
Abstract
Background
Brain arteriovenous malformations (BAVMs) are abnormal vessels that are apt to rupture, causing life-threatening intracranial hemorrhage (ICH). The estimated prevalence of BAVMs is 0.05% among otherwise healthy individuals. In this study, we aim to investigate the mutational spectrum of syndromic genes in sporadic BAVM.
Methods
We recruited a cohort of 150 patients with BAVM and performed whole-exome sequencing on their peripheral blood DNA. To explore the mutational spectrum of syndromic genes in sporadic brain arteriovenous malformation, we selected six genes according to the Online Mendelian Inheritance in Man (OMIM) and literature. All variants in the six candidate genes were extracted and underwent filtering for qualifying variants.
Results
There are a total of four patients with rare variants in hereditary hemorrhagic telangiectasia-related genes. In addition, we identified two patients have the variant of RASA1 gene in our database, which are also rare mutations that are absent from population databases. However, we did not find any patients with GNAQ mutations in our database.
Conclusions
In conclusion, we demonstrated that variants in syndromic vascular malformations play important roles in the etiology of sporadic BAVM.
Similar content being viewed by others

Background
Brain arteriovenous malformations (BAVMs) are abnormal vessels that are apt to rupture, causing life-threatening intracranial hemorrhage (ICH) [1]. The estimated prevalence of brain BAVMs is 0.05% among otherwise healthy individuals [2]. BAVMs account for 25% of hemorrhagic strokes in adults younger than 50 years of age, and up to 40% of BAVM patients die or remain functionally impaired within one year after ICH [3].
Although the pathogenesis of sporadic BAVM is largely unknown, some BAVM cases are associated with hereditary hemorrhagic telangiectasia (HHT) and capillary malformation-arteriovenous malformation (CM-AVM) [4, 5]. Moreover, rare congenital syndrome is present in featuring intracranial vascular malformations, such as Sturge-Weber syndrome [6]. Approximately 30% of affected syndromic individuals also have fast-flow AVM [7]. Life-threatening complications can arise from these fast-flow lesions including hemorrhage and neurological consequences requiring transarterial embolization or surgical treatment [8].
In recent years, whole exome sequencing (WES) has developed as a reliable technology for identifying coding mutations at a genome-wide level. WES makes it possible to identify predisposing variants for rare diseases such as BAVM. De novo variants in PITPNM3, SARS, and LEMD3 have been identified in sporadic BAVM in our research previously [9]. We also identified some compound heterozygous mutations through BAVM probands and their healthy parents [10]. These results opened a new avenue to further explore the pathogenesis of BAVMs.
In this study, we selected six candidate genes related to the syndromic BAVM to investigate the mutational spectrum of syndromic genes in sporadic BAVM.
Methods
Patient recruitment
Sporadic cases were consecutively recruited into this study following the including criteria: (1) bAVM lesion confirmed by both a neurosurgeon and a neuroradiologist according. (2) With written informed consent from the proband or familial members.
Exclusion criteria were (1) known diagnoses of hereditary hemorrhagic telangiectasia, capillary malformation-AVM, Sturge-Weber syndrome, or other Mendelian vascular disorders; and (2) incomplete clinical data.
Exome sequencing
Genome DNA was extracted from the peripheral blood for all individuals and available parental samples using DNeasy Blood Kits (QIAGEN, Eastwin Scientific, Inc. Beijing, China) according to the manufacturer’s instructions. SureSelect Human All Exon V6+UTRr2 core design was used for exome capture in cases and their parents. DNA sequencing was performed on Illumina HiSeq 4000 or Novaseq platform.
