
Abstract
Background
Spatial behavior, including home-ranging behaviors, habitat selection, and movement, can be extremely informative in estimating how animals respond to landscape heterogeneity. Responses in these spatial behaviors to features such as human land modification and resources can highlight a species’ spatial strategy to maximize fitness and minimize mortality. These strategies can vary on spatial, temporal, and individual scales, and the combination of behaviors on these scales can lead to very different strategies among species.
Methods
Harnessing the variation present at these scales, we characterized how species may respond to stimuli in their environments ranging from broad- to fine-scale spatial responses to human modification in their environment. Using 15 bobcat-years and 31 coyote-years of GPS data from individuals inhabiting a landscape encompassing a range of human land modification, we evaluated the complexity of both species’ responses to human modification on the landscape through their home range size, habitat selection, and functional response behaviors, accounting for annual, seasonal, and diel variation.
Results
Bobcats and coyotes used different strategies in response to human modification in their home ranges, with bobcats broadly expanding their home range with increases in human modification and displaying temporal consistency in functional response in habitat selection across both season and time of day. Meanwhile, coyotes did not expand their home ranges with increased human modification, but instead demonstrated fine-scale responses to human modification with habitat selection strategies that sometimes varied by time of day and season, paired with functional responses in selection behaviors.
Conclusions
These differences in response to habitat, resources, and human modification between the two species highlighted the variation in spatial behaviors animals can use to exist in anthropogenic environments. Categorizing animal spatial behavior based on these spatiotemporal responses and individual variation can help in predicting how a species will respond to future change based on their current spatial behavior.
Similar content being viewed by others


Background
Spatial behavior can be informative about how animals respond to the heterogeneity in their environment. These responses include broad-scale decisions about the size and location of the home range [10], as well as finer scale responses to heterogeneity in the environment through habitat selection decisions [34]. Animals often vary in their spatial behavior, which can be due in part to individual personality [35, 68] or behavioral plasticity [81, 83]. This variation can lead to the degree of selection of resources being dependent on the availability of resources in their environment, known as functional responses [25, 56], which can include responses to habitat [59], food resources [28, 82], or other stimuli. Functional responses in spatial behavior relative to anthropogenic features and activity have been documented in various species, specifically for habitat selection and proximity to humans in wolves (Canis lupus) [25], caribou (Rangifer tarandus caribou) [53], and moose (Alces alces) [7]. Habitat selection is particularly informative about a species' ecology because of the direct link between the spatial choices an individual makes and variability in the environment [33], and because it can have direct impact on reproduction and survival [52, 62]. Habitat selection and functional responses are especially important in the context of anthropogenic change, through which landscape composition is constantly undergoing modification.
Animals can display a wide range of responses to anthropogenic features. These responses can be broad-scale, such as consistently avoiding human activity or structures in their home range [40, 54], and which could lead to increased home range size for individuals inhabiting areas with greater levels of human land modification [63]. In contrast, species can also display fine-scale, nuanced responses to humans, such as only avoiding anthropogenic features within the home range and during a certain periods of the day [86], seasonally [32], or by using a combination of spatiotemporal responses [37]. These broad- and fine-scale behaviors do not only refer to the spatial scale (e.g., home range vs. landscape level) of the response, but also to the amount of spatial, temporal, and individual variation and complexity in their space use behaviors in response to a stimulus in their environment. Investigating individual and temporal variation in spatial behavior can elucidate broader patterns in behavior, linking spatial ecology and animal behavior [27], as well as help draw conclusions about population-level relationships with habitat [5]. Here, we propose that animals can vary in their behaviors on spatial, temporal, and individual axes at multiple scales in response to features on the landscape. These behaviors can be used to characterize animal responses to anthropogenic modification in their environment. Additionally, these responses can be particularly useful for wide-ranging species which use a variety of habitats with varying levels of human development.
