
Abstract
The discovery of small non-coding RNAs, such as miRNA and piRNA, has dramatically changed our understanding of the role RNA plays in organisms. Recent studies show that a novel small non-coding RNA generated from cleavage of tRNA or pre-tRNA, called tRNA-derived small RNA (tsRNA), serves as a new regulator of gene expression. tsRNA has been determined participate in regulating some specific physiological and pathological processes. Although knowledge regarding the biological roles of miRNA and piRNA is expanding, whether tsRNAs play similar roles remains poorly understood. Here, we review the current knowledge regarding the mechanisms of action and biological functions of tsRNAs in intracellular, extracellular and intergenerational inheritance, and highlight the potential application of tsRNAs in human diseases, and present the current problems and future research directions.
Similar content being viewed by others


Introduction
Small noncoding regulatory RNAs have emerged as vital players in various biological processes. Among them, microRNAs (miRNAs), which bind their complementary sites in the 3′-untranslated regions (UTRs) of target mRNAs, further inhibiting target gene expression at post-transcriptional level, is the most extensively studied [1, 2]. Recently, tRNA-derived small RNAs (tsRNA), a new type of small noncoding RNAs, have drawn great attention.
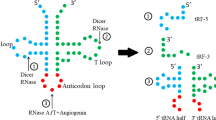
These small RNA fragments are not random degradation products, but specific cleavage products, which play important roles in various pathological and physiological conditions [3]. Collectively, there are two types of tsRNAs were produced from tRNAs. In the first, the 5′ and 3′ tRNA halves, also called tRNA-derived stress-induced RNAs (tiRNAs), are 30–40 nt long. tiRNAs are generated from cleavage of tRNAs by ribonucleases Rny1 and angiogenin (ANG) near or in the anticodon loop under pressure [4, 5]. Under pressure, for example, amino acid deficiency, hypoxia, UV radiation, oxidative damage, heat shock, phosphate starvation and viral infection [4, 6,7,8,9,10,11], the breakage of tRNAs happens. Nonetheless, this cleavege rarely occurs under normal conditions and very few tiRNAs are produced [12]. The other type of tsRNAs is tRNA-derived fragments (tRFs), tRFs are 18–22 nt and shorter than tiRNAs. tRFs are generated from mature and tRNA precursors by nucleases Dicer or RNase Z. According to their cutting site and sequence location on tRNAs, there are four types of tRFs at present, including 5-tRFs, 3-tRFs, 1-tRFs, and 2-tRFs [13,14,15,16,17,18,19] (Fig.1).

The structure of tRNA and classification of tsRNAs. a The schematic diagram of the secondary structure of tRNA. b The types of tsRNAs: 1-tRF(cleaved by RNase Z in the 3′ trailer of ribosomal RNA precursor), 2-tRF(generated by unknown ribonuclease), 3-tRFs and 5-tRFs, 5′ tiRNA and 3′ tiRNA are cleaved from the anticodon loop
As small non-coding RNAs, tsRNAs have a lot in common with miRNAs, such as length and mechanisms of action. In this study, we summarize the latest views about tsRNA functions and discuss the similarity and difference in biological roles and functional mechanism between miRNAs and tsRNAs.
Mechanisms of action of tsRNA
As we known, miRNAs function via base-pairing with complementary sites on the 3′-UTR of target mRNAs, further leading to mRNA degradation or translational repression [1]. More prominently, recent studies found the new action mode of miRNAs.
Unlike usual function in regulating mRNA stability on post-transcriptionally, miRNAs can also directly bind to Toll-like rec'eptors (TLRs) as a physiological ligand and trigger a specific biological action [20, 67]. Although the universal and specific mechanisms of action of tsRNAs are not very clear due to the limited studies on a few tsRNAs, tsRNA still showed some common features of small non-coding RNAs like miRNAs or piRNAs.
