
Abstract
Background
The organic nitrogen source is one of the key factors affecting Mortierella alpina cell growth as well as arachidonic acid (ARA) production. The aim of the present work is to achieve an optimized recipe of organic nitrogen source for ARA production by M. alpina by testing four organic nitrogen sources.
Results
In the flasks, the results showed yeast extract or corn steep liquor were the most suitable sole nitrogen sources for biomass and ARA yield. In the bioreactor, a biomass of 17.5 g L−1 and an ARA yield of 2.7 g L−1 were achieved when using sole yeast extract, while cell autolysis was induced when using sole corn steep liquor; When using the combined nitrogen source with corn steep liquor and yeast extract at a 3:7 weight ratio, the biomass and ARA yield were significantly improved to 37 and 7.8 g L−1, respectively.
Conclusions
This work evaluated whether using a mix of organic nitrogen sources could improve ARA production when scaling up from a flask to a bioreactor culture. The 3:7 ratio of corn steep liquor to yeast extract was quite favourable for large-scale ARA production, and as a result, this combination has great potential for improving fungal cultures.
Similar content being viewed by others


Background
Arachidonic acid (all-cis-5, 8, 11, 14-eicosatetraenoic acid, ARA) is classified as a polyunsaturated fatty acid and is an important fatty acid for human nutrition. It is a direct precursor of several key eicosanoid hormones, which are important biological regulators. Owing to its unique biological properties, ARA has been widely used for a variety of applications in medicine, pharmacology, cosmetics, the food industry, agriculture, and other fields (Birch et al. 2000; Dedyukhina et al. 2011).
Traditionally, ARA was extracted from animal livers, adrenal glands, and egg yolk. These feedstock are readily available; they only contain small amounts of ARA. Consequently, these feedstock cannot be able to meet the amazing rising market demand for ARA. There has recently been an increased interest in using microorganisms to produce ARA (Ratledge 2004; Eroshin et al. 2000). As an alternative source containing a high intracellular content of ARA, the filamentous fungus, Mortierella alpina, has been extensively studied for ARA production at various fermentation scales (Peng et al. 2010; Vali et al. 2003). The previous studies reported that cell growth, total fatty acid and ARA yields were influenced by many factors, such as cell morphology, carbon and nitrogen sources, mineral supplementation, and the dissolved oxygen concentration (Higashiyama et al. 2002; Jang et al. 2005). Among these parameters, nitrogen source was reported to be of critical importance in the cultivation of M. alpina. However, not all nitrogen sources enhance cell growth and ARA production equally well (Lu et al. 2011; Totani et al. 2000). Compared with inorganic nitrogen sources, organic nitrogen sources showed higher levels of cell growth, total fatty acids and ARA production. Replacement of sodium nitrate by corn steep liquor (CSL) significantly improved ARA production by M. alpina ATCC 32222 (Higashiyama et al. 2004). Nisha and Venkateswaran (2011) also studied the influence of various nitrogen sources on both the biomass and the production of ARA by M. alpina. They showed that yeast extract (YE) produced the maximum biomass (6.8 g L−1) and highest ARA content in lipids (35.28 %), peptone produced the highest lipid content in biomass (42.0 %), and the urea produced the lowest ARA content in lipids (12.06 %). The previous studies also reported that natural nitrogen sources had a marked effect on the morphology of M. alpina in submerged culture (Park et al. 2001) and that the morphology of the cells had an effect on cellular enzymes and secondary metabolites. They found that the morphology of M. alpina cells changed from pelleted shape to filamentous form. It was suggested that the fluffy pellet morphology was more suitable for ARA production than the smooth pellet or filamentous morphology.
The diversity of available organic nitrogen sources is important for cell growth and ARA production, so it is necessary to further study the effect of different nitrogen sources on biomass as well as on the ARA yield for M. alpina. Furthermore, few studies have focused on adopting a combination of organic nitrogen sources to M. alpina cultivation. The aim of this work is to achieve an optimized recipe of organic nitrogen source for ARA production by M. alpina.
