
Abstract
Background
The application of arbuscular mycorrhizal fungi (AMF) can induce the synthesis and accumulation of secondary metabolites in the tissues of host plants, thus impacting their allelopathic potential.
Materials and methods
The objective of this study was to determine the effect of three AMF species (Rhizoglomus intraradices, Funneliformis mosseae, Rhizoglomus fasciculatum) on photosynthetic pigments and secondary metabolites content in roots and reproductive organs of Ipomoea purpurea L., Digitaria sanguinalis L., and Solanum nigrum L. as a problematic weed species.
Results
Among compared weeds, the roots of D. sanguinalis associated with AMF accumulated the highest level of phenols. Higher content of flavonoids was obtained in roots of S. nigrum (7.46 mg g−1 FW) following colonization with R. intraradices. Berries of S. nigrum inoculated with R. intraradices had a higher concentration of terpenoids (21.45 mg 100 mL−1 of extract) than reproductive organs of D. sanguinalis and I. purpurea. Colonization with R. intraradices improved total phenolics in seeds of D. sanguinalis compared to the reproductive organs of other weeds. These compounds released from seeds help defend against pathogen infection, consequently increasing seed production. In addition, phenylalanine ammonia lyase enzyme activity in leaves of D. sanguinalis colonized by R. fasciculatum and F. mosseae was 55% and 67%, respectively, higher than I. purpurea plants, grown in the same condition.
Conclusions
Results suggest that AMF can play a crucial role in enhancing of secondary metabolites in these three weeds, thereby improving their allelopathic potential and competitive ability.
Graphic Abstract

Similar content being viewed by others


Background
Plants synthesize a broad and diverse assortment of natural products, the great majority of which do not appear to contribute directly in growth and development. These compounds have various functions such as in defence against herbivores and pathogens, attracting insects and protecting against UV light [1, 2]. In different plants, these compounds are synthesized and accumulated in roots, stems, leaves, fruits, and flowers. Most of these chemical compounds are accumulated in the vacuole, then polymerized or directly liberated, and eventually released to the environment, where they can act as allelopathic agents in the metabolism of neighbouring plants [3]. Allelochemicals are also released by weeds and inhibit growth and yield of crops [4].
Solanum nigrum L. (Solanaceae), or black nightshade, is a weed growing widely in worldwide. It mainly grows in tropical and temperate areas [5]. The berries of S. nigrum are revealed to have antiulcer, antioxidant, anti-inflammatory, antituberculosis, and antidiuretics effects [6]. Ipomoea purpurea L. Roth (Convolvulaceae), a troublesome weed of agronomic, horticultural and nursery crops, is often found in cotton, corn, and soybean fields. This weed is prolific, and can produce 8,000 seeds per season [7], which favours the infestation of fields by I. purpurea. Digitaria sanguinalis (L.) Scop. commonly known as crabgrass, is considered as an annual summer weed found in crops, turf, ruderal communities, and field margins in both tropical and temperature regions of the world [8]. Zhou et al. [9] identified three chemicals in the root exudates of D. sanguinalis, which may act as allelochemicals interfering with crop growth and affecting soil microbial communities.
Roots of D. sanguinalis, S. nigrum and I. purpurea can associate with arbuscular mycorrhizal fungi (AMF) present in soils [10, 11]. Previous studies indicated that AMF establishes mutualistic symbioses with flowering plants, ferns and bryophytes [12].
AMF can regulate chloroplast enzyme activity, decrease chlorophyll decomposition rate, accelerate the synthesis of important enzymes required for the chlorophyll peptide chain, promote chlorophyll synthesis, increase chlorophyll content plants, improve nutrient uptake, increasing the intensity of photosynthesis, which in turn reflects in increased biomass production [13]. In addition, some studies have also indicated that the AMF can increase the secondary metabolites content in plant organs such as seeds of Lallemantia iberica [14], fruits of Solanum lycopersicum L. [15] and leaves of Zea mays L. [12]. These chemical compounds also act as bioprotectants against pathogens and toxic stresses [16, 17]. For example, sesquiterpenes released from flowers of Arabidopsis thaliana defend plants against pathogen infection, reduce the cell death caused by the pathogen attack, and favour seed production [18]. The release of the flavonoids luteolin by seeds of Sesbania vesicaria can inhibit the growth of some edaphic fungi such as Pythium irregulare and Pythium ultimum [19].
Secondary metabolites in plants are usually accumulated in organs, tissues and structures critical for the survival of the plant itself (roots, functional leaves) and its offspring (flowers, seeds and fruits). As the allelochemicals in roots and reproductive organs of I. purpurea, D. sanguinalis and S. nigrum are unknown, the general objective of this study was to evaluate the impact of three AMF (Rhizoglomus intraradices, Rhizoglomus fasciculatum and Funneliformis mosseae), susceptible to establish a symbiotic association with those weeds, on the accumulation of secondary metabolites in roots and reproductive organs of those weed species.
