bassiana
- B. brevis :
-
Bacillus brevis
- B. cereus :
-
Bacillus cereus
- B. circulans :
-
Bacillus circulans
- B. megaterium :
-
Bacillus megaterium
- B. pumilus :
-
Bacillus pumilus
- B. subtilis :
-
Bacillus subtilis
- C. virulans :
-
Cymbalophora rivularis
- CFU:
-
Colony-forming units
- hrs:
-
Hours
- ITS:
-
Internal transcribed spacer
- ml:
-
Milliliter
- NA:
-
Nutrient agar
- NB:
-
Nutrient broth
- NF:
-
Nitrogen fixation
- PDA:
-
Potato dextrose agar
- PS:
-
Phosphate solubilization
- RH:
-
Relative humidity
- rpm:
-
Centrifugal rotation speed
- S. antiqua :
-
Sphenoptera antiqua
- S+:
-
Strong positive
- SDA:
-
Sabouraud dextrose agar
- SIM:
-
Similarity index
References
Demiroğlu Topçu G, Özkan ŞS (2017) General view to meadow-rangelands and forage crops cultivation of Aegean region and Turkey. COMU J Agric Facul 5(1):21–28 ((in Turkish))
Ekinci M, Turan M, Yildirim E, Güneş A, Kotan R, Dursun A (2014) Effect of plant growth promoting rhizobacteria on growth, nutrient, organic acid, amino acid and hormone content of cauliflower (Brassica oleracea L. var. botrytis) transplants. Acta Scientiarum Polonorum Hortorum Cultus 13(6):71–85
Goettel S, Eilenberg J, Glare T (2005) Entomopathogenic fungi and their role in the regulation of insect populations. In: Gilbert LB, Iatrou K (eds) Comprehensive molecular insect science. Elsevier Pergamon, Oxford, pp 361–406
Inglis GD, Goettel MS, Butt TM, Strasser H (2001) Use of hyphomycetous fungi for managing insect pests. In: Butt TM, Jackson C, Magan N (eds) Fungi as biocontrol agents: progress problems and potential, vol 3. CABI Publishing, pp 23–70
Karagöz K, Ateş F, Karagöz H, Kotan R, Çakmakçı R (2012) Characterization of plant growth-promoting traits of bacteria isolated from the rhizosphere of grapevine grown in alkaline and acidic soils. Eur J Soil Biol 50:144–150
Kılıç T, Ulutaş E, Taşkın T, Kaya A, Sokat Y (2013) Diseases, pests and weeds that are a problem in forage crops such as alfalfa, sainfoin and vetch. Türkiye Tohumcular Birliği Dergisi (TURKTOB) 6:47–53 ((in Turkish))
Kotan R, Sahin F, Ala A (2005) Identification and pathogenicity of bacteria isolated from pome fruits trees in eastern Anatolia region of Turkey. J Plant Dis Prot 113:8–13
Lehman LJ, McCoy RJ, Messenger BJ, Manker DC, Orjala JE, Lindhard D, Marrone PG (2001) Strain of Bacillus pumilus for controlling plant diseases caused by fungi. U.S. Patent 6:245–551
Mantzoukas S, Eliopoulos PA (2020) Endophytic entomopathogenic fungi: a valuable biological control tool against plant pests. Appl Sci 10:360
Prasanna VA, Kayalvizhi N, Rameshkumar N, Suganyan T, Krishnan M (2014) Characterization of amylase producing Bacillus megaterium from the gut microbiota of Silkworm Bombyx mori. Res J Chem Environ 18(7):38–45
Quesada-Moraga E, Ruiz-García A, Santiago-Alvarez C (2006) Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritidae). J Econ Entomol 99(6):1955–1966
Rishad KS, Rebello S, Shabanamol PS, Jisha MS (2017) Biocontrol potential of Halotolerant bacterial chitinase from high yielding novel Bacillus pumilus MCB-7 autochthonous to mangrove ecosystem. Pestic Biochem Physiol 137:36–41
Sharma R, Sharma P (2021) Fungal entomopathogens: a systematic review. Egypt J Biol Pest Control 31:57. https://doi.org/10.1186/s41938-021-00404-7
Suganthi M, Senthilkumar P, Arvinth S, Chandrashekara KN (2017) Chitinase from Pseudomonas fluorescens and its insecticidal activity against Helopeltis theivora. J Gen Appl Microbiol 63(4):222–227
Tozlu E, Kotan R, Tozlu G (2017) The Investigation of Beauveria bassiana (Ascomycota: Hypocreales) as a biocontrol agent of rose-stem sawfly, Syrista parreyssii (Spinola, 1843) (Hymenoptera: Symphyta; Cephidae) larvae. Fresenius Environ Bull 26(12):7091–7100
Tozlu E, Saruhan I, Tozlu G, Kotan R, Dadaşoğlu F, Tekiner N (2019) Potentials of some entomopathogens against the brown marmorated stink bug, Halyomorpha halys (Stål, 1855) (Hemiptera: Pentatomidae). Egypt J Biol Pest Control 29:76
Zhang M, Yan J, Reitz S, Wu S, Gao Y (2021) Beauveria bassiana as fungal endophyte for the potential control of the potato tuber moth Phthorimaea operculella on potatoes. Res Square 6:66. https://doi.org/10.21203/rs.3.rs-726671/v1
Acknowledgements
Not applicable.
Funding
No funding.
Author information
Authors and Affiliations
Contributions
ET, GT and RK conceived, designed, analyzed, wrote, corrected and approved the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tozlu, E., Tozlu, G. & Kotan, R. Biocontrol of Cymbalophora rivularis (Menetries) (Lepidoptera: Erebidae) larvae by entomopathogenic bacteria and fungi. Egypt J Biol Pest Control 32, 66 (2022). https://doi.org/10.1186/s41938-022-00565-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00565-z