Bioinformatic analysis
The variant-calling and annotation were performed by the in-house developed PUMP (Peking Union Medical college hospital Pipeline) [11, 12]. Single-nucleotide variants and internal duplications and/or deletions (indels) were called using the HaplotypeCaller of the Genome Analysis Toolkit, version 3.4.0. Annotated of the de novo, compound heterozygotes and recessive inherited variants were calculated with Gemini (version 0.19.1) for in silico subtraction of parental variants from the proband’s variants, with accounting for read number information extracted from BAM files. Computational prediction tools (GERP++ [13], CADD [14], SIFT [15], and Polyphen-2 [16]) were used to predict the conservation and pathogenicity of candidate variants. All variants were compared against publicly available databases such as the 1000 Genomes Project (http://www.internationalgenome.org/), the Exome variant server, NHLBI GO Exome Sequencing Project (ESP) (http://evs.gs.washington.edu/EVS/), and the genome aggregation database (gnomAD) (https://gnomad.broadinstitute.org/)
Candidate gene selection
To explore the mutational spectrum of syndromic genes in sporadic bAVM, we selected six genes according to the Online Mendelian Inheritance in Man (OMIM) (https://www.omim.org/) and literature. The syndromes include hereditary hemorrhagic telangiectasia (HHT, OMIM #PS187300) caused by germline, heterozygous loss of function (LoF) variants in either ENG [17], ACVRL1 [18], BMPR2 [19], or SMAD4 [20], capillary malformation-arteriovenous malformation (CM-AVM, OMIM #608354) which is caused by heterozygous LoF variants in RASA1 [21], as well as in Sturge-Weber syndrome (OMIM #185300) which is caused by somatic activating mutations in GNAQ [22].
Variant prioritization
All variants in the six candidate genes were extracted and underwent filtering for qualifying variants. We ask the variants to be either LoF variants (stop-gained, frameshift or canonical splice variants) or be deleterious missense varants/indels predicted to be deleterious (CADD score ≥ 15). Qualifying variants were also to have a minor allele frequency less than 0.001 in the gnomAD database.
Results
Cohort enrolment
A total of 150 probands with a clinical diagnosis of BAVM were consecutively enrolled between 2018 and 2020. Detailed clinical characteristics are presented in Table 1. Patients were independently reviewed by two experienced neuroradiologists to verify the diagnosis of BAVM by radiology imaging of the cerebrovascular system (magnetic resonance imaging/angiography (MRI/MRA), computed tomography angiography (CTA), and three-dimensional digital subtraction angiography (DSA)).
Mutational spectrum of gene related with HTT
According to previous studies in OMIM and literature, there are a total of four HHT-related genes included in our current study (Table 2). ENG encodes endoglin which is located on chromosome 9q and is associated with HHT1 (OMIM#187300); ACVRL1 encodes activin receptor-like kinase type 1 (ALK1), which is located on chromosome 12q and is associated with HHT2 (OMIM#600376) [17, 18]. Decapentaplegic homolog 4 (MADH4 or SMAD4) mutations can cause a syndrome which combines familial juvenile polyposis and HHT [20]. In addition, one study reported that BMPR2 gene analysis is indicated in patients affected with both HHT and heritable pulmonary arterial hypertension (HPAH) [19]. We identified 5 patients with mutations in these genes in our sporadic BAVM database.
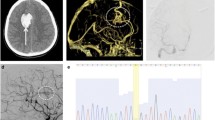
There is one patient with an ENG mutation in our cohort. BAVM1 is a 10-year-old male presented with severe headache accompanied with nausea and vomiting. Computed tomography (CT) showed subarachnoid hemorrhage (SAH). MRI showed an AVM in the area of right cerebellar hemisphere, and DSA revealed a 70*25*25mm AVM with Spetzler-Martin Grade IV (Fig. 1A). The missense variant c.901G>C from BAVM1 was predicted to be deleterious (CADD=16.89). The variant is absent from the gnomAD database (Table 3), indicating that it is extremely rare in the general population.

MRI, CT, and DSA imaging of patients carrying variants in syndromic genes. A Patient BAVM1 with ENG gene mutation c.901G>C. CT demonstrated subarachnoid hemorrhage in the distribution of posterior circulation, and DSA showed an AVM in the area of right cerebellar hemisphere. B Patient BAVM2 with ACVRL1 gene mutation c.1103C>T. MRI showed a hematoma and DSA showed an AVM in the right temporal lobe. C Patient BAVM3 with BMPR2 gene mutation c.775C>T. DSA revealed an AVM in the right side of the parietal lobe (black arrow) and an intracranial aneurysm in the internal carotid artery (white arrow). D, E Mutations of BMPR2 c.2678G>A shared by BAVM4 and BAVM5. DSA showed an AVM in the left parietal lobe in two patients. F, G Patient BAVM6 and BAVM7with RASA1 gene mutations c.1280G>A and c.3007G>A, respectively. MRI and DSA showed an AVM in the right basal ganglia and right parietal cerebral falx on two patients
A missense mutation c.1103C>T of ACVRL1 is identified in another patient BAVM2. The bioinformatics tools predict that this mutation is damaging (CADD=29.2). It does not exist in gnomAD East Asian (EAS) database (Table 3). Patient BAVM2 is a 15-year-old female who presented with headache, vomiting, and epilepsy. MRI showed a hematoma in the right temporal lobe and DSA revealed a 40*15*10mm AVM with Spetzler-Martin Grade III (Fig. 1B).