Materials and methods
Study design
In this study, we used seeds of D. sanguinalis, S. nigrum and I. purpurea obtained from naturally infested fields at Zanjan University Research Farm, Zanjan, Iran (36°0.41'N, 48°23' E; altitude 1,634 m). The seeds were surface sterilized with sodium hypochlorite (10%) for 5 min and subsequently washed with distilled water and then placed in Petri dishes to germinate. After seven days, the germinated seeds were transplanted into (11 cm diameter *14 cm height) plastic pots (two seedlings per pot) containing 1.1 kg autoclaved soil (for one hour at 121 °C on three consecutive days) obtained from Research Farm of University of Zanjan, New Biotechnology Research Center, Zanjan, Iran (36°0.41′ N and 48°0.28′ E; altitude 1,620 m). Soil was sandy [20] with features as follows: pH of 7.8, EC (electrical conductivity) of 0.9–1 ds/m, 0.5% organic matter [21], 0.03% nitrogen [22], 1.56 mg/kg available P [23], and 33.25 mg/kg potassium [24]. Soil pH and EC were determined using pH and EC meters (Jenway 4310, Lancashire, UK). Within each weed species, there were four treatments: (a) non-inoculated control plants (without AMF); (b) plants inoculated with Rhizoglomus intraradices (N.C. Schenck & G. S. Sm.) Sieverd, G.A. Silva& Oehl comb. nov.), (c) plants inoculated with Rhizoglomus fasciculatum (Thaxt.) Sieverd, G.A. Silva & Oehl comb. nov., and (d) plants inoculated with Funneliformis mosseae (Nicol. and Gerd.) Walker & Schüβler comb. nov. For each treatment four replicates were considered. The experiment was carried out twice, and data were obtained from both experimental rounds. AMF inoculum added to each plant consisted of 20 g of soil containing colonized root fragments from Zea mays and 40 spores per gram. A filtrate containing the microorganisms accompanying AMF was added to non-AMF plants. The filtrate was prepared by passing diluted mycorrhizal inoculum through a layer of 15–20 μm filter paper with particle retention of 2.5 μm (Whatman 42; GE Healthcare, Little Chalfont, UK). The plants were placed in a greenhouse at temperature of 26–29 °C (day/night) with photosynthetic photon flux density (PPFD) of 500–600 μmol m−2 s−1 and 45% relative humidity. Plants were irrigated every day with 200–250 mL of distilled water to keep the soil moisture at 75% FC and received once a week 80–100 mL of complete Hoagland solution [25].
Colonization rate (%)
Root samples of weeds were cleared and stained [26] and mycorrhizal colonization was determined by examining 1-cm root segments (50 fragments from each plant) under the microscope.
Secondary metabolites
Total phenolic content in plant organs was determined using 1 mL of each sample, mixed with 1 ml of 95% ethanol, 4 mL of deionized water, 0.5 mL of Folin–Ciocalteu reagent, and 1 mL of 0.5% sodium carbonate. Mixtures were then placed in the dark for 60 min and afterward, the absorbance rate was measured at 725 nm. Gallic acid was used as the standard solution, where the concentrations of soluble phenolic compounds were expressed as mg g−1 FW [27].
Total flavonoid contents of weeds were determined by the aluminium chloride colorimetric method [28]. Briefly, 0.5 mL of extract was mixed with 0.3 mL of 5% NaNO2, 4.5 mL of deionized water and 600 μL of 10% AlCl3. After 6 min, the reaction was stopped by adding 2 mL of 1 M NaOH and 2 mL of deionized water. The absorbance of the samples was read at 510 nm. Flavonoids concentrations were expressed as mg g−1 FW, where quercetin was used as the standard.
Total terpenoid concentration was performed according to Ghorai et al. [29].
Phenylalanine ammonia lyase (PAL)
PAL activity was measured in fresh leaves (0.3 g). Enzyme was extracted with 2 mL of 50 mM boracic acid buffer (pH 8.8), containing 8 mM mercaptoethanol and 2% (w/v) PVPP. The homogenate was centrifuged at 14,000×g for 20 min at 4 °C. PAL assay was carried out according to the procedure of Zucker [30].
DPPH radical scavenging
DPPH reagent prepared in methanol (5 mg/100 mL, 2.0 mL) was added to each test sample (1.5 mL) and mixed with 0.5 mL of methanol. The mixture was allowed to stand for 30 min in the dark and absorbance was measured at 517 nm.
Scavenging activity was performed according to Barros et al. [31]:
where A0 and A1 are the absorbance rate of the control and test sample, respectively.
Photosynthetic pigments and chlorophyll content index
Chlorophylls (chl a and b) and carotenoids were extracted according to the method of Arnon [32] from 0.1 g of fresh leaves in 80% acetone. Absorbance at 470, 645, 663 nm was determined using a PerkinElmer-Lambda 25 USA Spectrophotometer. Chl a, b and carotenoids concentrations were calculated by applying the equations of Lichtenthaler [33]. Chlorophyll meter (CL-O1, Hansatech instruments) was used to estimate the chlorophyll content index (CCI) in the middle part of the leaf at the beginning of the reproductive stage in each plant.
