
Abstract
Background
Coral reefs are one of the most biodiverse and productive ecosystems, providing habitat for a vast of species. Reef-building scleractinian corals with a symbiotic microbiome, including bacteria, archaea, viruses and eukaryotic microbes, are referred to coral holobionts. Among them, coral diseases, mainly caused by Vibrio spp., have significantly contributed to the loss of coral cover and diversity. Habitat filtering across the globe has led to a variety structure of marine bacterial communities. Coral species, quantity and characteristics are significant differences between the Xisha Islands and Daya Bay (Guangdong Province). Thus, the Vibrio communities may be distinct between coral rich and poor areas.
Results
Through comparison of Vibrio dynamics between coral-rich (Xisha Islands) and coral-poor (Daya Bay) locations, we uncovered differences in Vibrio abundance, diversity, community composition and assembly mechanisms associated with corals. The higher abundance of Vibrio in coral rich areas may indicate a strong interaction between vibrios and corals. V. campbellii, Paraphotobacterium marinum and V. caribbeanicus were widely distributed in both coral rich and poor areas, likely indicating weak species specificity in the coral-stimulated growth of Vibrio. Random-forest prediction revealed Vibrio species and Photobacterium species as potential microbial indicators in the coral rich and coral poor areas, respectively. Ecological drift rather than selection governed the Vibrio community assembly in the Xisha Islands. Comparatively, homogenizing selection was more important for the Daya Bay community, which may reflect a role of habitat filtration.
Conclusion
This study revealed the different distribution pattern and assembly mechanism of Vibrio spp. between coral rich and poor areas, providing the background data for the research of Vibrio community in coral reef areas and may help the protection of coral reef at the biological level. The main reasons for the difference were different number and species of corals, environmental (e.g., temperature) and spatial factors. It reflected the strong interaction between Vibrio and corals, and provided a new perspective for the investigation of Vibrio in coral reef ecosystem.
Similar content being viewed by others


Background
Coral reef is the most biologically diverse and productive ecosystem on earth, providing habitat for a vast array of species that build up a three-dimensional reef matrix by the calcium carbonate skeletons secreted from corals over time [1]. Though covering a small area of the ocean floor, coral reefs sustain a third of all described marine species [2]. For instance, the biomass of resident reef fish averages ~ 1000 kg/hectare [3]. Corals are usually symbiotic with algae, such as zooxanthellae for nutrient cycle to maintain the stability and growth of the reef [4]. Prokaryotic microorganisms also play important roles in the coral reef ecosystem, such as microbial-coral diseases, and interactions between coral symbiotic algae and microorganisms [5,6,7]. Bacteria can contribute to the physiology and health of the coral host through nutrient acquisition and metabolic cycling [8]. Shnit-Orland and Kushmaro have found that 25–70% of coral mucus-associated bacteria exhibited antibacterial activity, suggesting that coral mucus-associated bacteria may protect their coral hosts against pathogens [9]. Furthermore, nitrogen-cycling microbes may be critical to the stability of coral-algal symbiosis and the function of holobiont, and disturbances of microbial nitrogen cycling may be closely associated with coral bleaching and disease [10]. In addition, microbial processes can influence the resilience of coral reef ecosystems by influencing the colonization of larvae through chemical cues [11]. Thus, the diversity and metabolic activity of surrounding microorganisms in seawater may provide important solutions to potentially protect coral reefs.
Corals and their symbiotic algae may also exert influences on microbes in the surrounding seawater. For example, corals release mucus affecting microbial abundance and composition [12]. Coral mucus is rich in organic matter and nutrients, with concentrations of total organic carbon and nitrogen being 2–4 times higher than that of the surrounding seawater [13, 14]. Previous studies have shown a significant increase in microbial abundance in seawater with coral mucus presence [14]. Moreover, the exudates of coral symbiotic algae can transport sugars into the environment, promoting growth of planktonic bacteria [15], particularly Vibrio including pathogenic species such as Vibrio cholerae [16]. Comparison of microorganisms from four different environmental niches (Acropora palifera, Acropora aspera, sandy substrate and open water) in the Great Barrier Reef has shown increased occurrence frequency of Vibrio, Pseudoalteromonas and Alteromonas in coral seawater niches [17]. In a shallow reef in St. John, U.S. Virgin Islands, bacteria and archaea in the surrounding seawater showed significant temporal and spatial variations, but only few vibrios were present in the total community compositions [18]. However, the ecological distribution and community dynamics of Vibrio spp. in coral surrounding seawaters are still unclear.
Vibrio spp. is a group of Gram-negative rod-shaped bacteria belonging to the class Gammaproteobacteria with facultative fermentative metabolisms [19], which is highly heterogeneous and abundant in various aquatic environments [20, 21]. At present, the research on Vibrio in coral reefs mainly focuses on coral diseases, such as V. coralliilyticus causing white syndrome, V. alginolyticus causing yellow band disease and V. natriegens causing white spot disease [7]. Vibrio multiplies rapidly, has a short generation time, and responds quickly to nutrient pulses likely due to their highly plastic genomes and wide metabolic ranges [21]. Meanwhile, vibrios can be subject to predation by bacteriophages and protozoa, such as Vibrio alginolyticus and Vibrio natriegens [22, 23]. Based on these knowledges, the biogeochemical roles of Vibrio were underestimated for a long time. Vibrio community may rapidly respond to nutrient pulses produced by corals, which may cause the community dynamics in coral reef ecosystems differed from other marine environments. Previous study has found that V. hyugaensis, V. owensii and V. harveyi were the dominant species, and the influence of temperature on these species was evaluated in the coral reefs of Ishigaki (Japan) [24]. As a typical coral conservation area, the Vibrio community of Dongshan Bay (Fujian, China) was dominated by V. fortis, V. natriegens and V. pomeroyi, revealing a distance-decay pattern spanning four seasons [19]. The distribution pattern of corals in different coral reef environments may dominate the dynamics and assembly mechanisms of Vibrio community. However, the correlations between Vibrio spp. and corals needs to be further studied.
Corals can be divided into "reef" and "non-reef" coral communities, and the latter is characterized by the inability to accumulate calcium carbonate [25]. The coral species and the local reef environment can influence the composition of microbial community [26]. The majority of China’s coral reefs are found in the South China Sea, whereas only limited and scattered subtidal coral communities are found from the west coast of Leizhou Peninsula (Guangdong, China) to Dongshan Bay (Fujian, China) [27, 28]. The Xisha Islands (coral rich areas) mainly consisting of 36 atolls, is one of the four groups of islands located in the South China Sea, with plentiful coral reefs [29,30,31]. A total of 213 species of scleractinian corals (belonging to 43 genera and 16 families) have been discovered therein [32]. Daya Bay (coral poor areas) is one of the few coastal bays in China with coral distribution [33]. The corals scattered in the Daya Bay mainly appear in the form of communities and have not developed into coral reefs due to climate constraints [34]. A total of 44 species (belonging to 17 genera and 9 families) and 5 unclassified species of scleractinian corals are found there [35]. At present, the studies on microbial dynamics in Daya Bay and Xisha Islands mainly focus on the dynamic of total bacteria or the changes of culturable bacteria [36, 37], with no attention on the dynamic changes of the vibrionic community. Given that different environments affect microbial colonization [26], Vibrio may have different dynamic characteristics in these two coral environments. In this study, the dynamics of Vibrio communities in seawater from the coral rich and poor areas were investigated by qPCR and high-throughput sequencing techniques. The key factors affecting the community structure and assembly mechanism of Vibrio community in distinct coral environments were illustrated.