Three patients have mutations in the BMPR2 gene. The missense variant c.775C>T is absent from the gnomAD database (Table 3). Patient BAVM3 is a 23-year-old female presented with mild headache and epilepsy. DSA revealed a 60*45*30mm AVM with Spetzler-Martin Grade III in the right side of the parietal lobe and a intracranial aneurysm in the internal carotid artery (Fig. 1C). A research has shown that mutations of BMPR2 can cause the Loeys-Dietz syndrome, which is an inherited connective tissue disorder can accompanied with aneurysms [23]. Two patients BAVM4 and BAVM5 have the same missense mutation (c.2678G>A). The absence of this variant in the gnomAD database and the enrichment of this variant in our cohort support the association between c.2678G>A and BAVM. Interestingly, both patients had arteriovenous malformations in the left parietal lobe (Fig. 1D, E), which further strengthened the evidence for the pathogenicity of variants in this gene in BAVM. Therefore, our findings may represent a phenotypic expansion for this gene.
Mutational spectrum of gene related with CM-AVM
Capillary malformation-arteriovenous malformation (CM-AVM) syndrome is an autosomal dominant disorder due to germline heterozygous mutations in the RASA1 gene [21] (Table 2). Some studies have observed that 3% of the malformations occurred in the cerebral. We identified two patients have the variant of RASA1 gene in our database. They are both rare mutations (MAF<0.001) that are absent from the gnomAD database. CADD prediction indicated the mutations to be harmful (Table 3). BAVM6 is a 28-year-old male presented with severe headache accompanied with vomiting. DSA showed an AVM in the area of right basal ganglia (Fig. 1F). BAVM7 is a 27-year-old female presented with paroxysmal headache accompany with dizziness about 1 year, aggravating for 2 months. DSA showed a right parietal cerebral falx 28*30*25mm AVM belong to Spetzler-Martin Grade III (Fig. 1G). Our research indicates a more complex disease trait with potential digenic involvement in these patients.
Sturge-Weber syndrome
Sturge-Weber syndrome is a rare neurovascular disorder associated with seizures, capillary malformation, cognitive impairments, and stroke-like episodes (SLE), arising from a somatic activating mutation in GNAQ [22] (Table 2). Brain arteriovenous malformations can be found in patients with Sturge-Weber syndrome [24]. We did not find any patients with GNAQ mutations in our database. Therefore, we did not identify correlation between the sporadic BAVM and Sturge-Weber syndrome, which might be associated with the limited sample size.
Discussion
In this study, we utilized a WES dataset generated from a cohort of sporadic patients with BAVM. By inspecting genes associated with known vascular malformation syndromes, we found that deleterious variants in these genes are likely involved in the pathogenesis of sporadic BAVM. None of these variants have been reported in individuals with syndromic forms of BAVM or healthy individuals from public databases except two missense variants in RASA1.
The involvement of syndromic genes in sporadic cases have been reported in intellectual disability, which is difficult to diagnosis because of the absence of morphological clues and appropriate screening tools [25]. In a reported study, child-parent trios from ten centers in Switzerland and Germany were recruited. Of the 51 cases enrolled, 16 cases had de novo variants occurred in established intellectual disability-associated genes, including STXBP1, SCN2A, and SYNGAP1 [25]. Interestingly, some patients did not manifest the expected syndromic conditions associated with their genetic variants, implicating a strong over-description of the present clinical syndromes [25], and an obscure boundary between syndromic and sporadic conditions. Another study reported a stop-gain variant Q829X in OTOF, which is associated with auditory neuropathy, in patients with prelingual non-syndromic hearing loss [26]. The authors investigated 28 independent trios in Spain and genotyped the subjects for microsatellite markers. After identifying the linkage of the disease to a certain genetic marker, they sequenced the surrounding genome regions and identified this novel variant in OTOF. This variant is the third most frequent variant which cause the prelingual deafness so far reported in the Spanish population [26].