Bobcats (Lynx rufus) and coyotes (Canis latrans) are two carnivores that fill the role of top predator in the absence of large predators throughout much of North America [38, 42, 76]. Bobcats are strictly carnivorous and are thought to prefer habitats with ample cover, such as forest when available [42, 44]. Coyotes are more generalist in both diet and habitat and are found in all habitats along a forested-to-agricultural gradient [42, 67], and are more likely than bobcats to exploit agricultural landscapes [44, 61]. Both bobcats and coyotes have the potential to maintain larger home ranges in the presence of fragmentation, but coyotes are more plastic and adaptable to anthropogenic change, exploiting small resource patches on a landscape scale, regardless of connection [1, 73, 87]. Although small carnivores are a group expected to adapt better than other taxa to human development [13, 84], carnivore species vary in their ability to coexist with humans based on flexibility in diet and suitable habitat, as well as plasticity in behaviors like boldness and neophilia, leading to a variety of responses to anthropogenic land modification [50, 69].
Here we studied how variation in anthropogenic land modification shapes spatial behaviors of two carnivore species and characterized the scale and complexity at which they respond to human modification. Specifically, we investigated how a gradient of human land modification impacted home range size and habitat selection of both species. We also evaluated how habitat selection behaviors varied temporally by season and time of day and how individual variation in these behaviors could be linked to variation to the intensity of human modification for an individual (functional response). Overall, given the behavior of both carnivores, we expected bobcat responses to be marked by stronger and more consistent avoidance of human modification and overall larger home range when exposed to human modification due to their tendency to be more avoidant of human impact than coyotes. Meanwhile, we expected coyote responses to be more fine-scale, with home range size less affected by human modification, but with space use showing more individual variation, temporally-dependent selection behaviors, and complexity in their functional responses to human modification.
Methods
Study area
Our study occurred at two sites in Illinois. The southern Illinois study site consists of Touch of Nature Environmental Center (37.62762, − 89.15827) and Giant City State Park (37.60195, − 89.18925), making up a combined 28.6 km2 of land managed by Southern Illinois University and the state of Illinois. The landscape at the southern study site is largely forested (Jackson and Union Counties, Illinois, consist of 40% agriculture and 41% forest), with an average annual temperature of 14.1 °C and an average annual precipitation of 118 cm [58]. The central Illinois study site consists of properties managed by the state of Illinois and U.S. Army Corps of Engineers surrounding Lake Shelbyville (39.51856, − 88.70658). The landscape is dominated by row crop corn and soybean agriculture, with some lakeshore, small towns, and remnant forested patches (Moultrie and Shelby Counties, Illinois, consist of 79% agriculture and 12% forest). The central Illinois study site has an average annual temperature of 12.2 °C and average annual precipitation of 120 cm [58].
Capture and Handling
Bobcats and coyotes in both study sites were captured using cage traps (Tomahawk Live Trap, Hazelhurst, Wisconsin, Model 209.5, and homemade traps with similar dimensions, [6]) and rubber-padded foothold traps (Minnesota Trapline Products, Pennock, Minnesota, MB-650-RJ, [80]) during four winter capture seasons from January 2018 through March 2021. Bobcats were chemically immobilized with ketamine and xylazine and recovered inside a cage trap before release (ZooPharm, [6]). Coyotes were chemically immobilized with BAM™ (butorphanol tartrate, azaperone, and medetomidine hydrochloride) and the BAM™ was reversed post-handling with naltrexone and atipamezole before release (ZooPharm, [11]). All captured animals were fitted with LiteTrack Iridium 250 GPS collars (250 g, Lotek Wireless, Newmarket, Ontario, Canada) equipped with a release mechanism to drop off after one year. Thirty-two collars recorded GPS locations once every 1.5 h and 14 collars had a different schedule due to the GPS fix rate requirements of various project goals (1, 2, 3, or 4 h).
