
Abstract
Background
Animal social systems can be described through four main components: social structure, social organization, mating system, and care system. Social structure describes the relationships between individuals in a population, while social organization describes the group composition, size, and spatiotemporal variation of a population. We use the frameworks of social structure and social organization to study the social system of Microlophus atacamensis, a lizard found in the rocky intertidal zone along the Chilean coast. The area M. atacamensis inhabits poses specific challenges stemming from their use of two distinct habitat types in the intertidal zone: they forage in the cool areas near the water’s edge and use large rocks in more inland areas for basking and refuge.
Methods
Our assessment of their social system focused on two separate populations: one to characterize social structure by means of focal observations and social network analysis, and a second to assess social organization via home range and core area analyses. Further, we examined the social system in two habitat types that comprise the intertidal zone: cobble fields and interspersed large rocks.
Results
Social network analysis revealed an interconnected network with a few highly central individuals. Body size influenced the outcomes of aggressive interactions, with interactions being more common in cobble fields where males had more associates and more repeated interactions than females. Spatial analyses revealed that the social organization of M. atacamensis is characterized by (1) high home range overlap, specifically in the cobble fields and (2) relatively exclusive core areas dispersed across both habitat types.
Conclusion
A social system is composed of both cooperative and competitive behaviors. While our study focused on competitive interactions, the extent and influence of cooperative behaviors is still unclear and merits future research. We suggest that M. atacamensis has a variable social system in which territoriality on large rocks affects access to stationary resources in that habitat (e.g., basking sites and refuges), while competition in the cobble fields could lay the foundation for a system of dominance relationships controlling access to variable food resources in cobble field areas of the intertidal zone.
Similar content being viewed by others


Introduction
An animal’s social system influences many aspects of their ecology. Understanding social systems has been a topic of major interest, with various studies examining the evolution of social systems in relation to environmental factors (i.e., microhabitat and resource availability) and phylogeny [1,2,3]. Differential access to limited resources can result in variations in behavior [1] that in turn affects social interactions, and can be manifested in how animals use space or are socially connected to one another, either of which can affect the social system that emerges [4].
A newly proposed comprehensive framework describing social systems uses four core components: social structure, social organization, mating system, and care system [4]. Social structure describes how individuals in a social unit interact and the relationships that form from repeated interactions (i.e., competitive and cooperative relationships). Social organization describes the group size, composition, and kinship patterns of a social unit (i.e., solitary, pair-living, or group-living) including their spatiotemporal variation. The other two components are the mating system, often described with a categorical classification of their pattern of reproduction, and the care system, describing who cares for dependent young and the nature of parental and alloparental care, or lack of parental care [4].
Social Structure
Social structure is defined by the individuals in a social group interacting repeatedly, and the patterns that arise from their interactions. Social structure encompasses affiliative, cooperative, or tolerant social bonds as well as agonistic relationships [4]. The intensity, nature, and frequency of interactions contribute to the overall social structure, with dominance hierarchies sometimes, but not always, playing a role [5, 6]. The accumulation of varying types of interactions forms the basis of a social structure, and for participating individuals results in costs and benefits to fitness. Social structure is frequently tied to fitness, as interactions modulate access to resources. For example, dominance hierarchies and agonistic interactions influence which individuals can obtain resources like food, refuges, and mates [7]. In addition, cooperative and tolerant interactions can affect fitness by providing mutualistic benefits like increased fertilization success [8], predator avoidance and detection [8, 9], better thermoregulation [9], and, via effective communication of warning signals and information, higher foraging efficiency [10]. While not commonly observed in reptiles, allyship or “coalitions” between individuals can also form and confer benefits to involved members [11], as with male marine iguanas (Amblyrhynchus cristatus) that attain higher mating success when associating proximate to dominant males [5].
Reptiles, specifically lizards, have historically been thought of as “asocial” or simple in their intraspecific interactions in part owing to a relative lack of research on reptiles [3, 12]. However, agonistic displays have been relatively well-studied in lizards for the role they play in access to resources and mates [e.g., 5], perhaps because agonistic displays are more readily observed and defined in the field, and initiated under controlled experimental conditions. Despite the fact that many studies investigating sociality in reptiles do not make a clear distinction between social structure and organization, the complexity of reptile social behavior is increasingly being recognized [12] in both the overall social system [9], and in the frequency and complexity of cooperative behaviors (e.g., Pholidoscelis foraging behavior) [6]. Social network analysis provides a powerful tool for quantifying components of social structure, as networks are built by evaluating dyadic relationships with respect to phenotypes influencing sociality [13] and environmental influences like habitat complexity [14] and seasonality [15].
Social organization
Social organization often depends on numerous factors including population density, habitat structural complexity, and resource availability, and can be examined via group size, composition, and dynamics [1, 7]. When essential resources are scarce but spatiotemporally heterogeneous, animal distributions are often linked to the location of the resources [10], with habitat features related to thermoregulation being especially important for ectotherms, with the distribution of suitable locations for thermoregulation influencing group size and composition [16,17,18,19]. Examining microhabitat use can provide vital information on how spatial segregation between individuals can modulate social organization.
Space use is inexorably related to social organization as sharing space or excluding individuals from a location affects group composition, making spatial analyses a useful tool for understanding social organization [e.g., 20, 21]. Space use can be evaluated via an analysis of home range (i.e., the area traversed during daily activities [22,25,24]) and core area (i.e., an area of concentrated activity for an individual [25]). The extent to which home ranges or core areas overlap, which can be indicative of territoriality or social tolerance, provides an additional assessment of social organization [26, 27].