Methods
Water sampling and physicochemical parameter determination
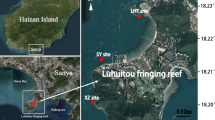
To explore the planktonic Vibrio communities in distinct coral areas, waters from the Xisha Islands and Daya Bay were collected by the R/V Yuezhanyuke10 from a total of 48 sites during 31 August and 30 September, 2020 (Fig. 1). A total of 85 samples were collected, and detailed information of the samples was recorded in Additional file 1: Table S1. At each sample, 1 L (L) of water was filtered through polycarbonate membranes (Millipore Corporation, Billerica, MA, USA) 3-μm for particle-associated (PA) and 0.22-μm for free-living (FL) microbes, respectively. All filters were stored in liquid nitrogen onboard and transferred to − 80 °C in the laboratory until DNA extraction. The physicochemical parameters of waters were determined in situ and in the laboratory. CTD was used to monitor the water chemistries such as temperature, salinity, pH, dissolved oxygen (DO) and suspended solid (SS). Water samples for dissolved inorganic nutrients (NO2–, NO3– and NH4+), and Chlorophyll a (Chl a) analyses were collected according to our previous work [38]. Dissolved inorganic nitrogen (DIN) were calculated as the sum of NO3−, NO2− and NH4+ [39]. After acid fumigation to remove the carbonate fraction, total nitrogen (TN) was determined by an elemental analyzer (vario MICRO cube EA, Elementar, Germany) which interfaced with a continuous flow isotope ratio mass spectrometer (Isoprime IRMS, Elementar, Germany) [40, 41]. Detection of the total phosphorus (TP) was conducted followed the description of Church et al. [42] and Varol et al. [43]. Additionally, the identification and abundance of coral species were provided by the Laboratory of Marine Biodiversity Research, Third Institute of Oceanography, Ministry of Natural Resources. Briefly, three transects (50 m in length) of different depths (5, 10 and 15 m) were set up at each sampling sites. A SCUBA diver performed Point Intercept Transect (PIT) video sampling at each transects, and another diver took close-up photographs of various corals under the tapeline and collected some specimens for species identification according to standard procedures [32]. Video transects were analyzed in the laboratory using a point sampling technique according to the taxonomic criteria [32], and the coral close-up photographs and the coral specimens were used to assist the species identification. The number of coral species were calculated based on the identification results.

Map showing locations of study area and sampling sites. [The map was created using Ocean Data View (version 5.5.2; R. Schlitzer, Ocean Data View, https://odv.awi.de, 2021.)]
DNA extraction and quantitative PCR (qPCR)
Genomic DNA was extracted according to the previous report [38]. The extracted DNA was resuspend using 50 μL TE buffer (1 M Tris–HCl, 0.5 M EDTA, pH 8.0) and preserved at – 80 ℃ until use. Through 16S rRNA gene-targeted quantitative PCR (qPCR) with SYBR-green detection, the absolute abundance of total Vibrio on 3- and 0.22-μm-pore-size membranes were tracked. Briefly, the StepOnePlus real-time PCR system (Applied Biosystems) and StepOne software version 2.2 was operated to performed the qPCR. V567F (5′-GGCGTAAAGCGCATGCAGGT-3′) and V680R (5′-GAAATTCTACCCCCCTCTACAG-3′) primers were utilized to quantify the Vibrio spp. [20, 44]. The reaction mixture and cycling conditions referred to the descriptions of Wang et al. [38] and Liang et al. [45]. The standard was prepared using 16S rRNA gene of Vibrio rotiferianus WXL191 (our laboratory) and the preparation methods followed our previous work [38]. qPCR was carried out in triplicates and all amplification efficiencies were between 95 and 105% with R2 values > 0.99.
High-throughput sequencing and reads processing
The Vibrio-specific 16S rRNA gene primers V169F (5′-GGATAACC/TATTGGAAACGATG-3′) and V680R (5′-GAAATTCTACCCCCCTCTACAG-3′) were utilized to determine the composition of Vibrio community [45]. The reaction mixture and cycling conditions followed the descriptions of Wang et al. [38]. After confirming positive amplification, sequencing was performed on the Illumina Miseq PE300 platform at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China). Raw reads were trimmed with FASTP, removing those short length sequences (< 100 bp) and low quality (< 20) [46]. Paired-end DNA sequences were then joined using FLASH with at least a 10-bp overlap and < 5% mismatches [47]. Moreover, chimeric sequences, barcode sequences and primers were removed using the DADA2 plug-in of software QIIME2 [48, 49]. Then, the sequences of Vibrio spp. were clustered into operational taxonomic units (OTUs) by QIIME2 software. The taxonomy of each OTUs representative sequences were assigned by local-blast against the SILVA v138 database (a minimum support threshold of 70%), and they were reassigned against the EzBioCloud database (https://www.ezbiocloud.net/) to acquire a more accurate taxonomic identification.
Statistical analysis
To minimize biases associated with sequencing coverage, the number of sequences for each sample was homogenized to the lowest number (32,029 reads) by running a script in R software. The differences of between separate groups of physicochemical parameters were calculated by Dunn's test. Alpha (α)-diversity was indicated by Shannon, Chao 1 and Simpsoneven indices, which were calculated using the “vegan” package. The differences between separate groups of samples were examined by t-test. For Beta (β)-diversity, the non-metric multidimensional scaling (NMDS) was performed at the OTU level by Canoco 5.0 software [50]. The subsequent analysis of similarities (ANOSIM) was performed using the anosim function with 999 permutations in “vegan”. The niche widths of the most thirty abundant OTUs were calculated by “vegan” and “spaa”. The random forest machine learning was performed with “randomForest” package in R. We used variation partition analysis (VPA), performed by the “vegan” package, to estimate the relative contributions of geographic distance, environmental factors and coral species to Vibrio community structure. To reveal the relationship between environmental factors and microbial communities, Mantel test based on Pearson’s correlations was carried out by the “ggcor” package. To determine the relationship between Vibrio species, α-diversity, the abundance of Vibrio and various factors, the software IBM SPSS Statistica (v 23.0.) was operated to calculate Spearman's rank correlation coefficient. Additionally, a null model analysis was carried out to quantify the relative contributions of different ecological processes, which was calculated using the “picante” package.