Regulating mRNA stability
As a type of small non-coding RNAs, tRFs have similar functional mechanisms with miRNAs. MiRNAs can directly combine with mRNA to regulate mRNA stability. Luo et al. demonstrated that most tsRNAs were abundant and conserved in Drosophila. Besides, tsRNAs could recognize mRNAs by binding to conserved complementary sequence and further inhibit target genes (key components of the general translation machinery) by translational inhibition [21]. Previous studies found that 3-tRFs have miRNA-like structure and function, directly binding to target mRNA and resulting in translational repression or mRNA degradation in mature B lymphocytes or non-small cell lung cancer cells [22, 23]. Interestingly, recent studies showed that tRFs can combine with Argonautes (Ago) 1, 3 and 4 but not Ago2 in human 293 cells. Although tRFs can’t bind to Ago2 (the main effector protein of miRNA function), they have similar characteristics with miRNAs, suggesting tRFs may involving in silencing RNA [24].
Binding to protein
Proteins which combine various RNA targets can serve as main regulators of gene expression to harmonize cellular processes and influence cell phenotype. For example, a recent study found that 1-tRFs and 3-tRFs take part in globally controlling small RNA silencing through competitively combining with Ago family, further influencing the silence efficiency of target genes [25]. In addition, a novel class of 2-tRFs derived from tRNA(Asp), tRNA(Tyr), tRNA(Gly) and tRNA(Glu) can competitively bind to YBX-1 and inhibit various oncogenic mRNAs stabilization in breast cancer (BRC) via transposing their 3′ UTRs [19]. Some studies found that specific tiRNAs could inhibit translation by displacing elF4G/elF4A from uncapped to capped RNAs depending on a terminal oligoguanine motif [26]. tiRNAs were packaged into a structure similar to G-quadruplex, competitively combining with eIF4G/eIF4A in the complex of translation initiation and further suppressing mRNA translation [27]. A previous study found that tRNA Val-GAC-produced 5-tRF in Haloferax volcanii could combine ribosomal subunit near mRNA channel and weaken synthesing protein integrally [28]. Further structural studies are needed to more sufficient explore how tRFs directly bind to protein.
Transposable elements
In general, transposable elements are transcriptionally inhibited by epigenetic factors, for instance, DNA methylation and histone modification [29]. In a recent study, a group determined that 18 or 22 nt tRFs could suppress reverse transcription, retrotransposon mobility (18 nt) and further silence the long terminal repeat (LTR) retrotransposon or post-transcription similar to miRNA (22 nt) [30]. Furthermore, Dicer-like 1 (DCL1)-cleaved and produced 5-tRFs were merged into Ago1, further participate in regulating the stability of genome by targeting transposon element mRNAs in plant Arabidopsis thaliana.
Regulating RNA reverse transcription
tsRNAs can also participate in regulating virus RNA reverse transcription. Ruggero et al. showed that tRF-3019, processed from tRNA-proline, exhibited perfect sequence complementarity to the binding sites in primers of human T-cell leukemia virus type 1 (HTLV-1), then priming HTLV-1 reverse transcription [31]. Meantime, infection of respiratory syncytial virus (RSV) can induce ANG to cut tRNAs to process tiRNAs in host cells. Host tiRNAs can serve as primers to promote RSV replication and enhance their infection efficiency [32,33,34]. Therefore, both tRFs and tiRNAs can be used as primers to regulate RNA reverse transcription. Further research on the functional mechanisms of tsRNAs may provide valuable ideas to explore the biological function of tsRNAs.
Biological function of tsRNAs
Biological function of intracellular tsRNAs
Intracellular miRNAs play a crucial part in regulating a variety of life activities, including individual development, cellular proliferation, apoptosis, metabolism and tumorigenesis, and their abnormal expressions make for the pathogenesis of many human diseases [35,36,37,38].
tsRNA and stress
Under stress stimulation, for instance, hypoxia, oxidative stress, UV irradiation and heat/cold shock, tRNA halves expression was induced [39]. Consequently, tRNA halves are also called tRNA-derived stress-induced RNA (tiRNA) [6]. ANG, a ribonuclease of RNase A superfamily, responsible for processesing tRNAs into tiRNAs [4]. The cleavage of tRNAs induced by stress has been found in Tetrahymena thermophile lack of nutrient [7], Streptomyces coelicolor [40], Trypanosoma cruzi [41], and in serum-deprived Giardia lamblia [9], spore-forming Aspergillus fumigatus [8], phosphate-depleted Arabidopsis thaliana [42], oxidatively stressed Saccharomyces cerevisiae [10] and Homo sapiens [39]. Several studies show that the tRNA fragments induced by stress could directly suppress protein biosynthesis. For example, 5′-tRNA fragments overexpression, but not 3′-tRNA fragments, had been shown to globally inhibit the translation in human U2OS cells [6] and trigger the package of cytoplasmic foci and stress granules (SGs) induced by translation initiation suppressors [43]. In a previous study, Ivanov et al. revealed that tiRNAs may combine with YB-1 to regulate the translation in stressed cells [26].