Spatial covariates
Several spatial covariates were used in seasonal delineation and resource selection analyses. Landcover covariates were sourced from a 30 m resolution National Land Cover Database classification [88] and reclassified to combine them into six landcover categories (water, exurban [denotes development], grassland and scrub, forest, agriculture, and wetland) for seasonal delineation (see below for seasonal delineation details) and four landcover categories (exurban, forest, agriculture, and other) for resource selection analyses. Grassland, scrub, and wetland were rare and so were combined with the other wetland category to avoid including resources not used by some individuals. We also included a separate human land modification covariate using a global layer which quantifies the degree of human land modification on a continuous scale, using 13 anthropogenic global stressors under the categories of human settlement, agriculture, transportation, mining and energy production, and electrical infrastructure at a 1 km resolution (see [36] for full details on this layer). We reprocessed a layer of Illinois streams and shorelines [29] to create a Euclidean distance to water covariate at 30 m resolution and took the natural logarithm of the Euclidean distances to account for decreasing impact of a water source with increasing distance from it [41]. Similarly, we reprocessed a layer of Illinois paved roads [30] to create a natural logarithm of the Euclidean distance to road covariate at 30 m resolution.
Home range size
To estimate the annual home ranges of bobcats and coyotes, we used autocorrelated kernel density estimation (AKDE), as developed by Fleming et al. [14]. We used the package ‘ctmm’ in Program R [12] to estimate home ranges. Home range sizes were calculated using the Ornstein–Uhlenbeck with foraging (OUF) model using a 0.95 quantile. Data from 11 individuals did not meet the requirements for AKDE and were excluded from home range size analyses, commonly due to instances of dispersal or short duration of tracking.
After log-transforming the home range sizes, we used a two-sample t-test to identify between-species differences in annual home range size [39, 41]. We performed similar tests between bobcat and coyote study sites and sexes, but due to uneven sample sizes between sex and study site within species, information on home range size comparisons by demographics can be found in Supporting Information. We used univariate linear models to evaluate how the degree of human modification, described above [36], in each home range impacted home range size in both species. We calculated the degree of human modification in each home range by averaging the human modification values within each home range and converting it from a 0 to 1 scale to a percentage. Intercept-only, linear, and quadratic regressions were performed and compared using the Akaike Information Criterion with correction for small sample size (AICC) to determine the top regression model [9].
Temporal period delineation
We used a clustering algorithm to define seasons ecologically in Program R [2]. To define bobcat and coyote seasons, the movement speed and turning angle between successive locations were calculated. Using a moving window of time, we calculated the mean speed and tortuosity, as well as the proportion of the locations in water, exurban, grassland and scrub, forest, agriculture, and wetland landcover areas within the moving window [88]. The DD-weighted gap method [92] was used to determine the optimal number of clusters (seasons). We then used K-means clustering analysis [24, 47] to identify clusters of similar space use behaviors to define seasons, adjusting bootstrap thresholds and windows of seasonal length to ensure continuous seasons of adequate length (longer than two weeks).
Day, night, and crepuscular diel periods were also delineated (diel period details in Supporting Information). Equinox and solstice dates [57] were used to divide the year into four periods, and the average sunrise and sunset time for each period was calculated [49] to account for changes in day length between the four periods [85]. Day was delineated as two hours after sunrise to one hour before sunset, night as two hours after sunset to one hour before sunrise, and crepuscular as the two lengths of time one hour before to two hours after sunrise and sunset [16, 31, 85]. The four species-specific seasons and three diel periods (day, night, crepuscular) were combined to create twelve bobcat temporal periods and twelve coyote temporal periods.
Resource selection functions
To determine individual-level habitat selection, we used a logistic regression to estimate resource selection functions (RSFs) for each individual within the annual home ranges [5, 48]. The individuals that were previously excluded from home range size analysis were included in the RSFs, using kernel density estimation (KDE) annual home range estimations (instead of AKDE) [91]. Twelve thousand random locations were generated within each of these home ranges. Each random location was also assigned a random date and time sampled from the dates and times of the used points [3], and the previously described temporal periods were applied to each used and random point so that each point was categorized based on its season and diel period. Landcover categories, human modification, distance to water, and distance to road covariates were extracted for each used and random point. Landcover categories included the dummy variables of forest (reference category), agricultural, exurban, and other. The continuous variables of human modification, distance to water, and distance to road covariates were scaled (divided by one standard deviation) so they could be easily compared [77].