Statistical analysis
Data were subjected to an analysis of variance (ANOVA) by using PROC GLM in SAS Software (Version 9.1, SAS Institute Inc., Cary, NC). The assumption of homogeneity of variance was tested before analysing the data. The significant differences were compared by Duncan’s multiple-range tests (P ≤ 0.05). The correlations between mycorrhizal colonization and secondary metabolites were tested with Pearson’s correlation coefficients.
Results
Mycorrhizal colonization
No mycorrhizal structures were found in roots of non-inoculated controls of any of the species (Fig. 1). Mycorrhizal colonization of weeds was significantly affected by the AMF species. The roots of S. nigrum and I. purpurea had higher colonization than D. sanguinalis plants. In D. sanguinalis, percentages of mycorrhizal colonization varied from 16 to 35% among different AMF species. Inoculation of I. purpurea with R. fasciculatum and R. intraradices increased colonization rate by 18.14 and 14.92%, respectively, as compared to F. mosseae (Fig. 1). In contrast, in S. nigrum F. mosseae and R. fasciculatum appeared as the most effective fungus for increasing the colonization rate.

AMF colonization rate (%) in roots of Solanum nigrum, Digitaria sanguinalis and Ipomoea purpurea at final harvest. Pots did not receive any mycorrhizal inoculum (control) or were inoculated with F. mosseae, R. fasciculatum or R. intraradices. Bars represent means (n = 4 plants) ± SE. Bars topped by the same letter indicate no significant differences between treatments at the 5% level using Duncan’s multiple-range test
Secondary metabolites
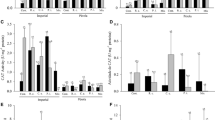
There was a significant effect of AMF on the levels of total soluble phenolic compounds and total flavonoids in the roots, and reproductive organs of the studied weeds (Fig. 2). R. intraradices induced the accumulation of total soluble phenolic compounds in both roots and fruits of S. nigrum and so did R. fasciculatum in fruits. The concentration of phenolic compounds in roots of R. intraradices-inoculated S. nigrum plants was 60.98% higher than that of non-AMF controls (Fig. 2). In D. sanguinalis roots were more influenced than seeds by AMF and all the tested AMF increased the concentration of total phenolic substances. I. purpurea also accumulated higher amount of phenolics after its inoculation with AMF, especially in their flowers. In the R. intraradices-inoculated plants, reproductive organs showed higher levels of phenolics than vegetative ones (roots). Colonization with F. mosseae, R. fasciculatum and R. intraradices improved phenolic compounds in flowers of I. purpurea (by 50%, 55.8% and 71%, respectively) compared to the respective non-AMF plants. However, this pattern changed after AMF inoculation, so that roots and seeds of D. sanguinalis had quite similar concentrations of these secondary metabolites and so did roots and fruits of S. nigrum associated with R. intraradices (Table 1).

Total soluble phenolic compounds (mg g−1 FW) and total flavonoids (mg g−1 FW) in different organs of Solanum nigrum, Digitaria sanguinalis and Ipomoea purpurea at final harvest. Pots did not receive any mycorrhizal inoculum (control) or were inoculated with F. mosseae, R. fasciculatum or R. intraradices. Bars represent means (n = 4 plants) ± SE. Bars topped by the same letter indicate no significant differences between treatments at the 5% level using Duncan’s multiple-range test
Roots of the three weed species had higher concentrations of flavonoids than reproductive organs (fruits, seeds or flowers) (Fig. 2). Colonization with F. mosseae and R. intraradices sharply promoted flavonoids in roots of S. nigrum (by 34% and 41%, respectively) compared to the respective control plants. Total flavonoids in roots of D. sanguinalis improved considerably when plants were associated with F. mosseae, so that D. sanguinalis plants colonized by R. intraradices species showed 25.43% more flavonoid content as compared to control plants. On the contrary, no difference in flavonoids was found in seeds of D. sanguinalis colonized by AMF and those collected from non-mycorrhizal plants. Colonization of I. purpurea with any of the three species of AMF used in this study promoted the accumulation of total flavonoids in roots and flowers. This increase is especially evident in the flowers of plants associated with R. intraradices (Fig. 2).
The levels of total terpenoids in the reproductive organs of weeds (fruits of S. nigrum, seeds of D. sanguinalis and flowers of I. purpurea) are shown in Fig. 3. The association of weeds with AMF significantly affected the levels of total terpenoids in S. nigrum and I. purpurea (Table 2). The highest amount of total terpenoids (21.45 mg 100 ml−1 of extract) was found in fruits of S. nigrum inoculated with R. intraradices. In I. purpurea, the greatest amount of terpenoids was measured in plants colonized by F. mosseae; the accumulation of these compounds in the flowers was 18% higher than in those of non-mycorrhized controls. In contrast, total terpenoids in seeds of D. sanguinalis were similar in plants colonized by F. mosseae or R. intraradices and in their respective non-mycorrhized control plants (Fig. 3).

