
Abstract
Background
For more than a century we have known that a high diversity of arthropod species lives in close relationship with army ant colonies. For instance, several hundred guest species have been described to be associated with the Neotropical army ant Eciton burchellii Westwood, 1842. Despite ongoing efforts to survey army ant guest diversity, it is evident that many more species await scientific discovery.
Results
We conducted a large-scale community survey of Eciton-associated symbionts, combined with extensive DNA barcoding, which led to the discovery of numerous new species, among them a highly specialized histerid beetle, which is formally described here. Analyses of genitalic morphology with support of molecular characters revealed that the new species is a member of the genus Nymphister. We provide a literature review of host records and host-following mechanisms of Eciton-associated Haeteriinae demonstrating that the new species uses an unusual way of phoretic transport to track the nomadic habit of host ants. Using its long mandibles as gripping pliers, the beetle attaches between the ants’ petiole and postpetiole. The beetles specifically attach to medium-sized ant workers, thus participating as hitchhikers in the regular colony emigrations of the single host species Eciton mexicanum Roger, 1863.
Conclusion
By providing tools for reliable species identification via morphology and DNA barcodes for hitherto unknown army ant guest species, we set the baseline for studies targeting the ecological and evolutionary dynamics in these species-rich host-symbiont communities.
Similar content being viewed by others

Background
Ants are among the most ubiquitous animals in terrestrial habitats, turning over a substantial amount of energy [1, 2]. This attracts diverse organisms engaging with ants in various interactions, ranging from mutualistic associations, as in ant-aphid interactions, to destructive predation of ant colonies by some vertebrates [1, 3, 4]. The most speciose group of ant-seeking organisms are arthropods with an estimated number of 10,000–100,000 species [5, 6]. Life in and around ant colonies is expected to be beneficial for arthropod guests, especially those that have managed to get along well with ants [1]. Among the latter are highly integrated species that inhabit ant nests, in which they are protected from their own predators, live in a stable microclimate, and have access to rich food sources [1, 7, 8]. Not surprisingly, exploiting ant societies is an evolutionarily ancient phenomenon [9–11] that dates back ~50 Ma as indicated by a recently described staphylinid beetle preserved in amber [5]. Those organisms that depend on ants for their own survival, at least during a part of their lifecycle, are known as ‘myrmecophiles’ (‘ant lovers’, [12]).
Some ant species are host to a particularly diverse myrmecophile fauna [3, 13–19]. Among them are the army ants [1, 3, 15]. As important arthropod predators, army ants are considered as keystone species in tropical forests [3, 20–22]. During raids, tens of thousands to millions of army ant workers roam the forest floor in search for arthropod prey [3, 22]. The retrieval of large amounts of high quality food [23] is supposedly a key factor explaining the abundance of army ant-associated species, which includes a diverse fauna of arthropods including millipedes, mites, bristletails, collembolans, beetles, and phorid flies [3, 15, 24, 25]. Army ant myrmecophiles often show astonishing adaptations [3] such as chemical and morphological mimicry of ant workers [3, 8, 24, 26, 27], protective morphologies [3, 7, 8, 28], as well as life history adaptations to the hosts’ life cycles [29, 30]. Neotropical army ant colonies undergo highly stereotypical cycles in which they alternate between a stationary and a nomadic phase, which, in Eciton, last about 3 and 2 weeks, respectively [3, 21, 25]. The colony stays at the same site during the stationary phase, and emigrates to a new nest site every night during the nomadic phase. Since army ants are nomadic, myrmecophiles exploiting their societies share the need to track their host colonies during the frequent colony emigrations to new bivouac sites [3, 21, 22]. They achieve this by riding on or being carried by the ants, or by walking independently on the ants’ emigration trail [3, 24, 30–33].
Beetles are among the most diverse groups of army ant myrmecophiles, and within beetles the families with the largest number of myrmecophilous species are the Staphylinidae and the Histeridae [8, 15]. Members of both beetle families make use of their glandular chemistry, which includes defensive and ant-appeasing compounds, allowing them to deal with host ants [7, 8]. Trichomes, for example, are found in members of both families. They are groups of setae that deliver glandular compounds which are licked by ant workers, thus facilitating peaceful interactions [8, 34]. While most staphylinid beetles also make use of their agility [8], histerid beetles rather rely on a protective morphology [28]. They have a robust, heavily chitinous body, and appendages that can be retracted into shallow grooves so that no potential points of attack are exposed to the host ants [8, 35] (e.g., Fig. 1). Interactions between histerids and army ants span from more facultative ones in species exploiting ant refuse deposits, to host-dependent ones in species that live within the army ant bivouac nest among the thousands of workers. Among the latter are highly integrated species (sensu [7]) with apparent behavioral and morphological adaptations [3, 7, 8, 28, 34]. For most army ant-associated histerids, however, the basic life history as well as the nature of the association is unknown.

Phoretic transport in Nymphister kronaueri. a Lateral and b dorsal view of Eciton mexicanum s. str. workers with N. kronaueri attached between the ants’ petiole and postpetiole. c, d Close-ups showing N. kronaueri grasping the trunk between petiole and postpetiole with its mandibles. Note how a the legs and c, d the antennae are completely retracted in surface depressions, a common feature in army ant-associated histerid beetles. Image credits: a Daniel Kronauer, b Munetoshi Maruyama
In the present study we describe a new histerid species from the subfamily Haeteriinae, whose members seem to be all associated with ants or termites [8, 36]. The new species uses phoretic transport to follow the frequent host colony emigrations. Using their strong mandibles, these beetles attach to the ants’ narrow waist between thorax and gaster allowing the beetles to be transported as hitchhikers to the ants’ new bivouac site (Fig. 1). Inspection of male genitalia with support of DNA barcoding data classified the new species to the genus Nymphister Reichensperger, 1933 [37]. Three species were previously classified in this genus: N. monotonus (Reichensperger, 1938) [38], N. rettenmeyeri Tishechkin and Mercado, 2012 [39], and N. simplicissimus Reichensperger, 1933 [37] (for host records see Table 1). Here we provide a taxonomic description of the new species, report its unusual mechanism of phoretic transport, and discuss some of its exceptional morphological characteristics in light of convergent trait evolution.
