
Abstract
In the Persian Gulf, Azooxanthellate Scleractinia remain understudied compared to their symbiotic counterparts. Here, we report the presence of azooxanthellate coral Truncatoflabellum mortenseni (Cairns & Zibrowius, 1997) for the first time in Iranian deep waters. This species was found on soft sediments at depths of 83 and 101 m. Given this report, the distribution range of T. mortenseni, previously known to exist only in the Indonesian-Philippine regions, is now expanded to the Persian Gulf.
Similar content being viewed by others


Introduction
Taxonomically, members of the azooxanthellate Scleractinia occur in 14 families (Cairns and Zibrowius 1997, Cairns 2007), and those ones occurring in deep waters (> 50 m), are referred to as cold-water or deep-sea corals (Cairns 2007). Among these families, the Caryophylliidae is the largest family, followed by the Dendrophylliidae, Flabellidae, and the Turbinoliidae (Cairns 2007).
The family Flabellidae Bourne, 1905 is comprised 10 genera, all of which are exclusively solitary and are widely distributed at depths ranging from shallow waters to 3200 m (Cairns 1999, Cairns 2007). Truncatoflabellum is one of the five living transversely-dividing flabellid genera which exhibits specific morphology and reproduction cycle (i.e., transverse division in anthocauli vs. sexual in anthocyathi) (Cairns 1988, Cairns 1989, Tokuda and Ezaki 2013, Cairns 2016). Transverse division is considered one of the main modes of asexual reproduction and is restricted to certain solitary corals living in soft substrates (Cairns and Zibrowius 1997, Cairns 1988, Cairns 2007, Tokuda et al. 2010, Tokuda et al. 2017).
The genus Truncatoflabellum currently has 34 recognized species (Hoeksema and Cairns 2020), all of which are solitary and azooxanthellate. Some of its species (e.g. T. formosum) are reported from many regions such as the Indonesian-Philippine region, Indian Ocean and Australia (Cairns 1989, Cairns and Keller 1993, Cairns 1998, Cairns 2016, Tenjing et al. 2019), whereas knowledge about distribution range of some species is currently inadequate, such as T. mortenseni (described for the first time from Indonesian/Philippine regions), (Cairns and Zibrowius 1997, Cairns 2016).
In the Persian Gulf, most studies have focused on the shallow-water corals, while deep-water corals are relatively poorly studied. According to literature, the reports of deep-water corals from this region are mainly related to specimens collected during the International Indian Ocean Expedition in 1963 (see Additional file 1). The present study reports the first record of free-living coral Truncatoflabellum mortenseni from deep water of the Persian Gulf.
Materials and methods
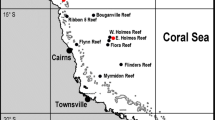
In May 2019 a scientific expedition was performed by the “Iranian National Institute for Oceanography and Atmospheric Science” (INIOAS). The objectives of the expedition were monitoring abiotic and biotic factors. In this regard, samples from both water-column and sediment were taken from 60 stations (Fig. 1). In particular, solitary coral samples were obtained from samples collected using Box-Corer (KC Denmark 1000 cm2). The coral specimens were found in two sites at depths 83 m (T6S4: 26.055552oN, 55.3548994°E) and 101 m (T0S5: 26.2100545oN, 54.458203°E), (Fig. 1). The samples were sorted onboard, preserved in 70% alcohol and transported to INIOAS’ laboratory. All coral samples were identified to the species level based on morphological traits according to available taxonomic guides (Cairns 1994, Cairns and Zibrowius 1997 and Cairns 2016). In sampling sites, temperature and salinity ranges were 22.8–20.69 °C and 39.5–40.3 PSU, respectively.

Map of the collection stations during “Persian Gulf Expedition” (PGE-1901) in Iranian waters, May 2019. The red triangles (T0S5 and T6S4) show places where Truncatoflabellum mortenseni specimens were collected
Result
A total of 20 solitary corals were identified as Truncatoflabellum mortenseni (Fig. 2). The species had laterally compressed corallum (fan shaped) which was brown to reddish color. Theca marked by fine light-brown longitudinal stripe that are equal in width. In addition, it was noted that the surface of adult anthocyathus were encrusted by small bryozoan and tube worms (Fig. 2 d-h) whereby their size indicated food restrictions in deep waters (Cairns and Keller 1993). Septa arranging in three orders consisting of 20 primary septa that reach columella and sometimes slightly exert, 20 secondary and about 40 tertiary that is short and limited to wall; total number of septa 76–80. The morphological features of anchocyathus were measured in large specimens: edge angle 45°-46°, face angle 24°-30°, height of corallum 19–22.30 mm, greater calicular diameter 20 mm, lesser scar diameter 4–5.1 mm, lesser calicular diameter 11.7–12.8 mm, greater scar diameter 6.5–8.1 mm, GSD:GCD 0.3–0.4 mm, H:GCD 0.95–1.1 mm and GCD:LCD 1.5–1.6 mm. Anthocaulus white to light-brown without thecae spines (Fig. 2 a-c), whereas anthocyatus bearing one pair of thecal spines basally. Typically, the mature anthocyatus of this species has one pair of thecal edge spine (Cairns and Zibrowius 1997, Cairns 2016), although one of the collected specimens had two pair of spines on the corallum (Fig. 1e). In one specimen, anthocyatus and anthocaulus remain together as one corallum which indicates transverse division has occurred (Fig. 2. c).

