
Abstract
Background
Previous studies reported differences in genotype frequency of the ACTN3 R577X polymorphisms (rs1815739; RR, RX and XX) in athletes and non-athletic populations. This systematic review with meta-analysis assessed ACTN3 R577X genotype frequencies in power versus endurance athletes and non-athletes.
Methods
Five electronic databases (PubMed, Web of Science, Scopus, Science Direct, SPORTDiscus) were searched for research articles published until December 31st, 2022. Studies were included if they reported the frequency of the ACTN3 R577X genotypes in power athletes (e.g., weightlifters) and if they included a comparison with endurance athletes (e.g., long-distance runners) or non-athletic controls. A meta-analysis was then performed using either fixed or random-effects models. Pooled odds ratios (OR) were determined. Heterogeneity was detected using I2 and Cochran's Q tests. Publication bias and sensitivity analysis tests were computed.
Results
After screening 476 initial registrations, 25 studies were included in the final analysis (13 different countries; 14,541 participants). In power athletes, the RX genotype was predominant over the two other genotypes: RR versus RX (OR 0.70; 95% CI 0.57–0.85, p = 0.0005), RR versus XX (OR 4.26; 95% CI 3.19–5.69, p < 0.00001), RX versus XX (OR 6.58; 95% CI 5.66–7.67, p < 0.00001). The R allele was higher than the X allele (OR 2.87; 95% CI 2.35–3.50, p < 0.00001) in power athletes. Additionally, the frequency of the RR genotype was higher in power athletes than in non-athletes (OR 1.48; 95% CI 1.25–1.75, p < 0.00001). The RX genotype was similar in both groups (OR 0.84; 95% CI 0.71–1.00, p = 0.06). The XX genotype was lower in power athletes than in controls (OR 0.73; 95% CI 0.64–0.84, p < 0.00001). Furthermore, the R allele frequency was higher in power athletes than in controls (OR 1.28; 95% CI 1.19–1.38, p < 0.00001). Conversely, a higher frequency of X allele was observed in the control group compared to power athletes (OR 0.78; 95% CI 0.73–0.84, p < 0.00001). On the other hand, the frequency of the RR genotype was higher in power athletes than in endurance athletes (OR 1.27; 95% CI 1.09–1.49, p = 0.003). The frequency of the RX genotype was similar in both groups (OR 1.07; 95% CI 0.93–1.24, p = 0.36). In contrast, the frequency of the XX genotype was lower in power athletes than in endurance athletes (OR 0.63; 95% CI 0.52–0.76, p < 0.00001). In addition, the R allele was higher in power athletes than in endurance athletes (OR 1.32; 95% CI 1.11–1.57, p = 0.002). However, the X allele was higher in endurance athletes compared to power athletes (OR 0.76; 95% CI 0.64–0.90, p = 0.002). Finally, the genotypic and allelic frequency of ACTN3 genes were similar in male and female power athletes.
Conclusions
The pattern of the frequencies of the ACTN3 R577X genotypes in power athletes was RX > RR > XX. However, the RR genotype and R allele were overrepresented in power athletes compared to non-athletes and endurance athletes. These data suggest that the RR genotype and R allele, which is associated with a normal expression of α-actinin-3 in fast-twitch muscle fibers, may offer some benefit in improving performance development in muscle strength and power.
Key Points
-
The ACTN3 gene encodes α-actinin-3, a structural protein of Z-lines of skeletal muscle fast-twitch (type II) actin filaments 4,5. Individuals with a ‘null’ XX in the p.R577X polymorphism of this gene are unable to express α-actinin-3, as opposed to individuals with the RR and RX genotypes.
-
This systematic review encompasses the meta-analysis of 23 studies that assessed ACTN3 R577X genotype frequencies in power athletes compared to endurance athletes and non-athletes. The meta-analysis reflected a higher prevalence of RR genotype and R allele in power athletes compared to non-athletes and endurance athletes.
-
These outcomes suggest that the RR genotype and the R allele, associated with the full expression of α-actinin-3 in type II muscle fibres may confer advantages for becoming an elite athlete in sports with high demands of muscle strength and power.
Similar content being viewed by others
Introduction
More than 200 genes and polymorphisms can alter the physical and physiological abilities of athletes [1], and it is estimated that genetic factors account for 40–60% of variations in cardiorespiratory parameters, 50–90% of the variations in anaerobic performance, and 30–70% of variations in muscle strength [2]. Among the many genes that may be related to athletic performance, the R577X polymorphism of the ACTN3 gene has been suggested to significantly influence sport performance [3].
The ACTN3 gene encodes α-actinin-3, a structural protein of Z-lines of skeletal muscle fast-twitch (type II) actin filaments [4, 5]. The R577X polymorphism is a single nucleotide polymorphism (SNPs) in the ACTN3 gene that affects the expression of α-actinin-3 [6]. Specifically, individuals with a ‘null’ 577XX (or simply ‘XX’) genotype are unable to express α-actinin-3, as opposed to individuals with the RR and RX genotypes [7]. The XX genotype does not cause harm, as the lack of α-actinin-3 expression is compensated by the overexpression of α-actinin-2 [7]. In fact, it is estimated that ~ 20% of the world population possess the XX genotype [8]. There is evidence that muscle phenotype may be partly determined by the ACTN3 R577X polymorphism [9].
The physical fitness qualities speed, power, and muscle strength are supported by the anaerobic metabolism [10]. Several studies investigated the impact of ACTN3 R577X polymorphisms on anaerobic performance in amateur and professional athletes [11,12,13], with the general finding of an overrepresentation of the RR genotype (and underrepresentation of the XX genotype) in athletes participating in anaerobic-based disciplines [14]. Specifically, ACTN3 RR/RX genotypes were associated with better speed, power and muscle strength performances, while the XX genotype was associated with increased endurance [12, 15,16,17,18]. Additionally, possessing the ACTN3 XX genotype has been associated with various phenotypes related to lower anaerobic performance and an increased risk of muscle damage associated with exercise [19,20,21]. Conversely, although a higher frequency of the ACTN3 XX genotype was found in endurance athletes in the first study on this topic [21], current evidence suggest no association between the XX genotype of ACTN3 gene and a higher level of endurance performance [22, 23]. A recent meta-analysis by Tharabenjasin et al. [24] reported a correlation between the R allele and RX genotype of the ACTN3 polymorphism in elite power athletes. However, the aforementioned meta-analysis did not compare power with endurance athletes in terms of genotypes and alleles of ACTN3 R577X polymorphism [24]. Furthermore, a significant dominance of the R allele over the X allele was observed (odds ratio [OR] = 1.35, 95% CI 1.18–1.53) in professional soccer players. These findings may suggest an association of the R allele of the ACTN3 gene and better performance in soccer [25].
Studies to date suggest ACTN3 R577X polymorphisms are strongly associated with anaerobic performance. However, the available studies have not yet been systematically aggregated in the form of a meta-analysis. Here, we aimed to systematically review and meta-analyze studies on ACTN3 R577X genotype frequencies in power athletes (disciplines of short duration, high intensity and dominated by muscle power) and compared data with endurance athletes (activities of long duration, low to moderate intensity, dominated by endurance and dependent on oxygen availability) and non-athletes. We hypothesized that the RR genotype and R allele would be overrepresented in power athletes compared to non-athletes and endurance athletes [12, 16, 18, 24, 25].