Bobcat and coyote RSFs were estimated using the package ‘IndRSA’ in Program R [5]. ‘IndRSA’ estimates an individual-level RSF for each individual and a population average in a second step [55]. Models were estimated for each permutation of species and temporal period. K-fold cross-validations were performed for each output (k = 5), and individuals with a k-fold value less than 0.2 were excluded from the results.
Impacts of human modification on carnivore behavior
We estimated how human modification directionally affects bobcat and coyote habitat selection behavior in the form of functional responses in habitat selection [25, 53]. We used univariate regressions with the individual RSF coefficients for five covariates (agriculture, exurban, and other landcover; distance to water; and distance to road) as the response variables and the degree of human modification in each home range as the explanatory variables. These regressions were separated based on temporal period to discern temporal effects on the functional responses. In addition, regressions of the means of coefficients for each covariate during all temporal periods for each degree of human modification in home range (the mean selection of each covariate for each individual) were performed to find if a broad functional response was present regardless of temporal period. Weighted regressions were used to account for uncertainty associated with the RSF coefficients [4]. Intercept-only, linear, and quadratic regressions were compared using AICC to determine the top regression model [9]. The “other” landcover category lacked biological meaning and so was not included in the results but can be found in the Supporting Information.
Results
Fifteen bobcat-years (central Illinois female n = 1, central Illinois male n = 3, southern Illinois female n = 6, southern Illinois male n = 5) and 31 coyote-years (central Illinois female n = 6, central Illinois male n = 17, southern Illinois female n = 5, southern Illinois male n = 3) of location data were collected. An average of 1,397 GPS locations were obtained from each bobcat (range 293–2,695) and an average of 1,736 locations were obtained from each coyote (range 213–3,596), dependent on the lifespan of the animal and the battery life of the collar (maximum one year). Details regarding data demographics, GPS fixes, and landcover category use for each animal can be found in the Supporting Information, Appendix 1. Fourteen bobcat-years and 28 coyote-years of GPS data where the individuals were tracked for over two months were used to calculate four bobcat and four coyote seasons. Bobcats had short, distinct seasons in fall, early winter, and late winter, but had one long season during spring and summer (season details in Supporting Information, Appendix 2). Coyotes had short seasons during early and late winter and two longer spring and summer/fall seasons.
Home range size
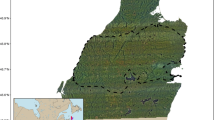
Thirty-five annual home ranges were estimated using AKDE (Fig. 1). Bobcat mean home range size was 32.3 km2 (central Illinois male n = 2, southern Illinois female n = 6, southern Illinois male n = 3, range 2.5–132.0 km2) and coyote mean home range size was 221.4 km2 (central Illinois female n = 5, central Illinois male n = 13, southern Illinois female n = 5, southern Illinois male n = 1, range 7.1–849.0 km2) (details on home range size uncertainty in Supporting Information, Appendix 3). Bobcat home ranges were significantly smaller than those of coyotes (bobcat μ = 32.3 km2, coyote μ = 221.4 km2, t = − 2.801, DF = 33, p = 0.009).

Population-level bobcat (orange) and coyote (purple) home range size estimations using AKDE. Asterisk indicates significant difference in home range sizes (α = 0.05)
The home range size of individual bobcats had a positive linear relationship with the degree of human modification within their home ranges (R2 = 0.66), with increased human modification being correlated with larger home ranges (Fig. 2). The intercept-only model was the top model for coyotes, indicating we found no relationship between the degree of human modification within home ranges and home range size (AICC table in Supporting Information, Appendix 4).