tsRNA and cancer
Dysregulation of tsRNA had been observed in various malignancies. For example, in breast cancer, hypoxic stress could induce the generation of tRFs, and this type of tRFs inhibits various oncogenic mRNA stabilization in BRC cells by replacing the 3′ UTRs from YBX1. In addition, highly metastatic cells could escape the pathways of tumor suppression by weakening production of the tRFs [19] (Table.1). In addition, Pekarsky et al. reported that two members of tsRNA family, ts-3676 and ts-4521, were inactivated in lung cancer and chronic lymphocytic leukemia (CLL) and interacted with piwi proteins. They also measured the expression of 120 tsRNAs via microarray chip and found that tsRNAs are dysregulated in lung cancer and CLL [44]. Honda et al. had found that a novel type of tsRNAs were abundant in estrogen receptor (ER)-positive BRC and prostate cancer cell lines and patient tissues with androgen receptor (AR)-positive, and increased by sex hormones and their receptors. Among them, 5′- tsRNAs, but not 3′-tsRNAs, has extensively participate in regulating cell proliferation. Maute et al. identified a miRNA-like tRNA-derived fragment downregulated in B cell lymphoma and inhibits cell proliferation. Moreover, the fragments regulate the DNA damage response through repressing mRNA transcripts in a sequence-specific mode [22]. Besides, Zhao et al. also found four 5′-tiRNAs were decreased in clear cell renal cell carcinoma tissues, indicating the potential role of tsRNA as a tumor suppressor [45]. Although several tsRNA had been showed play vital roles in cancer progression and tumorigenesis, the precious mechanism and common function of tsRNAs in cancer need further study.
tsRNA and neurodegenerative disease
Defects in tsRNA generation and processing enzymes were observed associated with a number of meurological disorder. For example, ANG mutants results in reduced RNase activity and serves as a potential predisposing genes for amyotrophic lateral sclerosis (ALS), a neurodegenerative disease characterized by adult-onset lack of motor neurons [46]. In 2012, a significantly higher frequency of ANG variants was also discovered in patients with Parkinson’s disease (PD) [47]. Recombinant ANG is beneficial for culturing motoneurons depend on RNase activity [48,49,50], suggested these effect due to the formation of tsRNAs. Actually, tiRNAs may promote motor neuron survival via inhibiting apoptosis [51] or promoting of SGs [26, 43].
tsRNAs also participate in neurodegeneration diseases [52,53,54]. CLP1 gene (R140A) mutations, the first RNA kinase to be identified in mammalian, are detected in neurological syndrome patients including peripheral nervous system (PNS) and central nervous system (CNS). Besides, the mice with CLP1 kinase-dead (CLP1K/K) show a progressive loss of spinal motor neurons correlated with axonal degeneration in denervation of neuromuscular junctions and peripheral nerves, finally leading to fatal respiratory failure, muscle weakness, paralysis, and impaired motor functions [52]. CLP1 mutants lose the kinase activity, tRNA endonuclease complex (TSEN) was nonstationary and the cleavage of tRNA precursor was further inhibited. In a previous study, in zebrafish, CLP1 null germline exhibited cerebellar neurodegeneration, this phenomenon could be rescued by overexpressing WT human CLP1, but couldn’t be rescued by mutant CLP1 overexpression. In patients, the neurons showed depleted mature tRNAs and accumulated unspliced tRNA precursors. When the patient cells were transfected with partially processed tRNA fragments, the oxidative stress-induced decrease of cell survival would further exacerbated. This research combines the maturation of tRNAs with neurodegeneration and neuronal development via impaired CLP1 function in human [54]. Recent study found that 5-tRFs accumulate in the CLP1 kinase-dead mice and further increase the cell sensitivity to oxidative stress and further promote cell apoptosis in a p53-dependent form [52]. Although most studies have focused on the details of the interaction between CLP1 activities, the mechanisms of tRNA splicing and maturation and tRNA precursor-derived tRFs functions are yet not clearly understood. The above studies provide an idea for the following research on combining deregulated tRNA metabolism with the development of neurodegeneration.