Methods
Microorganism and cultivation conditions
Mortierella alpina LU166 used in this study was a UV mutation of ATCC 3222, which was obtained from the American Type Culture Collection (ATCC, Rockville, USA). It was maintained on potato dextrose agar (PDA) slants at 4 °C and transferred monthly.
The seed medium contained (g L−1): glucose 30, yeast extract 4, KH2PO4 2.5 and MgSO4–7H2O 0.3. Inocula were prepared in 250 mL Erlenmeyer flasks containing 50 mL seed medium. The culture was grown for 72 h at 28 °C in a rotary shaker set at 150 rpm. The fermentation medium consisted of (g L−1): glucose 60, yeast extract 10, glutamic acid 1, MgSO4–7H2O 0.1, KH2PO4 0.5, CaCO3 0.05, and vitamin concentrate 1 mL (0.5 mg vitamin B12, 0.5 mg vitamin H, 1 mg vitamin B1, dissolved in 1 L of water). The pH of the medium was adjusted to 6.0 before autoclaving at 121 °C for 15 min. Scale-up batch cultures were carried out in a 3.6 L bioreactor containing 2 L fermentation medium. The culture was maintained at 28 °C with 0.5 vvm aeration and 200 rpm agitation. The pH was maintained at 4–6 in the exponential growth phase and at 7–8 in the subsequent fermentation period by feeding HCl or NaOH automatically. The experimental results comprise the average of two or three parallel experiments, and the data given in this paper are representative.
Analytical methods
The dinitrosalicylic acid method was used to assay the glucose concentration (Miller 1959). The biomass of fungal mycelia was harvested by filtration, washed with distilled water three times and then dried at 60 °C until the constant dry cell weight, (DCW) was obtained. The dried biomass was grinded into powder and used for extraction of the total lipids, which was determined gravimetrically. Fatty acid methyl esters (FAMEs) were prepared as follows: 5 mL 0.5 M KOH-methanol was added to a tube containing TLs. The tubes were heated in a water bath at 65 °C for 10 min, after then 5 mL 30 % BF3-ether was added. The tubes were then heated in a water bath at 65 °C for 30 min again, and then 5 mL hexane was added when the tubes cooled down to room temperature. It was settled for separation of two phases after adding 1 mL saturated sodium chloride solution for preventing emulsification. The upper phase containing FAMEs was applied to a gas chromatograph (Agilent GC 7890, USA) equipped with a 100 m × 0.25 mm capillary column (SP™-2560, USA). The column was increased from 140 to 240 °C at 3 °C/min and then maintained at 240 °C for further 30 min. The temperature of the injector and detector were both set at 260 °C, and nitrogen was used as the carrier gas at 20 cm/s. The quantity of ARA was estimated from the peak areas on the chromatogram using docosahexaenoic acid methyl ester as an internal standard.
The experimental design
Based on the literature (Lu et al. 2011; Nisha and Venkateswaran 2011), four types of organic nitrogen sources (YE, peptone, soybean meal, and CSL) were first tested individually in the flask culture at a concentration of 10 g L−1 to determine the optimal single nitrogen source. Then, the chosen optimal single nitrogen source was studied in the bioreactor culture to investigate its scale-up effect on ARA production. According to the results of the bioreactor culture using the sole nitrogen source, the proper nitrogen source mix based on the different weight ratios was tested for improving cell growth and ARA production from the flask culture to the bioreactor culture.