Results
Environmental conditions and coral distribution
Water temperature, salinity, SS, Chl a, TN, TP and pH varied significantly between Daya Bay and Xisha Islands (P < 0.05; Additional file 1: Fig. S1). Compared to the Daya Bay, the pH (8.11 ± 0.06), salinity (33.85 ± 0.07; practical salinity units [PSU]), TP (0.009 ± 0.003 mg L−1), SS (2.61 ± 0.60 mg L−1) and Chl a (0.38 ± 0.34 μg L−1) were significantly lower in the Xisha Islands. However, the Xisha Islands featured higher temperature (30.84 ± 0.41 ℃) and TN concentration (0.23 ± 0.07 mg L−1; Table 1). The abundance and number of corals species was higher in Xisha Islands than in Daya Bay. All corals were assigned to 15 families and 1 unclassified family, with 9 families of corals in Daya Bay, and 15 families and the unclassified coral in Xisha Islands. The number of corals in the Xisha Islands was nearly 6 times than that in Daya Bay. Difference in dominant corals was observed between Daya Bay and Xisha Islands (Additional file 1: Fig. S2). Poritidae was the most dominant coral in the Daya Bay, followed by Merulinidae and Acroporidae, whereas the dominant corals of the Xisha Islands were Merulinidae, followed by Poritidae, Acroporidae and Pocilloporidae. Several kinds of corals (i.e., Pocilloporidae, Oulastreidae, Coscinaraeidae, Fungiidae, Astrocoeniidae, Lobophylliidae and the unclassified one) were unique at the Xisha Islands. Detailed information on environmental parameters and coral numbers are provided in Additional file 1: Tables S2 and S3, respectively.
Total Vibrio abundance
Vibrio abundance in the Xisha Islands (2.92 ± 0.78 lg copies/mL) was significantly higher than that in Daya Bay (2.76 ± 0.46 lg copies/mL; P < 0.05; t-test), representing an obvious spatial distribution pattern (Fig. 2a). Meanwhile, both the average abundance of FL and PA Vibrio were higher in Xisha Islands than in Daya Bay, although the difference was not significant (Fig. 2a). Detailed information of Vibrio abundance was listed in Additional file 1: Table S4. This variation of Vibrio abundance between the two areas largely related to the corals. In both the two areas, the abundance of FL and PA Vibrio showed significantly positive correlations to several kinds of corals (i.e., Pocilloporidae, Poritidae, Oulastreidae, Agariciidae, Acroporidae, Merulinidae, Coscinaraeidae, Fungiidae, Euphylliidae and unclassified corals; P < 0.05 or 0.01; Fig. 2d). The Vibrio abundance was positively correlated with Pocilloporidae, Poritidae, Agariciidae, Acroporidae, Merulinidae, Coscinaraeidae and Fungiidae in Xisha Islands, whereas no correlation was found in Daya Bay (P < 0.05 or 0.01; Additional file 1: Fig. S3).

The abundance and diversity of Vibrio spp. a Total Vibrio abundance (lg copies/ml) across all samples and their differences among different groups. DY Daya Bay, XS Xisha Islands, PA particle-associated group, FL free-living group. *P < 0.05. b α-Diversity indices of Vibrio community. Student’s t-test showed significant difference between DY and XS. **P < 0.01; ***P < 0.001. c Nonmetric multidimensional scaling ordination of vibrios based on the Bray–Curtis dissimilarity. d Spearman correlations between Vibrio spp. abundance, α-diversity indices and the physicochemical parameters. *P < 0.05; **P < 0.01
Diversity estimators of Vibrio spp.
To analyze the diversity of the Vibrio community, 16S rRNA gene amplicons were sequenced by the Illumina Miseq PE300 platform. The high-throughput sequencing yielded 4,412,547 clean reads from 85 samples ranging from 32,029 to 82,544 reads per sample. After discarding the singletons and rarefaction, 32,029 sequences per sample were left. The total sequences were clustered into 549 operational taxonomic units (OTUs) based on a 97% sequence similarity level. And, the coverages of all samples were close to 99.9%, indicating that the current sequences could represent the real situation of Vibrio community in the studies sites.
For α-diversity indices, evenness, richness and diversity of Vibrio were variable between the two areas (Fig. 2b). Although the richness (Chao 1) had no significant difference between Daya Bay and Xisha Islands, Vibrio evenness in the Xisha Islands was higher than that in Daya Bay (Simpsoneven, t-test, P < 0.01), leading to a higher diversity level (Shannon, t-test, P < 0.001). Overall, correlation analysis demonstrated that Pocilloporidae, Agariciidae, Coscinaraeidae, Fungiidae, Plesiastreidae, Lobophylliidae and Euphylliidae were related to changes in α-diversity (Fig. 2d). However, the variation of α-diversity in the Xisha Islands showed association with Acroporidae, Psammocoridae, Coscinaraeidae and Astrocoeniidae, and the changes in the Daya Bay were associated with Poritidae, Acroporidae, Psammocoridae and Plesiastreidae (Additional file 1: Fig. S3). We also analysed the β-diversity among all samples. NMDS analysis based on Bray–Curtis distances showed spatial separation of Vibrio compositions between the two regions (Analysis of similarity [ANOSIM]; R = 0.2204; P = 0.001; Fig. 2c). The β-diversity of Vibrio community in the Xisha Islands was higher than that in Daya Bay (P < 0.001; Additional file 1: Fig. S4). Detailed information on the diversity parameters was recorded in Additional file 1: Table S4.
The community compositions of Vibrio spp.
After re-annotated against the EzBioCloud database, Vibrionaceae accounted for 52.60% of the total sequences, and Vibrio spp. accounted for 78.92% of Vibrionaceae sequences. The abundance of the top 30 OTUs accounted for 92.85% of all species (Fig. 3a). Among them, OTU30 and OTU493 were annotated to the genera Vibrio and Catenovulum, respectively.

Characteristics of microbial communities. a Compositions of Vibrio spp. at the species level. b The niche widths of the top 30 OTUs. c The most important 40 OTUs reducing the uncertainty in the prediction of geography (Daya Bay and Xisha Islands) based on their mean decrease in accuracy. d Relative proportions of the most important 40 OTUs in the Xisha Islands and Daya Bay
V. campbellii occupied the highest relative abundance across all samples (31.03%), and was more abundant in the Daya Bay than in the Xisha Islands (Additional file 1: Fig. S5; P < 0.001). Furthermore, Vibrio sp. OTU30, P. leiognathi, P. aquimaris OTU37 and V. sagamiensis also had higher abundance at the Daya Bay (P < 0.05), whereas V. neocaledonicus, V. xuii, V. aquimaris, V. fortis, V. navarrensis, V. parahaemolyticus, V. ponticus, V. alginolyticus, V. maritimus, and V. stylophorae were more abundant at the Xisha Islands (P < 0.05; Additional file 1: Fig. S5). Additionally, the Daya Bay could be divided into inside (DY01 to DY13) and outside sites (DY14 to DY23) according to the geographical location (Fig. 1). Five species had significant differences between the inside and outside sites by Dunn's test, i.e., Paraphotobacterium marinum (P < 0.01), Vibrio sp. OTU30 (P < 0.001), P. aquimaris OTU37 (P < 0.01), V. ponticus (P < 0.05) and P. aquimaris OTU530 (P < 0.01). Further, we calculated the niche widths of the top 30 OTUs to explore their adaptation. V. campbellii has the largest niche width, which may have the most adaptable to this environment (Fig. 3b).
To distinguish these two different coral environments, a random-forest machine-learning model was established to find the important features that could distinguish the coral rich and poor areas (Fig. 3c). The model had the highest accuracy (lowest out of bag (OOB) estimated error rate) when the number of top important features reached 40 OTUs (Additional file 1: Fig. S6). Therefore, the most characteristic OTUs (top 40) were selected as a group of biomarkers to distinguish the two habitats, such as P. aquimaris OTU37 representing coral poor area and V. owensii OTU505 representing coral rich area (Fig. 3c). Meanwhile, the relative abundance of 40 characteristic OTUs showed differences in the two regions (Fig. 3d).