There is growing attention on the research on the possible biological functions of tRNA-derived fragments. For instance, tsRNAs are potential systemic immune signaling molecules [55]. Virus infection also lead to abundant production of tsRNA, which inhibits target mRNA of cytoplasm and facilitates RSV replication [33]. Translational control in stem cells had been reported regulated by pseudouridylation of specific tsRNAs, influencing stem cell commitment during key developmental processes [56]. tsRNA also could combine with RPS15 and RPS28 ribosomal protein mRNAs to accelerate the translation and fine-tuned ribosome biogenesis [57].
Biological function of extracellular tsRNA
It is generally assumed that RNA is unstable due to the ubiquitous ribonuclease, now miRNA is known to circulate in the bloodstream and other body fluids in a stable, cell-free form. Importantly, extracellular miRNAs show abnormal expressions in serum, plasma and other body fluids in the development and progression of many diseases and serve as promising noninvasive biomarkers to estimate the pathological status of the body [58]. tsRNAs has also been found stability in the circulation. Joseph M et al. found that 5′-tiRNAs can stably exist in the blood and tiRNA levels are correlated with age and calorie restriction [59]. They thought that tiRNAs maintain stability in blood most likely by binding to a nucleoprotein complex. In addition, this group found the significant changes in the levels of specific 5′-tiRNAs associated with breast cancer [60]. Besides, Zhang et al. screened small RNAs (18–40 nt) in the multiple mouse organs, revealing that tsRNAs present in the serum in a surprisingly high percentage (~ 70%), more than miRNA reads in general [61]. They showed that the tsRNAs in serum were not concentrated in exosome but remained in the supernatant and be protected from rapid degradation via protein binding and nucleotide modifications [61]. The diagnostic values and biological functions of tsRNAs in serum are intriguing, which need more detailed research and possibly opens a new round of study focus on serum small RNAs.
tsRNAs in intergenerational inheritance
Once deemed heretical, recent evidence supports the view that acquired characteristics inheritance can occur via ancestral experiences or exposures and that some paternally acquired traits could be ‘memorized’ in the sperm as epigenetic information. It is very tempting to speculate that small RNAs, including tsRNAs and miRNAs, may also be mobile signals that can experience alterations from paternal environment and facilitate crosstalk between generations. The tsRNAs enrichment of mature sperm was first found by RNA-seq analysis in mice [62]. tsRNAs in sperm are produced from the 5′ end of tRNAs, 29–34 nt long and are more enriched than miRNAs, accounting for the majority of small ncRNAs in sperm [62,63,64]. Sperm tsRNAs expression profiles and RNA modifications changed significantly in a male mice model of high-fat diet (HFD). And injection the sperm tsRNA fraction of HFD male mice into normal zygotes leaded to metabolic disorder in the F1 offspring and changed metabolic pathways gene expression in early embryos and islets of F1 offspring, indicating tsRNAs in sperm stand for a paternal epigenetic factor that mediate intergenerational inheritance of diet-induced metabolic disorder. Sperm miRNAs are also found altered after paternal exposure to trauma and can mediate the intergenerational inheritance of certain traits in mice [65]. However, studying the contributions of tsRNAs or miRNAs in sperm to the acquired traits transfer is just the first step, the precise contribution may be more complex and needs step-by-step research.
Conclusions
The study of tsRNA is still in a very early stage. Maybe more aspects of tsRNA will be found in the near future, such as the specific mechanism of biogenesis process and the clinical application. In addition, the universality and specificity of tsRNA are still unclear. tsRNA are produced from tRNA and it is difficult to specifically change the tsRNAs content without affecting the expression level of mature tRNAs. Maybe tsRNA regulate translational process for this reason, further resulting in the some abnormal function. Better understanding the function of tsRNA will provide valuable views on human physiology and pathophysiology.
