
Abstract
After pubertal, cohort of small antral follicles enters to gonadotrophin-sensitive development, called recruited follicles. This study was aimed to identify candidate genes in follicular cyclic recruitment via analysis of protein-protein interaction (PPI) network. Differentially expressed genes (DEGs) in ovine granulosa cells of small antral follicles between follicular and luteal phases were accumulated among gene/protein symbols of the Ensembl annotation. Following directed graphs, PTPN6 and FYN have the highest indegree and outdegree, respectively. Since, these hubs being up-regulated in ovine granulosa cells of small antral follicles during the follicular phase, it represents an accumulation of blood immune cells in follicular phase in comparison with luteal phase. By contrast, the up-regulated hubs in the luteal phase including CDK1, INSRR and TOP2A which stimulated DNA replication and proliferation of granulosa cells, they known as candidate genes of the cyclic recruitment.
Similar content being viewed by others


Introduction
Small antral follicles represent the fundamental developmental unit of the mammalian ovary and, as such, serve the needs of the entire reproductive life span. After pubertal, cohort of small antral follicles enters to gonadotrophin-sensitive development upon FSH (Follicle Stimulating Hormone) stimulation, called cyclic recruited follicles [1, 2]. Cyclic recruitment, although obligatory, does not guarantee ovulation because growing follicles are vulnerable to atresia and thus may fall out from the growth trajectory [3]. Among somatic cells of the antral follicles, granulosa cells will undergo serious changes morphologically and physiologically during the processes of proliferation, differentiation, ovulation, lutenization and atresia [4].
To understand gene function of the cellular processes, genes must be studied in the context of networks. Emerging tools from systems biology going beyond simple gene ontology (GO) terms like causal network modelling linking gene expression analysis to gene interaction information are sorely needed [5, 6]. Nowadays, biological networks allow a comprehensive examination of the complex mechanisms for targeted empirical studies [7]. Numerous studies have been conducted in attempt to identify candidate genes of reproductive biology via gene networks analysis such as development of rat primordial follicles [8, 9], early embryo development in mouse [10] and bovine [11], gene networks of bovine oocytes [12] and bovine granulosa cells of small and large follicles [13]. Protein-protein interaction (PPI) networks are one the most known approaches for representation of candidate genes/proteins beyond of high-throughput studies [13,14,15,16,17,18,19].
In previous study, we surprisingly have shown differences in transcriptome profiles of ovine (sheep) granulosa cells between small antral follicles (1–3 mm) collected during the follicular and luteal phases [20]. Therefore, the specific purpose of the present study was to survey the differentially expressed genes (DEGs) from the previous study using the analysis of PPI networks in order to identification of candidate genes that probably those are impressive in cyclic recruitment of ovarian follicles.
Materials and methods
Dataset assessment
The main dataset of the present study was belonged to 663 DEGs in ovine granulosa cells between small antral follicles (1–3 mm in diameter) collected during the follicular and luteal phases [20]. These DEGs afterwards were annotated using a standard Ensembl gene annotation system.
Construction of protein-protein interaction (PPI) network
A large PPI network was reconstructed from the 646 genes/protein symbols (Additional file 1) of the Ensembl annotation (Ovis_aries.Oar_v3.1.91). In order to map pairwise interactions, all computational methods in STRING database Version. 10.0 [21] containing neighborhoods, gene fusion, co-occurrence, homology, co-expression, experiments, databases and text mining, were utilized with the medium confidence score (> 0.4).
Network analysis and modules selection
Cytoscape Version 3.1.1 [22] was used to plot and analyze the centralities, clustering and modularity of the PPI network. The MCODE (Molecular Complex Detection) v1.4.0-beta2 was performed to screen modules of PPI network with degree cutoff = 2, node score cutoff = 0.2, k-core = 2, and max. Depth = 100 [14]. This is a well-known automated algorithm to find highly interconnected subgraphs that detects densely connected regions in large PPI networks that may represent molecular complexes [23]. Also, the functional modules were chosen with the number of nodes ≥16 and nodes score [19].