The underlying mechanism underlying these findings may be the overlapping biological pathways perturbed in both syndromic and sporadic conditions. The pathogenesis of HHT is associated with the perturbation of BMP/TGF-β signaling pathway. TGF-β signaling is considered as crucial pathway in the physiological process of the vessels. Specifically, it has been shown to control proliferation, differentiation, apoptosis, migration, extracellular matrix (ECM) remodeling, immune functions, and tumor invasion/metastasis [27, 28]. Once TGF-β signaling activation occurs extracellularly, it is able to interact with and complex the type I and type II serine/threonine kinase receptors at the cell surface. During skeletal muscle development, the constitutively active type II receptor phosphorylates and activates the type I receptor, which in turn directly phosphorylates Smad3 to initiate signal transduction through the canonical cascades [29, 30]. In the non-canonical pathway, the type I receptor phosphorylates signal proteins that are involved in the activation of the mitogen-activated protein kinases (MAPKs), including non-canonical signaling through extracellular signal-related kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 [31, 32]. Activated non-canonical signaling then regulate transcription factors and/or the Smad proteins through direct interactions or via downstream proteins to promote vascular differentiation, migration, and ECM remodeling [33]. Our results suggest that perturbation of BMP/TGF-β also underly the pathogenesis of isolated BAVM.
The pathogenesis of CM-AVM is associated with the Ras-Raf-MEK-ERK pathway. This pathway is initiated by an extracellular mitogen which binds to the receptor on the cell membrane, which allows the small GTPase Ras to swap the GDP for the GTP [34]. The activated Ras then activates the activity of protein kinase of the RAF kinase. Then, the RAF kinase activates MEK, i.e., MEK1 and MEK2 by phosphorylating them [35]. MEK then phosphorylates and activates the mitogen-activated protein kinase (MAPK). After that, MAPK regulates the activation of several transcriptional factors [36]. MAPK also controls the transcription of C-Fos gene and leads to changes in transcription of genes key for the cell cycle, which is important for the normal growth of the vascular system [37].
Conclusions
In conclusion, we demonstrated that variants in syndromic vascular malformations play important roles in the etiology of sporadic BAVM. These may be due to the shared biological pathways between syndromic and sporadic forms of the disease.
Availability of data and materials
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- BAVM:
-
Brain arteriovenous malformation
- MAPK:
-
Mitogen-activated protein kinase
- ERK:
-
Extracellular signal-related kinase
- JNK:
-
c-Jun N-terminal kinase
- ICH:
-
Intracranial hemorrhage
- HHT:
-
Hereditary hemorrhagic telangiectasia
- WES:
-
Whole exome sequencing
- CM-AVM:
-
Capillary malformation-arteriovenous malformation
- LoF:
-
Loss of function
- OMIM:
-
Online Mendelian Inheritance in Man
- MRI/MRA:
-
Magnetic resonance imaging/angiography
- CTA:
-
Computed tomography angiography
- DSA:
-
Digital subtraction angiography
References
Kim H, Su H, Weinsheimer S, Pawlikowska L, Young W. Brain arteriovenous malformation pathogenesis: a response-to-injury paradigm. Acta Neurochir Supplement. 2011;111:83–92. https://doi.org/10.1007/978-3-7091-0693-8_14.
Morris Z, et al. Incidental findings on brain magnetic resonance imaging: systematic review and meta-analysis. BMJ (Clinical research ed.). 2009;339:b3016. https://doi.org/10.1136/bmj.b3016.
van Beijnum J, et al. Outcome after spontaneous and arteriovenous malformation-related intracerebral haemorrhage: population-based studies. Brain J Neurol. 2009;132:537–43. https://doi.org/10.1093/brain/awn318.
Barbosa Do Prado L, Han C, Oh S, Su H. Recent Advances in Basic Research for Brain Arteriovenous Malformation. Int J Mol Sci. 2019;20. https://doi.org/10.3390/ijms20215324.