Abbreviations
- Ago:
-
Argonaute
- ANG:
-
Angiogenin
- BRC:
-
Breast cancer
- CLL:
-
Chronic lymphocytic leukemia
- CNS:
-
Central nervous system
- DCL1:
-
Dicer-like 1
- LTR:
-
Long terminal repeat
- miRNA:
-
MicroRNA
- PNS:
-
Peripheral nervous system
- tiRNA:
-
tRNA-derived stress-induced RNA
- TLR:
-
Toll-like receptor
- tRF:
-
tRNA-Derived fragments
- tsRNA:
-
tRNA-Derived small RNA
- UTRs:
-
Untranslated regions
References
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.
Lee YS, Dutta A. MicroRNAs in Cancer. Annu Rev Pathol-Mechanisms of Disease. 2009;4:199–227.
Li S, Xu Z, Sheng J. tRNA-derived small RNA: a novel regulatory small non-coding RNA. Genes. 2018;9(5):246.
Fu H, Feng J, Liu Q, Sun F, Tie Y, Zhu J, Xing R, Sun Z, Zheng X. Stress induces tRNA cleavage by angiogenin in mammalian cells. FEBS Lett. 2009;583(2):437–42.
Thompson DM, Parker R. The RNase Rny1p cleaves tRNAs and promotes cell death during oxidative stress in Saccharomyces cerevisiae. J Cell Biol. 2009;185(1):43–50.
Yamasaki S, Ivanov P, G-f H, Anderson P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J Cell Biol. 2009;185(1):35–42.
Lee SR, Collins K. Starvation-induced cleavage of the tRNA anticodon loop in Tetrahymena thermophila. J Biol Chem. 2005;280(52):42744–9.
Joechl C, Rederstorff M, Hertel J, Stadler PF, Hofacker IL, Schrettl M, Haas H, Huettenhofer A. Small ncRNA transcriptome analysis from Aspergillus fumigatus suggests a novel mechanism for regulation of protein synthesis. Nucleic Acids Res. 2008;36(8):2677–89.
Li Y, Luo J, Zhou H, Liao J-Y, Ma L-M, Chen Y-Q, Qu L-H. Stress-induced tRNA-derived RNAs: a novel class of small RNAs in the primitive eukaryote Giardia lamblia. Nucleic Acids Res. 2008;36(19):6048–55.
Thompson DM, Lu C, Green PJ, Parker R. tRNA cleavage is a conserved response to oxidative stress in eukaryotes. Rna-a Publication of the Rna Society. 2008;14(10):2095–103.
Hsieh L-C, Lin S-I, Shih AC-C, Chen J-W, Lin W-Y, Tseng C-Y, Li W-H, Chiou T-J. Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol. 2009;151(4):2120–32.
Honda S, Loher P, Shigematsu M, Palazzo JP, Suzuki R, Imoto I, Rigoutsos I, Kirino Y. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc Natl Acad Sci U S A. 2015;112(29):E3816–25.
Kumar P, Mudunuri SB, Anaya J, Dutta A. tRFdb: a database for transfer RNA fragments. Nucleic Acids Res. 2015;43(D1):D141–5.
Lee YS, Shibata Y, Malhotra A, Dutta A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009;23(22):2639–49.
Li Z, Ender C, Meister G, Moore PS, Chang Y, John B. Extensive terminal and asymmetric processing of small RNAs from rRNAs, snoRNAs, snRNAs, and tRNAs. Nucleic Acids Res. 2012;40(14):6787–99.
Maraia RJ, Lamichhane TN. 3’processing of eukaryotic precursor tRNAs. Wiley Interdiscip Rev RNA. 2011;2(3):362–75.
Babiarz JE, Ruby JG, Wang Y, Bartel DP, Blelloch R. Mouse ES cells express endogenous shRNAs, siRNAs, and other microprocessor-independent, dicer-dependent small RNAs. Genes Dev. 2008;22(20):2773–85.
Liao J-Y, Ma L-M, Guo Y-H, Zhang Y-C, Zhou H, Shao P, Chen Y-Q, Qu L-H. Deep sequencing of human nuclear and cytoplasmic small RNAs reveals an unexpectedly complex subcellular distribution of miRNAs and tRNA 3’trailers. PLoS One. 2010;5(5):e10563.