Various factors governing the spatial dynamics of Vibrio spp.
To explore the key drivers shaping the Vibrio community, biotic and abiotic correlations were analyzed by VPA (Fig. 4). For Daya Bay, the pure effect of spatial factors (11.20%) was significantly greater than environmental (8.40%) and coralline (4.60%) variables (Fig. 4a), whereas the pure effect of coralline variables (23.66%) was greater than environmental (5.26%) and spatial (2.50%) factors in the Xisha Islands (Fig. 4c). However, 57.80% and 66.35% of the variation in Daya Bay and Xisha Islands were unexplained, respectively. Mantel test was used to evaluate the specific factors affecting Vibrio community structure. The Vibrio community in the Xisha Islands was significantly correlated with Pocilloporidae, Coscinaraeidae and Diploastraeidae (P < 0.05 or P < 0.01; Fig. 4). Detailed parameters were provided in Additional file 1: Table S5.

Effects of various factors on Vibrio community. a, c Variation partitioning analyses of the Vibrio community composition between biotic and abiotic variables. a, Daya Bay; c, Xisha Islands. E environmental factors, S spatial factors, C coralline factors. b, d Relationships between microbial community and other factors in different areas using Mantel test. b Daya Bay (DY); d Xisha Islands (XS). DO dissolved oxygen, DIN dissolved inorganic nitrogen, TN total nitrogen, TP total phosphorus, Chl a chlorophyll a, SS suspended solid, Tem temperature, Sal salinity
The Spearman’s rank correlation coefficients between the top 30 OTUs and factors were calculated (Fig. 5). As the most abundant specie in these two areas, V. campbellii showed significantly positive correlations with Poritidae, Acroporidae, Merulinidae, Dendrophylliidae, DO and SS in Daya Bay (P < 0.05; Fig. 5a), whereas negatively correlated to Pocilloporidae, Agariciidae, Acroporidae, Diploastraeidae, Euphylliidae and the unclassified corals in Xisha Islands (P < 0.05 or 0.01; Fig. 5b). The abundant species in the Daya Bay, i.e., Vibrio sp. OTU30, was positively correlated with Merulinidae and temperature, and significantly negative to pH, DO and salinity (P < 0.01; Fig. 5a). Similarly, V. neocaledonicus in the Xisha Islands only had positive correlation with salinity (P < 0.01; Fig. 5b).

The correlations of the most abundant 30 OTUs with abiotic and biotic factors. a Daya Bay; b Xisha Islands. *P < 0.05; **P < 0.01
Assembly process of the Vibrio community
We used the βNTI metric to quantify the relative importance of deterministic (|βNTI|> 2) or stochastic (|βNTI|< 2) factors to community structure. According to the description of Stegen et al., ecological processes are divided into deterministic processes (i.e., heterogeneous and homogeneous selection) and stochastic processes (i.e., homogenizing dispersal, ecological drift and dispersal limitation) [51]. There was a significant variation in βNTI values between Daya Bay and Xisha Islands via t-test (P < 0.001; Fig. 6b). The βNTI values of Vibrio in the Daya Bay were between − 4 and − 1, indicating that both deterministic and stochastic processes had an impact on Vibrio community structure. The βNTI values of Vibrio in the Xisha Islands were mostly between − 2 and 2, indicating the dominant effects from stochastic processes (Fig. 6a). The community of vibrios in the Daya Bay were jointly governed by ecological drift (47.9%) and homogeneizing selection (49.7%). It was noteworthy that ecological drift contributed the largest fraction (93.4%) to the community structure in the Xisha Islands (Fig. 6c).

Community assembly mechanisms of Vibrio communities. a The distribution of βNTI in different groups. b Comparison of βNTI between Daya Bay and Xisha Islands. ***P < 0.001. c The assembly processes of Vibrio communities with the null model. HeS Heterogeneous selection, HoS Homogenizing selection, DL Dispersal limitation, HD Homogenizing dispersal, ED Ecological drift
Discussion
Coral reefs are one of the most productive and biologically diverse ecosystems on earth. The distribution patterns of microorganism in distinct coral environments often show differences [52, 53]. Vibrio is related to the health of corals, so that the dynamics of Vibrio spp. may be various between different coral environments. Previous reports on the ecological distribution of Vibrio spp. mostly focused on estuarine, bay, or general marine environments [19, 38, 45, 54, 55]. Here, we assessed the abundance, distribution and community assembly mechanisms of Vibrio between coral rich (Xisha Islands) and coral poor (Daya Bay) environments. This is the first investigation regarding Vibrio distribution patterns and assembly mechanism in relation to coral species and density. Our study provides a foundation for study of the distribution dynamics of Vibrio associated with coral, and provides data support for coral conservation and restoration.
High Vibrio abundance in the coral rich areas may indicate their strong interaction with corals
Microbes in coral surrounding seawater are significantly different from those in ordinary seawater or sediments [17]. In our study, the average Vibrio abundance in coral reef areas was 1.57 × 103 copies/mL (Additional file 1: Table S4), higher than the value (1.17 × 103 copies/mL) in the South China Sea [55]. These abundance heterogeneities may be attributed to the presence of corals, in which high coral productivity [5] may provide nutrients for the growth of Vibrio. Moreover, the abundance of Vibrio in the Xisha Islands was higher than that in the Daya Bay (Fig. 2a, b). Differences in the number of corals may also affect Vibrio abundance. The number of corals in the Xisha Islands was nearly six times higher than that in the Daya Bay (Additional file 1: Fig. S2). Many interaction processes between microbes and corals have been reported. Corals can release dissolved and particulate organic matter into surrounding seawater in the form of coral mucus, which could aid heterotrophic feeding [13, 56], resulting higher bacterial abundance in coral mucus than in the surrounding seawater [57, 58]. Vibrio populations are able to rapidly respond to nutrient enrichment and use various quorum sensing (QS) signal molecules to regulate the process to colonize coral under different environmental conditions [59]. We also found positive correlation between the abundance of Vibrio and several corals in the Xisha Islands, while no correlation was found in the Daya Bay (Additional file 1: Fig. S3). Pollock et al. provided evidence of coral-microbe phylosymbiosis, in which the phylogeny of coral was related to the composition and richness of the coral microbiome [60]. Therefore, a higher Vibrio abundance may be required to maintain interactions with abundant corals in the Xisha Islands. In addition, coral bleaching occurred in the Xisha Islands in 2020 may be another reason for the high Vibrio abundance [32]. Bleached corals release twice as much particulate organic carbon (POC) and particulate nitrogen (PN) than normal corals [61], and coral pathogenic microbes may respond rapidly to these nutrient pulses [62, 63]. It has been reported that vibrios usually can respond to fluctuations in nutrient concentrations and show a tendency of explosive reproduction [21]. Indeed, during the 2015 to 2016 global coral bleaching event, the abundance of potential coral pathogens (such as Vibrio spp.) in the southern South China Sea increased from about 0.28 to ~ 52.92% [64].