Methods
Origin of specimens and taxonomic history
Ants and their associates were collected at several tropical sites (see Additional file 1: Table S1). For one site, La Selva Biological Station (LSBS) in Costa Rica (N10 25.847 W84 00.404, altitude 67 m), the only location where the new species was found, we provide comprehensive community data that allowed us to assess the specificity of host-myrmecophile interactions in great detail (for more information on collection methods see [30]). At LSBS we collected myrmecophiles during host colony emigrations from all of the six local Eciton species: E. burchellii foreli Mayr, 1886 (12 colonies), E. dulcium crassinode Borgmeier, 1955 (11 colonies), E. hamatum Fabricius, 1781 (10 colonies), E. lucanoides conquistador Weber, 1949 (2 colonies), E. mexicanum s. str. Roger, 1863 (10 colonies) and E. vagans angustatum Roger, 1863 (8 colonies). For ant identifications we used the keys and species distribution maps of Watkins [40, 41] and Longino [42]. The sampling took place from February to April 2013 and March to April 2014.
Morphological protocol
The species description follows the conventional terminology as used in current histerid taxonomic literature (e.g., [39, 43–47]). Body parts were measured using a Zeiss Stemi SV 11 stereomicroscope with an ocular micrometer. Measurements represent the range (minimum and maximum), are grouped at the beginning of the description to facilitate comparisons, and are abbreviated as follows: L (mm – dorsal length along midline from anterior pronotal margin to posterior elytral margin); W (mm – width at widest point); E/PnL (ratio – elytral length/pronotal length); E/PnW (ratio – elytral width/pronotal width); Pn W/L (ratio – pronotum width/length); EL/W (ratio – elytral length/width); Pr/Py (ratio – propygidium length/pygidium length); Thoracic sterna – pro-, meso-, metasternum (mm – lengths along midline); Tibiae – pro, meso, meta (mm - straight line length from base to apex, ignoring curvature). Primary type label information is listed verbatim, with the different labels separated by forward slashes.
We realized during collections that the new species was found primarily attached to workers of a certain size. To quantify this, we exclusively collected ant workers with the new beetle species attached for two different colonies. The head widths of those ant workers were then compared to a sample of workers from the same colonies that covered the entire body size range. Note that the ants used to assess body size were not random samples; rather they were obtained to gain an estimate of the range of body sizes present within a colony. A Leica Z16 APO stereomicroscope equipped with a Leica DFC450 camera and the processing software Leica Application Suite (version 4) were used to take frontal images of ant heads. A Nikon stage micrometer was used to calibrate the scale bars. Head width was defined as the straight line between the most distant points of the head, which was a line through the middle of the eyes. It was measured using the software ImageJ (version 1.47).
Molecular protocol
Morphological analysis was supplemented with genetic analysis for three reasons. First, we applied a broad-scale DNA barcoding approach to the entire Eciton-associated myrmecophile fauna (unpublished data and [30, 48]), which includes species from different taxonomic groups including millipedes, mites, silverfish, phorid flies, beetles, and wasps. DNA barcoding helped us to sort the specimens into pre-defined units, which were then carefully inspected by taxonomic experts. This streamlines the identification of species. Second, DNA barcoding allowed us to screen for possible cryptic diversity [30, 48]. Third, COI barcoding supported the generic placement of the new species, which was originally achieved by comparing morphological genus level synapomorphies [46]. While DNA barcoding is less effective in resolving taxonomic units above the species level [49, 50], it can still predict the taxon affiliation of an undescribed species at lower taxonomic scales, such as the genus, quite well [51].
We amplified the animal barcoding gene cytochrome oxidase I (COI) [52]. Forty-four specimens of the new species from eight different E. mexicanum colonies were genetically analyzed (range: 1–14 specimens per colony, mean = 5.5 ± 3.74 specimens per colony). In total, we performed genetic analyses on 58 histerid specimens including taxa expected to be closely and distantly related to our candidate genus for the new species, i.e. Nymphister. These taxa were chosen based on phylogenetic analyses of morphological and genetic characters of the subfamily Haeteriinae [34, 46, 53], as well as on specimen availability.
DNA was extracted from whole specimens using the QIAGEN® DNeasy® Tissue Kit for 96-well plates. We followed the standard protocol except for a shortened digestion step of 2–3 h, which allowed us to keep all samples as voucher specimens in absolute ethanol. The mitochondrial COI barcode region (658 base pairs (bp)) was amplified in standard polymerase chain reactions (PCRs). For N. kronaueri specimens (for species identifications see result section) we used the primers LCO1490 and HCO2198 [54]. These primers did not amplify consistently for other taxa, and therefore we used a combination of newly designed primers that are located a few base pairs inwards of the COI barcode fragment: Ecc1F (5’AACYTTATAYTTTATCTTTGGNGCWT3’; forward primer), Ecc2F (5’GCAGGAATAGTAGGAACATCTCTTAG3’; forward primer) and Ecc1R (5’AAWAGRTGTTGRTATARAATAGGGTC3’, reverse primer). PCRs were set up as described previously [30]. Purification and sequencing of PCR products were outsourced to Macrogen USA. All PCR products were sequenced in both directions.
Analysis of molecular data
Workflows, including information about collection, DNA extraction, PCR and DNA sequencing, were tracked with the laboratory management software Geneious (version R10.0.5) (http://www.geneious.com, [55]) with the plugin ‘biocode’ (version 3.0.1) [56]. Sequences were trimmed and aligned in Geneious. A neighbor-joining tree based on Tamura-Nei distances with bootstrap support (1000 replicates) was created using the Geneious Tree Builder. To support the generic placement of the new species, we were seeking for the most similar sequence in a set of potential candidates using a distance-based tree method. Note that our approach did not intend to fully resolve the phylogenetic relationships of the species studied here.