Individual bobcat and coyote home range size estimations paired with the degree of human modification within the home ranges. Bobcat home range size had a linear relationship with human modification (trendline and confidence interval shadow shown), while coyote home range size had no relationship with human modification (with no trendline to display)
Habitat selection
No individual bobcats were tracked during each diel period for the entire duration of the early winter season, so that season was excluded from the bobcat RSF results. Bobcats avoided agriculture during the spring/summer season and late winter night, weakly avoided (confidence intervals overlapping 0) it during late winter day and crepuscular periods, weakly selected it during fall crepuscular and night periods, and selected it during fall day (n = 15, k-fold mean = 0.62, range 0.28–0.92) (Fig. 3). Bobcats generally did not respond to exurban habitat or human modification. Bobcats tended to select areas farther from roads and closer to water in periods when they had any response. The "other" landcover category lacks biological meaning, so from here onward we report the coefficients but will not interpret the meaning.

Population-level bobcat RSF coefficients of agricultural landcover, exurban landcover, other landcover, human modification, distance to water, and distance to road covariates in reference to forested landcover with 95% confidence interval bars. The results are divided by bobcat seasons (late winter, spring/summer, fall) and diel period (day, night, crepuscular) for coefficients representing nine temporal periods
Coyotes generally avoided agriculture regardless of season, but the strength of avoidance varied by temporal period; avoidance decreased at night regardless of season (sometimes resulting in positive selection) (n = 31, k-fold mean = 0.67, range 0.20–0.96) (Fig. 4). Coyotes generally avoided exurban habitat, with weak avoidance at night and stronger avoidance during the day and crepuscular periods, regardless of season. Coyote avoidance of most landcover categories during most temporal periods indicated they mainly preferred forest over alternative habitat types. Coyotes generally did not select for or avoid human modification. They generally selected areas closer to water and weakly selected areas farther from roads during most temporal periods.

Population-level coyote RSF coefficients of agricultural landcover, exurban landcover, other landcover, human modification, distance to water, and distance to road covariates in reference to forested landcover with 95% confidence interval bars. The results are divided by coyote seasons (early winter, late winter, spring, summer/fall) and diel period (day, night, crepuscular) for coefficients representing twelve temporal periods
Impacts of human land modification on carnivore behavior
We investigated whether there were functional responses between carnivore habitat selection and the degree of human modification in their home range (mean selection coefficients regardless of temporal period, hereafter referred to as “mean”), in addition to separating out selection by temporal period to gain insight about whether there was a temporal aspect to functional responses (Fig. 5). Bobcats exposed to higher degrees of human modification in their home ranges selected for more agriculture. There was a quadratic relationship in the spring/summer day period and a positive linear relationship in the spring/summer crepuscular and night temporal periods. Exurban selection and human modification exhibited a negative linear relationship during the spring/summer night, indicating that bobcats avoided exurban habitat with increased human modification during that period. Bobcats also exhibited negative linear relationships of human modification and distance to water in the mean and the spring/summer day and crepuscular periods, indicating selection to areas near water with increased human modification. Distance to road regressions yielded intercept-only top models for all temporal periods and means for bobcats, indicating no relationship between intensity of human modification and distance to road (AICC table in Supporting Information, Appendix 5).

Top regression model trendlines for bobcat (A–C) and coyote (D–G) functional responses. Individual-level RSF coefficients for agricultural landcover, exurban landcover, distance to water, and distance to roads were regressed against the response to the degree of human modification present in each individual’s home range. Trendlines are displayed when the relationship was linear or quadratic and are not displayed if the relationship was intercept-only based on AICC model selection (see Supporting Information, Appendix 6). Trendlines are displayed for specific temporal periods (solid) or mean of all temporal periods (bolded and dashed). R2 values are displayed for each regression
Coyote selection for agriculture varied in response to the degree of human modification. Agriculture selection had a positive linear relationship in the early winter day, spring crepuscular, and summer/fall night periods and a negative linear relationship in the spring day and night periods with increasing human modification, indicating coyote functional responses may be dependent on season and/or time of day (Fig. 5). There was a positive linear mean relationship between coyote selection for exurban habitat and human modification, a negative linear trend in the spring day period, and a quadratic relationship in the late winter crepuscular period. Coyote selection had a mean quadratic relationship between human modification and distance to water, as well as slightly negative linear trends in spring day, crepuscular, and night periods. There was a strong negative linear trend in the summer/fall crepuscular period, indicating that, in general, coyotes selected areas farther from water as human modification increased. Coyotes displayed a quadratic relationship between distance to roads and human modification only in the mean, but not in any specific temporal periods.