Methods
The protocol for our review was registered in PROSPERO (registration number: CRD42022360255). The applied systematic searches and meta-analyses were conducted in accordance with the recommendations outlined in the Cochrane Handbook for Systematic Review and Meta-analysis of Interventions [26]. A bibliographic search strategy was performed using the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) Statement [27].
Working Definitions
Aerobic-based sports (endurance athletes) generally last longer than 5 min and involve an exercise intensity similar to or lower than VO2max. Aerobic-based sports are related to the ability of the respiratory and circulatory systems to provide energy and replenish adenosine triphosphate (ATP) through oxidative metabolism [28, 29]. For the purpose of this meta-analysis, studies were included examining road cyclists, marathoners, and long-distance triathletes. In contrast, anaerobic-based sports (power athletes) require an intensity higher than VO2max and energy provision for muscle contraction depend on different metabolic pathways related to the duration of the exercise, including anaerobic glycolysis and the ATP/phosphocreatine system [30]. In the context of this meta-analysis, studies were included with track sprinters, long jumpers, weightlifters.
Elite athletes refer to well-trained athletes who train at least four times per week and compete on an international level.
Regarding the sub-elite athlete category, we refer to athletes who participate in national or state leagues/tournaments and develop the skills necessary to play the sport at a high (national) level, including biomechanics, ball handling skills, and decision-making elements.
Non-athletes and healthy individuals engage in multiple sports and/or types of physical activity in accordance with the World Health Organization (WHO), with at least 150 to 300 min of moderate-intensity activity or 75 to 150 min of vigorous-intensity activity per week and muscle-strengthening activities 2 or more days per week.
Eligibility Criteria
Only studies that examined the link between the ACTN3 R577X polymorphism and anaerobic-based exercise performance (e.g., muscle strength, power, speed) were included in our analysis. For this purpose, we selected studies assessing the frequency of the ACTN3 R577X genotypes in athletes of anaerobic-based sports in which a comparison with either endurance athletes or non-athletic controls was made. We used the following criteria for studies that were included in our systematic review: (1) published in peer-reviewed journals; (2) participants aged 14 years or older; (3) involved athletes of elite or sub-elite level; (4) used validated methods for the characterization of athletes related to anaerobic-based performances (e.g., strength-, power-, or speed-based sports) and contrasted those with endurance athletes (e.g., long-distance runners) or non-athletes; (5) presented data on the frequency of the ACTN3 R577X polymorphisms (such as RR, RX and XX). Studies were excluded if they (1) did not meet the minimum requirements of an experimental study design (e.g., case reports), (2) did not meet the minimum requirements to classify the sample as athletic; (3) were not written in English; or (4) presented data on the frequency of the ACTN3 R577X polymorphism only as RR vs X-allele carriers or R-carriers vs XX. Moreover, systematic or narrative review articles were not included in the current systematic review and meta-analysis.
Literature Search Strategy
Systematic literature searches were conducted in five electronic databases (PubMed, Web of Science, Scopus, Science Direct and SPORTDiscus) from inception until December 31st, 2022, with no restriction on publication dates. The following key terms (and synonyms searched using the MeSH database) were included and combined using the operators “AND”, “OR”, “NOT”: (Alpha-actinin-3 OR ACTN3) OR (“ACTN3 gene” OR “ACTN3 R577X polymorphism” OR “ACTN3 R577X” OR “ACTN3 R577R” OR “ACTN3 577XX genotype*” OR “ACTN3 577RR genotype*” OR “ACTN3 577RX genotype*”) AND (“anaerobic performance” OR “muscle strength*” OR “muscle power” OR speed OR jump) AND (correlation study OR association OR relationship). In addition, the reference lists and citations (Google Scholar) of the identified studies were explored to detect additional relevant studies.
Study Selection
The screening and study selection was realized by two investigators (EMEO and BB) using the inclusion and exclusion criteria introduced above. If the title of the article was of potential relevance, the abstract was examined and if still found eligible, the full text was reviewed for evaluation. A third-party consensus meeting was held with a third author (HZ) if the two investigators were unable to reach an agreement on the inclusion of the respective article.
Data Extraction
Once the inclusion/exclusion criteria were applied, data extraction was conducted to collect information about participants, interventions, comparisons, outcomes and study design (PICOS) in agreement with PRISMA methodology. The following relevant data from each study were extracted: study details (author, year of publication, country), study population (sample size, age, sex), the method used to determine the genotyping of ACTN3 R577X polymorphism (rs1815739), and the number of participants with RR, RX and XX genotypes in each group. For articles in which these information were not present, associations were calculated using raw data, if available. Two authors (EMEO and BB) extracted data from the included studies, and, a third (HZ) was consulted in case of uncertainty.
Quality Assessment
The methodological quality of the included studies was assessed using the Physiotherapy Evidence Database (PEDro) scale, which has good reliability and validity [31]. The PEDro scale has eleven possible items and examines external validity (criterion 1) and internal validity (criteria 2–9) of controlled trials and whether there is sufficient statistical information for interpreting results (criteria 10–11). A cut-off threshold of six points on the PEDro scale was used to indicate high-quality studies, as this has been reported to be sufficient to determine the methodological quality level in previous studies. Two independent researchers (EMEO and BB) assessed the quality of the studies, if any disagreements arose, a third researcher (HZ) was contacted, and a unanimous decision was achieved. Any study with a PEDro score below four points would have been excluded from the systematic review and meta-analysis, although none of the studies reviewed were excluded by this criterion.
Statistical Analyses
To compute associations between the ACTN3 R577X polymorphism in power athletes compared with endurance athletes and controls (non-athletes), ORs with 95% confidence intervals (CI) and forest plots were calculated using the number of participants with the RR, RX and XX genotype in each study. With these data, we identified the individual and pooled effects of the studies. The degree of heterogeneity between the results of the study was evaluated using the I2 test, where I2 values of 25%, 50% and 75% were considered as low, medium and high levels of heterogeneity, respectively. The application of fixed or random effects models for each analysis was based on the level of heterogeneity revealed via the I2 statistics (< 50% = fixed-effects model; > 50% = random-effects model) and Cochran’s Q [32] test, (significance level at p < 0.05). We followed the Cochrane recommendations [33] to detect publication bias in our meta-analysis, and a visual inspection of the funnel plot was performed. Data were analyzed using Cochrane Review Manager (RevMan) version 5.4.1. We performed a one-way ANOVA to calculate the mean percentage distribution and 95% CI for the RR, RX and XX genotypes. Statistical analyses were carried out with GraphPad Prism 9.2.0 (GraphPad Software Inc, San Diego, USA).
Results
Selection of Studies and Characteristics of the Included Studies
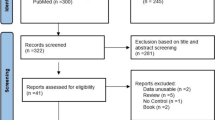
We identified 476 potentially relevant articles, of which 96 studies were duplicates, 63 were review articles and 198 articles were considered ineligible based on inclusion/exclusion criteria and the titles and abstracts (Fig. 1). After reviewing 119 articles, a further 94 were excluded based on the results of the full-text assessment for the following reasons: participants aged under 14 years (n = 32), no elite or sub-elite athletes (n = 33) were included in the study, missing data or uncertain data in one or more genotypes (n = 18) and high risk of bias (n = 11). Based on our inclusion/exclusion criteria and quality assessment, 25 eligible studies were included in our systematic review with meta-analysis [9, 10, 12, 23, 34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54]. These studies were conducted in 13 different countries including Czech Republic, Israel, Brazil, Russia, Spain, Australia, Lithuania, India, Japan, Korea, Italy, Poland and China, with a total sample size of 14,541 participants (7,080 athletes and 7,461 controls). Eleven studies included only males and 14 studies included both male and female participants. The general characteristics of the 25 included studies in this systematic review and meta-analysis were summarized in Table 1.