Social System
The social system of a species can vary considerably as a result of their evolutionary history in combination with the environmental characteristics of their habitat [1, 2, 27, 28]. Because there is a focus on agonistic behaviors over cooperative behaviors in lizards, social systems have been described in terms of dominance hierarchies and territoriality. If a dominance hierarchy exists, social structure will depend on relationships between individuals independent of location, with the result of aggressive interactions often predictable based on the ranks of individuals [29]. By contrast, if a territorial system exists social organization is affected, with activity centered around locations where individuals exclude competitors from resources [29]. Social structure and social organization describe different characteristics of social systems and as such, dominance hierarchies and territoriality are not mutually exclusive [30]. Social network analyses can be used to identify and quantify the social structure, including an evaluation of whether dominance hierarchies exist, whereas habitat use and spatial analyses can be used to reveal whether the social organization includes territoriality. Linkages between network and spatial analyses can reveal nuanced aspects of social systems that would not be evident if only one component was assessed [6].
The ecology of Microlophus atacamensis, a lizard endemic to the intertidal zone in coastal Chile, provides a unique opportunity to assess location-specific aspects of their social system. The species occupies two habitats in the intertidal zone—shoreline cobble fields that are thermodynamically unfavorable but provide good food sources, and the adjacent, more inland areas of the Atacama Desert with large rock outcrops that provide better basking and shelter options. Ocean currents circulate cold waters year-round that keep the intertidal zone cool but subsidized with significant food items (e.g., algae and organic matter, plus the attracted insect food sources [31, 32]) in an unpredictable frequency, location, and density. The defense of food resources is only beneficial at intermediate abundance levels because when too low the costs of defense exceed the gains, while at very high abundances the benefits of defense are minimal [7]. Further, the heterogeneity of resources in space and time also can affect whether territoriality is part of the social system, as the further apart resource patches are the larger the territory necessary to defend, and the higher the costs of defense [10]. The instability and variation in abundance of the shoreline food resources make defense of marine subsidies less likely, while defense of basking locations and refuges on the inland rocks are much more probable as they are more stable and less likely to change across time and space.
The separation of food and thermoregulatory resources presents an opportunity to assess the overarching idea that understanding the social system of M. atacamensis requires an awareness of both social structure (social network) and social organization (space use). The extent to which cooperation or agonistic behavior shape the social system will rely on environmental conditions and are based on the costs and benefits to individuals resulting from changing group dynamics. Cooperation can provide benefits like higher inclusive fitness in harsh environments [9]. Likewise, agonistic behaviors can increase individual fitness, but the prevalence of conflicts and their effect on fitness will vary with resource distribution and abundance. Outcomes of agonistic encounters often provide reproductive benefits for victors and decrease reproductive prospects for losers [7]. We explore the interplay between social structure and social organization in M. atacamensis to better understand their social system. We test two main hypotheses. First, that social structure is independent of habitat type (e.g., cobble fields vs. rock outcrops). We predict that the social structure present in the cobble fields (e.g., social network) will be preserved on the large inland rocks. Second, we hypothesize that social organization, as reflected in patterns of space use, will vary with habitat type and by sex. We predict that social organization will consist of home ranges that contain exclusive use areas, with the extent of space use exclusivity and the size of home ranges varying with sex.
Methods
We studied the social system of M. atacamensis in two field sites separated by 1.5 km along the rocky intertidal coastline near Peña Blanca, ca. 40 km south of Huasco, Huasco Province, Atacama Region, Chile (Site 1: 28.6979°S, 71.3182°W; Site 2: 28.6901°S, 71.3076°W; datum = WGS84). The distance between the two sites ensured that the lizards at each site represented separate populations. Both sites were characterized by two distinct habitat types: (1) cobble fields, which contained cobble-sized stones and comprised central arenas bounded by large rock outcrops and (2) large rocks, which consisted of isolated boulders (3–6 m in height) and rock walls (> 3 m in height) forming the boundaries of cobble fields aligned approximately east-west from the Pacific Ocean inland to the edge of the Atacama Desert (Fig. 1A). Site 1 consisted of a single cobble field bounded by large rocks. Site 2 was composed of multiple cobble fields, each bounded by large rocks that collectively formed a contiguous block. At site 2, some interior large rocks bounded two cobble fields. The two sites were chosen based on their different functions. Site 1 was small enough to observe daily interactions but was not logistically practical for collecting spatial data as many large rocks could be surveyed only from within the site in a way that would have disturbed behavior. Site 2 was more suited to spatial analysis because the rocks could be traversed without disturbing the animals. Although we studied M. atacamensis at two different sites, the purpose of our study was not to compare behavior between sites, but rather to characterize the social system of the species by primarily exploring different aspects of the social system at each site.

(A) A depiction of the two habitat types, with a large cobble field surrounded by rocks walls on either side, and a large boulder in the foreground (depicting area outlined in red in Fig. 5). The transition from sandy desert to the cobble field is also visible in the foreground and the transition into the ocean in the background. Most of the interactions we observed occurred in the cobble fields. Photo by M. Eifler. (B) Spatial proximity of a marked male and female M. atacamensis. Photo by C. Staley
Data Collection
Prior to collecting data, we marked most of the animals in the study populations. Lizards on both sites were captured using a lasso attached to an extendable pole. They were then measured (mass (g) and snout-vent length (SVL, mm)), sexed via cloacal probing, and uniquely marked using non-toxic paint pens. Lizards were released at their initial sighting location within 5 h. Unmarked lizards that we used for data collection were captured after sampling. If multiple unmarked lizards were seen during the observation period, they were excluded from data collection and marked after the day’s observation period concluded. Data collection started once lizards had become active for the day, which varied with the weather.