Data/specimen storage and research permits
In total, 67 voucher images from 19 specimens including dorsal, ventral, lateral views, as well as frontal images of the head were uploaded together with collection records and DNA sequences to the Barcode of Life database (http://www.boldsystems.org/; Additional file 1: Table S1). GenBank accession numbers for COI sequences can be found in Additional file 1: Table S1. Images were taken with the Leica Z16 APO stereomicroscope described above and processed using the software Helicon Focus (v. 5.3). Specimens were deposited at the following institutions: the Louisiana State Arthropod Museum, Baton Rouge, USA (LSAM), the National Museum of Costa Rica, San Jose, Costa Rica (MNCR), the Field Museum of Natural History, Chicago, USA (FMNH), the Connecticut State Museum of Natural History, USA (CSMNH), and the private collection of A. Allen, Boise, ID, USA (AA). Ant workers of all colonies are vouchered in CvB’s personal collection. Research and specimen export permits for Costa Rica were issued by the ‘Ministry of the Environment, Energy and Technology’ (MINAET; permit numbers: 192-2012-SINAC and R-009-2014-OT-CONAGEBIO).
Nomenclatural act
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature (ICZN), and hence the new species name contained herein is available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank Life Science Identifiers (LSID) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/“. The LSID for this publication is: urn:lsid:zoobank.org:pub:0D0FD20C-D73E-4326-B574-F3523EBD96F4.
Results
Species description Nymphister kronaueri von Beeren & Tishechkin sp. nov. (LSID: urn:lsid:zoobank.org:act:B2736288-50F7-45A0-B87A-02222237B201)
Material examined
Holotype (Fig. 2a-d). A male, mounted on a point and labeled “COSTA RICA: Heredia, ~3.5 km SE Puerto Viejo, La Selva Biological Station 10.4305°N 84.0064°W /Eciton mexicanum emigration, attached to workers. 15. April 2013. C. von Beeren & D. Kronauer. cvb722higa001/LSAM0263619/HOLOTYPE Nymphister kronaueri sp. n. A. K. Tishechkin & C. von Beeren des. 2015” (FMNH).

Nymphister kronaueri habitus and male and female genitalia. a Dorsal, b lateral, c ventral view and d frontal head view of the Nymphister kronaueri sp. nov. holotype (male). Scale bars are 1 mm except for d where it is 0.2 mm. Male genitalia: e Ventral and f dorsal view of the 8th sternite-tergite complex. g Dorsal and h lateral view of aedeagus. i Lateral and j ventral view of the 9th tergite and k ventral and l lateral view of the 9th sternite. Images are at the same scale. Female genitalia: m Ventral view of female genitalia with 8th sternite omitted. n Ventral view of the 8th sternite. Images are at the same scale
Paratypes (N = 20). Collected at the same locality on 04.iii.2014: cvb683higa001-cvb683higa003 (all LSAM); 19.iii.2014: cvb591higa003 (MNCR), cvb591higa004, cvb591higa005 (both FMNH), cvb591higa006 (LSAM); 20.iii.2014: cvb594higa002 (FMNH), cvb594higa005, cvb594higa008-cvb594higa009 (all LSAM), cvb594higa006, cvb594higa007 (both CSMNH); 21.iii.2014: cvb597higa002, cvb597higa003, cvb597higa005, cvb597higa006 (all LSAM), cvb597higa003 (AA); 04.iv.2014: cvb692higa001 (MNCR), cvb692higa003 (FMNH); all specimens were collected during Eciton mexicanum s. str. emigrations by C. von Beeren, S. Pohl, G. Burke and D. Kronauer (for more details see Additional file 1: Table S1).
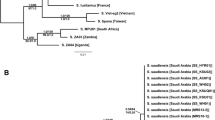
Differential Diagnosis. The long basal piece of the aedeagus combined with short parameres (Fig. 2g, h) placed N. kronaueri into the tribe Nymphistrini [46]. Within Nymphistrini, Nymphister kronaueri can easily be distinguished from all other globular, subspherical species with short, broad legs and absence of trichomes by its peculiar frontal structure, i.e. the fronto-clypeal margin is keeled and frontal and clypeal planes meet at an angle of almost 90° (Figs. 1d and 2d). We assigned the new species to the genus Nymphister based on a comparative morphological analysis. In the most recent and complete key to the genera of Nymphystrini [46], the new species ran to couplet 73/74 (pp. 158–159). However, it could not be identified there as Nymphister because of the presence of both, dorsal striae and dense body surface punctation (Table 2; Fig. 2a-c). Besides these characters and the presence of dorsal setae, all other external characters, e.g. the overall globular body shape with short legs (Fig. 2a-c), the prosternal keel (Fig. 2c), the meso- and metasternite structures, and especially the development of lateral metasternal striae (Fig. 2c) corresponded perfectly with typical Nymphister species (for an overview and additional characters see Table 2). In particular, the following genitalic characters, which represent putative generic synapomorphies, allowed classification of the new species as a member of the genus Nymphister: the reduced halves of the male 8th sternite bearing velae (Fig. 2e, f), the overall structure of the male 9th tergite (Fig. 2i, j), the shape of the parameres and the basal piece of the aedeagus (Fig. 2g, h), the deeply cleft 8th female sternite with its well-developed basal bridge (Fig. 2n), the split 8th female tergite, the shape and the degree of connections of female coxites and the 9th tergite (Fig. 2m) (for an overview see Table 2). A COI neighbor-joining tree additionally supported that the new species is genetically most closely affiliated to the genus Nymphister (Fig. 3).

Molecular support for the generic classification of Nymphister kronaueri. Shown is a Neighbor-Joining Tree based on Tamura–Nei distances (scale bar) for COI barcodes of members of the tribe Haeteriini (Histeridae: Haeteriinae). Margarinotus striola (Histeridae: Histerinae: Histerini) served as an outgroup (GenBank accession number: KM448233.1 [76]). Bootstrap support values (1000 replicates) are given at major nodes. Branches of bootstrap support values ≤ 50 were collapsed and shown as a polytomy
Description. For all measurement ranges Nspecimens = 21: L: 1.40–1.58; W: 1.18–1.40; E/Pn L: 1.00–1.35; E/Pn W: 1.02–1.15; Pn W/L: 1.11–1.30; E L/W: 0.59–0.71; Pr/Py: 1.10–1.19; pro-, meso-, and metasterna, respectively: 0.38–0.42, 0.06–0.09, 0.32–0.38; pro-, meso-, and metatibiae, respectively: 0.48–0.54, 0.52–0.58, 0.64–0.68. Body round, convex dorsally, subspherical, dark reddish-brown; body surface with fine punctate-alutaceous microsculpture throughout; punctures on dorsal surface each bearing a pale yellow, relatively short, relatively inconspicuous seta (see Fig. 2a-d).