Flowchart of selecting eligible studies for systematic review and meta-analysis
Study Quality Assessment
The eligible studies had satisfactory methodological quality, as summarized in Table 2. We used the PEDro scale to identify one low quality study (4 points), 21 studies were of moderate quality (5 or 6 points) and three studies were of high quality (7 points or more).
Meta-Regression and Cumulative Meta-analysis
For power athletes, our search identified 19 eligible studies [9, 10, 12, 23, 34,35,36,37,38,39,40,41,42,43,44,45,46, 52, 53] with 2,212 athletes genotyped the ACTN3 R577X polymorphism. Another 15 studies [9, 10, 12, 23, 35,36,37,38,39,40,41,42, 44, 46, 46, 49] were identified with power athletes (n = 2310) and controls (n = 6203) genotyped the ACTN3 R577X polymorphism. Eight studies [9, 10, 12, 37,38,39, 46, 49] were included with power athletes (n = 1722) and endurance athletes (n = 1357) genotyped the ACTN3 R577X polymorphism In addition, five studies compared ACTN3 R577X genotypes in male (n = 564) and female (n = 296) power athletes [47, 48, 50, 51, 54].
These studies included fencers, track sprinters, long jumpers, weightlifters, football players, basketball players, speed skaters, sprint swimmers, ice hockey players, volleyball players, rugby players, sprint canoers, wrestlers, gymnasts, mixed samples of athletes (such as jumpers, sprinters and decathlon athletes) or samples with unspecified power-based sports. All of these sport disciplines were classified as muscle power dominated in the context of this meta-analyses. Additionally, the samples including long-distance runners, marathoners, road cyclists, endurance rowers, and long-distance triathletes, and the samples with unspecified endurance-based sports were categorized as endurance dominated for the purpose of this meta-analysis.
ACTN3 R577X Polymorphism in Power Athletes
The pooled data on studies of power athletes showed a lower frequency of the RR genotype compared to the RX genotype (OR 0.70; 95% CI 0.57–0.85, p = 0.0005) with a moderate heterogeneity of 59% (Fig. 2). The pooled data also indicated a higher frequency of RR in power athletes compared to XX counterparts (OR 4.26; 95% CI 3.19–5.69, p < 0.00001) with a moderate heterogeneity of 67% (Fig. 3). Additionally, the frequency of RX in power athletes was higher than XX counterparts (OR 6.58, 95% CI 5.66–7.67, p < 0.00001) with a moderate heterogeneity of 51% (Fig. 4). The pooled data showed a higher frequency of the R allele compared to the X allele (OR 2.87; 95% CI 2.35–3.50, p < 0.00001, 79% heterogeneity) (Fig. 5).




ACTN3 R577X Polymorphism in Power Athletes Compared to Controls
The pooled data comparing ACTN3 R577X polymorphism in power athletes versus controls (non-athletes) showed a greater frequency of the RR genotype in athletes than in controls (OR 1.48, 95% CI 1.25–1.75, p < 0.00001, moderate heterogeneity of 51%) (Fig. 6). The frequency of the RX genotype was similar in both groups (OR 0.84, 95% CI 0.71–1.00, p = 0.06, moderate heterogeneity of 61%) (Fig. 7). In contrast, the frequency of the XX genotype was lower in power athletes than in controls (OR 0.73, 95% CI 0.64–0.84, p < 0.00001, low heterogeneity of 30%) (Fig. 8). Furthermore, there was higher frequency of the R allele in power athletes compared to controls (OR 1.28; 95% CI 1.19–1.38; p < 0.00001) with 25% heterogeneity (Fig. 9). Conversely, a high frequency of the X allele was observed in the control group compared to power athletes (OR 0.78; 95% CI 0.73–0.84, p < 0.00001) with 25% heterogeneity (Fig. 10).





ACTN3 R577X Polymorphism in Power Athletes Compared to Endurance Athletes
When comparing the ACTN3 R577X polymorphism in power athletes versus endurance athletes, the pooled data indicated that the frequency of the RR genotype was higher in power athletes than in endurance athletes (OR 1.27, 95% CI 1.09–1.49, p = 0.003, low heterogeneity of 43%) (Fig. 11), while no significant differences were observed between groups of athletes for the frequency of the RX genotype (OR 1.07, 95% CI 0.93–1.24, p = 0.36, low heterogeneity of 15%) (Fig. 12). The frequency of the XX genotype in power athletes was lower than in endurance athletes (OR 0.63, 95% CI 0.52–0.76, p < 0.00001, low heterogeneity of 29%) (Fig. 13). In addition, a high frequency of the R allele occurred in power athletes compared to endurance athletes (OR 1.32; 95% CI 1.11–1.57, p = 0.002) with 53% of heterogeneity (Fig. 14). Conversely, the frequency of allele X was higher in endurance athletes than in power athletes (OR 0.76; 95% CI 0.64–0.90, p = 0.002) with a heterogeneity of 53% (Fig. 15).





ACTN3 R577X Polymorphism in Male and Female Power Athletes
The pooled data indicated that the frequency of the RR genotype was similar in males versus female power athletes (OR 0.82, 95% CI 0.61–1.10, p = 0.18, 0% of heterogeneity) (Fig. 16A), while the RX genotype was slightly more frequent in male than in female power athletes, although this was not statistically significant (OR1.32, 95% CI 0.99–1.77, p = 0.06, 0% heterogeneity) (Fig. 16B). The frequencies of the XX genotype were similar in males and females power athletes (OR 0.79, 95% CI 0.49–1.28, p = 0.34, 0% heterogeneity) (Fig. 16C). Moreover, no significant differences were observed in the frequencies of R and X alleles in male (OR 0.94, 95% CI of 0.76–1.17, p = 0.57, 1% of heterogeneity) (Fig. 17A) and female power athletes (OR 1.06, 95% CI 0.86–1.32, p = 0.57, 1% heterogeneity) (Fig. 17B).


Genotypic Frequency of ACTN3 R577X Polymorphism in all Groups
Subsequently, we performed further analyses by calculating the mean percentage distribution and determined the 95% CI for RR, RX, and XX genotypes in the various groups included in this meta-analysis. The main objective was to identify the dominant genotype in power and endurance athletes as well as in controls (non-athletes). The results of this analysis are presented in Table 3 and represented visually in Fig. 18.

Percentage distribution of RR, RX and XX genotypes in the different groups included in this meta-analysis. Power athletes (A) [9, 10, 12, 23, 34,35,36,37,38,39,40,41,42,43,44,45,46, 52, 53], controls (B) [9, 10, 12, 23, 34,35,36,37,38,39,40,41,42, 44, 46], endurance athletes (C) [9, 10, 12, 37,38,39, 46, 49], male power athletes (D) [47, 48, 50, 51, 54] and female power athletes (E) [47, 48, 50, 51, 54]
Sensitivity Analyses
We used both the fixed-effects model and a random-effects model based on the percentage of heterogeneity in our initial sensitivity analysis. As the use of fixed-effects or random-effects independently of heterogeneity reflects the same between-groups differences for the comparison of populations, we next conducted another sensitivity analysis by removing one study at a time to determine the impact of the deleted data on the ORs and heterogeneity. One study [49] was excluded from the meta-analysis due to excessive heterogeneity caused by a large sample size. The corresponding OR for the genotype comparisons did not change with this additional sensitivity analysis, indicating the robustness of our data.