Results and discussion
Effect of a single nitrogen source on ARA production in the flask culture
Table 1 revealed that different type of nitrogen source in the cultivation media had a profound significant effect on cell growth, ARA production and mycelial morphology. CSL gave the maximum biomass and ARA yield in the culture, which also has the lowest residual glucose content at the end of fermentation. A slightly lower biomass and ARA yield were obtained in the medium containing YE, which is a typical nitrogen source for the cultivation of microorganisms, owing to the presence of metal ions and required micronutrients vital for the growth of microorganisms (Lan et al. 2002; Sakuradani et al. 2004). It is also showed that a high ARA percentage, as a fraction of total lipids (approximately 46 %), could also be obtained in medium supplemented with either CSL or YE as the sole nitrogen source. Although the medium containing soybean meal was favourable for cell growth, lipid and ARA production were much lower than that containing YE or CSL. This might be due to that soybean meal contains insoluble components which are not beneficial for the accessibility of cells. Table 1 also showed that peptone was unfavourable for M. alpina strain in this study. There was the highest residual glucose in the medium containing peptone, which limited the cell growth and almost totally inhibited the production of ARA. In addition, it was observed that peptone induced filamentous forms of mycelia during the cultivation process, whereas the other three nitrogen sources induced the formation of fluffy pellets. The former morphology correlates with a small ARA yield, and the latter morphology correlates with a considerably higher productivity of ARA. This result indicated that the fluffy pellet morphology was beneficial for the production of ARA, which is in agreement with previous studies (Higashiyama et al. 2002; Lu et al. 2011). Taking all factors into consideration, YE and CSL were chosen as the most suitable nitrogen sources for M. alpina scale-up culture, among the nitrogen sources tested.
Effect of a single nitrogen source on ARA production in the bioreactor culture
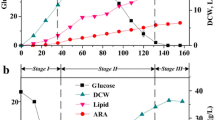
The culture of M. alpina was scaled up based on the results of the flask culture. When using YE as the sole nitrogen source, the mycelia of M. alpina grew normally in the bioreactor, and the morphology was maintained in a pellet form throughout the cultivation, similar to that in the flask culture, which is favourable for ARA production. The results in Fig. 1 showed that cells reached the maximum concentration at 108 h, which is an increase of 29.6 % over that of the flask culture. Total lipids and ARA achieved the maximum concentrations at 156 h, which individually increased by 114.3 and 107.7 %. Figure 1 also showed there was a high concentration of residual glucose (approximately 37 % of the original glucose concentration) in the fermentation broth, which means the carbon source in this medium could not be efficiently used to improve cell growth. It has been proven that the residual glucose level can affect the intracellular metabolite productivity (Shang et al. 2003), so the uptake of glucose should be enhanced in the fermentation. These results suggested that YE has a positive effect on the scale-up of cultures of M. alpina, especially with respect to lipid and ARA accumulation.

Profiles of glucose concentrations, DCW, total lipids and ARA yield in a 3.6-L bioreactor by M. alpina cultured in the medium using yeast extract as the sole nitrogen source. Black square, glucose; black circle, DCW; black star, total lipids; black up-pointing triangle, ARA yield
When CSL was used as the sole nitrogen source in the bioreactor, the painful problem of cell autolysis occurred during the culturing process of M. alpina. It was found that the cell morphology was filamentous, which was unfavourable for ARA production. CSL, as one type of quick-acting nitrogen source, was consumed so fast that it may have disturbed the consumed C/N ratio balance. A proper consumed C/N ratio also influences cell growth and lipid accumulation in the culture of M. alpina (Koike et al. 2001). However, the results in Table 1 showed that when CSL was used as the sole nitrogen source in flask culture, biomass, lipid and ARA yields were the highest, especially the ARA content in lipid. These results indicated CSL was more favourable for ARA production. In view of the results of the flask culture and the bioreactor culture, the combined nitrogen sources of CSL + YE were considered for the following experiment.
Effect of combined nitrogen sources on ARA production in the flask culture
As mentioned in the preceding section, a combination of nitrogen sources (CSL and YE) in shaking cultures were first tested. On the basis of several previous reports (Nisha and Venkateswaran 2011) and our experimental results, it is considered that the suitable substrate concentration as the nitrogen source in the medium is 10 g L−1. The different weight ratios of CSL and YE that were used ranged from 10:0 to 0:10. Figure 2 showed that the biomass, total lipids and ARA yield obtained from the culture using 7 g L−1 CSL and 3 g L−1 YE or 3 g L−1 CSL and 7 g L−1 YE were all higher than those obtained from the other combination cultures. Therefore, the combination of 7 g L−1 CSL and 3 g L−1 YE as well as of 3 g L−1 CSL plus 7 g L−1 YE were chosen as the mixed nitrogen sources for the bioreactor culture.