Temperature and TN are also found to affect Vibrio abundance. The importance of temperature in regulating Vibrio abundance has been widely reported [44, 45]. For TN, a higher concentration was found in Xisha Islands than in Daya Bay (Additional file 1: Fig. S1). In general, the nitrogen cycle is of great importance to coral environments. Coral symbiotic algae can provide photosynthates to the coral host, but these photosynthates have been referred to as "junk food" due to a high C:N ratio and additional nitrogen is required to sustain coral growth [65]. The high efficiency of nitrogen absorption by coral holobiont enables this symbiotic relationship to effectively utilize nitrogen compounds in the surrounding seawater [10]. In addition, strains of Vibrio spp. and Photobacterium spp. showed the ability of nitrogen fixation [66,67,68]. A study have found that Vibrio spp. and Photobacterium spp. may be involved in the nitrogen-fixing process in the sediments of the South China Sea, and Vibrio spp. may also be involved in the dissimilatory nitrate reduction to ammonium [69]. Thus, Vibrio may co-exist with coral by participating in the nitrogen cycle process. We also found that the abundance of PA Vibrio in the Xisha Islands was higher than that in Daya Bay (Fig. 2a), which may be related to the particulate matter or attachment environment provided by corals [56].
Corals may weaken the species specificity of Vibrio spp. in the Chinese coral reef areas
Microbes may have strong adaptabilities facilitating their wide spread across various marine environments [70]. In the non-coral areas, including the South China Sea, the Chinese northern marginal seas and the Changjiang estuary, 160, 74 and 40 Vibrio OTUs were reported, respectively [38, 45, 55]. Comparatively, in this study, we obtained a total of 549 OTUs, suggesting that the presence of coral significantly increase the diversity of Vibrio spp. due to weak species-specific stimulation. Indeed, the abundant group in the coral-poor area, including V. campbellii, Paraphotobacterium marinum and V. caribbeanicus, were also dominant in the coral-rich area, corresponding their high niche widths. Predominance of these species in both areas may indicate their strong environmental adaptablities. V. campbellii is widely distributed, even in the oligotrophic deep sea, and can acquire gene horizontally to enhance adaptation of environment [70]. As a common pathogen of shrimp [71], V. campbellii encodes a sugar-specific porin in the outer membrane responsible for the transport of chitooligosaccharides, allowing it to grow rapidly in aquatic environments with chitin as a nutrient [72]. The environmental adaptability of V. campbellii was also reflected by its strong ability to degrade macromolecule substances [70], possession of quorum sensing for interbacterial cell killing and biofilms formation [73], and expression of rpoS ortholog gene (rpoS1) and rpoS xenolog gene (rpoS2) [74]. Paraphotobacterium marinum possesses the activity of a variety of enzymes including alkaline phosphatase, esterase (C4), leucine arylamidase and acid phosphatase, which can utilize D-Glucose and D-maltose, and reduce nitrate to nitrite [75]. Similarly, V. caribbeanicus also has the activity of various enzymes such as alkaline phosphatase and esterase lipase (C8), and can use a variety of amino acids and sugars [76]. The multiple enzyme activities and strengthening substrates utilization capabilities of Paraphotobacterium marinum and V. caribbeanicus may favor their growth in two areas.
It has been reported that the community compositions of Vibrio usually separated in various marine environments [21, 77]. In this study, except V. campbellii, Paraphotobacterium marinum and V. caribbeanicus, the rest species (e.g., rare taxa) showed differences in their relative abundances between coral rich and poor areas (Fig. 3), which may be related to habitat differentiation. We obtained 40 indicator species as marker species that can distinguish the two coral environments using random-forest machine-learning model (Fig. 3c). Among the indicator species in the Daya Bay, temperature may affect P. aquimaris OTU37 and OTU530. The growth temperature of P. aquimaris is 10–25 ℃, and cannot grow at 30 and 37 ℃ [78], whereas the temperature of the Xisha Islands is more than 30 ℃. Additionally, P. leiognathi was isolated from the luminescent organs of fishes in family Leiognathidae [79]. Fishes of family Leiognathidae are common fish resources in the southern coast of China, and studies have found that eggs of family Leiognathidae account for the largest proportion of all fish eggs in the Daya Bay [80, 81]. Thus, the special fish resources and temperature may explain why most of the indicator species in the Daya Bay are of the Photobacterium spp. Coral pathogenic Vibrio species, e.g., V. owensii, V. rotiferianus and V. alginolyticus, were among the indicated species of the Xisha Islands. V. owensii, V. parahaemolyticus and V. rotiferianus are the potential coral-pathogens that cause diseases of corals [7, 24]. V. alginolyticus and P. rosenbergii are detected from unhealthy corals at Tioman Island Marine Park, Malaysia [82]. The relative abundances of these coral pathogenic vibrios in the Xisha Islands are higher than those in the Daya Bay (Fig. 3d). The 2020 coral bleaching event in the Xisha Islands may cause the increasements of coral pathogenic vibrios [63]. With high coral number, Vibrio in the Xisha Islands may be developed as the dominant group by influencing coral health. In sum, the distribution patterns of Vibrio spp. reflected differences between coral poor and rich areas.
Distinct community structure of Vibrio spp. may be affected by biological factors and assembly mechanism
Previous reports on community structure of Vibrio mainly focused on the influence of physicochemical parameters, showing that temperature and salinity are important factors [38, 45]. However, the interaction between Vibrio and corals may play an important role in coral reef areas. In the Daya Bay, the main Vibrio species showed a positive correlation with corals (Fig. 5a), which may be due to the fact that corals provide nutrients for their growth [56]. The community structure of Vibrio in the Xisha Islands was greatly affected by corals (Fig. 4c), and the main groups showed negatively correlation with coral (Fig. 5b). The Xisha Islands face the problem of coral bleaching, and there is a close relationship between coral bleaching and Vibrio, perhaps causing a negative correlation of them [32, 63]. Meanwhile, vibrios in Xisha Islands were driven by ecological drift, whereas that in Daya Bay was mainly influenced by ecological drift and homogenizing selection (Fig. 6c), which might be related to the habitat filtration. Habitat filtering across the globe has been reported to cause the various structure of marine bacterial communities [83]. Ecological drift, used to describe random fluctuations in species abundance over time, is a process that can lead to different microbial communities between host species as these communities apart from each other over evolutionary time [84]. For instance, ecological drift drives interindividual diversification of gut microbes in early infants, causing an increase in β-diversity of microbial community [85]. Meanwhile, microbes in coral ecosystems are diverse, and even within the same coral individual, the microbial communities are different [86]. High β-diversity of Vibrio communities was also discovered in the Xisha Islands (Additional file 1: Fig. S4). Thus, the large number of different corals species in the Xisha Islands may have influenced the surrounding seawater to produce a distinct Vibrio community composition, making ecological drift a major community assembly process. Homogenizing selection refers to the selective effect of similar environment on the formation of microbial community, and leads to the convergence of microbes [87]. Lower β-diversity of Vibrio in the Daya Bay (Additional file 1: Fig. S4) indicated that Vibrio community tended to be more consistent than that in Xisha Islands. Furthermore, it has been found that more similar environmental conditions in smaller spatial ranges can promote the effect of homogenizing selection [88]. Thus, the less number and species of corals and the relatively small spatial range in the Daya Bay may provide a relatively single living environment for Vibrio spp., which may weaken the ecological drift and enhance the homogenizing selection.