Enrichment analysis for the functional modules
Biological significance of these predicted modules were inferred by ClueGO [24] plugin of Cytoscape. The statistical significance of the biological terms analyzed was calculated with Right-sided enrichment hypergeometric test and Benjamini and Hochberg P-value correction [25] to reduce false positives- and negatives. Kappa statistics were used to link and grouping of the enriched terms and functional grouping of them as described by Bindea et al. [24]. The minimum connectivity of the pathway network (kappa score) was set to 0.4 units.
Identification of hub genes/proteins and its directed interaction network
In this study, all networks were utilized to identify hub genes/proteins which important in folliculogenesis during the ovine estrous cycle. Hubs were detected by calculating the node degree distribution [16] using the Network Analyzer (http://apps.cytoscape.org/apps/networkanalyzer) plugin of Cytoscape [24]. Degree gives a simple count of the number of interactions of a given node [26]. Additionally, we utilized several centrality parameters including stress, betweenness and closeness instead of using degree centrality itself. The mathematical formulas for the calculation of centrality parameters including stress, betweenes and clossness, are available in Additional file 2 that has been retrieved from Zhuang et al. [26]. By extract direction of PPI from STRING Version. 10.0 [21], a directed gene network was reconstructed from the hub genes/proteins. These important proteins were extracted from hub genes/proteins via network analysis and modularity of the network. This small, but important, graph is visualized by Cytoscape Version 3.1.1 [22]. Using CluePedia v.1.1.7. [24] plugin of Cytoscape, hub genes/proteins were placed in a directed gene network based on its molecular function.
Result and discussion
Among 646 DEGs (Additional file 1), 498 proteins were annotated on the Total PPI network. Also, 2191 edges containing neighborhoods, gene fusion, co-occurrence, homology, co-expression, experiments, databases and text mining, were interacted between such genes/proteins (Fig. 1 and Additional file 3). The statistics of network containing network density, network diameter and clustering coefficient were 0.018, 9 and 0.275, respectively. The power law of degree distribution was followed with an R2 = 0.895. Meanwhile, R2 is computed on logarithmized values. Some proteins in this network had high values in all degree, stress and betweenness centralities, such as FYN, CDK1, RAC1, ACTG2, FGR, APP, MMP2, SYK, CDH1, TGFB1, PTPN6, ITGB7, FOS, ACTN1, GNAI2, INSRR, BMP4, BMP2, LYN and HCK (Additional file 3). This list was utilized to identify some candidate genes among hubs as major regulators in cyclic recruitment of ovine small antral follicles.

The Un-directed PPI network for total genes/proteins differentially expressed (cut-off of P < 0.05 in corrected tests of Benjamini-Hochberg) in luteal vs. follicular phases of ovine estrous cycle. The PPI network illustrates degree (node size) and betweenness centrality (node color) and layout option of edge weighted spring embedded. Up- and down-regulated genes in luteal phase vs. follicular phase are shown with triangles and V shapes, respectively
From the total PPI network, thirteen modules were extracted, among which five subnetworks (modules of 1, 2, 4, 6 and 12) were detected with the intra-connection nodes ≥16 and node score > 2.0 (Table 1). A module/subnetwork is a group of closely related proteins that act in concert to perform specific biological functions through PPI network that occurs in time and space [15]. In the biological processes (BP), the highest significant terms for module 1, module 2, module 4, module 6 and module 12 were related to the regulation of B cell receptor signaling pathway, Fc receptor mediated stimulatory signaling pathway, positive regulation of reactive oxygen species metabolic process, innate immune response activating cell surface receptor signaling pathway and positive regulation of DNA replication, respectively (Table 2). As opposite performances among these modules, we have surprisingly identified four modules of 1, 2, 4 and 6, in relevant to immune system in comparison with module 12 in relevant to cell proliferation (Table 2).
Among identified hub genes/proteins in Table 3, twenty-five hub genes (FYN, RAC1, FGR, MMP2, SYK, LYN, TGFB1, PTPN6, FOS, CDH1, ITGB7, HCK, APP, ACTN1, GNAI2, BMP4, BMP2, PTPRC, NFKB1, VAV1, IL1B, COL1A1, TIMP1, PDGFRA, BTK), were up-regulated in ovine granulosa cells of small antral follicles during the follicular phase (up-regulated genes in Follicular has been shown in Additional file 1). Interestingly, these hubs were significantly connected to immune system and phagocytosis (Table 2). On the contrary, three (CDK1, INSRR and TOP2A) were up-regulated in ovine granulosa cells of small antral follicles during the luteal phase (up-regulated genes in Luteal has been shown in Additional file 1).