Goodarzi H, Liu X, Nguyen HCB, Zhang S, Fish L, Tavazoie SF. Endogenous tRNA-derived fragments suppress breast Cancer progression via YBX1 displacement. Cell. 2015;161(4):790–802.
Chen X, Liang H, Zhang J, Zen K, Zhang C-Y. microRNAs are ligands of toll-like receptors. RNA-a Publication of the Rna Society. 2013;19(6):737–9.
Luo SQ, He F, Luo JJ, Dou SQ, Wang YR, Guo AN, Lu J. Drosophila tsRNAs preferentially suppress general translation machinery via antisense pairing and participate in cellular starvation response. Nucleic Acids Res. 2018;46(10):5250–68.
Maute RL, Schneider C, Sumazin P, Holmes A, Califano A, Basso K, Dalla-Favera R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc Natl Acad Sci U S A. 2013;110(4):1404–9.
Shao Y, Sun Q, Liu X, Wang P, Wu R, Ma Z. tRF-Leu-CAG promotes cell proliferation and cell cycle in non-small cell lung cancer. Chem Biol Drug Des. 2017;90(5):730–8.
Kumar P, Anaya J, Mudunuri SB, Dutta A. Meta-analysis of tRNA derived RNA fragments reveals that they are evolutionarily conserved and associate with AGO proteins to recognize specific RNA targets. BMC Biol. 2014;12:14.
Haussecker D, Huang Y, Lau A, Parameswaran P, Fire AZ, Kay MA. Human tRNA-derived small RNAs in the global regulation of RNA silencing. RNA-a Publication of the Rna Society. 2010;16(4):673–95.
Ivanov P, Emara MM, Villen J, Gygi SP, Anderson P. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol Cell. 2011;43(4):613–23.
Ivanov P, O'Day E, Emara MM, Wagner G, Lieberman J, Anderson P. G-quadruplex structures contribute to the neuroprotective effects of angiogenin-induced tRNA fragments. Proc Natl Acad Sci U S A. 2014;111(51):18201–6.
Gebetsberger J, Wyss L, Mleczko AM, Reuther J, Polacek N. A tRNA-derived fragment competes with mRNA for ribosome binding and regulates translation during stress. RNA Biol. 2017;14(10):1364–73.
Slotkin RK, Martienssen R. Transposable elements and the epigenetic regulation of the genome. Nat Rev Genet. 2007;8(4):272–85.
Schorn AJ, Gutbrod MJ, LeBlanc C, Martienssen R. LTR-retrotransposon control by tRNA-derived small RNAs. Cell. 2017;170(1):61–+.
Ruggero K, Guffanti A, Corradin A, Sharma VK, De Bellis G, Corti G, Grassi A, Zanovello P, Bronte V, Ciminale V, et al. Small noncoding RNAs in cells transformed by human T-cell leukemia virus type 1: a role for a tRNA fragment as a primer for reverse transcriptase. J Virol. 2014;88(7):3612–22.
Deng J, Ptashkin RN, Chen Y, Cheng Z, Liu G, Phan T, Deng X, Zhou J, Lee I, Lee YS, et al. Respiratory syncytial virus utilizes a tRNA fragment to suppress antiviral responses through a novel targeting mechanism. Mol Ther. 2015;23(10):1622–9.
Wang Q, Lee I, Ren J, Ajay SS, Lee YS, Bao X. Identification and functional characterization of tRNA-derived RNA fragments (tRFs) in respiratory syncytial virus infection. Mol Ther. 2013;21(2):368–79.
Zhou J, Liu S, Chen Y, Fu Y, Silver AJ, Hill MS, Lee I, Lee YS, Bao X. Identification of two novel functional tRNA-derived fragments induced in response to respiratory syncytial virus infection. J Gen Virol. 2017;98(7):1600–10.
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–33.
Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11(9):597–610.
Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009;10(10):704–14.
Esteller M. Non-coding RNAs in human disease. Nat Rev Genet. 2011;12(12):861–74.