Frons flat, impunctate except for four small setiferous punctures along the occipital border; frontal stria narrowly interrupted in the middle, with prominent latero-marginal frontal carina, supraorbital stria absent. Fronto-clypeal juncture carinate, clypeus distinctly concave, its plane perpendicular to the frontal plane, clypeo-labral suture indistinct throughout (Fig. 2d). Mandibles wide, stout, with weakly inwardly curved long tips. Antennae with scape thick, angulate, pyramid-shaped, bearing several stiff, short setae; antennal clubs oval, with dense pubescence except for sclerotized areas in dorsal basal, outer lateral and outer ventral surfaces.
Pronotum (Fig. 3a) with posterior margin obtusely roundly angulate, sides almost straight, narrowing anteriorly, with shallow, narrow incisions in anterior thirds; pronotal disc convex, with sparse, shallow, indistinctly outlined punctures; lateral marginal stria complete, but inconspicuous; outer lateral striae present as sharp, low keels in basal halves aligned very close to lateral margins; anterior margin shallowly emarginate behind the head; antennal cavities not visible in dorsal view. Prosternum with prosternal lobe short, deeply concave medially, separated from rest of prosternum by stria; with complete fine marginal stria, small, but conspicuous lateral foveae and shallow lateral notches; prosternal keel broad, weakly convex, with two pairs of closely situated carinal striae along outer edges of the keel abbreviated around midpoint; lateral prosternal striae distinct, weakly diverging anteriorly; posterior margin triangularly emarginate, depressed medially (Fig. 2c).
Scutellum minute, triangular; elytra convex, widest at anterior third, weakly narrowing arcuately posteriorly; outer subhumeral striae present in anterior fourth, inner subhumeral striae complete, oblique subhumeral striae barely traceable as narrow shallow folds; all dorsal and sutural striae more or less complete, all elytral striae marked by a sparsely spaced, shallow, large, postobsolete punctures (see definition in [46]) (Fig. 2a).
Mesosternite with medial triangular projection on anterior margin; marginal stria fine, present only medially; disc of mesosternite with a few small punctures along anterior margin medially and transverse bisinuate keeled discal stria narrowly interrupted in the middle, running close to posterior margin; meso-metasternal sutures fine but conspicuous. metasternite with median suture distinct and complete, keeled in anterior half; disc of metasternite flat to weakly convex both in females and males, with widely scattered sparse, shallow, indistinctly outlined punctures; outer lateral striae complete, continuous with their long recurrent arms, inner lateral striae abbreviated halfway to metacoxae; longitudinal discal striae long, almost reaching metacoxae; anterior ends of outer lateral and longitudinal discal striae curved inwards; recurrent arms of inner lateral striae present, long, originating at metacoxae; recurrent arms and outer lateral striae situated upon sharp, low keels along the margins of the lateral depressions that house the mid- and hindlegs in repose . First visible abdominal ventrite unmodified, weakly convex, with row of punctures along the posterior margin, similar to those on metasternite, and paired long postmetacoxal striae laterally (Fig. 2c).
Pro-, meso- and metafemora elongate rectangular, with a row of sparse punctures in anterior third, coxae flattened. Protibiae elongate oval, widest at midlength, rugose, outer margins with few very weak, shallow emarginations, almost straight, with 7–8 small spines along outer margins, apices rounded, inner margins with row of setae; meso- and metatibiae paddle-shaped, outer margins obtusely angulate at anterior third, inner margins with row of setae. Tarsi laterally flattened, tarsomeres each with pair of setae on the ventral side; tarsal claws relatively short, almost straight (Fig. 2a-c).
Propygidium hexagonal, with traces of well-spaced, poorly outlined, shallow punctures; marginal stria indistinct. Pygidium convex, relatively short, trapezoidal; punctation similar to that on propygidium; no marginal stria present; disc somewhat more convex in males than in females. Male and female genitalia as illustrated (Fig. 2e-n).
Etymology. We dedicate this species to Daniel Kronauer, an avid field biologist and long-time army ant researcher, who discovered the species during an Eciton mexicanum s. str. colony emigration.
Distribution. Known only from the type locality, i.e. La Selva Biological Station, a lowland Atlantic rainforest in Costa Rica.
Molecular analysis - Nymphister kronaueri sp. nov.
COI barcodes were successfully obtained for 44 N. kronaueri specimens (including the holotype and paratypes; for GenBank accession numbers see Additional file 1: Table S1). All sequences were high-quality, full-length reads of 658 bp. No genetic variability was found in COI sequences, i.e. all specimens had the same COI haplotype.
Host specificity, abundance and phoretic transport of Nymphister kronaueri sp. nov.
In total, we collected 145 N. kronaueri specimens (including the holotype and paratypes). All specimens were collected from Eciton mexicanum s. str. colonies and no specimens were found in colonies of any other Eciton species at LSBS. Specimens were collected from 9 out of 10 Eciton mexicanum s. str. colonies during emigrations. Nymphister kronaueri showed a relatively high abundance compared to other haeteriines, which are generally rare ([36, 57, 58] and personal observations, CvB and AKT). We collected up to 60 N. kronaueri specimens from a single host colony (mean ± SD abundance in colonies with N. kronaueri = 16 ± 19; median = 12; N = 9 colonies).
Nymphister kronaueri specimens were attached between the petiole and postpetiole of host ants (Fig. 1). They were primarily found on ant workers of medium size (Fig. 4). On three occasions we found a specimen attached to an E. mexicanum s. str. major worker and once we detected an individual attached to a minor worker. In contrast to a variety of other Eciton-associated myrmecophiles (Tab. 1), we did not find N. kronaueri specimens walking in the emigration column independently. In almost all cases (˃95%), specimens were found toward the end of E. mexicanum s. str. colony emigrations when ant traffic was light and no ant brood was being carried anymore.