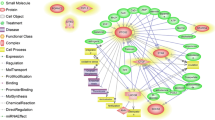
As shown in Fig. 2, the hub genes of PTPN6 and FYN are revealed the highest in-degree and out-degree, respectively (Additional file 4, and Fig. 2). Regarding to protein differential expression in normal tissues from HIPED (the Human Integrated Protein Expression Database), PTPN6 is overexpressed in Peripheral blood mononuclear cells, Lymph node, Blymphocyte, and Monocytes (http://www.genecards.org/). Moreover, FYN is another hub with highest out-degree, is also overexpressed in Peripheral blood mononuclear cells (http://www.genecards.org/). Regardless, PTPN6 and FYN being up-regulated in the ovine granulosa cells of small antral follicles during the follicular phase, represents an accumulation of blood immune cells into small antral follicles of the follicular phase in comparison with luteal phase. Therefore, the protein of FYN can be known as an upstream regulator in inhibition of ovarian folliculogenesis. This protein is belonged to the protein tyrosine kinase (PTK) family as illustrated in regulatory model of molecular function in Fig. 2. These signaling molecules regulate a variety of cellular processes including cell growth, differentiation, mitotic cycle, and oncogenic transformation (http://www.genecards.org/).

The directed PPI network for the identified hub genes/proteins. The nodes with small red circles and blue triangles are down- and up-regulated hub genes/proteins in luteal phase vs. follicular phase, respectively. The green- and red vectors are revealed the molecular function of activation and inhibition, respectively. The hub genes/proteins in direction with molecular function are shown with red labels
As shown in Table 2, the subnetwork 2 with the eight hubs were became the highest in relative to the others. The most significant term of BP in module 2 was belonged to Fc receptor mediated stimulatory signaling for the hub genes/proteins, including FYN, HCK, RAC1, SYK and VAV1 (Table 2). This can regulate immune responses through interacting with Fc receptors [27]. Moreover, the up-regulated hubs of PDGFRA, COL1A1, BMP2, CDH1, ACTN1, FOS, FYN, and TIMP1 in the follicular phase, they also were up-regulated in bovine and porcine granulosa cells of small atretic follicles [28, 29]. By contrast, the up-regulated hubs of CDK1 and TOP2A in the luteal phase, they also were up-regulated in the bovine granulosa cells of small heathy follicles [28]. These genes were cohesively interconnected around up-regulated nodes in classical mitosis checkpoint controllers [30]. Furthermore, TOP2A is expressed mainly during the S to G2 /M phase of the cell cycle and is likely to play a major role in DNA catenation during mitosis [31]. Interestingly, such hubs all were belonged to module 12 whose been enriched in positive regulation of DNA, cell cycle and progesterone-mediated oocyte (Table 2). Therefore, the hubs including CDK1, INSRR and TOP2A, represented the proliferation of ovine granulosa cells from small antral follicles during the luteal phase [20], whose probably are crucial in cyclic recruitment of small antral follicles. This is closer to the second theory in recruitment of antral follicles during menstrual cycle by Baerwald et al. [32].
Conclusion
In this study, we evidenced that cyclic recruitment of small antral follicles mostly occurs in the luteal phase in comparison with follicular phase during the ovine estrous cycle. Based on analysis of PPI network and its modulation, we identified some biomarkers whose potentially impress on cyclic recruitment of ovarian follicles. Surprisingly, FYN was identified as upstream regulator that probably inhibits the proliferation of granulosa cells. By contrast, hub genes of CDK1, INSRR and TOP2A, were known as inducers in proliferation of granulosa cells among genes were up-regulated in luteal phase in comparison with follicular phase. These results may provide valuable genetic markers for increasing ewe prolificacy with focus on cyclic recruitment of ovarian small follicles. Nevertheless, further studies using an experimental approach and a greater number of individuals are warranted for the verification of such candidate genes.