Revencu N, et al. Parkes Weber syndrome, vein of Galen aneurysmal malformation, and other fast-flow vascular anomalies are caused by RASA1 mutations. Hum Mutat. 2008;29:959–65. https://doi.org/10.1002/humu.20746.
Gökçe E, Acu B, Beyhan M, Celikyay F, Celikyay R. Magnetic resonance imaging findings of developmental venous anomalies. Clin Neuroradiol. 2014;24:135–43. https://doi.org/10.1007/s00062-013-0235-9.
Hershkovitz D, Bercovich D, Sprecher E, Lapidot M. RASA1 mutations may cause hereditary capillary malformations without arteriovenous malformations. Brit J Dermatol. 2008;158:1035–40. https://doi.org/10.1111/j.1365-2133.2008.08493.x.
Lacalm A, et al. Prenatal diagnosis of cerebral and extracerebral high-flow lesions revealing familial capillary malformation-arteriovenous malformation (CM-AVM) syndrome. Ultrasound Obstet Gynecol. 2018;51:409–11. https://doi.org/10.1002/uog.17460.
Wang K, et al. Perturbations of BMP/TGF-beta and VEGF/VEGFR signalling pathways in non-syndromic sporadic brain arteriovenous malformations (BAVM). J Med Genet. 2018;55:675–84. https://doi.org/10.1136/jmedgenet-2017-105224.
Zhang M, et al. Exome sequencing of 112 trios identifies recessive genetic variants in brain arteriovenous malformations. J Neurointerventional Surg. 2020. https://doi.org/10.1136/neurintsurg-2020-016469.
Zhao S, et al. Diagnostic yield and clinical impact of exome sequencing in early-onset scoliosis (EOS). J Med Genet. 2021;58:41–7. https://doi.org/10.1136/jmedgenet-2019-106823.
Chen N, Zhao S, Jolly A, Wang L, Pan H, Yuan J, et al. Perturbations of genes essential for Müllerian duct and Wölffian duct development in Mayer-Rokitansky-Küster-Hauser syndrome. Am J Hum Genet. 2021;108(2):337-345. https://doi.org/10.1016/j.ajhg.2020.12.014.
Davydov EV, et al. Identifying a high fraction of the human genome to be under selective constraint using GERP++. PLoS Comput Biol. 2010;6:e1001025. https://doi.org/10.1371/journal.pcbi.1001025.
Kircher M, et al. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. 2014;46:310–5. https://doi.org/10.1038/ng.2892.
Vaser R, Adusumalli S, Leng SN, Sikic M, Ng PC. SIFT missense predictions for genomes. Nat Protoc. 2016;11:1–9. https://doi.org/10.1038/nprot.2015.123.
Adzhubei IA, et al. A method and server for predicting damaging missense mutations. Nat Methods. 2010;7:248–9. https://doi.org/10.1038/nmeth0410-248.
McAllister KA, et al. Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat Genet. 1994;8:345–51. https://doi.org/10.1038/ng1294-345.
Johnson DW, et al. Mutations in the activin receptor-like kinase 1 gene in hereditary haemorrhagic telangiectasia type 2. Nat Genet. 1996;13:189–95. https://doi.org/10.1038/ng0696-189.
Rigelsky CM, et al. BMPR2 mutation in a patient with pulmonary arterial hypertension and suspected hereditary hemorrhagic telangiectasia. Am J Med Genet A. 2008;146A:2551–6. https://doi.org/10.1002/ajmg.a.32468.
Gallione CJ, et al. A combined syndrome of juvenile polyposis and hereditary haemorrhagic telangiectasia associated with mutations in MADH4 (SMAD4). Lancet. 2004;363:852–9. https://doi.org/10.1016/S0140-6736(04)15732-2.
Eerola I, et al. Capillary malformation-arteriovenous malformation, a new clinical and genetic disorder caused by RASA1 mutations. Am J Hum Genet. 2003;73:1240–9. https://doi.org/10.1086/379793.
Shirley MD, et al. Sturge-Weber syndrome and port-wine stains caused by somatic mutation in GNAQ. N Engl J Med. 2013;368:1971–9. https://doi.org/10.1056/NEJMoa1213507.