Phoretic transport of N. kronaueri on workers of a certain size. Size range of Eciton mexicanum s. str. head widths (N = 93) support the findings of Powell & Franks [77], i.e. a bimodal distribution consisting of the typical E. mexicanum workers scaling over a broad head width range, and major workers with a larger and distinct range in head widths. b Mean (dot), standard deviation (solid line), and range (dotted line) of Eciton mexicanum s. str. workers with N. kronaueri attached. Specimens with N. kronaueri attached were collected from two different ant colonies (N1 = 12 workers with N. kronaueri; N2 = 60 workers with N. kronaueri). Image credits: Daniel Kronauer
Discussion
Among the diverse menagerie of army ant-associated arthropods are numerous species awaiting scientific discovery. Here we have described one of them, the histerid beetle Nymphister kronaueri with an unusual mechanism of phoretic transport (Table 1). The beetles specifically grip the trunk between the ants’ two petiolar nodes (petiole and postpetiole) with their long mandibles (Fig. 1c, d). Possibly, the unique fronto-clypeal structure of this species helps to ‘anchor’ the beetle to the ant tightly (Fig. 1d). The beetles attach preferentially to workers of medium body size. This way they participated as hitchhikers in the frequent colony emigrations of their single host species Eciton mexicanum s. str.
Various myrmecophiles hitchhike in army ant emigrations by either attaching at different ant body parts or by attaching on brood or booty that is being carried by the ants (Table 1) [24, 58, 59]. Similar to N. kronaueri, some myrmecophiles are very specific in the location at which they attach to ants. Mites, in particular, are transportation specialists partitioning the ant bodies into ˈphoretic microhabitatsˈ [59]. For instance, a mite species of the genus Circocylliba Sellnick, 1926 [60] attaches specifically at soldier mandibles while another species, Macrocheles rettenmeyeri Krantz, 1962 [61], is only found at the pulvili of Eciton dulcium legs [59]. We are aware of only one other myrmecophile that attaches at the same position as N. kronaueri, the histerid Latronister rugosus Reichensperger, 1932 [58, 62]. It also attaches between the two petiolar segments of its host workers E. hamatum and E. vagans, but it rides in a ventral position. Interestingly, like N. kronaueri, it rides specifically on medium-sized workers but not on majors or small workers [58]. Irrespective of the exact mechanism, phoretic transport can be interpreted as a more advanced, energy-saving host-following mechanism compared to myrmecophiles following the colony emigrations on foot [3, 15].
To a human eye, attached N. kronaueri beetles are difficult to detect during army ant emigrations as they somewhat resemble the gaster of host ants in size and shape (Fig. 1). If this resemblance fools potential predators and as a consequence increases the beetles’ fitness, it constitutes a case of adaptive resemblance. We cannot exclude this possibility, but it seems unlikely to us. Nymphister kronaueri was exclusively collected during nocturnal ant emigrations and we did not detect it during daily ant raids. Hence, it seems unlikely that nocturnal predators, which barely rely on visual cues, have caused selection pressures on N. kronaueri to resemble a host gaster. In this context, it is also worth noting that other histerids with a generally similar body shape do not attach at this particular position (see for example Ecclisister bickhardti costaericae Reichensperger, 1935 in Fig. 3). Nonetheless, imitating a part of an ant’s body, here a gaster, might hamper the recognition by ants themselves, facilitating social integration of myrmecophiles into ant societies (e.g., [7, 27, 62]; see also discussion about tactile mimicry below).
Besides its unique mechanism of phoresy, Nymphister kronaueri’s external morphology, particularly its cuticular microsculpture and high density of macrosetae, constitutes an exception within the genus Nymphister (Table 2; see for comparison the smooth cuticular surface and lack of macrosetae in N. simplicissimus, Fig. 3). These morphological characters are also found in Ecclisister beetles, another group of Eciton-associated histerids of the tribe Nymphistrini that shows a similar host specificity on Eciton burchellii, and a similar caste specific phoresy on the underside of major worker heads (Table 1). The close morphological similarity of N. kronaueri and Ecclsister beetles led us to speculate about the possibility of convergent evolution deriving from occupying the same selective environment (for a discussion about possible selection pressures acting on myrmecophilous beetles see [8]). Frequent and close contact with army ant host workers, which principally represent potential predators for symbionts, might be responsible for the evolution of a high setal density on the cuticle of these beetle inquilines. This is because one of the primary functions of setae is mechano-reception [63, 64], possibly providing the necessary mechano-sensory input for nest-inhabiting myrmecophiles to avoid being caught by host ants. Frequent host contact might also be related to the second characteristic shared among these beetles, the integumental microsculpture. In our opinion, the microsculpture of N. kronaueri and Ecclisister is fairly similar to that of their host ants, a phenomenon that has been described for numerous myrmecophiles [1, 3, 7], including histerids [35]. Reichensperger noticed that histerid species with a rather loose, facultative association with army ants rather resemble the more typical histerid microsculpture of a smooth and shiny cuticle, whereas obligate myrmecophiles generally show a high degree of microsculpture similarity to host ants [35]. Upon tactile inspection by ant workers, a cuticular microsculpture resembling those of host ants might help to fool the host into accepting the inimical guests as nestmates (tactile mimicry; [1, 7, 65]).
Finally, the analysis of DNA barcodes in N. kronaueri deserves discussion. Sharing the same COI haplotype among species is rare [66], which is the reason why DNA barcoding is a useful tool to screen for species boundaries and to detect cryptic diversity [65–68]. For instance, we discovered morphologically cryptic species in Eciton-associated Vatesus beetles with the help of DNA barcodes (Staphylinidae: Tachyporinae) [30]. In contrast to Vatesus, we did not detect distinct COI clades, i.e. evidence for cryptic diversity, in N. kronaueri. Instead of genetic divergence, we only found a single COI haplotype for 44 analyzed specimens in the population at LSBS, an interesting and exceptional case of missing genetic variability among Eciton myrmecophiles at LSBS ([30, 48], and unpublished data, CvB). A complete lack of variance in COI is unusual in native arthropod populations and the possible causes are diverse [67–74]. A possible explanation is a recent genetic bottleneck, maybe due to a recent dispersal event followed by the expansion of a small genetically uniform founder population, a population genetic pattern typical for invasive species [71–75]. More elaborate population genetic markers and the inclusion of different geographic areas will be required to explore reasons for this lack of variability in N. kronaueri.