Conclusion
We studied the distribution pattern and assembly mechanism of Vibrio spp. in both coral rich (Xisha Islands) and poor areas (Daya Bay) by molecular approaches. The abundance of Vibrio in the Xisha Islands was significantly higher than that in Daya Bay, which may be related to the interaction with corals. Community composition analysis showed that coral might stimulate the growth of the whole Vibrio genus resulting in less species specificity, in which V. campbellii, Paraphotobacterium marinum and V. caribbeanicus with strong environmental adaptability were dominant in both two areas. However, there were still heterogeneity of these two areas as assessed through random forest. The indicator species of coral poor areas were mainly Photobacterium spp., whereas those in coral rich areas were dominated by Vibrio spp. The assembly mechanisms of Vibrio communities appeared dissimilarly between the two regions. In the coral poor areas, homogenizing selection and ecological drift dominated the assembly of Vibrio communities, whereas in the coral rich areas, only ecological drift dominated the assembly of Vibrio communities. Different number and species of corals, environmental (e.g., temperature) and spatial factors between these two coral reef environments may be the primary causes. However, our study only assessed the dynamics of Vibrio in the coral environments by molecular approaches, and the possible colonization status of Vibrio in corals should be further evaluated experimentally to enrich the background data of coral reef protection.
Availability of data and materials
All sequencing data were stored in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA), with the accession numberSRP420698 and bioproject number PRJNA930676.
References
Hoegh-Guldberg O, Poloczanska ES, Skirving W, Dove S. Coral reef ecosystems under climate change and ocean acidification. Front Mar Sci. 2017;4:158.
Nelson HR, Kuempel CD, Altieri AH. The resilience of reef invertebrate biodiversity to coral mortality. Ecosphere. 2016;7(7): e01399.
MacNeil MA, Graham NAJ, Cinner JE, Wilson SK, Williams ID, Maina J, Newman S, Friedlander AM, Jupiter S, Polunin NVC, McClanahan TR. Recovery potential of the world’s coral reef fishes. Nature. 2015;520:341–4.
Stat M, Morris E, Gates RD. Functional diversity in coral–dinoflagellate symbiosis. Proc Natl Acad Sci USA. 2008;105(27):9256–61.
Hoppe H-G, Schramm W, Bacolod P. Spatial and temporal distribution of pelagic microorganisms and their proteolytic activity over a partly destroyed coral reef. Mar Ecol Prog Ser. 1988;44(1):95–102.
Kelly LW, Williams GJ, Barott KL, Carlson CA, Dinsdale EA, Edwards RA, Haas AF, Haynes M, Lim YW, McDole T, Nelson CE, Sala E, Sandin SA, Smith JE, Vermeij MJA, Youle M, Rohwer F. Local genomic adaptation of coral reef-associated microbiomes to gradients of natural variability and anthropogenic stressors. Proc Natl Acad Sci USA. 2014;111(28):10227–32.
Kemp KM, Westrich JR, Alabady MS, Edwards ML, Lipp EK. Abundance and multilocus sequence analysis of Vibrio associated with diseased elkhorn coral, Acropora palmata, of the Florida Keys. Appl Environ Microb. 2017;84(2):AEM.01035-17.
Galand PE, Ruscheweyh H-J, Salazar G, Hochart C, Henry N, Hume BCC, et al. Diversity of the Pacific Ocean coral reef microbiome. Nat Commun. 2023;14:3039.
Shnit-Orland M, Kushmaro A. Coral mucus-associated bacteria: a possible first line of defense. FEMS Microbiol Ecol. 2009;67(3):371–80.
Rädecker N, Pogoreutz C, Voolstra CR, Wiedenmann J, Wild C. Nitrogen cycling in corals: the key to understanding holobiont functioning? Trends Microbiol. 2015;23(8):490–7.
Sun W, Anbuchezhian R, Li Z. Association of coral-microbes, and the ecological roles of microbial symbionts in corals. In: Goffredo S, Dubinsky Z, editors. The cnidaria, past, present and future. Cham: Springer; 2016. https://doi.org/10.1007/978-3-319-31305-4_22.
Taniguchi A, Yoshida T, Eguchi M. Bacterial production is enhanced by coral mucus in reef systems. J Exp Mar Biol Ecol. 2014;461:331–6.
Tanaka Y, Ogawa H, Miyajima T. Bacterial decomposition of coral mucus as evaluated by long-term and quantitative observation. Coral Reefs. 2011;30:443–9.
Nakajima R, Yoshida T, Azman BAR, Zaleha K, Othman BHR, Toda T. In situ release of coral mucus by Acropora and its influence on the heterotrophic bacteria. Aquat Ecol. 2009;43:815–23.
Nelson CE, Goldberg SJ, Wegley Kelly L, Haas AF, Smith JE, Rohwer F, Carlson CA. Coral and macroalgal exudates vary in neutral sugar composition and differentially enrich reef bacterioplankton lineages. ISME J. 2013;7:962–79.
Takemura A, Chien D, Polz M. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front Microbiol. 2014;5:38.
Tout J, Jeffries TC, Webster NS, Stocker R, Ralph PJ, Seymour JR. Variability in microbial community composition and function between different niches within a coral reef. Microb Ecol. 2014;67:540–52.
Weber L, Apprill A. Diel, daily, and spatial variation of coral reef seawater microbial communities. PLoS ONE. 2020;15(3): e0229442.
Xu W, Gong L, Yang S, Gao Y, Ma X, Xu L, Chen H, Luo Z. Spatiotemporal dynamics of Vibrio communities and abundance in Dongshan Bay, South of China. Front Microbiol. 2020;11: 575287.
Thompson JR, Randa MA, Marcelino LA, Tomita-Mitchell A, Lim E, Polz MF. Diversity and dynamics of a north Atlantic coastal Vibrio community. Appl Environ Microb. 2004;70(7):4103–10.
Zhang X-H, Lin H, Wang X, Austin B. Significance of Vibrio species in the marine organic carbon cycle. A review. Sci China Earth Sci. 2018;61:1357–68.
Fu J, Li Y, Zhao L, Wu C, He Z. Characterization and genomic analysis of a bacteriophage with potential in lysing Vibrio alginolyticus. Viruses. 2023;15(1):135.
Pfeifer E, Michniewski S, Gätgens C, Münch E, Müller F, Polen T, Millard A, Blombach B, Frunzke J. Generation of a prophage-free variant of the fast-growing bacterium Vibrio natriegens. Appl Environ Microb. 2019;85:e00853-e919.
Amin AKMR, Feng G, Al-saari N, Meirelles PM, Yamazaki Y, Mino S, Thompson FL, Sawabe T, Sawabe T. The first temporal and spatial assessment of Vibrio diversity of the surrounding seawater of coral reefs in Ishigaki, Japan. Front Microbiol. 2016;7:1185.
Couce E, Ridgwell A, Hendy EJ. Environmental controls on the global distribution of shallow-water coral reefs. J Biogeogr. 2012;39:1508–23.