DCW, total lipids and ARA yield in shaking culture using a different weight ratio of CSL and YE.  , DCW;
, DCW;  , Total lipids;
, Total lipids;  , ARA yield
, ARA yield
Effect of combined nitrogen sources on ARA production in the bioreactor culture
Considering there is the risk of cell autolysis in the cultures with high concentrations of CSL, the ratio of CSL to YE of 3:7 was chosen as the first combination to test in the bioreactor culture. It was found that M. alpina grew well without cell autolysis during the fermentation process and the mycelial morphology was also maintained in a fluffy pellet form throughout the culture period. Further details were shown in Fig. 3 and Table 2. The change curves of residual glucose also showed that almost all the glucose was exhausted when using CSL + YE culture (the bioreactor culture using 3 and 7 g L−1 YE as the combined nitrogen source), while a significant amount of glucose was still unconsumed at the end of fermentation in YE culture (the bioreactor culture using YE as the sole nitrogen source, Fig. 1). It is indicated that CSL enhanced the accessibility of glucose to cells in the culture contained CSL + YE. As the amount of glucose consumed increased, the biomass achieved the highest concentration of 37 g L−1 at 108 h, the increase of which is approximately 111 % higher than that obtained from YE culture. As previously mentioned, CSL plays a key role during the early fermentation period by providing a nitrogen source for fast cell growth, and YE plays a main role during the late fermentation period for keeping normal cell growth and ARA accumulation (Wynn et al. 2001). The total lipids and ARA concentration were also significantly increased by 167–189 % higher than that obtained from YE culture, respectively. The lipid content and ARA content in DCW were also individually improved to 43.2 and 21.1 %, which were higher than those contents in the YE culture, as shown in Table 2. It is revealed that the combined nitrogen source of the ratio of CSL to YE of 3:7 led to an obvious enhancement in cell growth, lipid and ARA production by M. alpina.

Profiles of glucose concentrations, DCW, total lipids and ARA yield in a 3.6-L bioreactor by M. alpina cultured in the medium using the combination of organic nitrogen sources containing 3 g L−1 corn steep liquor and 7 g L−1 yeast extract. Black square, glucose; black circle, DCW; black star, total lipids; black up-pointing triangle, ARA yield
CSL is a nitrogen source that contains abundance of soluble proteins micronutrients, both of which can be used rapidly by microorganisms to promote cell growth. YE would be accessed slowly. As a result, cells can be supplied nutrition continually, which in turn helps prevent cell autolysis, a phenomenon caused mainly by a C/N ratio imbalance. As presented in Figs. 1 and 3, the cell growth reached 22 g L−1 at 48 h when using CSL + YE, which was higher than the highest value of 17.5 g L−1 obtained at 96 h using YE only. Furthermore, at 48 h, 59 % of total biomass had been produced in CSL + YE culture, while only 40 % of total biomass had been obtained in YE culture. The time courses of the lipid and ARA accumulation show that the lipid and ARA reached their highest concentrations at 144 h in CSL + YE culture, which was 12 h earlier than when peak concentrations reached in YE culture. This result indicated that CSL + YE culture can shorten cultivation duration, which is favourable for large-scale production.
In the other culture, which had a CSL:YE ratio of 7:3, it was found that M. alpina grew well in the incipient stages of fermentation but the cells still underwent autolysis quickly on the 3rd day and onward (Figure omitted). It was concluded that the bioreactor provided a better environment for M. alpina growth than the culture flask. When the percentage of CSL was higher than YE in the medium, most of the nitrogen source was consumed rapidly and the C/N ratio was disturbed. Consequently, the cells underwent autolysis because of insufficient nutrition.