Site 1
Our site for examining social structure (area = 0.134 ha) was characterized by a central cobble field bordered by 12 large rocks (Fig. 1A). From 22 Dec 2019–4 Jan 2020 between 1300 and 1830 h, four observers simultaneously monitored the lizards in the cobble field to record all interactions. The sampling periods lasted 30–120 min, depending on lizard activity levels. For behavioral sampling, one person was positioned in each corner of the cobble field to ensure that the entire central arena could be monitored. Simultaneously, a fifth observer surveyed the bordering large rocks every 30 min, scanning each with binoculars to record occupancy and compare behavior of lizards on rocks to lizards in the cobble fields. To ensure that surveys did not interfere with behavioral sampling, the observer walked around the edges of the large rocks as far from the central cobble field as possible. After the observation period concluded, all five observers reviewed notes and interactions to ensure that all recorded interactions were unique. Interactions between two or more lizards were categorized as spatial proximity (Fig. 1B), chases, fights, displacements, or retreats (Table 1).
Site 2
Our site for examining social organization (area = 1.02 ha), consisted of six large rocks interspersed with five cobble fields. Site 2 was larger than site 1, but was contiguous so that a single lizard could still travel across multiple large rocks and cobble fields (Fig. 1A). We studied home ranges and space use of M. atacamensis by obtaining location information through surveys (1–4 per day) conducted every 90 min from 10 to 17 Jan 2020, between 1000 and 1900 h. Four surveyors walked evenly spaced transects parallel to the shore, using binoculars to scan for and identify individuals. Each time a marked animal was observed, we recorded their identity, time, and location (i.e., large rock or cobble field and GPS coordinates (myTracks GPS-logger for iPhone X)). To ensure independence, we only used locations for individuals that were separated by at least 1 h. Each day, we alternated whether surveys began at the north or south end of the study site. We moved in the same direction throughout each day, to ensure that animals had ca. 90 min to recover from any disturbance that our presence might have caused.
Analysis
Social structure
We constructed and analyzed social networks for site 1 animals with Ucinet (ver. 6.671; [33]) and the igraph package in R (ver. 4.0.0; [34]), treating all lizards as nodes and considering individuals observed interacting as having a social link (= network edge). Social links were directional for all interaction types (i.e., representing dominant-subordinate encounters) except spatial proximity, which is non-directional (Fig. 1B; Table 1). To measure linear dominance in the population, we calculated Landau’s index (h) using directional social links [35]:
where n is the number of individuals in the population and \({{\rm{v}}_{\rm{I}}}\) is the number of individuals dominated by individual I. Landau’s index ranges from 0 to 1 where 1 represents a completely linear dominance hierarchy and 0 indicates that there is no consistent pattern of dominance. To assess a lizard’s centrality, a measure of the relative importance of the position of the individual to the structure of the social network [13], we calculated degree, strength, betweenness, and assortativity (Table 2) [13]. To assess general network properties, we compared the degree distribution to both Poisson and power-law distributions using chi-square tests and regressions [13, 36]. For the power-law distribution, we determined the relationship between log(degree) and log(probability(degree)) [13]. We also determined network density, calculated as the proportion of social links present relative to the total number of possible social links, and identified network components, which are sets of interconnected nodes separate from the rest of the network. On site 1, after filtering the dataset to include only individuals with degree ≥ 1 [13, 37], we analyzed the social network in three ways: (1) population-level network analysis using all interactions collected for all individuals (= overall network), (2) interactions observed on large rocks only (= rock network), and (3) interactions observed in the cobble field only (= cobble field network).
Social metrics (betweenness, degree, and strength) were not normally distributed so we used non-parametric tests for comparisons. We used paired Wilcoxon tests to compare social metrics between habitat type on site 1 (i.e., large rock network vs. cobble field network) for lizards present in both networks. We compared the frequency of interactions, as well as social links within and between the sexes, using chi-square goodness of fit tests, examining standardized residuals (> 2) to identify observations that strongly differed from expectations. For some analyses, the different types of directional interactions were pooled to compare with non-directional interactions (i.e., spatial proximity). The expected values were calculated based on the sex ratio present in the overall network. To assess the significance of the assortativity values, we used permutation tests (2000 permutations) where social links were randomly reassigned in our network. We then compared our observed values to the randomized distribution when identifying statistical significance. Additionally, to examine for differences between the cobble field and large rock habitats, we calculated assortativity using the interactions seen in each habitat type and used permutation tests to check significance. General linear models were developed to compare social metrics (degree, betweenness, and strength) to demographic measures. A step-wise model selection process was employed, using sex, SVL, and their interaction for the model examining relationships to degree, adding in degree for strength and betweenness models. We used a square root transformation for normality.
Social Organization
We calculated home range estimates on site 2 using Ranges (ver. 7; [38]) and analyzed them statistically in R (ver. 4.0.0; [34]). Home range size was estimated using minimum convex polygons (MCP; [41]) and neighbors were defined as individuals whose home range polygons exhibited any overlap. We calculated percent home range overlap for each individual relative to all other individuals in the population and summed pairwise overlap, resulting in many individuals with percent overlap values > 100%. Core areas were estimated using 50% MCPs (i.e., the smallest polygon containing 50% of an individual’s locations; [39]). Core area overlap was calculated for each lizard relative to all other lizards in the population, using each individual’s 50% MCP. We developed general linear models to examine space use measures with sex and body size as factors in a step-wise model selection process, which resulted in six models, one each for home range size, core area size, home range overlap, core area overlap, number of home range neighbors, and number of core area neighbors. In addition, we tested for nested influence, where aspects of home range predicted aspects of core area, and whether overlap predicted number of neighbors, as well as per capita overlap. Per capita home range overlap and per capita core area overlap was calculated as the average home range or core area overlap for each individual with its neighbors. We used paired Wilcoxon tests to examine the variation in the per capita male and female home range or core area overlap.