Total terpenoids (mg 100 mL−1 of extract) in fruit of Solanum nigrum, seeds of Digitaria sanguinalis and flowers of Ipomoea purpurea, total carotenoids (mg g−1 FW) and phenylalanine ammonia lyase (PAL) (µmol cinnamic acid h −1 protein −1) in leaves of weeds at final harvest. Pots did not receive any mycorrhizal inoculum (control) or were inoculated with F. mosseae, R. fasciculatum or R. intraradices. Bars represent means (n = 4 plants) ± SE. Bars topped by the same letter indicate no significant differences between treatments at the 5% level using Duncan’s multiple-range test
Our results showed that the mycorrhizal association can significantly affect the concentrations of total carotenoids in leaves of the three studied weeds (Table 2). While R. intraradices induced the accumulation of carotenoids in S. nigrum, R. fasciculatum increased the concentrations of these pigments in leaves of D. sanguinalis and I. purpurea, suggesting that R. fasciculatum and R. intraradices are very efficient in improving total carotenoids in weeds. The content of total carotenoids in leaves of I. purpurea, which was unaffected by F. mosseae and R. intraradices, was significantly higher when associated with R. fasciculatum, so that I. purpurea inoculated with R. fasciculatum showed 16.81% more total carotenoids than its respective non-AMF control (Fig. 3).
The effect of AMF and weed species on PAL activity was significant (Table 2). In leaves of both D. sanguinalis and S. nigrum, the activity of the enzyme PAL clearly enhanced when plants were inoculated with either R. fasciculatum or F. mosseae. Likewise, the PAL activity in leaves of I. purpurea colonized by F. mosseae, R. fasciculatum or R. intraradices was, respectively, 61%, 69%, and 56% higher than that measured in non-AMF control plants. Contrariwise, PAL activity in leaves of D. sanguinalis inoculated with R. intraradices was 54% lower as compared to non-mycorrhizal control plants (Fig. 3).
Total antioxidant capacity differed between vegetative and reproductive organs in the studied weeds. In S. nigrum and D. sanguinalis DPPH activity was higher in the reproductive (fruits and seeds) than in the vegetative (root) organs. In contrast, roots of I. purpurea exhibited higher DPPH activity than flowers. Moreover, the DPPH radical scavenging activity in the organs of the three weed species was significantly affected by AMF. The three tested AMF increased the antioxidant capacity in the reproductive organs of S. nigrum and D. sanguinalis. DPPH activity in fruits of S. nigrum colonized by with F. mosseae, R. fasciculatum and R. intraradices AMF species was 6.89, 8.98 and 2.46%, respectively, higher as compared to non-AMF control plants. Similarly, R. intraradices and R. fasciculatum also improved the antioxidant activity in the flowers of I. purpurea, and so did R. fasciculatum and F. mosseae in roots of this weed (Fig. 4, Table 3).

DPPH radical scavenging activity (%) in different parts of Solanum nigrum, Digitaria sanguinalis and Ipomoea purpurea at final harvest. Pots did not receive any mycorrhizal inoculum (control) or were inoculated with F. mosseae, R. fasciculatum or R. intraradices. Bars represent means (n = 4 plants) ± SE. Bars topped by the same letter indicate no significant differences between treatments at the 5% level using Duncan’s multiple-range test
The effect of AMF on the concentrations of chlorophylls (a, b) in leaves depended on the weed species. While the highest concentrations of chlorophylls a and b were observed in S. nigrum associated with either R. intraradices or F. mosseae, AMF did not significantly affect the concentrations of chlorophyll a in D. sanguinalis and I. purpurea (Table 4). The chlorophyll index (SPAD) was increased by AMF colonization in the three species of weeds (Table 4).
Discussion
Unlike D. sanguinalis, mycorrhizal colonization of S. nigrum and I. purpurea plants with AMF species reached a high percentage (47–70%), making these weeds plants relatively stronger AMF hosts compared to D. sanguinalis. Since weeds are one of the major threats to the natural environment, the widespread occurrence of AMF and their important role in communities and ecosystems makes the interaction between weed and AMF key for the ecosystem functioning [34, 35].
Our results showed that roots and reproductive organs of the three investigated weeds inoculated with AMF were rich in total soluble phenolic compounds and flavonoids, substances with high allelopathic capacity, whose regulation and composition often differ below and above ground plant organs [36, 37]. Results of this study allow us to hypothesize that the secondary metabolites accumulated in the mycorrhized roots of weeds may be released into the soil through the external fungal mycelium and impact the roots of the surrounding plants [38]. Increased amounts of secondary metabolites in roots following mycorrhizal colonization may reinforce the allelopathic potential of weeds thus negatively affecting crops [39]. In forest ecosystems, chemical compounds released by invasive species can limit the growth of competing vegetation providing the invader competitive advantage [40, 41].