The present results were compared with those of previous studies by others, which are summarized in Table 3. To the best of the authors’ knowledge, during batch cultures, only one 7.5 L bioreactor cultivation of M. alpina R807 after 7 days was reported to produce a higher ARA yield (8.29 g L−1) than our work, which applied a novel membrane gas distributor to provide sufficient oxygen in the culture. Among fed-batch cultures, three studies obtained higher ARA concentration than the results reported here, typically by combining glucose feeding with other strategies to improve ARA production. These studies with fed-batch cultures provided us with a significant amount of information and insight regarding how to further enhance the culture of M. alpina in a bioreactor.
Conclusions
The experimental results indicate that YE and CSL are favourable for the morphology of M. alpina LU 166 to form the fluffy circular pellets, which is linked to high biomass and ARA yields. And the combined nitrogen source containing 3 g L−1 CSL and 7 g L−1 YE was found to be the most suitable composition to avoid cell autolysis and promote high cell density and ARA production during scale-up of the culture in the bioreactor. The corresponding biomass and ARA yield are 37.0 and 7.8 g L−1, respectively. In addition, extensive efforts, including research on the mechanism of morphology formation and on fed-batch culture strategies (including glucose and nitrogen source feeding), are in progress to facilitate the large-scale industrial ARA production.
References
Birch EE, Garfield S, Hoffman DR, Uauy R, Birch DG (2000) A randomized controlled trial of early dietary supply of long-chain polyunsaturated fatty acids and mental development in term infants. Dev Med Child Neurol 42:174–181
Dedyukhina EG, Chistyakova TI, Vainshtein MB (2011) Biosynthesis of arachidonic acid by micromycetes (review). Appl Biochem Microbiol 47:109–117
Eroshin VK, Satroutdinov AD, Dedykhina EG, Chistyakova TI (2000) Arachidonic acid production by Mortierella alpina with growth-coupled lipid synthesis. Process Biochem 35:1171–1175
Higashiyama K, Fujikawa S, Park EY, Shimizu S (2002) Production of arachidonic acid by Mortierella Fungi. Biotechnol Bioprocess Eng 7:252–262
Higashiyama K, Yaguchi T, Akimoto K, Shimizu S (2004) Media for culturing microorganisms and process for producing unsaturated fatty acids or lipids containing the same. US patent 6, 746, 857, 8 Jun 2004
Hwang BH, Kim JW, Park CY, Park CS, Kim YS, Ryu YW (2005) High-level production of arachidonic acid by fed-batch culture of Mortierella alpina using NH4OH as a nitrogen source and pH control. Biotechnol Lett 27:731–735
Jang HD, Lin YY, Yang SS (2005) Effect of culture media and conditions on polyunsaturated fatty acids production by Mortierella alpina. Bioresour Technol 96:1633–1644
Ji XJ, Zhang AH, Nie ZK, Wu WJ, Ren LJ, Huang H (2014) Efficient arachidonic acid-rich oil production by Mortierella alpina through a repeated fed-batch fermentation strategy. Bioresour Technol 170:356–360
Jin MJ, Huang H, Xiao AH, Zhang K, Liu X, Li S, Peng C (2008) A novel two-step fermentation process for improved arachidonic acid production by Mortierella alpina. Biotechnol Lett 30:1087–1091
Koike Y, Cai HJ, Higashiyama K, Fujikawa S, Park EY (2001) Effect of consumed carbon to nitrogen ratio on mycelia morphology and arachidonic acid production in cultures of Mortierella alpina. Biosci Bioeng 91:382–389
Lan WZ, Qin WM, Yu LJ (2002) Effect of glutamate on arachidonic acid production from Mortierella alpina. Lett Appl Microbiol 35:357–360
Li ZY, Lu Y, Yadward VB, Ward OP (1995) Process for production of arachidonic acid concentrate by a strain of Mortierella alpina. Can J Chem Eng 73:135–139