Weber L, Gonzalez-Díaz P, Armenteros M, Apprill A. The coral ecosphere: a unique coral reef habitat that fosters coral–microbial interactions. Limnol Oceanogr. 2019;64:2373–88.
Fu X, Wang C, Shao C, Han L, Li G, Zeng X, Guan H. Investigation on the status of coral reef resources and medicinal research in China I. Coral reef resources and ecological functions. J Ocean U China. 2009;39:676–84.
Yu K. Coral reefs in the South China Sea: their response to and records on past environmental changes. Sci China Earth Sci. 2012;55:1217–29.
Huang H, You F, Lian J, Yang J, Li X, Dong Z, Zhang C, Yuan T. Species diversity and distribution of scleractinian coral at Xisha Islands, China. Biodivers Sci. 2011;19:710–5.
Zhao M, Yu K, Shi Q, Chen T, Zhang H, Chen T. Coral communities of the remote atoll reefs in the Nansha Islands, southern South China Sea. Environ Monit Assess. 2013;185:7381–92.
Zuo X, Su F, Zhao H, Zhang J, Wang Q, Wu D. Regional hard coral distribution within geomorphic and reef flat ecological zones determined by satellite imagery of the Xisha Islands, South China Sea. Chin J Ocean Limnol. 2017;35:501–14.
Xiao J, Wang W, Wang X, Tian P, Niu W. Recent deterioration of coral reefs in the South China Sea due to multiple disturbances. PeerJ. 2022;10: e13634.
Zhao J, Qiu Y, Zhang H, Cheng T. Distribution and ecological characters of neritic stone corals at Daya Bay. J Trop Oceanogr. 2007;26:63–7.
Li S, Yu K, Shi Q, Chen T, Zhao M, Zhao J. Interspecies and spatial diversity in the symbiotic zooxanthellae density in corals from northern South China Sea and its relationship to coral reef bleaching. Chin Sci Bull. 2008;53:295–303.
Guo F, Xiao J, Tian P, Wang W, Wang X, Huang D, Wang J, Niu W. Status of scleractinian corals in Daya Bay and along the coast of Dapeng Peninsula and ecological vulnerability assessment. J Appl Oceanogr. 2022;41:568–82.
Shi R, Xu S, Qi Z, Zhu Q, Huang H, Weber F. Influence of suspended mariculture on vertical distribution profiles of bacteria in sediment from Daya Bay, Southern China. Mar Pollut Bull. 2019;146:816–26.
Zhang X, Zhang Z, Li J, Lu Y, Zheng P, Xian J. Isolation and identification of marine Vibrio from Xisha Yongle Islands, China. J Fish Res. 2022;44(1):1–8.
Wang X, Liu J, Liang J, Sun H, Zhang X-H. Spatiotemporal dynamics of the total and active Vibrio spp. populations throughout the Changjiang estuary in China. Environ Microbiol. 2020;22:4438–55.
Bohlen L, Dale AW, Sommer S, Mosch T, Hensen C, Noffke A, Scholz F, Wallmann K. Benthic nitrogen cycling traversing the Peruvian oxygen minimum zone. Geochim Cosmochim Acta. 2011;75:6094–111.
Wang J, Yao P, Bianchi TS, Li D, Zhao B, Cui X, Pan H, Zhang T, Yu Z. The effect of particle density on the sources, distribution, and degradation of sedimentary organic carbon in the Changjiang Estuary and adjacent shelf. Chem Geol. 2015;402:52–67.
Zhao B, Yao P, Li D, Yu Z. Effects of river damming and delta erosion on organic carbon burial in the Changjiang Estuary and adjacent East China Sea inner shelf. Sci Total Environ. 2021;793: 148610.
Church C, Spargo J, Fishel S. Strong acid extraction methods for “total phosphorus” in soils: EPA method 3050B and EPA method 3051. Agric Environ Lett. 2017;2: 160037.
Varol M, Sen B. Assessment of nutrient and heavy metal contamination in surface water and sediments of the upper Tigris River, Turkey. CATENA. 2012;92:1–10.
Vezzulli L, Brettar I, Pezzati E, Reid PC, Colwell RR, Höfle MG, Pruzzo C. Long-term effects of ocean warming on the prokaryotic community: evidence from the vibrios. ISME J. 2012;6:21–30.
Liang J, Liu J, Wang X, Lin H, Liu J, Zhou S, Sun H, Zhang X-H. Spatiotemporal dynamics of free-living and particle-associated vibrio communities in the Northern Chinese Marginal Seas. Appl Environ Microb. 2019;85(9):e00217.
Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(17):884–90.
Magoč T, Salzberg SL. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics. 2011;27(21):2957–63.
Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. 2016;13:581.
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–6.
ter Braak CJF, Smilauer P. Canoco reference manual and user's guide: software for ordination, version 5.0. Microcomputer Power. 2012.
Stegen JC, Lin X, Konopka AE, Fredrickson JK. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012;6:1653–64.
Kooperman N, Ben-Dov E, Kramarsky-Winter E, Barak Z, Kushmaro A. Coral mucus-associated bacterial communities from natural and aquarium environments. FEMS Microbiol Lett. 2007;276(1):106–13.
Schöttner S, Hoffmann F, Wild C, Rapp HT, Boetius A, Ramette A. Inter- and intra-habitat bacterial diversity associated with cold-water corals. ISME J. 2009;3:756–9.
Li N, Dong K, Jiang G, Tang J, Xu Q, Li X, Kang Z, Zou S, Chen X, Adams JM, Zhao H. Stochastic processes dominate marine free-living Vibrio community assembly in a subtropical gulf. FEMS Microbiol Ecol. 2020;96(11):fiaa198.
Wang X, Liu J, Zhao W, Liu J, Liang J, Thompson F, Zhang X-H. Fine-scale structuring of planktonic Vibrio spp. in the Chinese marginal seas. Appl Environ Microb. 2022;88(23):e01262.
Brown BE, Bythell JC. Perspectives on mucus secretion in reef corals. Mar Ecol Prog Ser. 2005;296:291–309.
Garren M, Azam F. New method for counting bacteria associated with coral mucus. Appl Environ Microb. 2010;76(18):6128–33.
Allers E, Niesner C, Wild C, Pernthaler J. Microbes enriched in seawater after addition of coral mucus. Appl Environ Microb. 2008;74(10):3274–8.
Tait K, Hutchison Z, Thompson FL, Munn CB. Quorum sensing signal production and inhibition by coral-associated vibrios. Environ Microbiol Rep. 2010;2:145–50.
Pollock FJ, McMinds R, Smith S, Bourne DG, Willis BL, Medina M, Thurber RV, Zaneveld JR. Coral-associated bacteria demonstrate phylosymbiosis and cophylogeny. Nat Commun. 2018;9:4921.
Niggl W, Glas M, Laforsch C, Mayr C, Wild C. First evidence of coral bleaching stimulating organic matter release by reef corals. In: Proceedings of the 11th international coral reef symposium, Ft. Lauderdale, Florida, 7–11 July 2008, Session 19. 2009.
Radice VZ, Fry B, Dove SG, Hoegh-Guldberg O. Biogeochemical variability and trophic status of reef water column following a coral bleaching event. Coral Reefs. 2021;40(1):1–7.