Abbreviations
- BP:
-
Biological process
- CDK1:
-
Cyclin-dependent kinase 1
- DEGs:
-
Differentially expressed genes
- FYN:
-
Proto-oncogene tyrosine-protein kinase
- GO:
-
Gene Oncology
- INSRR:
-
Insulin Receptor Related Receptor
- PPI:
-
Protein-Protein Interaction
- PTPN6:
-
Protein tyrosine phosphatase, non-receptor type 6
- TOP2A:
-
Topoisomerase II Alpha
References
McGee EA, Hsueh AJ. Initial and cyclic recruitment of ovarian follicles. Endocr Rev. 2000;21:200–14.
Shimizu T. Molecular and cellular mechanisms for the regulation of ovarian follicular function in cows. J Reprod Dev. 2016;62:323–9.
Strauss JF, Barbieri RL. Yen & Jaffe's reproductive endocrinology E-book: physiology, pathophysiology, and clinical management (Expert Consult - Online and Print). 7th Edition. Philadelphia: Elsevier Inc.; 2013.
Cuiling L, Wei Y, Zhaoyuan H, Yixun L. Granulosa cell proliferation differentiation and its role in follicular development. Chin Sci Bull. 2005;50:2665.
Beiki H, Nejati-Javaremi A, Pakdel A, Masoudi-Nejad A, Hu ZL, Reecy JM. Large-scale gene co-expression network as a source of functional annotation for cattle genes. BMC Genomics. 2016;17:846.
Chindelevitch L, Ziemek D, Enayetallah A, Randhawa R, Sidders B, Brockel C, et al. Causal reasoning on biological networks: interpreting transcriptional changes. Bioinformatics. 2012;28:1114–21.
Amiri M, Jafari M, AzimzadehJamalkandi S, Davoodi SM. Atopic dermatitis associated protein interaction network lead to new insights in chronic sulfur mustard skin lesion mechanisms. Expert Rev Proteomics. 2013;10:449–60.
Nilsson EE, Savenkova MI, Schindler R, Zhang B, Schadt EE, Skinner MK. Gene bionetwork analysis of ovarian primordial follicle development. PLoS One. 2010;5:e11637.
Nilsson E, Zhang B, Skinner MK. Gene bionetworks that regulate ovarian primordial follicle assembly. BMC Genomics. 2013;14:496.
Xue Z, Huang K, Cai C, Cai L, Jiang CY, Feng Y, et al. Genetic programs in human and mouse early embryos revealed by single-cell RNA sequencing. Nature. 2013;500:593–7.
Jiang Z, Sun J, Dong H, Luo O, Zheng X, Obergfell C, et al. Transcriptional profiles of bovine in vivo pre-implantation development. BMC Genomics. 2014;15:756.
Labrecque R, Fournier E, Sirard MA. Transcriptome analysis of bovine oocytes from distinct follicle sizes: insights from correlation network analysis. Mol Reprod Dev. 2016;83:558–69.
Bahrami A, Miraie-Ashtiani SR, Sadeghi M, Najafi A. miRNA-mRNA network involved in folliculogenesis interactome: systems biology approach. Reproduction. 2017;154:51–65.
Kotni MK, Zhao M, Wei DQ. Gene expression profiles and protein-protein interaction networks in amyotrophic lateral sclerosis patients with C9orf72 mutation. Orphanet J Rare Dis. 2016;11:148.
Lin CY, Lee TL, Chiu YY, Lin YW, Lo YS, Lin CT, et al. Module organization and variance in protein-protein interaction networks. Sci Rep. 2015;5:9386.
Malik A, Lee EJ, Jan AT, Ahmad S, Cho KH, Kim J, et al. Network analysis for the identification of differentially expressed hub genes using myogenin knock-down muscle satellite cells. PLoS One. 2015;10:e0133597.
Nai W, Threapleton D, Lu J, Zhang K, Wu H, Fu Y, et al. Identification of novel genes and pathways in carotid atheroma using integrated bioinformatic methods. Sci Rep. 2016;6:18764.
Talebi R, Ahmadi A, Afraz F, Abdoli R. Parkinson's disease and lactoferrin: analysis of dependent protein networks. Gene Rep. 2016;4:177–83.
Xu Z, Zhou Y, Cao Y, Dinh TL, Wan J, Zhao M. Identification of candidate biomarkers and analysis of prognostic values in ovarian cancer by integrated bioinformatics analysis. Med Oncol. 2016;33:130.