Some secondary metabolites belonging to the phenolics increase cell membrane permeability and induce lipid peroxidation, which finally results in plant death [42]. Earlier studies demonstrated that the increase of electrolyte leakage represents membrane integrity damage [43]. Among these phenolic compounds with allelopathic potential, catechin has been found in roots of the weed Centaurea stoebe and it has shown strong phytotoxicity against Festuca idahoensis and Arabidopsis thaliana [44]. Similarly, catechin has found in Melia azedarach fruit play an important role in its allelopathic potential [45]. In addition, other phenolic compounds including gallic acid, p-coumaric acid, p-hydroxybenzoic acid and ferulic acid, which cause reduced growth of rice, have been detected in the rhizosphere of Ageratum conyzoides L. [46]. Gmerek and Politycka [47] reported that the lipid peroxidation in roots of Z. mays L., Raphanus sativum L. and Pisum sativum L. was attributed to p-coumaric and ferulic acids.
Flavonoids are compounds with high antioxidant activity [48], whose biosynthesis can be inhibited by gibberellic acid [49]. They are known to inhibit the electron transport chain in the mitochondrial membrane [50]. These compounds suppress root growth, may be due to the breakage of cell homeostasis leading to allelopathic stress. Endogenous and exogenous flavonoids in various doses can affect auxin transport in roots and induce lateral root growth under stress conditions [51]. The allelopathic potential of Dittrichia viscosa L. may be attributed to flavonoids accumulated in its tissues [52]. Allelochemicals isolated from extracts of Xanthium strumarium L. exert inhibitory effects on growth of Gram-positive and Gram-negative bacterial strains and had antioxidant activity [53]. According to our results, the fruits of S. nigrum plants inoculated with AMF have high concentrations of phenolics and terpenoids and R. fasciculatum and R. intraradices are the most effective fungal species to improve phenolic compounds, terpenoids and DPPH. Consequently, we can infer that they can also exhibit increased antioxidant activity acting as free radical scavengers [56]. This higher accumulation of secondary metabolites in the fruits of S. nigrum can have practical applications for the phytotherapeutic industry.
A high concentration of secondary metabolites, such as phenols, flavonoids, anthocyanin and terpenoids, was also reported from S. nigrum and D. sanguinalis leaves [10], in addition, inoculation with F. mosseae species increased phenol, anthocyanin, and total terpenoid content in S. nigrum plants much more than D. sanguinalis.
The enhanced production of secondary metabolite concentrations in AM plants may be (1) due to improved mineral nutrition, and/or (2) a result of plant reaction to fungal colonization [54, 55]. Both of these mechanisms are possible explanations for the effect of AMF on the production of phenols, flavonoids and terpenoids in weeds in our study.
Carotenoids concentrations in leaves of D. sanguinalis and I. purpurea were improved by the association of these plants with R. fasciculatum. Carotenoids are known to be non-enzymatic antioxidant molecules that prevent the photo-oxidative damage of chlorophylls. In our study, the amount of carotenoids was enhanced in most part of the mycorrhizal plants in comparison with their respective non-mycorrhizal controls, which agrees with findings of Kumar et al. [57] working with Vigna radiata L.
PAL is a key enzyme in the biosynthesis of phenols, flavonoids and isoflavonoids in plants. Increased PAL activity in weeds associated with AMF may probably induce the production of flavonoids and other phenolic compounds production thus increasing their allelopathic potential. Altered gene expressions in hosts as a result of AMF colonization influence their metabolism and lead to the induction of chemical defence [58]. It was found that roots colonized by AMF had increased levels of transcripts encoding phenylalanine ammonia lyase (PAL). PAL is the first enzyme of the phenolics/phenylpropanoid pathway [59]. Since phenolic compounds are produced in weed as, D. sanguinalis, S. nigrum and I. purpurea, defence metabolites, the improved concentrations of these chemicals in weeds in our experiment might be explained by this mechanism.
Arbuscular mycorrhiza association promoted changes in chlorophyll concentration of the leaves of weeds. This result is likely due to improved nutrient uptake, resulting in overall higher photosynthetic capability [60]. Our results showed an increase in chlorophyll contents in S. nigrum associated with R. intraradices. Increased concentration of chlorophylls in Calendula officinalis associated with AMF was founded by Kheyri et al. [54]. A large amount of chlorophyll content in the leaves of AMF weeds, which allows plants to achieve more energy from light, could be related to enhanced uptake of phosphorus and magnesium increased transpiration, stomatal conductance, and carbon assimilation [61].