Li XY, Liu RJ, Li J, Chang M, Liu YF, Jin QZ, Wang XG (2015) Enhanced arachidonic acid production from Mortierella alpina combining atmospheric and room temperature plasma (ARTP) and diethyl sulfate treatments. Bioresour Technol 177:134–140
Lu JM, Peng C, Ji XJ, You JY, Cong LL, Ouyang PK, Huang H (2011) Fermentation characteristics of Mortierella alpina in response to different nitrogen sources. Appl Biochem Biotechnol 164:979–990
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31:426–428
Nie ZK, Ji XJ, Shang JS, Zhang AH, Ren LJ, Huang H (2014) Arachidonic acid-rich oil production by Mortierella alpina with different gas distributors. Bioprocess Biosyst Eng 37:1127–1132
Nisha A, Venkateswaran G (2011) Effect of culture variables on mycelia arachidonic acid production by M. alpina. Food Bioprocess Technol 4:232–240
Park EY, Koike Y, Cai HJ, Higashiyama K, Fujikawa S (2001) Morphological diversity of Mortierella alpina: effect of consumed carbon to nitrogen ratio in flask culture. Biotechnol Bioprocess Eng 6:161–166
Peng C, Huang H, Ji XJ, Liu X, You JY, Lu JM, Cong LL, Xu XK, Ouyang PK (2010) A temperature-shift strategy for efficient arachidonic acid fermentation by Mortierella alpina in batch culture. Biochem Eng J 10:101–104
Ratledge C (2004) Fatty acid biosynthesis in microorganisms being used for single cell oil production in cultures of Mortierella alpina. Biochimie 86:807–815
Sakuradani E, Hirano Y, Kamada N, Nojini M, Ogawa J, Shimizu S (2004) Improvement of arachidonic acid production by mutants with lower n-3 desaturation activity derived from Mortierella alpina 1S-4. App Microbiol Biotechnol 66:243–248
Shang L, Jiang M, Chang HN (2003) Poly(3-hydroxybutyrate) synthesis in fed-batch culture of Ralstonia eutropha with phosphate limitation under different glucose concentrations. Biotechnol Lett 25:1415–1419
Singh A, Ward OP (1997) Production of high yields of arachidonic acid in a fed-batch system by Mortierella alpina ATCC 32222. Appl Microbiol Biotechnol 48:1–5
Totani N, Hyodo K, Yamaguchi A, Ueda T (2000) A study on new nitrogen source for cultivation of genus Mortierella. J Jpn Oil Chem Soc 49:479–485
Vali SR, Sheng HY, Ju YH (2003) An efficient method for the purification of arachidonic acid from fungal single-cell oil (ARASCO). J Am Oil Chem Soc 7:725–730
Wynn JP, Hamid AA, Li Y, Ratledge C (2001) Biochemical events leading to the diversion of carbon into storage lipids in the oleaginous fungi Mucor circinelloides and Mortierella alpina. Microbiology 147:2857–2864
Zhu M, Yu LJ, Li W, Zhou PP, Li CY (2006) Optimization of arachidonic acid production by fed-batch culture of Mortierella alpina based on dynamic analysis. Enzyme Microb Technol 38:735–740
Authors’ contributions
XPL and SYZ carried out the main experiments. CXC and XTL helped in the cultivation of the strain and fatty acids assay. XPL, SYZ and YHL are involved in the drafting and revision of the manuscript. YHL has given final approval of the version to be published. All authors read and approved the final manuscript.
Acknowledgements
This work was funded by grants from the Science and Technology Program of Xiamen, China (No. 3502Z20153005), and from the National High Technology Research and Development Program of China (No. 2014AA021701).
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ling, Xp., Zeng, Sy., Chen, Cx. et al. Enhanced arachidonic acid production using a bioreactor culture of Mortierella alpina with a combined organic nitrogen source. Bioresour. Bioprocess. 3, 43 (2016). https://doi.org/10.1186/s40643-016-0121-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-016-0121-9