We examined home range overlap and core area overlap in relation to the habitat type in ArcGIS Pro (version 2.7.26828). For each lizard, 100% MCPs were plotted using their GPS coordinates, then all the individual polygons were overlaid to compute the number of individuals overlapping in each cell (1 × 1 m) throughout the study site. The same method was used for core areas (50% MCPs).
Results
As expected, males were larger in body size than females (Mann-Whitney U test: SVL: W = 1636.5, P < 0.05, males: 121 (64–138 mm) vs. females: 97 (69–112 mm); mass: W = 1659, P < 0.05, males: 63 (9–99 g); vs. females: 34 (11–56 g)). The population size on both sites was similar (site 1, n = 44; site 2, n = 43), as was the number of males and females (19 males on each site; 25 females on site 1, 24 females on site 2), but site 2 was almost eight times larger than site 1 (1.02 ha vs. 0.134 ha, respectively) resulting in a density on site 1 that was almost eight times higher than site 2 (328.4 vs. 42.2 lizards/ha, respectively). The sex ratio of adults did not differ significantly from 1:1 (1-sample proportion test: site 1, P = 0.45; site 2, P = 0.54) and was the same on both sites (Fisher’s exact test: P = 1).
Social Structure
Site 1 Social Network analyses
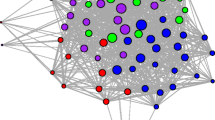
Of the 44 individuals on site 1, 30 (68%) lizards had social links and were part of the network, while 14 lizards were unconnected to the network. On site 1, the network had one main component and two smaller components consisting of pairs of individuals (Fig. 2). In the main component, both sexes were equally represented and the individuals with the highest degree included both males and females. There was no sex difference in the proportion of individuals with a degree of 0 (i.e., unconnected to the network; females (n = 10/25; 40%) vs. males (n = 4/19; 21%); Fisher’s exact test: P = 0.21). We observed 172 interactions on site 1, 37 of which (22%) occurred on the large rocks and 135 (78%) in the cobble field. The interactions composed 68 social links yielding a network density of 15.6%.

Site 1 social network (n = 30 individuals). Each circle is a node and represents a unique individual, with males in gray and females in black. Highly central individuals (degree ≥ 10) are represented by nodes with white circles. Lines between nodes are network edges, representing social links between lizards. Edges are weighted by the number of interactions that were observed between a pair of individuals. For social links that occurred only once, weight = 1, for those occurring 2–5 times, weight = 2, and for those occurring 6–14 times, weight = 3
Network structure
The degree distribution did not follow a Poisson distribution (X2 = 82.9; df = 11; P < 0.001) but approximated a power-law distribution with a slope of 1.1 (F1,11 = 30.4, P < 0.001, R2 = 75.3%). Most lizards in the network were somewhat peripheral, having relatively low degree, betweenness, and strength values, while a small subset of individuals in the network were highly interconnected (Figs. 2 and 3). Male and female degree (X̄ = 4.6 ± 0.9; 4.5 ± 0.9, respectively), betweenness (X̄ = 16.1 ± 6.0; 13.7 ± 5.1), and strength (X̄ = 13.4 ± 3.6; 9.5 ± 2.6) distributions did not differ (Chi-square test: degree, X2 = 0.6, df = 2, P = 0.76; betweenness, X2 = 1.4, df = 2, P = 0.49; strength, X2 = 3.9, df = 2, P = 0.14). In the step-wise model selection process degree was not related to SVL, sex, or their interaction. The best fit model for betweenness included SVL, which was not significant (F1,27 = 2.4, P = 0.13) but indicated that betweenness was strongly related to degree (F1,27 = 45.6, P < 0.001; overall R2 = 66.4%). In the best fit model for strength in the social network analysis, strength was positively related to both SVL (F1,27 = 5.3; P = 0.03) and degree (F1,27 = 131.5; P < 0.001; overall R2 = 84.4%). Examining associative tendencies for sex, SVL, and degree yielded values ≈ 0 (Table 3), indicating no strong inclination to associate with the same sex, similar-sized lizards, or with lizards of similar connectedness, respectively. We did not detect a linear dominance hierarchy in M. atacamensis (Landau’s index: h = 0.02), but the dominance matrix used for the calculation was largely unpopulated (i.e., low network density), which could preclude a determination of existing dominance patterns.

Distribution of social metrics for site 1 (n = 30). (A) Degree denotes the number of individuals interacting with a specific lizard. For M. atacamensis, highly central individuals are the five individuals with the highest degree values (i.e., the right tail of the x-axis). (B) Betweenness indicates the number of shortest paths between all possible pairs of animals that pass through a focal animal. A few highly central individuals have high betweenness values, while a majority of lizards have betweenness closer to 0. (C) Strength values indicate the total number of interactions for each individual
Interactions
About half of the total interactions were spatial proximity (Fig. 1B; n = 88, 51%), with the remaining interactions resulting in a clear winner (= directional; n = 84, 49%). The proportions of spatial proximity (49% vs. 51%) and directed interactions (51% vs. 49%) were similar in cobble fields and on large rocks, respectively. Male-male directional and male-female spatial proximity interactions were more common than random expectations (Chi-square test: Χ2 = 8.97, df = 2, P = 0.011; Table 4).