Talebi R, Ahmadi A, Afraz F, Sarry J, Plisson-Petit F, Genet C, Fabre S. Transcriptome analysis of ovine granulosa cells reveals differences between small antral follicles collected during the follicular and luteal phases. Theriogenology. 2018;108:103–17.
Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, et al. STRING v10: protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015;43:447–52.
Lotia S, Montojo J, Dong Y, Bader GD, Pico AR. Cytoscape app store. Bioinformatics. 2013;29:1350–1.
Bader GD, Hogue CW. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinformatics. 2003;4:2.
Bindea G, Mlecnik B, Hackl H, Charoentong P, Tosolini M, Kirilovsky A, et al. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics. 2009;15:1091–3.
Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995;57:289–300.
Zhuang DY, Jiang L, He QQ, Zhou P, Yue T. Identification of hub subnetwork based on topological features of genes in breast cancer. Int J Mol Med. 2015;35:664–74.
Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immune responses. Nat Rev Immunol. 2008;8:34–47.
Hatzirodos N, Hummitzsch K, Irving-Rodgers HF, Harland ML, Morris SE, Rodgers RJ. Transcriptome profiling of granulosa cells from bovine ovarian follicles during atresia. BMC Genomics. 2014;15:40.
Terenina E, Fabre S, Bonnet A, Monniaux D, Robert-Granié C, SanCristobal M, et al. Differentially expressed genes and gene networks involved in pig ovarian follicular atresia. Physiol Genomics. 2017;49:67–80.
Pérez-Guijarro E, Karras P, Cifdaloz M, Martínez-Herranz R, Cañón E, Graña O, et al. Lineage-specific roles of the cytoplasmic polyadenylation factor CPEB4 in the regulation of melanoma drivers. Nat Commun. 2016;7:13418.
Tamaichi H, Sato M, Porter AC, Shimizu T, Mizutani S, Takagi M. Ataxia telangiectasia mutated-dependent regulation of topoisomerase II alpha expression and sensitivity to topoisomerase II inhibitor. Cancer Sci. 2013;104:178–84.
Baerwald AR, Adams GP, Pierson RA. Ovarian antral folliculogenesis during the human menstrual cycle: a review. Hum Reprod Update. 2012;18:73–91.
Funding
This work was supported by “Bu-Ali Sina University” and “Agricultural Biotechnology Research Institute” from Iran.
Availability of data and materials
The part of data generated or analyzed during the current study are available in the supplemental files, and other parts from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
RT, AA and FA conceived the study. RT performed the bioinformatics analyses. RT, AA and FA wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All procedures were approved (approval number 01171.02) by the French Ministry of Teaching and Scientific Research and local ethical committee C2EA-15 (Animal Health and Science) in accordance with the European Union Directive 2010/63/EU on the protection of animals used for scientific purposes.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
The 646 genes of differentially expressed (DEGs) in luteal vs. follicular phase (P < 0.05 in corrected test of Benjamini-Hochberg) from the genes/protein symbols (Additional file 1) of the Ensembl annotation (Ovis_aries.Oar_v3.1.91). The 455 up-regulated genes in ovine granulosa cells of the small antral follicles in the follicular phase. The 191 up-regulated genes in ovine granulosa cells of the small antral follicles in the Luteal phase (Talebi et al. [20]). (XLS 108 kb)
Additional file 2:
The mathematical formulas for the analysis of topological parameters like network centrality options such as stress, betweenness and closeness centralities (Zhuang et al. [26]). (DOCX 16 kb)
Additional file 3:
The list of 498 proteins as nodes was annotated on the Total PPI network. Edges containing neighborhoods, gene fusion, co-occurrence, homology, coexpression, experiments, databases and text mining, were interacted between such genes/proteins (XLS 839 kb)
Additional file 4:
The list of hub genes/proteins from the directed PPI network. The criteria of Indegree and Outdegree have been used for the identification of downstream and upstream regulators in directed PPI network, respectively. (XLS 31 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Talebi, R., Ahmadi, A. & Afraz, F. Analysis of protein-protein interaction network based on transcriptome profiling of ovine granulosa cells identifies candidate genes in cyclic recruitment of ovarian follicles. J Anim Sci Technol 60, 11 (2018). https://doi.org/10.1186/s40781-018-0171-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40781-018-0171-y