Publication Bias
A visual inspection of the funnel plots for each meta-analysis indicated no publication bias for any of the meta-analyses included in this investigation. The funnel plots for the comparison of genotype frequencies are included in the Additional file 1.
Discussion
This systematic review with meta-analysis assessed ACTN3 R577X genotype frequencies in power athletes compared with populations of endurance athletes and non-athletes. Our meta-analysis showed that the frequency of ACTN3 R577X genotypes in power athletes followed an RX > RR > XX pattern, which is similar to that in non-athletes and endurance athletes. However, the RR genotype was over-represented in power athletes compared to non-athletes and endurance athletes. In contrast, the XX genotype was under-represented in power athletes compared to non-athletes and endurance athletes. Additionally, the RX genotype was slightly more frequent in male than in female power athletes, but this was not statistically significant. Collectively, these data demonstrate the frequency of the ACTN3 R577X genotype in elite and sub-elite strength athletes, which is characterized by a higher proportion of RR athletes and a lower prevalence of XX genotype. The genotype ACTN3 R577X affects the expression of α-actinin-3 in fast-twitch fibres [7], suggesting that a full expression of α-actinin-3 associated with the RR genotype may increase the likelihood of reaching the status of elite athletes in anaerobic sporting events.
ACTN3 R577X Polymorphism in Power Athletes Compared to Controls
Our meta-analysis indicates that the RR genotype and the R allele are more frequent in power athletes than in controls, which is supported by other reports [24, 35, 45, 48]. Thus, we provide additional evidence for the important role of the RR genotype in performance in speed/power sports. Studies of R and X alleles in athletes and controls and in athletes of different sport types (mainly power-based versus endurance-based) suggest that the R allele is more frequent in power athletes than in control (non-athletes) [23, 38] or endurance athletes [55]. Other studies also reported that the RR genotype is over-represented while the XX genotype is under-represented in strength and power athletes compared to control groups [21, 38, 56].
Our finding that the RR genotype and the R allele of the ACTN3 R577X polymorphism promote power and speed phenotypes [57] is supported by findings in young male Chinese soldiers where the RR genotype is associated with better anaerobic performance, while those with the XX genotype had lower grip strength [58]. Muscle strength and size are greater for RR and RX genotypes than for XX genotypes [59]. Additionally, other studies show that athletes with the ACTN3 RR genotype have better sprint times and a higher proportion of type II muscle fibers compared to those with the XX genotype [16, 60]. The percentage and number of IIx fibers are higher in RR-genotype athletes than in XX-genotype athletes, and the α-actinin-3 protein content is higher in type IIx fibers than in type IIa [61]. Finally, athletes with the RR genotype have higher levels of testosterone [16], which may explain, in part, the association between the ACTN3 RR genotype, skeletal muscle hypertrophy and power athlete status, as athletes carrying the RR genotype are more likely to be power athletes. These observations have led to the ACTN3 gene being labeled the “speed gene” [14]. Given that the RR genotype is overrepresented in power athletes, it is possible that this genotype augments the chance of being an elite athlete in sports requiring power, speed and strength [19]. The α-actinin-3 protein allows the traction of actin filaments attached to the Z line at high amplitudes of muscle contraction in type II fibers, raising the possibility that athletes with RR and RX genotypes (who express the α-actinin-3) perform better in strength and power activities than those with XX genotypes [36].
There are some findings that do not support the contention that ACTN3 R577X genotypes are associated with strength and power in weightlifting, powerlifting and throwing events [40]. For example, differences in the distribution of ACTN3 R577X alleles/genotypes were not observed in a group cohort of athletes or in each group in Russian and Lithuanian weightlifters and throwers [40]. Other studies suggest no benefits of the ACTN3 R577X polymorphism on muscle strength and other power-based phenotypes [62,63,64,65,66], as also shown when jump heights were measured in volleyball, basketball and rugby players [48, 67].
Collectively, these studies indicate that the ACTN3 R577X genotype contributes to the likelihood of becoming an elite athlete in aerobic sports but that possessing the XX genotype does not directly negate becoming an elite athlete in these sports disciplines. This is important as the genotyping of the ACTN3 R577X should not be used to detect talents in anaerobic-sports. The selection of RR athletes in the talent detection process should not occur at the expense of talented athletes with a XX genotype as there may be additional factors to also consider (physical and physiological abilities, psychological attributes, environment and motivation). For physical fitness qualities, the level of inheritance ranges from 20% for (balance) to 70–80% for alactic anaerobic power which is in line with our findings [68]. The practical application of a genotype score to is currently exploratory in nature, and its use in talent prediction is premature and has in fact proven to be ineffective [64,65,66,67,68], likely because the collection of genotypic data from the global population from athletes from different sporting disciplines.
ACTN3 R577X Polymorphism in Power Athletes Compared to Endurance Athletes
Our findings are in agreement with other studies in which the frequency of the RR genotype and the presence of the R allele was higher in athletes specializing in strength-power sports [21]. Additionally, the frequency of the RR genotype was higher in sprint/power, martial arts, and ball game sports athletes [46]. These findings suggest a crucial role for α-actinin-3, which is expressed only in individuals with the R-allele, in elite level sporting activities with a large anaerobic component. A study of top Finish sprinters demonstrated none of them possessed the XX genotype [69] Soccer players with the XX genotype showed higher VO2 peak levels than their RR genotype counterparts [45]. Conversely, individuals with the RR genotype showed reduced times in 10, 20 and 30m sprints, as well as better performance in jumping tests [45]. The XX genotype is underrepresented in Russian endurance athletes [70], which may indicate that in addition to endurance capacity, power and speed components are also necessary for success in endurance sports [71,72,73]. On the other hand, data from Asia and Africa suggests that a deficiency of ACTN3 is not related to endurance performance [74, 75]. Athletes with the RR genotype have higher explosive leg muscle power in jump tests compared to those with the XX genotype, but some studies report no difference between RX and XX genotypes [9].
In contrast, some studies reported that homozygous XX athletes had higher explosive power than athletes with a RR genotype [67], or that was no association between the ACTN3 gene and power/strength phenotypic traits [50]. The latter study had a limitation because the tests were not specific to the activity. ACTN3 knockout mice show improved endurance performance, characterized by high oxidative enzyme activity and a transition to slow-twitch type I muscle fibers [76, 77], which are highly vascularized and contain many mitochondria [78]. A deficiency of the XX genotype leads to a lack of α-actinin-3 protein, with reductions in athletic power and speed [19]. Additionally, the changes in type II muscle fibers due to the absence of ACTN3 reduces sprint performance and improves endurance performance in athletes with a XX genotype [21, 69]. Individuals with the XX genotype have superior physical characteristics compared to those with the R allele, due to a higher proportion of Type I fibers [77], a higher ventilation threshold respiratory compensation point [79], a higher fatigue index [80], decreased glycogen depletion in type II fibers after exercise [81], and a higher VO2peak [82]. Moreover, the proportion of subjects with aerobic energy metabolism for maximal and submaximal exercise was high for the athletes with a XX genotype [77, 79, 82].