Thompson DM, Parker R. Stressing out over tRNA cleavage. Cell. 2009;138(2):215–9.
Haiser HJ, Karginov FV, Hannon GJ, Elliot MA. Developmentally regulated cleavage of tRNAs in the bacterium Streptomyces coelicolor. Nucleic Acids Res. 2008;36(3):732–41.
Garcia-Silva MR, Frugier M, Tosar JP, Correa-Dominguez A, Ronalte-Alves L, Parodi-Talice A, Rovira C, Robello C, Goldenberg S, Cayota A. A population of tRNA-derived small RNAs is actively produced in Trypanosoma cruzi and recruited to specific cytoplasmic granules. Mol Biochem Parasitol. 2010;171(2):64–73.
Hsieh L-C, Lin S-I, Kuo H-F, Chiou T-J. Abundance of tRNA-derived small RNAs in phosphate-starved Arabidopsis roots. Plant Signal Behav. 2010;5(5):537–9.
Emara MM, Ivanov P, Hickman T, Dawra N, Tisdale S, Kedersha N, Hu G-F, Anderson P. Angiogenin-induced tRNA-derived stress-induced RNAs promote stress-induced stress granule assembly. J Biol Chem. 2010;285(14):10959–68.
Pekarsky Y, Balatti V, Palamarchuk A, Rizzotto L, Veneziano D, Nigita G, Rassenti LZ, Pass HI, Kipps TJ, Liu C-G, et al. Dysregulation of a family of short noncoding RNAs, tsRNAs, in human cancer. Proc Natl Acad Sci U S A. 2016;113(18):5071–6.
Zhao C, Tolkach Y, Schmidt D, Kristiansen G, Mueller SC, Ellinger J. 5 ‘-tRNA halves are dysregulated in clear cell renal cell carcinoma. J Urol. 2018;199(2):378–83.
Greenway MJ, Andersen PM, Russ C, Ennis S, Cashman S, Donaghy C, Patterson V, Swingler R, Kieran D, Prehn J, et al. ANG mutations segregate with familial and ‘sporadic’ amyotrophic lateral sclerosis. Nat Genet. 2006;38(4):411–3.
van Es MA, Schelhaas HJ, van Vught PWJ, Ticozzi N, Andersen PM, Groen EJN, Schulte C, Blauw HM, Koppers M, Diekstra FP, et al. Angiogenin variants in Parkinson disease and amyotrophic lateral sclerosis. Ann Neurol. 2011;70(6):964–73.
Sebastia J, Kieran D, Breen B, King MA, Netteland DF, Joyce D, Fitzpatrick SF, Taylor CT, Prehn JHM. Angiogenin protects motoneurons against hypoxic injury. Cell Death Differ. 2009;16(9):1238–47.
Kieran D, Sebastia J, Greenway MJ, King MA, Connaughton D, Concannon CG, Fenner B, Hardiman O, Prehn JHM. Control of Motoneuron survival by Angiogenin. J Neurosci. 2008;28(52):14056–61.
Wu D, Yu WH, Kishikawa H, Folkerth RD, Iafrate AJ, Shen YP, Xin WM, Sims K, Hu GF. Angiogenin loss-of-function mutations in amyotrophic lateral sclerosis. Ann Neurol. 2007;62(6):609–17.
Saikia M, Jobava R, Parisien M, Putnam A, Krokowski D, Gao X-H, Guan B-J, Yuan Y, Jankowsky E, Feng Z, et al. Angiogenin-cleaved tRNA halves interact with cytochrome c, protecting cells from apoptosis during osmotic stress. Mol Cell Biol. 2014;34(13):2450–63.
Hanada T, Weitzer S, Mair B, Bernreuther C, Wainger BJ, Ichida J, Hanada R, Orthofer M, Cronin SJ, Komnenovic V, et al. CLP1 links tRNA metabolism to progressive motor-neuron loss. Nature. 2013;495(7442):474–80.
Karaca E, Weitzer S, Pehlivan D, Shiraishi H, Gogakos T, Hanada T, Jhangiani SN, Wiszniewski W, Withers M, Campbell IM, et al. Human CLP1 mutations Alter tRNA biogenesis, affecting both peripheral and central nervous system function. Cell. 2014;157(3):636–50.