Among the directional interactions, larger individuals won interactions more often (79%) than smaller individuals (21%). For same-sex interactions, larger lizards won most often (n (% larger victors): males 34 (97%), females 13 (77%)). In male-female directional interactions (n = 37), larger lizards won 62% of the time and among interactions where larger individuals won, the winners were male in all but one interaction (96%). In male-female interactions where the smaller individual won, all winners were female (Table 5). Of the male-female directional interactions, females won 25% of 16 chases, 67% of 12 displacements, 33% of three fights, and 33% of six retreats (Table 4).
Between habitats
Of the 68 social links we observed, most occurred on the cobble field (59%), where there were almost twice as many as on the large rocks (32%); only six pairs of lizards were seen interacting in both habitats (9%; Table 6). In the cobble field and on large rocks, same sex and intersex social link frequencies were similar to random expectations (Chi-square goodness of fit test: cobble field, X2 = 1.62, df = 2, P = 0.44; large rocks, X2 = 2.16, df = 2, P = 0.34). Male degree and strength were significantly larger in the cobble fields than on the large rocks (paired Wilcoxon test: n = 8; degree, V = 0; P = 0.022; strength, V = 1; P = 0.016) but did not differ for females (n = 7; degree, V = 3.5; P = 0.341; strength, V = 2; P = 0.093). Betweenness did not differ significantly between habitats for either sex (males, V = 8, P = 0.35; females, V = 7, P = 0.529). When split by habitat, assortativity values for sex, SVL, and degree were ≈ 0, indicating that there was no trend in a lizard’s assortative mixing, except for degree on large rocks where lizards with similar degree values interacted more than randomly expected (P = 0.042; Table 3).
Social Organization
Space use
We conducted 21 surveys on site 2, using 557 locations obtained from lizards with > 3 sightings for the home range analyses (n = 39 lizards; X̄ = 14, range = 6–22 sightings per lizard). Mean home range size was 715 m2, while mean core area was 89 m2 (Table 7). Home range size was positively related to the number of locations used to calculate the MCPs (F1,27 = 11.63, P = 0.002; overall R2 = 23.9%), with the mean number of sightings likely to underestimate home range size for some animals (Fig. 4). Core area size was positively related to both the number of locations used (F1,27 = 19.83, P < 0.001) and home range size (F1,27 = 11.38, P = 0.002 overall R2 = 55.8%). No included variable was strongly related to home range overlap, but we retained home range size in the model F1,27 = 2.22, P = 0.145; overall R2 = 5.7%). Core area overlap was positively related to home range overlap (F1,27 = 19.59, P < 0.001; overall R2 = 35.2%). The number of home range neighbors was significantly related to home range size (F1,27 = 19.92, P < 0.001; overall R2 = 35%), while the number of core area neighbors was related to home range overlap (F1,27 = 6.4, P = 0.016), core area overlap (F1,27 = 17.14, P < 0.001), and core area size (F1,27 = 5.96, P = 0.02; overall R2 = 63.2%). Sex and body size were not related to any of the metrics.

Relationship between the number of sightings and the final home range estimation. The number of sightings used to calculate home range size versus percent of the final home range estimate. Black points represent female lizards, while gray points represent male lizards. The blue line is the average percent of the final home range estimate for animals with a given number of points used to assess home range
Overlap
Core areas overlapped significantly less than home ranges (paired Wilcoxon test: V = 741, P < 0.05; Fig. 5). Lizards shared a home range with X̄ = 14.5 neighbors, but only shared a core area with X̄ = 2.5 neighbors. When examining the extent of overlap in home range compared to core area, a similar difference in magnitude is evident: lizards had a mean home range overlap of 458%, but a mean core area overlap of only 62% (Table 7; Fig. 5). When examining overlap by location on the study site (Fig. 5), most home range overlap occurred in the largest cobble field, with up to 14 lizards sharing space (i.e., home ranges). Most areas outside of the largest cobble field were used by 3–5 individuals and areas of high overlap extended onto the edges of the large rocks along the cobble field borders (Fig. 5). When core areas were mapped by location, most core areas were exclusive with a maximum of three individuals overlapping (Fig. 5).

Map of space use across site 2. (A) 100% MCPs (i.e., home ranges), with overlap concentrated in the largest cobble field outlined in red (depicted in Fig. 1 A). (B) 50% MCPs (i.e., core areas); note the much lower overlap among polygons. Location of study site relative to the ocean is illustrative of position and not to scale
Sex differences
The sexes did not differ in the number of home range or core area neighbors (Mann-Whitney test: home range, W = 339.5, P = 0.573; core area, W = 301.5 P = 0.386). They also had similar levels of overlap in their home ranges and core areas (Mann-Whitney test: home range, W = 354.0, P = 0.777; core area, W = 323.0, P = 0.814). Among males, the average home range overlap with male neighbors did not differ from the average home range overlap with female neighbors (paired Wilcoxon test: V = 68, P = 0.7119). Among females, the average home range overlap with female neighbors was greater than the average overlap with male neighbors (paired Wilcoxon test: V = 39, P = 0.006). However, for both males and females, the average core area overlap with male neighbors did not differ from the average core area overlap with female neighbors (paired Wilcoxon test: males, V = 13, P = 0.546; females, V = 44, P = 0.46).
Discussion
The social system of M. atacamensis is complex and can be understood better by examining both social structure and social organization. Our analysis of social structure indicated that the social network is held together by a few key individuals, with connections maintained by male-male competition and male-female associations that vary across the two habitats. Our examination of their social organization indicated that individuals overlapped broadly in their use of space, but maintained small exclusive areas in the large rock habitat. In combination, social structure and social organization point towards a social system that is not strictly territorial, but is composed of location-dependent territoriality (on large rocks) and location-independent dominance relationships (in the cobble fields), likely driven by the heterogenous distribution of resources.