Conclusions
Despite the enduring efforts to explore the microcosm of army ant-associated arthropods, a large proportion of unknown biodiversity still exists today. Myrmecophile communities of army ant species other than E. burchellii and E. hamatum have not been intensively studied, and thus it can be expected that many more species with fascinating adaptations still await scientific discovery. The present study is an example demonstrating the benefits of a combined approach, using morphology and DNA barcodes, to discover and describe new species in ant-myrmecophile communities.
References
Hölldobler B, Wilson EO. The ants. Cambridge: Harvard University Press; 1990.
Wilson EO. Success and dominance in ecosystems: the case of the social insects. Oldendorf/Luhe: Ecology Institute; 1990.
Gotwald WH. Army ants: the biology of social predation. Ithaca: Comstock Pub. Associates; 1995.
Stadler B, Dixon AFG. Ecology and evolution of aphid-ant interactions. Annu Rev Ecol Evol Syst. 2005;36:345–72.
Parker J, Grimaldi DA. Specialized myrmecophily at the ecological dawn of modern ants. Curr Biol. 2014;24:2428–34.
Thomas JA, Schönrogge K, Elmes GW. Specializations and host associations of social parasites of ants. In: Fellowes MDE, Holloway GJ, Rolff J, editors. Insect evolutionary ecology. Reading, UK: CABI; 2005.
Kistner DH. Social and evolutionary significance of social insect symbionts. In: Hermann HR, editor. Social insects. New York: Academic; 1979. p. 339–413.
Parker J. Myrmecophily in beetles (Coleoptera): evolutionary patterns and biological mechanisms. Myrmecol News. 2016;22:65–108.
Geiselhardt SF, Peschke K, Nagel P. A review of myrmecophily in ant nest beetles (Coleoptera: Carabidae: Paussinae): linking early observations with recent findings. Naturwissenschaften. 2007;94:871–94.
Philips TK, Mynhardt G. Description of Electrognostus intermedius, the first spider beetle from Dominican amber with implications on spider beetle phylogeny (Coleoptera Ptinidae). Entomapeiron (PS). 2011;4:37–51.
Engel MS, Chatzimanolis S. An oxyteline rove beetle in Dominican amber with possible African affinities (Coleoptera: Staphylinidae: Oxytelinae). Ann Carnegie Mus. 2009;77:425–9.
Wilson EO. The insect societies. Cambridge: Harvard University Press; 1971.
Eidmann H. Die Gäste und Gastverhältnisse der Blattschneiderameise Atta sexdens L. Zoomorphology. 1937;32:391–462.
Navarrete-Heredia JL. Beetles associated with Atta and Acromyrmex ants (Hymenoptera: Formicidae: Attini). Trans Am Entomol Soc. 2001;127:381–429.
Rettenmeyer CW, Rettenmeyer ME, Joseph J, Berghoff SM. The largest animal association centered on one species: the army ant Eciton burchellii and its more than 300 associates. Insect Soc. 2011;58:281–92.
Parmentier T, Dekoninck W, Wenseleers T. A highly diverse microcosm in a hostile world: a review on the associates of red wood ants (Formica rufa group). Insect Soc. 2014;61:229–37.
Pérez-Lachaud G, Lachaud J-P. Arboreal ant colonies as “Hot-points” of cryptic diversity for myrmecophiles: the weaver ant Camponotus sp. aff. textor and its interaction network with its associates. Plos One. 2014;9:e100155.
Lachaud J-P, Pérez-Lachaud G. Ectaheteromorph ants also host highly diverse parasitic communities: a review of parasitoids of the neotropical genus Ectatomma. Insect Soc. 2015;62:121–32.
Witek M, Barbero F, Markó B. Myrmica ants host highly diverse parasitic communities: from social parasites to microbes. Insect Soc. 2014;61:307–23.
Kaspari M, O’Donnell S. High rates of army ant raids in the Neotropics and implications for ant colony and community structure. Evol Ecol Res. 2003;5:933–9.
Schneirla TC. Army ants. A study in social organization. In: Topoff HR, editor. San Francisco: W. H. Freeman & Co.; 1971.
Kronauer DJC. Recent advances in army ant biology (Hymenoptera: Formicidae). Myrmecol News. 2009;12:51–65.
Powell S. How much do army ants eat? On the prey intake of a neotropical top-predator. Insectes Soc. 2011;58:317–24.
Witte V, Leingärtner A, Sabaß L, Hashim R, Foitzik S. Symbiont microcosm in an ant society and the diversity of interspecific interactions. Anim Behav. 2008;76:1477–86.
Rettenmeyer CW. Arthropods associated with neotropical army ants with a review of the behavior of these ants (Arthropoda; Formicidae: Dorylinae). Lawrence: University of Kansas; 1961.
von Beeren C, Schulz S, Hashim R, Witte V. Acquisition of chemical recognition cues facilitates integration into ant societies. BMC Ecol. 2011;11:30.
von Beeren C, Hashim R, Witte V. The social integration of a myrmecophilous spider does not depend exclusively on chemical mimicry. J Chem Ecol. 2012;38:262–71.
Kistner DH. The social insects’ bestiary. In: Hermann HR, editor. Social insects. New York: Academic; 1982. p. 1–244.
Akre RD, Torgerson RL. Behavior of Vatesus beetles associated with army ants (Coleoptera: Staphylinidae). Pan Pac Entomol. 1969;45:269–81.
von Beeren C, Maruyama M, Kronauer DJC. Cryptic diversity, high host specificity and reproductive synchronization in army ant-associated Vatesus beetles. Mol Ecol. 2016;25:990–1005.
Akre RD, Rettenmeyer CW. Trail-following by guests of army ants (Hymenoptera: Formicidae: Ecitonini). J Kans Entomol Soc. 1968;41:165–74.
von Beeren C, Maruyama M, Hashim R, Witte V. Differential host defense against multiple parasites in ants. Evol Ecol. 2011;25:259–76.
Ott R, von Beeren C, Hashim R, Witte V, Harvey MS. Sicariomorpha, a new myrmecophilous goblin spider genus (Araneae, Oonopidae) associated with Asian army ants. Am Mus Novit. 2015;3843:1–14.
Helava JVT, Howden HF, Ritchie AJ. A review of the new world genera of the myrmecophilous and termitophilous subfamily Hetaeriinae (Coleoptera: Histeridae). Sociobiology. 1985;10:127–386.