A significant positive relationship between mycorrhizal colonization and DPPH (0.832**) and terpenoids (0.853**) in fruits of S. nigrum was observed (Additional file 1: Table S2). In roots of D. sanguinalis, the root colonization is significantly correlated with total phenolics compounds (0.727**) and flavonoids (0.550*) and a positive correlation was observed between colonization rate and flavonoids and DPPH in seeds of this weed (Additional file 1: Tables S3, S4). Highly significant positive correlations were found between the mycorrhizal colonization and total phenolics, flavonoids and DPPH in roots and flowers of I. purpurea (Additional file 1: Tables S5, S6).
Conclusion
In conclusion, the application of AMF is a way of improving the contents of secondary metabolites, thereby increasing the allelopathic potential of these weeds. In between three AMF of species, R. intraradices had the highest effect in improving secondary metabolites in roots of S. nigrum. The higher production of secondary metabolites in the fruit of R. intraradices-inoculated S. nigrum can have practical applications in the phytotherapeutic industry. These results indicate that the establishment of AM symbiosis induces secondary metabolite accumulation, increases PAL enzyme activity, which may be of biological significance in the interactions of colonized plants with their environments.
Availability of data and materials
All the data are included in this manuscript.
Abbreviations
- AMF:
-
Arbuscular mycorrhizal fungi
- PAL:
-
Phenylalanine ammonia lyase
References
Rodríguez-Calzada T, Qian M, Strid Å, Neugart S, Schreiner M, Torres-Pacheco I, Guevara-González RG. Effect of UV-B radiation on morphology, phenolic compound production, gene expression, and subsequent drought stress responses in chili pepper (Capsicum annuum L.). Plant Physiol Biochem. 2019;1(134):94–102.
Berini JL, Brockman SA, Hegeman AD, Reich PB, Muthukrishnan R, Montgomery RA, Forester JD. Combinations of abiotic factors differentially alter production of plant secondary metabolites in five woody plant species in the boreal-temperate transition zone. Front Plant Sci. 2018;5(9):1257.
Durán AG, Chinchilla N, Molinillo JM, Macías FA. Structure-activity relationship studies on naphthoquinone analogs. The search for new herbicides based on natural products. Pest Manag Sci. 2019;75(9):2517–29.
Cheng F, Ali M, Liu C, Deng R, Cheng Z. Garlic allelochemical diallyl disulfide alleviates autotoxicity in the root exudates caused by long-term continuous cropping of tomato. J Agric Food Chem. 2020;68(42):11684–93.
Chauhan SH, Yadav S, Takahashi T, Łuczaj Ł, D’Cruz L, Okada K. Consumption patterns of wild edibles by the Vasavas: a case study from Gujarat, India. J Ethnobiol Ethnomed. 2018;14(1):1–20.
Wang L, Gao G, Bai Y, Luo W, Lin C, Jia Q. Fingerprint quality detection of Solanum nigrum using high-performance liquid chromatography–evaporative light scattering detection. Pharm Biol. 2011;49(6):595–601.
Chaney L, Baucom RS. The costs and benefits of tolerance to competition in Ipomoea purpurea, the common morning glory. Evol. 2014;68(6):1698–709.
Holm LG, Plucknett DL, Pancho JV, Herberger JP. The world’s worst weeds. Hawaii USA: Distribution and biology. University Press of Hawaii. Honolulu; 1977.
Zhou B, Kong CH, Wang P, Li YH. Chemical constituents of the essential oils of wild oat and crabgrass and their effects on the growth and allelochemical production of wheat. Weed Biol Manag. 2013;13(2):62–9.
Rashidi S, Yousefi AR, Pouryousef M, Goicoechea N. Total phenol, anthocyanin, and terpenoid content, photosynthetic rate, and nutrient uptake of Solanum nigrum L. and Digitaria sanguinalis L. as affected by arbuscular mycorrhizal fungi inoculation. Weed Biol Manag. 2020;20(3):95–108.
Sharda JN, Koide RT. Exploring the role of root anatomy in P-mediated control of colonization by arbuscular mycorrhizal fungi. Botany. 2010;88(2):165–73.
Begum N, Qin C, Ahanger MA, Raza S, Khan MI, Ashraf M, Ahmed N, Zhang L. Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front Plant Sci. 2019;10:1–15.
Eulenstein F, Tauschke M, Behrendt A, Monk J, Schindler U, Lana MA, Monk S. The application of mycorrhizal fungi and organic fertilisers in horticultural potting soils to improve water use efficiency of crops. Horticulturae. 2017;3(1):1–8.
Heydari S, Pirzad A. Efficiency of Funneliformis mosseae and Thiobacillus sp. on the secondary metabolites (essential oil, seed oil and mucilage) of Lallemantia iberica under salinity stress. J Hortic Sci Biotechnol. 2021;96(2):249–59.
Pasković I, Soldo B, Ban SG, Radić T, Lukić M, Urlić B, Mimica M, Bubola KB, Colla G, Rouphael Y, Major N. Fruit quality and volatile compound composition of processing tomato as affected by fertilisation practices and arbuscular mycorrhizal fungi application. Food Chem. 2021;359:129961.