Social Structure
The social network of M. atacamensis contained many interconnected individuals, but most had only a few associates (i.e., low degree; Fig. 3) and few of the possible social links were realized (i.e., low network density (15.6%)), indicating that the network is held together by indirect connections maintained through a few highly central individuals (Figs. 2 and 3). The degree distribution approximated a power law, pointing out that our network resembles the “fit-get-rich” model, in which a few nodes with high degree values link nodes of lesser degree [40]. However, our interpretation is tentative, as the scale of our network is considerably smaller than other networks analyzed similarly (e.g., the world wide web) [41]. The indirect connections we observed might arise from the separation of resources (e.g., basking sites and feeding areas) that results in highly central individuals interacting with different lizards in the two habitat types. The extent to which “central” individuals actively pursue interactions or simply move in ways that increase the likelihood of encounters requires further investigation.
Social systems can be variable when food is inconsistent or unpredictable [1, 26]. While the intertidal zone can contain high food availability, territoriality is unlikely in the cobble fields due to the unpredictability of marine subsidies, which lowers the net benefit of defending food resources [7, 42] but does not preclude a stable dominance system controlling access to food in the cobble fields. In our population of M. atacamensis, larger individuals engaged in more repeat interactions (i.e., higher strength) and won more of their encounters, indicating that body size could form the basis of a dominance hierarchy. Body size can be important in determining which individuals access, acquire, or defend resources [6, 43, 44]. Larger M. atacamensis might have higher resource holding potential and be better able to defend small, high-quality, food areas that frequently change location within the cobble fields. Defending or controlling entry to small areas in the intertidal zone can occur as food density and competitor abundance changes, as when Sanderlings (Calidris alba) adjust their territorial defense with shifting feeding sites [45]. Despite not finding evidence of linear dominance (via Landau’s index) in our population, the timeframe of our study could have influenced our ability to detect a dominance hierarchy (days vs. months [46]). The extent to which dominance relationships form can be appraised by more intensive data sampling. Additionally, future investigations into the characteristics associated with resource holding potential and whether individuals defend small areas in the cobble fields, as well as the extent to which food abundance and locations shift is merited.
Most lizards did not interact with the same associates on both large rocks and in the cobble field (Table 6), indicating the importance of habitat-specific interactions (i.e., cobble field or large rocks). When comparing habitat-specific networks, most social links occurred in the cobble field. Further, males were more socially engaged (i.e., higher degree and strength) in the cobble fields than on large rocks, while females interacted similarly in both habitats. Although not quantified, we often observed larger male lizards displaying from high points in the cobble field and seeming to control which individuals accessed the splash zone [pers. obs.]. More elevated display locations are often associated with greater dominance as they can could enhance effective communication distance by providing greater visibility [20, 47].
In terms of social links, same sex and intersex social links were similar to random expectations in the cobble field (Table 6), supporting the idea that males and females use the cobble fields in similar ways. However, overall male-male aggressive and male-female non-aggressive interactions were most common (Table 4). Male-male aggression is common when there is competition for resources [48], while male-female associations involving spatial proximity could be a way of strengthening reproductive pairs or alliances. In the Australian sleepy lizard, Tiliqua rugosa, long-term male-female associations increase reproductive success [8]. Ultimately, the high frequency of male-male and male-female interactions indicates the importance of male competition and opposite sex cooperative or tolerant associations in shaping the social system.
Social Organization
Our prediction that social organization, reflected by patterns of space use and overlap, would vary with habitat was supported. Lizards exhibited relatively high home range overlap, primarily in the cobble fields rather than on large rocks (Fig. 5). Core areas overlapped much less, tended to be exclusive, and were more dispersed across the two habitat types than home ranges (Fig. 5). Overall, the cobble fields were areas of high use, as indicated by high home range overlap, whereas the large rocks were isolated areas where M. atacamensis spent a majority of their time, as evidenced by low core area overlap. The resource dispersion hypothesis asserts that when resources are distributed unevenly, larger areas are needed by individuals to encompass the variation in necessary resources across the habitat [10], which could explain M. atacamensis space use. Home ranges are necessarily large to access food in the cobble fields, while also covering enough area that individuals can maintain exclusive core areas on the large rocks for refuge and basking. The low overlap in core areas provides circumstantial support for the notion that M. atacamensis could be territorial on the large rocks. We hypothesize that their core areas function as refuge and basking locations, but quantifying specific microhabitat use was beyond our scope. Future research could test the hypothesis and assess the extent of competition for specific rock features.
Sex differences in Social Organization
Home range and core area size estimates and overlap did not differ significantly between the sexes, indicating an overall similarity in space use for males and females. The tendency for greater female per capita home range overlap with female neighbors points to a social organization of overlapping home ranges with non-random associations between females, indicating that females are more tolerant of sharing space with each other than males. However, all individuals regardless of sex had much more isolated core areas.
Conclusion
Animals can exhibit different behavioral strategies in different locations within their home range based on resource abundance [2, 49]. For M. atacamensis, the cobble field habitat was important to the social system, as activity in the cobble fields was greater than on large rocks when examining both social structure (i.e., higher frequency of interactions), and social organization (i.e., greater home range overlap). Although not all social systems are organized by aggression, we observed many agonistic interactions. Variation in agonistic behavior with habitat, body size, and sex points to the possibility that M. atacamensis could have a variable social system, with territoriality organizing the social system on large rocks, while aggressive interactions, potentially organized through dominance hierarchies, play a larger role in the social system in cobble fields. Microlophus atacamensis is not, by far, the only lizard species with a complicated ecology, thus the complexity we observed in their social system, specifically social structure and organization, is unlikely to be unique. We recognize that many other squamates likely have complex social systems that deserve further comprehensive study that would aid in the understanding of lizard socioecology.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Stamps JA. Sexual selection, sexual dimorphism, and territoriality. In: Huey RB, Pianka ER, Schoener TW, editors. Lizard Ecology: Studies of a Model Organism. Cambridge: Harvard University Press; 1983. pp. 169–204.