Reichensperger A. Neue südamerikanische Histeriden als Gäste von Wanderameisen und Termiten. II Teil. Rev Suisse Zool. 1924;31:117–52.
Helava JVT. A new genus and two species of ant-associated histerids from Panama (Coleoptera: Histeridae: Hetaeriinae). Coleopt Bull. 1989;43:121–6.
Reichensperger A. Ecitophilen aus Costa Rica (II), Brasilien und Peru (Staph. Hist. Clavig.). Revista de Entomologia (Rio de Janeiro). 1933;3:179–194.
Reichensberger A. Beiträge zur Kenntnis der Myrmekophylen- und Termitophilenfauna Brasiliens und Costa Ricas v. (Col. Hist., Staph.). Revista de Entomologia (Rio de Janeiro). 1938;9:74–97.
Tishechkin AK, Mercado-Cárdenas A. Description of three new species of Nymphistrini (Coleoptera: Histeridae: Haeteriinae) from Central America. Zootaxa. 2012;3500:36–48.
Watkins II JF. The army ants of Mexico (Hymenoptera: Formicidae: Ecitoninae). J Kansas Entomol Soc. 1982;55:197–247.
Watkins JF. The identification and distribution of New World army ants (Dorylinae: Formicidae). Waco: The Markham Press Fund of Baylor University Press; 1976.
Longino J. Ants of Costa Rica. [Internet]. http://ants.biology.utah.edu/~longino/AntsofCostaRica.html. 2010.
Lackner T. Revision of the attaphilous genus Phoxonotus (Coleoptera: Histeridae: Saprininae). Eur J Entomol. 2016;113:240–58.
Caterino MS, Tishechkin AK. A systematic revision of Operclipygus Marseul (Coleoptera, Histeridae, Exosternini). Zookeys. 2013;271:1–401.
Caterino MS, Tishechkin AK. A systematic revision of Baconia Lewis (Coleoptera, Histeridae, Exosternini). Zookeys. 2013;343:1–297.
Tishechkin AK. Phylogenetic revision of the genus Mesynodites (Coleoptera: Histeridae: Hetaeriinae) with description of new tribes, genera and species. Sociobiology. 2007;49:1–167.
Tishechkin AK, Caterino MS. A new north American genus of Hetaeriinae (Coleoptera: Histeridae), with descriptions of six new species from the USA and Mexico. Zootaxa. 2009;2311:1–18.
von Beeren C, Maruyama M, Kronauer DJC. Community sampling and integrative taxonomy reveal new species and host specificity in the army ant-associated beetle genus Tetradonia (Coleoptera, Staphylinidae, Aleocharinae). Plos One. 2016;11:e0165056.
Waugh J. DNA barcoding in animal species: progress, potential and pitfalls. Bioessays. 2007;29:188–97.
Greenstone MH, Rowley DL, Heimbach U, Lundgren JG, Pfannenstiel RS, Rehner SA. Barcoding generalist predators by polymerase chain reaction: carabids and spiders. Mol Ecol. 2005;14:3247–66.
Kress WJ, Erickson DL. DNA barcodes: Methods and protocols. New York, USA: Springer Science+Busines Media; 2012.
Hebert PDN, Ratnasingham S, de Waard JR. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc B. 2003;270:S96–9.
Caterino MS, Tishechkin AK. DNA identification and morphological description of the first confirmed larvae of hetaeriinae (Coleoptera: Histeridae). Syst Entomol. 2006;31:405–18.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Marine Biol Biotechnol. 1994;3:294–9.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28:1647–9.
Parker M, Stones-Havas S, Starger C, Meyer C. Laboratory information management systems for DNA barcoding. Methods Mol Biol. 2012;858:269–310.
Akre RD. Behavior of Euxenister and Pulvinister histerid beetles associated with army ants. Pan pac entomol. 1968;44:87–101.
Rettenmeyer CW. The diversity of arthropods found with Neotropical army ants and observations on the behavior of representative species. Proc NC Branch-ESA. 1962;17:14–15.
Gotwald Jr WH. Mites that live with army ants: a natural history of some myrmecophilous hitch-hikers, browsers, and parasites. J Kansas Entomol Soc. 1996;69:232–237.
Sellnick M. Alguns novos acaros (Uropodidae) myrmecophilos e termitophilos. Arq Mus Nac Rio De J. 1926;26:29–56.
Krantz GW. Descriptions of three myrmecophilous Macrochelidae (Acarina: Mesostigmata) from Panama, British Guiana and the British West Indies. J Kansas Entomol Soc. 1962;35:349–57.
Reichensperger A. Ecitophilen und Termitophilen Aus Costa Rica I, nebst Bemerkungen über Fühlerbildung bei Mimeciton (Col. Hist. Staph.). Rev Entomol Rio Jan. 1932;2:6–14.
Winterton SL. Scales and setae. In: 1, Resh VH, Cardé RT, editors. Encyclopedia of insects. Amsterdam : Academic Press; 2003. p. 1017–20.
Katta S, Krieg M, Goodman MB. Feeling force: physical and physiological principles enabling sensory mechanotransduction. Annu Rev Cell Dev Biol. 2015;31:347–71.
Wasmann E. Die Ameisenmimikry: ein exakter Beitrag zum Mimikryproblem und zur Theorie der Anpassung. Berlin: Borntraeger; 1925. p. 944–51.
Hebert PDN, Gregory TR. The promise of DNA barcoding for taxonomy. Syst Biol. 2005;54:852–9.
Roman JOE, Palumbi SR. A global invader at home: population structure of the green crab, Carcinus maenas, in Europe. Mol Ecol. 2004;13:2891–8.
Darling JA, Bagley MJ, Roman JOE, Tepolt CK, Geller JB. Genetic patterns across multiple introductions of the globally invasive crab genus Carcinus. Mol Ecol. 2008;17:4992–5007.
Cognato AI, Sun J-H, Anducho-Reyes MA, Owen DR. Genetic variation and origin of red turpentine beetle (Dendroctonus valens LeConte) introduced to the People’s Republic of China. Agric for Entomol. 2005;7:87–94.