ACTN3 R577X Polymorphism in Male and Female Power Athletes
Athletic performance can be influenced by sex-specific genetic factors such as differences in the physical, physiological and psychological abilities, including factors specific to males and females [83, 84]. Our meta-analysis shows that the RX genotype is slightly more frequent in male than in female power athletes (p = 0.06), while no differences were observed for the expression of RR and XX genotypes. In contrast to our findings, the distribution of RX was reported by others to be greater in female than in male sprinters (57% versus 39%) [85]. Yang et al. [12] found female sprint/power athletes lacked the XX genotype. Peak power was greater in male athletes with the R dominant allele compared to the XX group, but this difference was not present in female athletes [86]. Additionally, there is a higher frequency of the R allele in elite female sprint swimmers than in national level female swimmers and controls [87]. However, the ACTN3 XX genotype was similarly under-represented in both male and female artistic gymnasts compared to controls [54]. Female weightlifters have a higher ratio of the XX genotype (16% in females compared to 14% in males) [51]. On the other hand, no significant difference between sexes were observed in the frequency of genotypes between those with the best and worst performance in terms of power [66]. A study by Clarkson et al. [88] reported no correlation between ACTN3 R577X polymorphisms and isometric elbow flexion strength in males, although females with the XX genotype had lower isometric strength than those with a heterozygous RX. A study by Orysiak et al. [50] found no association between sports disciplines and ACTN3 genotype variants for female and male athletes, and the distribution of ACTN3 R577X polymorphisms did not differ between male and female athletes. Some studies have reported that the absence of the actinin-3 protein negatively influences the maximal isokinetic knee extension torque in middle-aged women, but not in males [89]. However, as genotypes can have different effects on athletic performance in female and male athletes, gene expression may be constructed differently for each sex [90].
Strengths and Limitations
This study expands on previous studies on the association between ACTN3 R577X polymorphisms and performance in sports of a short and intense nature. Our meta-analysis included 25 eligible studies conducted in 13 different countries with controls and athletes from different specialties. An assessment of the association between ACTN3 R577X polymorphisms and anaerobic exercise in various sports (power athletes, power athletes vs. controls, power athletes vs. endurance athletes, and male vs. female power athletes) indicates that the RR genotype and the R allele has greater benefits in power athletes, while the XX genotype and the X allele may be less favorable. Our meta-analyses were strengthened by a low heterogeneity in the studies, where seven forest plots indicated low heterogeneity (< 50%) and five forest plots demonstrated moderate heterogeneity (< 70%).
Our systematic review and meta-analysis has some limitations: (1) we identified only five studies that included both sexes with relatively small total sample sizes of males (n = 564) and females (n = 296), suggesting caution in discussing the role of sex on the distribution of ACTN3 R577X genotypes in power athletes; (2) although this study analyzed a large number of eligible studies (25 studies) with a large total sample size (14,541 participants), more studies are needed to support any recommendations, particularly related to the future use of genetic data in the orientation, optimization, adaptation of training and planning of the workload of athletes.
Conclusions
Our systematic review with meta-analysis provides additional evidence to support the notion that genetic variations contribute to athletic status and sports performance. Findings of this study suggest that the frequency of the ACTN3 R577X genotypes in power athletes followed a RX > RR > XX pattern, with a dominance of the R allele over the X allele. However, the RR genotype was overrepresented in power athletes compared non-athletes and endurance athletes, while the XX was under-represented in power athletes. Collectively, the study results suggest that the RR genotype, associated with a normal expression of α-actinin-3 in fast-twitch fibers, could be useful to elite performers of anaerobic-based exercise, and can increase athletic abilities in short and highly intense sports.
From a practical perspective, ACTN3 genotyping in athletes may allow coaches to better tailor training programs in terms of frequency and intensity to the athlete's genetic constitution and optimize performance and reduce the likelihood of injury. For example, an athlete with an RR genotype could benefit from strength and power training to improve vertical/horizontal jump performances and linear as well as change-of-direction speed, while an athlete with an XX genotype could benefit from endurance training to improve aerobic capacity. However, the type of training more suitable for each athlete according to their ACTN3 genotype is still under debate and further research is warranted in this topic. In any case, genotyping should not be used to select those athletes who could have a higher probability of becoming elite for either power or endurance sports, as this would introduce selection bias [91]. Regarding future studies, we propose a comprehensive exploration of the ACTN3 gene and various biomarkers in elite athletes from various sporting disciplines. This approach aims to elucidate the physiological implications of the ACTN3 R577X polymorphism and its potential influence on athletic performance.
Availability of Data and Materials
All data supporting the findings of this study are available in this published article.
References
Bray MS, Hagberg JM, Pérusse L, et al. The human gene map for performance and health-related fitness phenotypes: the 2006–2007 update. Med Sci Sports Exerc. 2009;41(1):35–73. https://doi.org/10.1249/mss.0b013e3181844179.
Bouchard C, Hoffman EP, editors. Genetic and molecular aspects of sport performance. 1st ed. London: Wiley; 2011. https://doi.org/10.1002/9781444327335.
Routledge Handbook of Sport and Exercise Systems Genetics. Routledge & CRC Press. Accessed 26 Mar 2023. https://www.routledge.com/Routledge-Handbook-of-Sport-and-Exercise-Systems-Genetics/Lightfoot-Hubal-Roth/p/book/9780367731311.
Koku FE, Karamizrak SO, Ciftci AS, Taslidere H, Durmaz B, Cogulu O. The relationship between ACTN3 R577X gene polymorphism and physical performance in amateur soccer players and sedentary individuals. Biol Sport. 2019;36(1):9–16. https://doi.org/10.5114/biolsport.2018.78900.
Mills M, Yang N, Weinberger R, et al. Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: implications for the evolution of functional redundancy. Hum Mol Genet. 2001;10(13):1335–46. https://doi.org/10.1093/hmg/10.13.1335.
North KN, Beggs AH. Deficiency of a skeletal muscle isoform of alpha-actinin (alpha-actinin-3) in merosin-positive congenital muscular dystrophy. Neuromuscul Disord. 1996;6(4):229–35. https://doi.org/10.1016/0960-8966(96)00361-6.
Del Coso J, Hiam D, Houweling P, Pérez LM, Eynon N, Lucía A. More than a “speed gene”: ACTN3 R577X genotype, trainability, muscle damage, and the risk for injuries. Eur J Appl Physiol. 2019;119(1):49–60. https://doi.org/10.1007/s00421-018-4010-0.
Domańska-Senderowska D, Szmigielska P, Snochowska A, et al. Relationships between the expression of the ACTN3 gene and explosive power of soccer players. J Hum Kinet. 2019;69:79–87. https://doi.org/10.2478/hukin-2019-0020.
Orysiak J, Busko K, Michalski R, et al. Relationship between ACTN3 R577X polymorphism and maximal power output in elite Polish athletes. Medicina (Kaunas). 2014;50(5):303–8. https://doi.org/10.1016/j.medici.2014.10.002.
Massidda M, Bachis V, Corrias L, et al. ACTN3 R577X polymorphism is not associated with team sport athletic status in Italians. Sports Med Open. 2015;1:6. https://doi.org/10.1186/s40798-015-0008-x.
Cięszczyk P, Eider J, Ostanek M, et al. Association of the ACTN3 R577X polymorphism in polish power-orientated athletes. J Hum Kinet. 2011;28:55–61. https://doi.org/10.2478/v10078-011-0022-0.