Truncatofelabellum mortenseni collected from the Persian Gulf: (a-b) Anthocaulus stages; (c) transverse division; (d-f) juvenile anthocyatus stage; (g-h) adult anthocyathus stage, their theca encrusted by tube worm and bryozoan; (i) septal arrangement and basal scale in adult anthocyathus. All scale bars: approximately 1 mm
Discussion
Although deep-water corals are reported from many regions across the Indo-Pacific Ocean, knowledge of species distribution and occurrence in the Persian Gulf remains undetermined. Truncatoflabellum mortenseni has so far been reported from Philippines/Indonesia and New Caledonia at depths between 50 and 450 m (Cairns and Zibrowius 1997, Cairns 2016). This new report extends its geographical distribution to deep waters of the Persian Gulf for the first time. Moreover, this is the deepest record of the azooxanthellate coral in the Persian Gulf.
During this survey, T. mortenseni has been found on soft sediments in two sites where amphipoda, large polychaete and bivalvia shells were present. The type of substrate is considered to be a vital factor in predicting the distribution of many deep-water corals (Tokuda et al. 2010, Kinlan et al., 2016). In this regard, different adaptive strategies have evolved in free-living corals to live on various substrates (see Tokuda et al. 2010, Tokuda and Ezaki 2013). For examples, the basal spines in T. mortenseni are considered as adaptive strategies to stabilize the life position in soft substrates (Tokuda et al. 2010). Asexual reproduction (transverse division) is another adaptive advantage for living on soft substrate (Cairns 1988, Tokuda et al. 2010, Tokuda et al. 2017), as less energy is required. Moreover, such reproduction is essential in environments where sexually reproducing units may be low in number and far in distance especially in deep-sea environments (Cairns 1988).
During the survey in Iranian waters, this species has been exclusively found in two sites even though all sampling locations were noted to have similar substrates. Failure to find this species in other locations or even more coral specimens might be related to the intensity of sampling effort or even sampling equipment. Further, the occurrence of deep-water corals can be influenced by other biological and ecological factors. Habitat suitability correlates with a variety of environmental variables including bottom geomorphology, temperature, salinity, and surface productivity (Kinlan et al., 2016).
This study and previous reports from the Red Sea (Roder et al. 2013, Qurban et al. 2014, Qurban et al. 2020) reveal that some deep-water corals are able to withstand extreme conditions where temperature and salinities are considerably higher than those reported elsewhere for deep-water coral (Roder et al. 2013, Qurban et al. 2014). Therefore, further studies are required to understand and determine the factors which influence species occurrence and distribution in the deep-water of the Persian Gulf.
Conclusion
The present report constitutes the first mention of T. mortenseni for the Persian Gulf. With the addition of T. mortenseni, a new species is added to azooxanthellate scleractinia inventory of this region. This observation indicates further investigations are needed to better understand the occurrence and distribution of deep-water corals in this region.
Availability of data and materials
The data generated during this study are included in this published article [Additional file 1]. The collected specimens are deposited at the INIOAS museum and are available upon request.
References
Cairns SD. A key to the genera and species of the transversely-dividing Flabellidae (Anthozoa, Scleractinia, Flabellidae), with a guide to the literature, and the description of two new species. ZooKeys. 2016;562:1.
Cairns SD. Deep-water corals: an overview with special reference to diversity and dis-tribution of deep-water scleractinian corals. Bull Mar Sci. 2007;81(3):311–22.
Cairns SD. Species richness of recent Scleractinia. Atoll Res Bull. 1999;459:1–12.
Cairns SD. Azooxanthellate Scleractinia (Cnidaria: Anthozoa) of Western Australia. Rec Aust Mus. 1998;18:361–417.
Cairns SD, Zibrowius H. Cnidaria Anthozoa: azooxanthellate Scleractinia from the Philipine and Indonesian regions. Mémoires du Muséum national d'Histoire naturelle. 1997.
Cairns SD. Scleractinia of the temperate North Pacific. Smithsonian Contributions to Zoology. 1994;557:150. https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=+Scleractinia+of+the+temperate+North+Pacific.+Smithsonian+Contributions+to+Zoology&btnG=.