Martins EP. Phylogenetic perspective on the evolution of lizard territoriality. In: Vitt LJ, Pianka ER, editors. Lizard Ecology: Historical and Experimental Perspectives. Princeton: Princeton University Press; 1994. pp. 117–44.
Doody JS, Burghardt GM, Dinets V. Breaking the social-non-social dichotomy: a role for reptiles in vertebrate social behavior research? Ethology. 2013;119(2):95–103.
Kappeler PM. A framework for studying social complexity. Behav Ecol Sociobiol. 2019;73:1–14.
Partecke J, Haeseler A, Von, Wikelski M. Territory establishment in lekking marine iguanas, Amblyrhynchus cristatus: support for the hotshot mechanism. Behav Ecol Sociobiol. 2002;51(6):579–87.
Eifler D, Eifler M, Malela K, Childers J. Social networks in the Little Scrub Island ground lizard (Ameiva corax). J Ethol. 2016;34(3):343–8.
Maher CR, Lott DF. A review of ecological determinants of territoriality within vertebrate species. Am Midl Nat. 2000;143(1):1–29.
Leu ST, Burzacott D, Whiting MJ, Bull CM. Mate familiarity affects pairing behaviour in a long-term monogamous lizard: evidence from detailed bio-logging and a 31-year field study. Ethology. 2015;121(8):760–8.
Gardner MG, Pearson SK, Johnston GR, Schwarz MP. Group living in squamate reptiles: a review of evidence for stable aggregations. Biol Rev. 2016;91(4):925–36.
Johnson DDP, Kays R, Blackwell PG, Macdonald DW. Does the resource dispersion hypothesis explain group living? Trends Ecol Evol. 2002;17(12):563–70.
Silk J, Cheney D, Seyfarth R. A practical guide to the study of social relationships. Evol Anthropol. 2013;22(5):213–25.
Doody JS, Dinets V, Burghardt GM. The secret social lives of reptiles. Johns Hopkins University Press; 2021.
Croft DP, James R, Krause J. Exploring animal social networks. Princeton: Princeton University Press; 2008.
Leu ST, Farine DR, Wey TW, Sih A, Bull CM. Environment modulates population social structure: experimental evidence from replicated social networks of wild lizards. Anim Behav. 2016;111:23–31.
Mbizah MM, Farine DR, Valeix M, Hunt JE, Macdonald DW, Loveridge AJ. Effect of ecological factors on fine-scale patterns of social structure in African lions. J Anim Ecol. 2020;89(11):2665–76.
Schlesinger CA, Shine R. Selection of diurnal retreat sites by the nocturnal gekkonid lizard Oedura lesueurii. Herpetologica. 1994;50(2):156–63.
Eifler MA, Marchand R, Eifler DA, Malela K. Habitat use and activity patterns in the nocturnal gecko, Chondrodactylus turneri. Herpetologica. 2017;73(1):43–7.
Hedman HD, Chuga SC, Eifler DA, Hanghome GPK, Eifler MA. Microhabitat use of two sympatric geckos, Turner’s thick-toed gecko (Chondrodactylus turneri) and the common Namib day gecko (Rhoptropus afer). J Arid Environ. 2021;188:104448.
Croak BM, Pike DA, Webb JK, Shine R. Three-dimensional crevice structure affects retreat site selection by reptiles. Anim Behav. 2008;76(6):1875–84.
Eifler DA, Eifler MA, Eifler EN. Habitat use and movement patterns in the graceful crag lizard, Pseudocordylus capensis. Afr Zool. 2007;42(2):152–7.
Vasconcelos R, Santos X, Carretero MA. High temperatures constrain microhabitat selection and activity patterns of the insular Cape Verde wall gecko. J Arid Environ. 2012;81:18–25.
Burt WH. Territoriality and home range concepts as applied to mammals. J Mammal. 1943;24(3):346–52.
Börger L, Dalziel BD, Fryxell JM. Are there general mechanisms of animal home range behaviour? A review and prospects for future research. Ecol Lett. 2008;11(6):637–50.
Passos DC, Galdino CAB, Rocha CFD. Challenges and perspectives for studies on home range of lizards from South America. South Am J Herpetol. 2015;10(2):82–9.
Vander Wal E, Rodgers AR. An individual-based quantitative approach for delineating core areas of animal space use. Ecol Modell. 2012;224(1):48–53.
Fox SF, Shipman PA. Social behavior at high and low elevations: Environmental release and phylogenetic effects in Liolaemus. In: Fox SF, McCoy JK, Baird TA, editors. Lizard Social Behavior. Baltimore: The Johns Hopkins University Press; 2003. pp. 310–434.
Stamps JA. Social behavior and spacing patterns in lizards. In: Gans C, Tinkle DW, editors. Biology of the Reptilia. New York: Academic Press; 1977. pp. 265–334.
Stone PA, Snell HL, Snell HM. Island biogeography of morphology and social behavior in the lava lizards of the Galapagos Islands. In: Fox SF, McCoy JK, Baird TA, editors. Lizard Social Behavior. Baltimore: Johns Hopkins University Press; 2003. pp. 190–239.
Maher CR, Lott DF. Definitions of territoriality used in the study of variation in vertebrate spacing systems. Anim Behav. 1995;49(6):1581–97.
Effenberger E, Mouton PLFN. Space use in a multi-male group of the group-living lizard. J Zool. 2007;272(2):202–8.