Helgason T, Fitter A. The ecology and evolution of the arbuscular mycorrhizal fungi. Mycologist. 2005;19(3):96–101.
Ismail Y, McCormick S, Hijri M. A fungal symbiont of plant-roots modulates mycotoxin gene expression in the pathogen Fusarium sambucinum. PLoS ONE. 2011;6(3):e17990.
Huang M, Sanchez-Moreiras AM, Abel C, Sohrabi R, Lee S, Gershenzon J, Tholl D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defence against a bacterial pathogen. New Phytol. 2012;193(4):997–1008.
Hartwig UA, Joseph CM, Phillips DA. Flavonoids released naturally from alfalfa seeds enhance growth rate of Rhizobium meliloti. Plant Physiol. 1991;95(3):797–803.
Gee GW, Bauder JW. Particle-size Analysis. p 383–411 InA. Klute (ed.) Methods of Soil Analysis Part 1. Soil Science Society of America Book Series. 1986;5.
Nelson RE. Carbonate and gypsum.--p. 181–197. En: Methods of soil analysis: part 2; chemical and microbiological properties.--Winsconsin, US: American Society of Agronomy, 1986; 1982.
Bremner JM, Mulvaney CS. Nitrogen-total. In ‘Methods of soil analysis. Part 2. Chemical and microbiological properties.’(Ed. AL Page) pp. 1085–1121. Soil Science Society of America: Madison. 1982.
Olsen SR. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. US Department of Agriculture; 1954.
Helmke PA, Sparks DL. Lithium, sodium, potassium, rubidium, and cesium. Methods of soil analysis: Part 3 chemical methods. 1996;5:551–74.
Arnon DI, Hoagland DR. A comparison of water culture and soil as media for crop production. Science. 1939;89(2318):512–4.
Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc. 1970;55(1):158.
Waterman PG, Mole S. Analysis of phenolic plant metabolites. Oxford: Blackwell Scientific; 1994.
Miguel MG, Nunes S, Dandlen SA, Cavaco AM, Antunes MD. Phenols and antioxidant activity of hydro-alcoholic extracts of propolis from Algarve South of Portugal. Food Chem Toxicol. 2010;48(12):3418–23.
Ghorai N, Chakraborty S, Gucchait S, Saha SK, Biswas S. Estimation of total Terpenoids concentration in plant tissues using a monoterpene, Linalool as standard reagent. Protocol exchange. 2012. 005.
Zucker M. Induction of phenylalanine deaminase by light and its relation to chlorogenic acid synthesis in potato tuber tissue. Plant Physiol. 1965;40(5):779–84.
Barros L, Baptista P, Ferreira IC. Effect of Lactarius piperatus fruiting body maturity stage on antioxidant activity measured by several biochemical assays. Food Chem Toxicol. 2007;45(9):1731–7.
Arnon AN. Method of extraction of chlorophyll in the plants. Agron J. 1967;23(1):112–21.
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth Enzymol. 1987;148(34):350–82.
Rinaudo V, Bàrberi P, Giovannetti M, Van Der Heijden MG. Mycorrhizal fungi suppress aggressive agricultural weeds. Plant Soil. 2010;333(1):7–20.
Rashidi S, Yousefi AR, Pouryousef M, Goicoechea N. Mycorrhizal impact on competitive relationships and yield parameters in Phaseolus vulgaris L.—weed mixtures. Mycorrhiza. 2021;31(5):599–612.
Hartmann T. From waste products to ecochemicals: fifty years research of plant secondary metabolism. Phytochemistry. 2007;68(22–24):2831–46.
Johnson SN, Erb M, Hartley SE. Roots under attack: contrasting plant responses to below-and aboveground insect herbivory. New Phytol. 2016;210(2):413–8.
Blum U. Allelopathy: A soil system perspective. In: Reigosa MJ, Pedrol N, González L, editors. Allelopathy: a physiological process with ecological implications. Springer; 2006. p. 299–340.
Putnam AR. Weed allelopathy. In: Duke SO, editor. Weed Physiology, vol. I. Reproduction and Ecophysiology. Boca Raton, Florida: CRC Press; 1985, p. 131–155.
Callaway RM, Aschehoug ET. Invasive plants versus their new and old neighbors: a mechanism for exotic invasion. Science. 2000;290(5491):521–3.
Ridenour WM, Callaway RM. The relative importance of allelopathy in interference: the effects of an invasive weed on a native bunchgrass. Oecologia. 2001;126(3):444–50.
Li Y, Han D, Hu G, Sommerfeld M, Hu Q. Inhibition of starch synthesis results in overproduction of lipids in Chlamydomonas reinhardtii. Biotechnol Bioeng. 2010;107(2):258–68.
Poonpaiboonpipat T, Pangnakorn U, Suvunnamek U, Teerarak M, Charoenying P, Laosinwattana C. Phytotoxic effects of essential oil from Cymbopogon citratus and its physiological mechanisms on barnyardgrass (Echinochloa crus-galli). Ind Crops Prod. 2013;41:403–7.