Schaffer AE, Eggens VRC, Caglayan AO, Reuter MS, Scott E, Coufal NG, Silhavy JL, Xue Y, Kayserili H, Yasuno K, et al. CLP1 founder mutation links tRNA splicing and maturation to cerebellar development and neurodegeneration. Cell. 2014;157(3):651–63.
Anderson P, Ivanov P. tRNA fragments in human health and disease. FEBS Lett. 2014;588(23):4297–304.
Guzzi N, Ciesla M, Phuong Cao Thi N, Lang S, Arora S, Dimitriou M, Pimkova K, Sommarin MNE, Munita R, Lubas M, et al. Pseudouridylation of tRNA-derived fragments steers translational control in stem cells. Cell. 2018;173(5):1204–1216.e26.
Kim HK, Fuchs G, Wang S, Wei W, Zhang Y, Park H, Roy-Chaudhuri B, Li P, Xu J, Chu K, et al. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature. 2017;552(7683):57–+.
Liang H, Gong F, Zhang S, Zhang C-Y, Zen K, Chen X. The origin, function, and diagnostic potential of extracellular microRNAs in human body fluids. Wiley Interdiscip Rev RNA. 2014;5(2):285–300.
Dhahbi JM, Spindler SR, Atamna H, Yamakawa A, Boffelli D, Mote P, Martin DIK. 5’tRNA halves are present as abundant complexes in serum, concentrated in blood cells, and modulated by aging and calorie restriction. BMC Genomics. 2013;14:14.
Dhahbi JM, Spindler SR, Atamna H, Boffelli D, Martin DI. Deep sequencing of serum small RNAs identifies patterns of 5′ tRNA half and YRNA fragment expression associated with breast Cancer. Biomarker Cancer. 2014;6:37–47.
Zhang YF, Zhang Y, Shi JC, Zhang H, Cao ZH, Gao X, Ren WH, Ning YN, Ning LN, Cao YJ, et al. Identification and characterization of an ancient class of small RNAs enriched in serum associating with active infection. J Mol Cell Biol. 2014;6(2):172–4.
Peng H, Shi J, Zhang Y, Zhang H, Liao S, Li W, Lei L, Han C, Ning L, Cao Y, et al. A novel class of tRNA-derived small RNAs extremely enriched in mature mouse sperm. Cell Res. 2012;22(11):1609–12.
Chen Q, Yan M, Cao Z, Li X, Zhang Y, Shi J, Feng G-h, Peng H, Zhang X, Zhang Y, et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science. 2016;351(6271):397–400.
Sharma U, Conine CC, Shea JM, Boskovic A, Derr AG, Bing XY, Belleannee C, Kucukural A, Serra RW, Sun F, et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science. 2016;351(6271):391–6.
Gapp K, Jawaid A, Sarkies P, Bohacek J, Pelczar P, Prados J, Farinelli L, Miska E, Mansuy IM. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat Neurosci. 2014;17(5):667–9.
Schopman NCT, Heynen S, Haasnoot J, Berkhout B. A miRNA-tRNA mix-up tRNA origin of proposed miRNA. RNA Biol. 2010;7(5):573–6.
Yanbo Wang, Hongwei Liang, Fangfang Jin, Xin Yan, Guifang Xu, Huanhuan Hu, Gaoli Liang, Shoubin Zhan, Xiuting Hu, Quan Zhao, Yuan Liu, Zhen-You Jiang, Chen-Yu Zhang, Xi Chen, Ke Zen. Injured liver-released miRNA-122 elicits acute pulmonary inflammation via activating alveolar macrophage TLR7 signaling pathway. Proceedings of the National Academy of Sciences. 2019;116(13):6162-6171.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 81872284).
Availability of data and materials
Not applicable.
Author information
Authors and Affiliations
Contributions
FFJ drafted the manuscript. ZGG contributed to designing and revising the manuscript. They both approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
We have obtained consents to publish this paper from all the participants of this study.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Jin, F., Guo, Z. Emerging role of a novel small non-coding regulatory RNA: tRNA-derived small RNA. ExRNA 1, 39 (2019). https://doi.org/10.1186/s41544-019-0036-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41544-019-0036-7