Wellbrock J, et al. Overexpression of Gremlin-1 in patients with Loeys-Dietz syndrome: implications on pathophysiology and early disease detection. PloS one. 2014;9:e104742. https://doi.org/10.1371/journal.pone.0104742.
Nishino K, Ito Y, Sorimachi T, Shimbo J, Fujii Y. Sturge-Weber syndrome associated with arteriovenous malformation in a patient presenting with progressive brain edema and cyst formation. J Neurosurg Pediatr. 2010;5:529–34. https://doi.org/10.3171/2010.1.Peds09140.
Rauch A, et al. Range of genetic mutations associated with severe non-syndromic sporadic intellectual disability: an exome sequencing study. Lancet. 2012;380:1674–82. https://doi.org/10.1016/S0140-6736(12)61480-9.
Migliosi V, et al. Q829X, a novel mutation in the gene encoding otoferlin (OTOF), is frequently found in Spanish patients with prelingual non-syndromic hearing loss. J Med Genet. 2002;39:502–6. https://doi.org/10.1136/jmg.39.7.502.
Wu M, Chen G, Li YP. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016;4:16009. https://doi.org/10.1038/boneres.2016.9.
Burks TN, Cohn RD. Role of TGF-β signaling in inherited and acquired myopathies. Skelet Muscle. 2011;1:19. https://doi.org/10.1186/2044-5040-1-19.
Liu D, Black BL, Derynck R. TGF-beta inhibits muscle differentiation through functional repression of myogenic transcription factors by Smad3. Genes Dev. 2001;15:2950–66. https://doi.org/10.1101/gad.925901.
Shi Y, Massagué J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113:685–700. https://doi.org/10.1016/s0092-8674(03)00432-x.
Rahimi RA, Leof EB. TGF-beta signaling: a tale of two responses. J Cell Biochem. 2007;102:593–608. https://doi.org/10.1002/jcb.21501.
Zhang YE. Non-Smad pathways in TGF-β signaling. Cell Res. 2009;19:128–39. https://doi.org/10.1038/cr.2008.328.
Guo X, Wang XF. Signaling cross-talk between TGF-beta/BMP and other pathways. Cell Res. 2009;19:71–88. https://doi.org/10.1038/cr.2008.302.
Orton RJ, et al. Computational modelling of the receptor-tyrosine-kinase-activated MAPK pathway. Biochem J. 2005;392:249–61. https://doi.org/10.1042/BJ20050908.
Schulze WX, Deng L, Mann M. Phosphotyrosine interactome of the ErbB-receptor kinase family. Mol Syst Biol. 2005;1:2005–0008. https://doi.org/10.1038/msb4100012.
Downward J. Targeting RAS signalling pathways in cancer therapy. Nat Rev Cancer. 2003;3:11–22. https://doi.org/10.1038/nrc969.
McCubrey JA, et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim Biophys Acta. 2007;1773:1263–84. https://doi.org/10.1016/j.bbamcr.2006.10.001.
Acknowledgements
We thank Beijing Ekitech Co., Ltd., for support in bioinformatic analyses.
Funding
This study was sponsored by the National Key Research and Development Plan of China (grant number: 2016YFC1300800), the National Natural Science Foundation of China (grant numbers: 81901178, 82072036 and 81822030), the Special Research Project for Capital Health Development (grant number: 2018-4-1077), and Beijing Natural Science Foundation (JQ20032).
Author information
Authors and Affiliations
Contributions
KW, NW, and XY performed the study design and organization; KW, MZ, YZ, JL, and YZ performed the patient recruitment; KW, SZ, MZ, and ZX drafted the manuscript; YZ, JL, YZ, XY, and NW critically revised the manuscript. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All procedures in this study were approved by the ethics committee at Tiantan Hospital (KY2017-017-01). Written informed consent was provided by each participant.
Consent for publication
All the participants gave consent for publication of their deidentified clinical information.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, K., Zhang, M., Zhao, S. et al. Mutational spectrum of syndromic genes in sporadic brain arteriovenous malformation. Chin Neurosurg Jl 8, 4 (2022). https://doi.org/10.1186/s41016-022-00270-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41016-022-00270-8