Yang R, Shen X, Wang Y, et al. ACTN3 R577X gene variant is associated with muscle-related phenotypes in elite Chinese sprint/power athletes. J Strength Cond Res. 2017;31(4):1107–15. https://doi.org/10.1519/JSC.0000000000001558.
Moreno-Pérez V, Machar R, Sanz-Rivas D, Del Coso J. ACTN3 R577X genotype in professional and amateur tennis players. J Strength Cond Res. 2020;34(4):952–6. https://doi.org/10.1519/JSC.0000000000003501.
Houweling PJ, Papadimitriou ID, Seto JT, et al. Is evolutionary loss our gain? The role of ACTN3 p.Arg577Ter (R577X) genotype in athletic performance, ageing, and disease. Hum Mutat. 2018;39(12):1774–87. https://doi.org/10.1002/humu.23663.
Ma F, Yang Y, Li X, et al. The association of sport performance with ACE and ACTN3 genetic polymorphisms: a systematic review and meta-analysis. PLoS ONE. 2013;8(1):e54685. https://doi.org/10.1371/journal.pone.0054685.
Ahmetov II, Egorova ES, Gabdrakhmanova LJ, Fedotovskaya ON. Genes and athletic performance: an update. Med Sport Sci. 2016;61:41–54. https://doi.org/10.1159/000445240.
Pickering C, Kiely J. ACTN3: More than just a gene for speed. Front Physiol. 2017;8:1080. https://doi.org/10.3389/fphys.2017.01080.
Silva HH, Silva MRG, Cerqueira F, Tavares V, Medeiros R. Genomic profile in association with sport-type, sex, ethnicity, psychological traits and sport injuries of elite athletes. J Sports Med Phys Fitness. 2022;62(3):418–34. https://doi.org/10.23736/S0022-4707.21.12020-1.
Baltazar-Martins G, Gutierrez-Hellin J, Aguilar-Navarro M, et al. Effect of ACTN3 genotype on sports performance, exercise-induced muscle damage, and injury epidemiology. Sports. 2020;8(7):99. https://doi.org/10.3390/sports8070099.
Del Coso J, Valero M, Salinero JJ, et al. ACTN3 genotype influences exercise-induced muscle damage during a marathon competition. Eur J Appl Physiol. 2017;117(3):409–16. https://doi.org/10.1007/s00421-017-3542-z.
Yang N, MacArthur DG, Gulbin JP, et al. ACTN3 genotype is associated with human elite athletic performance. Am J Hum Genet. 2003;73(3):627–31. https://doi.org/10.1086/377590.
Papadimitriou ID, Lockey SJ, Voisin S, et al. No association between ACTN3 R577X and ACE I/D polymorphisms and endurance running times in 698 Caucasian athletes. BMC Genom. 2018;19(1):13. https://doi.org/10.1186/s12864-017-4412-0.
Eynon N, Duarte JA, Oliveira J, et al. ACTN3 R577X polymorphism and Israeli top-level athletes. Int J Sports Med. 2009;30(9):695–8. https://doi.org/10.1055/s-0029-1220731.
Tharabenjasin P, Pabalan N, Jarjanazi H. Association of the ACTN3 R577X (rs1815739) polymorphism with elite power sports: a meta-analysis. PLoS ONE. 2019;14(5):e0217390. https://doi.org/10.1371/journal.pone.0217390.
McAuley ABT, Hughes DC, Tsaprouni LG, et al. The association of the ACTN3 R577X and ACE I/D polymorphisms with athlete status in football: a systematic review and meta-analysis. J Sports Sci. 2021;39(2):200–11. https://doi.org/10.1080/02640414.2020.1812195.
Furlan AD, Pennick V, Bombardier C, van Tulder M, Editorial Board, Cochrane Back Review Group. 2009 updated method guidelines for systematic reviews in the Cochrane Back Review Group. Spine (Phila Pa 1976). 2009;34(18):1929–41. https://doi.org/10.1097/BRS.0b013e3181b1c99f.
Moher D, Liberati A, Tetzlaff J, Altman DG, Group TP. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLOS Med. 2009;6(7):e1000097. https://doi.org/10.1371/journal.pmed.1000097.
Bassett DR, Howley ET. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med Sci Sports Exerc. 2000;32(1):70–84. https://doi.org/10.1097/00005768-200001000-00012.
Joyner MJ, Coyle EF. Endurance exercise performance: the physiology of champions. J Physiol. 2008;586(1):35–44. https://doi.org/10.1113/jphysiol.2007.143834.
Chamari K, Padulo J. “Aerobic” and “Anaerobic” terms used in exercise physiology: a critical terminology reflection. Sports Med Open. 2015;1(1):9. https://doi.org/10.1186/s40798-015-0012-1.
Maher CG, Sherrington C, Herbert RD, Moseley AM, Elkins M. Reliability of the PEDro scale for rating quality of randomized controlled trials. Phys Ther. 2003;83(8):713–21.
Whitehead A, Whitehead J. A general parametric approach to the meta-analysis of randomized clinical trials. Stat Med. 1991;10(11):1665–77. https://doi.org/10.1002/sim.4780101105.
Sterne JA, Egger M. Funnel plots for detecting bias in meta-analysis: guidelines on choice of axis. J Clin Epidemiol. 2001;54(10):1046–55. https://doi.org/10.1016/s0895-4356(01)00377-8.
Balkó I, Balkó Š, Týnková H, Kohlíková E. The occurence of R577X polymorphism of ACTN3 gene in a selected group of elite fencers. AUC Kinanthropologica. 2015;50(2):5–12. https://doi.org/10.14712/23366052.2015.11.
Ben-Zaken S, Eliakim A, Nemet D, Meckel Y. Genetic variability among power athletes: the stronger versus the faster. J Strength Cond Res. 2019;33(6):1505–11. https://doi.org/10.1519/JSC.0000000000001356.
Coelho DB, Pimenta E, Rosse IC, et al. The alpha-actinin-3 r577x polymorphism and physical performance in soccer players. J Sports Med Phys Fitness. 2016;56(3):241–8.
Eynon N, Alves AJ, Meckel Y, et al. Is the interaction between HIF1A P582S and ACTN3 R577X determinant for power/sprint performance? Metabolism. 2010;59(6):861–5. https://doi.org/10.1016/j.metabol.2009.10.003.
Eynon N, Ruiz JR, Femia P, et al. The ACTN3 R577X polymorphism across three groups of Elite Male European Athletes. PLoS ONE. 2012;7(8):e43132. https://doi.org/10.1371/journal.pone.0043132.
Eynon N, Banting LK, Ruiz JR, et al. ACTN3 R577X polymorphism and team-sport performance: a study involving three European cohorts. J Sci Med Sport. 2014;17(1):102–6. https://doi.org/10.1016/j.jsams.2013.02.005.
Gineviciene V, Jakaitiene A, Aksenov MO, et al. Association analysis of ACE, ACTN3 and PPARGCIA gene polymorphisms in two cohorts of European strength and power athletes. Biol Sport. 2016;33(3):199–206. https://doi.org/10.5604/20831862.1201051.
Grover VK, Verma JP, Kumar A, Sharma N, Tiwari PK. A statistical model for ACTN3 genotype in elite power and speed athletes. J Stat Manag Syst. 2020;23(2):239–48. https://doi.org/10.1080/09720510.2020.1724624.
Kim H, Song KH, Kim CH. The ACTN3 R577X variant in sprint and strength performance. J Exerc Nutr Biochem. 2014;18(4):347–53. https://doi.org/10.5717/jenb.2014.18.4.347.