Cairns SD, Keller NB. New taxa and distributional records of azooxanthellate Scleractinia (Cnidaria, Anthozoa) from the tropical south-west Indian Ocean,with comments on their zoogeography and ecology. Annals of the South African Museum. 1993;103:PART 5. 289p. ISSN 0303-2515.
Cairns SD. A revision of the ahermatypic Scleractinia of the Philippine Islands and adjacent waters, Part 1: Fungiacyathidae, Micrabaciidae, Turbinoliinae, Guyniidae, and Flabel-lidae. Smithsonian Contributions to Zoology. Washington, D.C. 1989;486:140.
Cairns SD. Asexual reproduction in solitary Scleractinia. Proceedings of the 6th International Coral Reef Symposium. Australia 2; 1988. p. 641–6.
Hoeksema BW, Cairns S. 2020. World list of Scleractinia. Truncatoflabellum Cairns 1989. Accessed through: world register of marine species at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=135118 on 2020-07-07.
Kinlan B, Poti M, Dorfman D, Caldow C, Drohan A, Packer D, Nizinski M. Model output for deep-sea coral habitat suitability in the U.S. North and Mid-Atlantic from 2013 (NCEIAccession 0145923). NOAA National Centers for Environmental Information. 2016. In: Zimmerman AN, Johnson CC, Bussberg NW, Dalkilic MM, editors. Stability and decline in deep-sea coral biodiversity, Gulf of Mexico and US West Atlantic. Coral Reefs; 2020. p. 1–5.
Qurban MA, Krishnakumar PK, Joydas TV, Manikandan KP, Ashraf TT, Sampath G, Thiyagarajan D, He S, Cairns SD. Discovery of deep-water coral frameworks in the northern Red Sea waters of Saudi Arabia. Sci Rep. 2020;10(1):1–8.
Qurban MA, Krishnakumar PK, Joydas TV, Manikandan KP, Ashraf TT, Quadri SI, Wafar M, Qasem A, Cairns SD. In-situ observation of deep water corals in the northern Red Sea waters of Saudi Arabia. Deep-Sea Res I Oceanogr Res Pap. 2014;89:35–43.
Roder C, Berumen ML, Bouwmeester J, Papathanassiou E, Al-Suwailem A, Voolstra CR. First biological measurements of deep-sea corals from the Red Sea. Sci Rep. 2013;3:2802.
Tenjing SY, Krishnan P, Deepak Samuel V, Purvaja R. Truncatoflabellum madrasensis sp. nov.-A new Ahermatypic coral species from the India and a checklist of species of the genus Truncatoflabellum from the Indian Ocean. VIE ET MILIEU - Life and Environment. 2019;69(2–3):89–93.
Tokuda Y, Haraguchi H, Ezaki Y. First real-time observation of transverse division in azooxa thellate scleractinian corals. Sci Rep 2017;7 (1):1–0.
Tokuda Y, Ezaki Y. Asexual reproduction of pliocene solitary scleractinian coral truncatofl bellum: a morphological and biometric study. J Paleontol. 2013;86(2):268–72.
Tokuda Y, Ikeno T, Goto SG, Numata H, Ezaki Y. Influence of different substrates on the evlution of morphology and life-history traits of azooxanthellate solitary corals (Scleractinia: Flabellidae). Biol J Linn Soc. 2010;101(1):184–92.
Acknowledgements
We wish to thank the Iranian National Institute for Oceanography and Atmospheric Science (INIOAS), and Iran National Science Foundation (INSF) for supporting and providing laboratory facility during the present study. The survey carried out on-board the vessel Kavooshgar Khalig Fars (Persian Gulf Explorer-PGE 1901- Program No: INIOAS-397-06-026-02). The authors thank the anonymous reviewers for their valuable comments on our manuscript.
Funding
This work was supported by the Iran National Science Foundation INSF under Grant [number 97004513].
Author information
Authors and Affiliations
Contributions
AM (Principal Investigator) carried out the field survey and resources. PAS identified the specimens and wrote the original draft. FO provided information about substrates and financial aid and. All authors read, revised and approved the final manuscript. The authors did not have any conflict about the order of authors’ names.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
No potential conflict of interest was reported by the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
List of azooxanthellate Scleractinia (Cnidaria, Anthozoa) reported from deep waters of the Persian Gulf and Oman Sea.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Maghsoudlou, A., Alidoost Salimi, P. & Owfi, F. First record and range extension of the transversely-dividing azooxanthellate coral Truncatoflabellum mortenseni Cairns and Zibrowius, 1997 (Scleractinia, Flabellidae) in the Persian Gulf, Western Indo-Pacific Ocean. Mar Biodivers Rec 14, 8 (2021). https://doi.org/10.1186/s41200-021-00203-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41200-021-00203-x