Bais HP, Kaushik S. Catechin secretion & phytotoxicity: fact not fiction. Commun Integr Biol. 2010;3(5):468–70.
Chiffelle I, Huerta Fuentes A, Lizana RD. Physical and chemical characterization of Melia azedarach L. fruit and leaf for use as botanical insecticide. Chil J Agric Res. 2009;69:38–45.
Batish DR, Kaur S, Singh HP, Kohli RK. Role of root-mediated interactions in phytotoxic interference of Ageratum conyzoides with rice (Oryza sativa). Flora. 2009;204(5):388–95.
Gmerek J, Politycka B. Response of maize, pea and radish roots to allelochemical stress. Acta Biol Crac Ser Bot. 2011;53(1):32–7.
AitChaouche FS, Mouhouche F, Hazzit M. Antioxidant capacity and total phenol and flavonoid contents of Teucrium polium L. grown in Algeria. Med J Nutrition Metab. 2018;11(2):135–44.
Hinderer W, Petersen M, Seitz HU. Inhibition of flavonoid biosynthesis by gibberellic acid in cell suspension cultures of Daucus carota L. Planta. 1984;160(6):544–9.
Smith CO, Nehrke K, Brookes PS. The Slo (w) path to identifying the mitochondrial channels responsible for ischemic protection. Biochem J. 2017;474(12):2067–94.
Maloney GS, DiNapoli KT, Muday GK. The anthocyanin reduced tomato mutant demonstrates the role of flavonols in tomato lateral root and root hair development. Plant Physiol. 2014;166(2):614–31.
Grande M, Piera F, Cuenca A, Torres P, Bellido IS. Flavonoids from Inula viscosa. Planta Med. 1985;51(05):414–9.
Ghahari S, Alinezhad H, Nematzadeh GA, Tajbakhsh M, Baharfar R. Biochemical composition, Antioxidant and biological activities of the essential oil and fruit extract of Xanthium strumarium Linn. From Northern Iran. J Agric Sci Technol. 2017;19(7):1603–16.
Kheyri Z, Moghaddam M, Farhadi N. Inoculation efficiency of different mycorrhizal species on growth, nutrient uptake, and antioxidant capacity of Calendula officinalis L.: a Comparative Study. J Soil Sci Plant Nutr. 2022;6(20):1–13.
Sbrana C, Avio L, Giovannetti M. Beneficial mycorrhizal symbionts affecting the production of health-promoting phytochemicals. Electrophoresis. 2014;35(11):1535–46.
Rice-Evans CA, Miller NJ, Paganga G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med. 1996;20(7):933–56.
Kumar A, Kumar Yadav D, Ulla R. Role of arbuscular mycorrhizal fungi on plant growth and photosynthetic pigments in Vigna radiata. Mycopath. 2021;18:2.
Al-Arjani AB, Hashem A, Abd-Allah EF. Arbuscular mycorrhizal fungi modulates dynamics tolerance expression to mitigate drought stress in Ephedra foliata Boiss Saudi. J Biol Sci. 2020;27(1):380–94.
Yang YH, Wang CJ, Li RF, Zhang ZY, Yang H, Chu CY, Li JT. Overexpression of RgPAL family genes involved in phenolic biosynthesis promotes the replanting disease development in Rehmannia glutinosa. J Plant Physiol. 2021;257:153339.
de Souza Campos PM, Borie F, Cornejo P, Meier S, López-Ráez JA, López-Garcia Á, Seguel A. Wheat root trait plasticity, nutrient acquisition and growth responses are dependent on specific arbuscular mycorrhizal fungus and plant genotype interactions. J Plant Physiol. 2021;1(256):153297.
Zai XM, Zhu SN, Qin P, Wang XY, Che L, Luo FX. Effect of Glomus mosseae on chlorophyll content, chlorophyll fluorescence parameters, and chloroplast ultrastructure of beach plum (Prunus maritima) under NaCl stress. Photosynthetica. 2012;50(3):323–8.
Acknowledgements
Not applicable.
Funding
This research was funded by the University of Zanjan, Iran.
Author information
Authors and Affiliations
Contributions
ARY: methodology, software, conceptualization, supervision, writing and editing. MP, NG: software, writing and editing. SR: investigation, methodology, software, writing—original draft. ARY, MP: funding acquisition. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All authors contributed in design and preparation of the research, and they have read the final version of the manuscript.
Consent for publication
We declare our agreement.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Additional tables.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Rashidi, S., Yousefi, A.R., Pouryousef, M. et al. Effect of arbuscular mycorrhizal fungi on the accumulation of secondary metabolites in roots and reproductive organs of Solanum nigrum, Digitaria sanguinalis and Ipomoea purpurea. Chem. Biol. Technol. Agric. 9, 23 (2022). https://doi.org/10.1186/s40538-022-00288-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-022-00288-1