Pasqualetti M, Onori ME, Canu G, et al. The relationship between ACE, ACTN3 and MCT1 genetic polymorphisms and athletic performance in elite rugby union players: a preliminary study. Genes. 2022;13(6):969. https://doi.org/10.3390/genes13060969.
Petr M, Thiel D, Katerina K, et al. Speed and power-related gene polymorphisms associated with playing position in elite soccer players. Biol Sport. 2022;39(2):355–66. https://doi.org/10.5114/biolsport.2022.105333.
Pimenta EM, Coelho DB, Veneroso CE, et al. Effect of Actn3 Gene on strength and endurance in soccer players. J Strength Cond Res. 2013;27(12):3286–92. https://doi.org/10.1519/JSC.0b013e3182915e66.
Akazawa N, Ohiwa N, Shimizu K, et al. The association of ACTN3 R577X polymorphism with sports specificity in Japanese elite athletes. Biol Sport. 2022;39(4):905–11. https://doi.org/10.5114/biolsport.2022.108704.
Druzhevskaya AM, Ahmetov II, Astratenkova IV, Rogozkin VA. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. Eur J Appl Physiol. 2008;103(6):631–4. https://doi.org/10.1007/s00421-008-0763-1.
Garatachea N, Verde Z, Santos-Lozano A, et al. ACTN3 R577X polymorphism and explosive leg-muscle power in elite basketball players. Int J Sport Physiol Perform. 2014;9(2):226–32. https://doi.org/10.1123/IJSPP.2012-0331.
Kikuchi N, Miyamoto-Mikami E, Murakami H, et al. ACTN3 R577X genotype and athletic performance in a large cohort of Japanese athletes. Eur J Sport Sci. 2016;16(6):694–701. https://doi.org/10.1080/17461391.2015.1071879.
Orysiak J, Mazur-Różycka J, Busko K, Gajewski J, Szczepanska B, Malczewska-Lenczowska J. Individual and combined influence of ACE and ACTN3 genes on muscle phenotypes in Polish athletes. J Strength Cond Res. 2018;32(10):2776–82. https://doi.org/10.1519/JSC.0000000000001839.
Chen W, Zhang B, Li Y, Liang T. Association between rs1815739 polymorphism of ACTN3 gene and athletic ability in chinese sprinters. J Sci Sport Exerc. 2020;2(2):113–9. https://doi.org/10.1007/s42978-020-00058-1.
Melian Ortiz A, Laguarta-Val S, Varillas-Delgado D. Muscle work and its relationship with ACE and ACTN3 polymorphisms are associated with the improvement of explosive strength. Genes. 2021;12(8):1177. https://doi.org/10.3390/genes12081177.
Papadimitriou ID, Lucia A, Pitsiladis YP, et al. ACTN3 R577X and ACE I/D gene variants influence performance in elite sprinters: a multi-cohort study. BMC Genom. 2016;17(1):285. https://doi.org/10.1186/s12864-016-2462-3.
Massidda M, Vona G, Calò C. Association between the ACTN3 R577X polymorphism and artistic gymnastic performance in Italy. Genet Test Mol Biomark. 2009;13(3):377–80. https://doi.org/10.1089/gtmb.2008.0157.
Juffer P, Furrer R, González-Freire M, et al. Genotype distributions in top-level soccer players: a role for ACE? Int J Sports Med. 2009;30(5):387–92. https://doi.org/10.1055/s-0028-1105931.
Alfred T, Ben-Shlomo Y, Cooper R, et al. ACTN3 genotype, athletic status, and life course physical capability: meta-analysis of the published literature and findings from nine studies. Hum Mutat. 2011;32(9):1008–18. https://doi.org/10.1002/humu.21526.
Erskine RM, Williams AG, Jones DA, Stewart CE, Degens H. The individual and combined influence of ACE and ACTN3 genotypes on muscle phenotypes before and after strength training. Scand J Med Sci Sports. 2014;24(4):642–8. https://doi.org/10.1111/sms.12055.
Shang X, Zhang F, Zhang L, Huang C. ACTN3 R577X polymorphism and performance phenotypes in young Chinese male soldiers. J Sports Sci. 2012;30(3):255–60. https://doi.org/10.1080/02640414.2011.619203.
Broos S, Malisoux L, Theisen D, et al. Evidence for ACTN3 as a speed gene in isolated human muscle fibers. PLoS ONE. 2016;11(3):e0150594. https://doi.org/10.1371/journal.pone.0150594.
Moran CN, Yang N, Bailey MES, et al. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. Eur J Hum Genet. 2007;15(1):88–93. https://doi.org/10.1038/sj.ejhg.5201724.
Vincent B, De Bock K, Ramaekers M, et al. ACTN3 (R577X) genotype is associated with fiber type distribution. Physiol Genom. 2007;32(1):58–63. https://doi.org/10.1152/physiolgenomics.00173.2007.
Santiago C, Rodríguez-Romo G, Gómez-Gallego F, et al. Is there an association between ACTN3 R577X polymorphism and muscle power phenotypes in young, non-athletic adults? Scand J Med Sci Sports. 2010;20(5):771–8. https://doi.org/10.1111/j.1600-0838.2009.01017.x.
Bell W, Colley JP, Evans WD, Darlington SE, Cooper SM. ACTN3 genotypes of Rugby Union players: distribution, power output and body composition. Ann Hum Biol. 2012;39(1):19–27. https://doi.org/10.3109/03014460.2011.632648.
Delmonico MJ, Zmuda JM, Taylor BC, et al. Association of the ACTN3 genotype and physical functioning with age in older adults. J Gerontol A Biol Sci Med Sci. 2008;63(11):1227–34. https://doi.org/10.1093/gerona/63.11.1227.
McCauley T, Mastana SS, Hossack J, Macdonald M, Folland JP. Human angiotensin-converting enzyme I/D and alpha-actinin 3 R577X genotypes and muscle functional and contractile properties. Exp Physiol. 2009;94(1):81–9. https://doi.org/10.1113/expphysiol.2008.043075.
Ruiz JR, Fernandez del Valle M, Verde Z, et al. ACTN3 R577X polymorphism does not influence explosive leg muscle power in elite volleyball players. Scand J Med Sci Sports. 2011;21(6):E34–41. https://doi.org/10.1111/j.1600-0838.2010.01134.x.
Ginevičienė V, Pranculis A, Jakaitienė A, Milašius K, Kučinskas V. Genetic variation of the human ACE and ACTN3 genes and their association with functional muscle properties in Lithuanian elite athletes. Medicina. 2011;47(5):40. https://doi.org/10.3390/medicina47050040.
Issurin VB. Evidence-based prerequisites and precursors of athletic talent: a review. Sports Med. 2017;47(10):1993–2010. https://doi.org/10.1007/s40279-017-0740-0.
Niemi AK, Majamaa K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. Eur J Hum Genet. 2005;13(8):965–9. https://doi.org/10.1038/sj.ejhg.5201438.
Ahmetov II, Druzhevskaya AM, Astratenkova IV, Popov DV, Vinogradova OL, Rogozkin VA. The ACTN3 R577X polymorphism in Russian endurance athletes. Br J Sports Med. 2010;44(9):649–52. https://doi.org/10.1136/bjsm.2008.051540.
Gibala MJ, McGee SL. Metabolic adaptations to short-term high-intensity interval training: a little pain for a lot of gain? Exerc Sport Sci Rev. 2008;36:2. https://doi.org/10.1097/JES.0b013e318168ec1f.