Zhao W, Chen L, Liu M, Huang K, Ding Y, Xiao J, Tian P, Liu J, Zhang X-H, Niu W, Wang X. Sedimentary Vibrio blooms in the Xisha Islands may associate with the 2020 coral bleaching event. Appl Environ Microb. 2023;89(7):e00543.
Qin Z, Yu K, Liang J, Yao Q, Chen B. Significant changes in microbial communities associated with reef corals in the southern south china sea during the 2015/2016 global-scale coral bleaching event. J Geophys Res Oceans. 2020;125:e2019JC015579.
Falkowski PG, Dubinsky Z, Muscatine L, Porter JW. Light and the bioenergetics of a symbiotic coral. Bioscience. 1984;34(11):705–9.
Kang SR, Srinivasan S, Lee S-S. Vibrio oceanisediminis sp. nov., a nitrogen-fixing bacterium isolated from an artificial oil-spill marine sediment. Int J Syst Evol Microbiol. 2015;65(10):3552–7.
Rowan-Nash AD, Korry BJ, Mylonakis E, Belenky P. Cross-domain and viral interactions in the microbiome. Microbiol Mol Biol R. 2019;83(1):e00044-e118.
Shieh WY, Simidu U, Maruyama Y. A photobacterium-like bacterium able to fix nitrogen. Antonie Van Leeuwenhoek. 1990;57:51–4.
Yu T, Li M, Niu M, Fan X, Liang W, Wang F. Difference of nitrogen-cycling microbes between shallow bay and deep-sea sediments in the South China Sea. Appl Microbiol Biotechnol. 2018;102:447–59.
Liang J, Liu J, Wang X, Sun H, Zhang Y, Ju F, Thompson F, Zhang X-H. Genomic analysis reveals adaptation of Vibrio campbellii to the Hadal Ocean. Appl Environ Microb. 2022;88(16):e00575.
Wang L, Chen Y, Huang H, Huang Z, Chen H, Shao Z. Isolation and identification of Vibrio campbellii as a bacterial pathogen for luminous vibriosis of Litopenaeus vannamei. Aquac Res. 2015;46:395–404.
Aunkham A, Suginta W. Probing the physiological roles of the extracellular loops of chitoporin from Vibrio campbellii. Biophys J. 2021;120(11):2124–37.
Simpson CA, Petersen BD, Haas NW, Geyman LJ, Lee AH, Podicheti R, Pepin R, Brown LC, Rusch DB, Manzella MP, Papenfort K, van Kessel JC. The quorum-sensing systems of Vibrio campbellii DS40M4 and BB120 are genetically and functionally distinct. Environ Microbiol. 2021;23:5412–32.
Barekzi N, Wang Z, Rubin RA, Rahbar AM, Thach DC, Meador CE, Vora GJ. Transcriptomic, proteomic and phenotypic analysis of the Vibrio campbellii stationary phase alternative sigma factors RpoS1 and RpoS2.
Huang Z, Dong C, Shao Z. Paraphotobacterium marinum gen. nov., sp. nov., a member of the family Vibrionaceae, isolated from surface seawater. Int J Syst Evol Microbiol. 2016;66(8):3050–6.
Hoffmann M, Monday SR, Allard MW, Strain EA, Whittaker P, Naum M, McCarthy PJ, Lopez JV, Fischer M, Brown EW. Vibrio caribbeanicus sp. nov., isolated from the marine sponge Scleritoderma cyanea. Int J Syst Evol Microbiol. 2012;62(8):1736–43.
Chen X, Zhao H, Jiang G, Tang J, Xu Q, Huang L, Chen S, Zou S, Dong K, Li N. Responses of free-living vibrio community to seasonal environmental variation in a subtropical Inland Bay. Front Microbiol. 2020;11: 610974.
Yoshizawa S, Wada M, Kita-Tsukamoto K, Yokota A, Kogure K. Photobacterium aquimaris sp. nov., a luminous marine bacterium isolated from seawater. Int J Syst Evol Microbiol. 2009;59(6):1438–42.
Dunlap PV. Osmotic control of luminescence and growth in Photobacterium leiognathi from ponyfish light organs. Arch Microbiol. 1985;141:44–50.
Liu J, Zhuang H, Liu Y, Pharmacy SO. Detection of muscular fatty acids in 8 ponyfish species in Zhanjiang ocean. J Guangdong Med Coll. 2014;32:773–80.
Lin Z, Wang X, Jiang Y. Distribution and species composition of fish eggs in Daya Bay. J Fish Sci China. 2010;17:543–50.
Khodzori FA, Saad S, Mansor NN, Nasir NANM, Khalid NNNA, Rawi FZ. Pathogenic Vibrio spp. identified for white syndrome coral disease in Tioman Island Marine Park, Malaysia. Malays J Microbiol. 2021;17(1):69–79.
Pontarp M, Canbäck B, Tunlid A, Lundberg P. Phylogenetic analysis suggests that habitat filtering is structuring marine bacterial communities across the globe. Microb Ecol. 2012;64:8–17.
Kohl KD. Ecological and evolutionary mechanisms underlying patterns of phylosymbiosis in host-associated microbial communities. Philos Trans R Soc B. 2020;375:20190251.
Seki D, Schauberger C, Hausmann B, Berger A, Wisgrill L, Berry D. Individuality of the extremely premature infant gut microbiota is driven by ecological drift. mSystems. 2022;7(3):e00163-e222.
Zhang J, Hu A, Sun Y, Yang Q, Dong J, Long L, Huang S. Dispersal limitation expands the diversity of coral microbiome metacommunity in the South China Sea. Front Mar Sci. 2021;8: 658708.
Stegen JC, Lin X, Fredrickson JK, Konopka AE. Estimating and mapping ecological processes influencing microbial community assembly. Front Microbiol. 2015;6:370.
Logares R, Tesson SVM, Canbäck B, Pontarp M, Hedlund K, Rengefors K. Contrasting prevalence of selection and drift in the community structuring of bacteria and microbial eukaryotes. Environ Microbiol. 2018;20:2231–40.
Acknowledgements
We appreciate all the crew members and the divers for their great efforts and help in sample collection on the R/V Yuezhanyuke 10 during the expedition.
Funding
This work was funded by the National Natural Science Foundation of China (No. 42006085); the Fundamental Research Funds for the Central Universities (202312034); the Marine S&T Fund of Fujian Province (2021J05094); the Scientific Research Foundation of the Third Institute of Oceanography, MNR (No. 2020005) and the Fund of Key Laboratory of Marine Ecological Conservation and Restoration, Ministry of Natural Resources/ Fujian Provincial Key Laboratory of Marine Ecological Conservation and Restoration (EPR2023001).
Author information
Authors and Affiliations
Contributions
WX and LJ developed the study concept and design; WX, TP and NW collected the samples; ZW completed the DNA isolations; ZW processed the raw sequencing data; ZW and CX performed the data analysis; ZW wrote the original manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplementary Material Figs. S1-S6, Tables S1-S5.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhao, W., Chen, X., Liu, R. et al. Distinct coral environments shape the dynamic of planktonic Vibrio spp.. Environmental Microbiome 18, 77 (2023). https://doi.org/10.1186/s40793-023-00532-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40793-023-00532-7