Mayer F, Piel FB, Cassel-Lundhagen A, Kirichenko N, Grumiau L, Økland B, et al. Comparative multilocus phylogeography of two palaearctic spruce bark beetles: influence of contrasting ecological strategies on genetic variation. Mol Ecol. 2015;24:1292–310.
Rubinoff D, Holland BS, Shibata A, Messing RH, Wright MG. Rapid invasion despite lack of genetic variation in the erythrina gall wasp (Quadrastichus erythrinae Kim). Pac Sci. 2010;64:23–31.
Simonsen TJ, Brown RL, Sperling FA. Tracing an invasion: phylogeography of Cactoblastis cactorum (Lepidoptera: Pyralidae) in the United States based on mitochondrial DNA. Ann Entomol Soc Am. 2008;101:899–905.
Scheffer SJ, Grissell EE. Tracing the geographical origin of Megastigmus transvaalensis (Hymenoptera: Torymidae): an African wasp feeding on a south American plant in north America. Mol Ecol. 2003;12:415–21.
Slade RW, Moritz C. Phylogeography of Bufo marinus from its natural and introduced ranges. Proc R Soc B. 1998;265:769–77.
Simberloff D. Invasive species: what everyone needs to know. New York: Oxford University Press; 2013.
Hendrich L, Morinière J, Haszprunar G, Hebert PD, Hausmann A, Köhler F, et al. A comprehensive DNA barcode database for central European beetles with a focus on Germany: adding more than 3500 identified species to BOLD. Mol Ecol Resour. 2015;15:795–818.
Powell S, Franks NR. Ecology and the evolution of worker morphological diversity: a comparative analysis with Eciton army ants. Funct Ecol. 2006;20:1105–14.
Tishechkin AK. New distribution records of neotropical Hetaeriinae (Coleoptera: Histeridae). Sociobiology. 2003;41:673–84.
Reichensperger A. Neue Beiträge zur Artenkenntnis und zur Lebensweise myrmekophiler Histeriden. Verh d III int Kongr Ent (Zürich). 1926;2:184–203.
Mann WM. Guests of Eciton hamatum (Fab.) collected by professor WM Wheeler. Psyche. 1925;32:166–77.
Bruch C. Nuevos histéridos ecotófilos (Col.). Rev Mus la Plata. 1926;29:17–33.
Reichensperger A. Neue südamerikanische Histeriden als Gäste von Wanderameisen und Termiten. I. Systematischer Teil Mitt Schweiz Entomol Ges. 1923;13:313–36.
Reichensperger A. Beitrag zur Kenntnis der Myrmekophilenfauna Brasiliens und Costa Ricas III. (col. staphyl. hist.). Arb Morph Taxon Ent Berlin-Dahlem. 1935;2:188–218.
Dégallier N. Coleoptera Histeridae de Guyane Française. IV. Myrmécophiles et termitophiles de la sous-famille des Hetaeriinae: notes techniques, faunistiques et taxonomiques. Bull Soc Entomol Fr. 2004;109:293–316.
Dégallier N. Coleoptera Histeridae Hetaeriinae: description de nouveaux taxons, désignation de lectotypes et notes taxonomiques. Bonn Zool Beitr. 1998;47:345–79.
Bruch C. Estudios mirmecológicos con la descripción de neuvas especies de dipteros (“Phoridae”) por los RR. PP. H. Schmitz y Th. Borgmeier y de une araña (“Gonyleptidae”) por El. Doctor Mello-leitão. Rev Mus la Plata. 1923;28:172–209.
Reichensperger A. Neue südamerikanische Histeriden als Gäste von Wanderameisen und Termiten. Zeitschr Ins Biol. 1923;18:243–52.
Reichensperger A. Die Wirte der Mesynodites-gruppe nebst Beschreibung neuer ecitophiler und termitophiler Histeridenarten (Coleopt. Hist.). Zool Jahrb. 1931;61:263–84.
Reichensperger A. Beiträge zur Kenntnis der Myrmecophilen-und Termitophilenfauna Brasiliens und Costa Ricas. VI. (Col. Hist. Staph.). Rev Entomol Rio Jan. 1939;10:97–137.
Reichensperger A. Systematische und ökologische Myrmekophilen-Beiträge (Staph., Hist., Pauss.). Zool Anz. 1929;82:257–68.
Acknowledgements
We thank two BMC editors, Laurence Packer and Terry Ord, as well as four anonymous reviewers for their helpful comments on earlier drafts of the manuscript. We thank Daniel Kronauer, Sebastian Pohl and Griffin Burke for help during field work. Special thanks go to Daniel Kronauer for hosting CvB as a postdoc in his laboratory and for providing comments on the manuscript. Thanks also go to Nico Blüthgen for providing office space for CvB, and Adrian Pinto, Carlos de la Rosa, Bernal Matarrita Carranza, Ronald Vargas and Danilo Brenes Madrigal, as well as the entire staff of La Selva Biological Station for their generous support.
Funding
A Carl & Marian Rettenmeyer Ant-Guest Endowment Award supported this work. In addition, CvB was supported by the German Science Foundation (DFG), the National Geographic Society’s Committee for Research and Exploration, and a Bristol-Myers Squibb Postdoctoral Fellowship from The Rockefeller University.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and its additional files, and are deposited at GenBank and BOLD (see Additional file 1: Table S1 for accession numbers).
Authors’ contributions
CvB designed the study; CvB and AKT performed field-work; CvB led the writing of the manuscript; both authors worked on the manuscript draft; CvB and AKT processed collections; CvB performed the molecular work; AKT provided morphological descriptions; both authors gave final approval for publication.
Competing interests
The authors declare that they have no competing interests.
Ethics approval and consent to participate
The present study complies with institutional, national, and international ethical guidelines. Research and specimen export permits for Costa Rica were issued by the ‘Ministry of the Environment, Energy and Technology’ (MINAET; permit numbers: 192-2012-SINAC and R-009-2014-OT-CONAGEBIO).
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1: Table S1.
Specimen Information. (XLS 90 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
von Beeren, C., Tishechkin, A.K. Nymphister kronaueri von Beeren & Tishechkin sp. nov., an army ant-associated beetle species (Coleoptera: Histeridae: Haeteriinae) with an exceptional mechanism of phoresy. BMC Zool 2, 3 (2017). https://doi.org/10.1186/s40850-016-0010-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40850-016-0010-x