Gillen JB, Gibala MJ. Is high-intensity interval training a time-efficient exercise strategy to improve health and fitness? Appl Physiol Nutr Metab. 2014;39(3):409–12. https://doi.org/10.1139/apnm-2013-0187.
Hughes DC, Ellefsen S, Baar K. Adaptations to endurance and strength training. Cold Spring Harb Perspect Med. 2018;8(6):a029769. https://doi.org/10.1101/cshperspect.a029769.
Yang N, Macarthur DG, Wolde B, et al. The ACTN3 R577X polymorphism in east and west African athletes. Med Sci Sports Exerc. 2007;39(11):1985–8. https://doi.org/10.1249/mss.0b013e31814844c9.
Shang X, Huang C, Chang Q, Zhang L, Huang T. Association between the ACTN3 R577X polymorphism and female endurance athletes in China. Int J Sports Med. 2010;31(12):913–6. https://doi.org/10.1055/s-0030-1265176.
Seto JT, Quinlan KGR, Lek M, et al. ACTN3 genotype influences muscle performance through the regulation of calcineurin signaling. J Clin Invest. 2013;123(10):4255–63. https://doi.org/10.1172/JCI67691.
MacArthur DG, Seto JT, Raftery JM, et al. Loss of ACTN3 gene function alters mouse muscle metabolism and shows evidence of positive selection in humans. Nat Genet. 2007;39(10):1261–5. https://doi.org/10.1038/ng2122.
Stuart CA, McCurry MP, Marino A, et al. Slow-twitch fiber proportion in skeletal muscle correlates with insulin responsiveness. J Clin Endocrinol Metab. 2013;98(5):2027–36. https://doi.org/10.1210/jc.2012-3876.
Pasqua LA, Bueno S, Artioli GG, et al. Influence of ACTN3 R577X polymorphism on ventilatory thresholds related to endurance performance. J Sports Sci. 2016;34(2):163–70. https://doi.org/10.1080/02640414.2015.1040823.
Broos S, Van Leemputte M, Deldicque L, Thomis MA. History-dependent force, angular velocity and muscular endurance in ACTN3 genotypes. Eur J Appl Physiol. 2015;115(8):1637–43. https://doi.org/10.1007/s00421-015-3144-6.
Norman B, Esbjörnsson M, Rundqvist H, Österlund T, Glenmark B, Jansson E. ACTN3 genotype and modulation of skeletal muscle response to exercise in human subjects. J Appl Physiol. 2014;116(9):1197–203. https://doi.org/10.1152/japplphysiol.00557.2013.
Silva MSM, Bolani W, Alves CR, et al. Elimination of influences of the ACTN3 R577X variant on oxygen uptake by endurance training in healthy individuals. Int J Sports Physiol Perform. 2015;10(5):636–41. https://doi.org/10.1123/ijspp.2014-0205.
Zilberman-Schapira G, Chen J, Gerstein M. On sports and genes. Recent Pat DNA Gene Seq. 2012;6(3):180–8. https://doi.org/10.2174/187221512802717367.
Malczewska-Lenczowska J, Orysiak J, Majorczyk E, et al. Total hemoglobin mass, aerobic capacity, and HBB gene in polish road cyclists. J Strength Cond Res. 2016;30(12):3512–9. https://doi.org/10.1519/JSC.0000000000001435.
Delmonico MJ, Kostek MC, Doldo NA, et al. Alpha-actinin-3 (ACTN3) R577X polymorphism influences knee extensor peak power response to strength training in older men and women. J Gerontol A Biol Sci Med Sci. 2007;62(2):206–12. https://doi.org/10.1093/gerona/62.2.206.
Kikuchi N, Nakazato K, Min SK, Ueda D, Igawa S. The ACTN3 R577X polymorphism is associated with muscle power in male Japanese athletes. J Strength Cond Res. 2014;28(7):1783–9. https://doi.org/10.1519/JSC.0000000000000338.
Chiu LL, Wu YF, Tang MT, Yu HC, Hsieh LL, Hsieh SSY. ACTN3 genotype and swimming performance in Taiwan. Int J Sports Med. 2011;32(06):476–80. https://doi.org/10.1055/s-0030-1263115.
Clarkson PM, Devaney JM, Gordish-Dressman H, et al. ACTN3 genotype is associated with increases in muscle strength in response to resistance training in women. J Appl Physiol. 2005;99(1):154–63. https://doi.org/10.1152/japplphysiol.01139.2004.
Walsh S, Liu D, Metter EJ, Ferrucci L, Roth SM. ACTN3 genotype is associated with muscle phenotypes in women across the adult age span. J Appl Physiol. 2008;105(5):1486–91. https://doi.org/10.1152/japplphysiol.90856.2008.
Pranckeviciene E, Gineviciene V, Jakaitiene A, Januska L, Utkus A. Total genotype score modelling of polygenic endurance-power profiles in Lithuanian elite athletes. Genes. 2021;12(7):1067. https://doi.org/10.3390/genes12071067.
Tanisawa K, Wang G, Seto J, et al. Sport and exercise genomics: the FIMS 2019 consensus statement update. Br J Sports Med. 2020;54(16):969–75. https://doi.org/10.1136/bjsports-2019-101532.
Acknowledgements
Not applicable.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was supported by the Deutsche Forschungsgemeinschaft (DFG) and Open Access Publishing Fund of the University of Freiburg, Germany.
Author information
Authors and Affiliations
Contributions
Study design, data analysis and manuscript writing involved the collaboration of EE, BB and HZ. JDC, ACH, AM, IL, and UG contributed to data analysis, manuscript writing, and data evaluation. All authors have carefully reviewed and accepted the order of authors and the final version of the manuscript.
Corresponding authors
Ethics declarations
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing interests
The authors declare that they have no competing interest relevant to the content of this review.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
The funnel plots for the comparison of genotype frequencies: Figure S1. Funnel plot of ACTN3 R577X polymorphism in power athletes (RR vs. RX genotypes). Figure S2. Funnel plot of ACTN3 R577X polymorphism in power athletes (RR vs. XX genotypes). Figure S3. Funnel plot of ACTN3 R577X polymorphism in power athletes (RX vs. XX genotypes). Figure S4. Funnel plot of ACTN3 R577X polymorphism in power athletes (R vs. X alleles). Figure S5. Funnel plot of RR genotype expression in power athletes versus controls. Figure S6. Funnel plot of RX genotype expression in power athletes versus controls. Figure S7. Funnel plot of XX genotype expression in power athletes versus controls. Figure S8. Funnel plot of R allele in power athletes versus controls. Figure S9. Funnel plot of X allele in power athletes versus controls. Figure S10. Funnel plot of RR genotype expression in power versus endurance athletes. Figure S11. Funnel plot of RX genotype expression in power versus endurance athletes. Figure S12. Funnel plot of XX genotype expression in power versus endurance athletes. Figure S13. Funnel plot of R allele in power athletes versus endurance athletes. Figure S14. Funnel plot of X allele in power athletes versus endurance athletes.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
El Ouali, E., Barthelemy, B., Del Coso, J. et al. A Systematic Review and Meta-analysis of the Association Between ACTN3 R577X Genotypes and Performance in Endurance Versus Power Athletes and Non-athletes. Sports Med - Open 10, 37 (2024). https://doi.org/10.1186/s40798-024-00711-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40798-024-00711-x