Spiller DA, Piovia-Scott J, Wright AN, Yang LH, Takimoto G, Schoener TW, et al. Marine subsidies have multiple effects on coastal food webs. Ecology. 2010;91(5):1424–34.
Wright AN, Piovia-Scott J, Spiller DA, Takimoto G, Yang LH, Schoener TW. Pulses of marine subsidies amplify reproductive potential of lizards by increasing individual growth rate. Oikos. 2013;122(10):1496–504.
Borgatti SP, Everett MG, Freeman LC. Ucinet for Windows: Software for Social Network Analysis. Harvard: Analytic Technologies; 2002.
R Core Team. R: A language and environment for statistical computing [Internet]. Vienna: R Foundation for Statistical Computing; 2020.
Whitehead H. Analyzing animal societies: quantitative methods for vertebrate social analysis. Chicago: University of Chicago Press; 2008.
Almaas E, Vazquez A, Barabási L. Scale-free networks in biology. In: Kepes F, editor. Biological Networks. World Scientific Publishing Co. Pte. Ltd.; 2007. pp. 1–19.
Bejder L, Fletcher D, Bräger S. A method for testing association patterns of social animals. Anim Behav. 1998;56(3):719–25.
Kenward RE, Casey NM, Walls SS, South AB. Ranges9: For the analysis of tracking and location data. Wareham: Anatrack Ltd.; 2014.
Rose BR. Lizard home ranges: methodology and functions. J Herpetol. 1982;16(3):253–69.
Albert R, Barabási AL. Statistical mechanics of complex networks. Rev Mod Phys. 2002;74(1):47–97.
Barabási AL. Scale-free networks: A decade and beyond. Science. 2009;325(5939):412–3.
Connors PG, Myers JP, Connors CSW, Pitelka FA. Interhabitat movements by sanderlings in relation to foraging profitability and the tidal cycle. Auk. 1981;98:49–64.
Torr GA, Shine R. Patterns of dominance in the small scincid lizard Lampropholis guichenoti. J Herpetol. 1996;30(2):230–7.
Noble DWA, Wechmann K, Keogh JS, Whiting MJ. Behavioral and morphological traits interact to promote the evolution of alternative reproductive tactics in a lizard. Am Nat. 2013;182(6):726–42.
Myers J, Connors P, Pitelka F. Territory size in wintering sanderlings: the effects of prey abundance and intruder density. Auk. 1979;96(3):551–61.
Wittig RM, Boesch C. Food competition and linear dominance hierarchy among female chimpanzees of the Taï National Park. Int J Primatol. 2003;24(4):847–67.
Tokarz RR. Body size as a factor determining dominance in staged agonistic encounters between male brown anoles (Anolis sagrei). Anim Behav. 1985;33(3):746–53.
Knell RJ. Population density and the evolution of male aggression. J Zool. 2009;278:83–90.
Evans LT. Field study of the social behavior of the black lizard, Ctenosaura pectinata. Am Museum Novit. 1951;1493:1–26.
Acknowledgements
We are especially grateful to Juan Segundo Tapia who facilitated logistical support and secured permission from Marlene Rojas, Presidente del Sindicato de Algueros Costa Sur, SITRALSUR for use of their cabin. We thank the Museo Nacional de Historia Natural in Santiago, Chile for providing camp logistical equipment, and D. Smith and J. Soberon for their comments and help in reviewing early drafts of the manuscript. Finally, we are grateful to F. Patricio Ojeda and two anonymous reviewers whose detailed suggestions and comments substantially improved our manuscript. The research was done in partial fulfillment of an MA degree for Kaera Utsumi from the Department of Ecology and Evolutionary Biology at the University of Kansas.
Funding
Financial support was provided by Erell Institute and The Dunlap Charitable Fund. The Dunlap Charitable Fund and Erell Institute do not have competing interests and were not involved in research planning. The article processing charges related to the publication of this article were supported by The University of Kansas (KU) One University Open Access Author Fund sponsored jointly by the KU Provost, KU Vice Chancellor for Research & Graduate Studies, and KUMC Vice Chancellor for Research and managed jointly by the Libraries at the Medical Center and KU - Lawrence.
Author information
Authors and Affiliations
Contributions
H.N. and D.A.E. developed the general project idea and identified the field work location. All authors (K.U., C.S., H.N., M.A.E., and D.A.E.) conceived and designed the specific research questions and methods. H.N. arranged for logistical support for the field work, including assisting us in obtaining in-country permits, equipment, and housing at the field site. All authors (K.U., C.S., H.N., M.A.E., and D.A.E.) collected the field data. K.U., M.A.E., and D.A.E. analyzed and interpreted the results. K.U. drafted the original manuscript, with all other authors providing editorial and content revisions. M.A.E. and D.A.E. provided the critical revisions for intellectual content. All authors read and approved the final manuscript. K.U. is the guarantor and attests that all the listed authors meet the authorship criteria and that none have been omitted.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Our work adhered to the current Guidelines for the use of live amphibians and reptiles in field and laboratory research by the Herpetological Animal Care and Use Committee of the American Society of Ichthyologists and Herpetologists under the approval of Erell Institute’s Animal Care and Use Committee (IACUC proposal no. 2019-01). Research in Chile was authorized by permit No. 9223/2019 issued to H. Núñez by SAG Ministerio de Agricultura, Chile.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Utsumi, K., Staley, C., Núñez, H. et al. The social system of the lava lizard, Microlophus atacamensis: the interplay between social structure and social organization. Rev. Chil. de Hist. Nat. 95, 9 (2022). https://doi.org/10.1186/s40693-022-00113-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40693-022-00113-x