Discussion
Using a gradient of human modification within carnivore home ranges in two study sites, we aimed to better understand how landcover types and intensity of human modification affect carnivore behaviors both spatially and temporally. As hypothesized, we found notable differences between bobcats and coyotes in the degree of complexity in their responses to human modification. While differences in wildlife species’ response to human activity has been studied before (e.g., [17]), our work characterized specific responses to anthropogenic disturbances based on several behaviors across spatial and temporal contexts. Bobcat responses included larger home ranges with increased human modification within them in addition to weak habitat selection responses to agriculture, exurban areas, and human modification. Bobcats also displayed functional responses in their habitat selection choices that were relatively temporally consistent. In contrast, coyote home range size did not increase with human modification, but instead they displayed complex responses including stronger avoidance of agriculture, exurban areas, and human modification than bobcats, indicating more fine-scale avoidance behaviors within the home range. Coyote habitat selection functional responses were more nuanced, temporally-dependent, and sometimes changed direction depending on the amount of human modification (quadradic relationships). Our work provides evidence that species inhabiting the same landscape, and even filling a similar trophic role as top predators in this system, can vary widely in the degree of scale and complexity of their behavioral response to their environment. Specifically, our work shows the importance of investigating spatial and temporal variation in habitat selection and functional responses to better understand the complexity in how extrinsic factors shape wildlife behavior.
Carnivore spatial behavior and response to human modification
Overall, higher degrees of human modification within home range were correlated with larger home range size in bobcats. Bobcats tend to use larger home ranges in more fragmented and developed landscapes [73, 87] and lynx (Lynx lynx) have been found to expand their home ranges to increase hunting efforts in areas with declining prey abundance [78]. Therefore, the fragmentated, patchy landscape and increased human modification in the central Illinois site could be leading to low-quality resources for bobcats, causing them to expand their home ranges to maintain access to necessary resources [61, 87]. Coyotes had larger home ranges than bobcats, but a large amount of variation was present within the coyote population [20, 23]. In addition, there was no correlation between human modification within their home ranges and home range size in coyotes. This lack of response is likely due to coyotes adjusting to human modification in their home ranges in other ways, such as spatial choices within their home ranges [19] or temporal adaptations to human activity [18, 79]. The great potential for behavioral plasticity in coyotes [8] could lead to the high levels of individual variation in home range size and adjustments within the home range that we observed.
Coyotes displayed a higher degree of temporal adjustment in their habitat selection than bobcats; their selection responses were more varied depending on the temporal period, both by time of day and by season, than bobcats. We observed bobcat tolerance of human modification and exurban habitat regardless of temporal period, which was unexpected based on previous studies (e.g., [70, 73]). Bobcats could be diluting the human density within their home ranges by expanding their home ranges in response to human modification, becoming less negatively affected by human modification and exurban areas overall. This dilution could be possible as long as human modification is below an animal’s threshold for acceptable modification within their home range [60, 64].
Bobcats and coyotes both adjusted their responses to agriculture, exurban habitat, and water depending on the degree of human modification around them. Bobcat functional responses to human modification in their home ranges were straightforward, selecting more agriculture, less exurban habitat, and areas closer to water as human modification increased. This means that human modification does impact bobcat behavior, causing them to adjust their use of habitat accordingly, which was expected based on other work (e.g., [15]). The directionality of these trends (positive or negative trend in selection with increasing human modification) was consistent when they were present regardless of the temporal period, although the strength of the trend sometimes varied by temporal period. Coyotes had a more varied response to human modification and mean annual responses did not always reflect the same trends present in specific temporal periods, although often trends within the same season were similar. These differences in functional response trends based on temporal period could be due to factors such as differences in prey availability depending on season or time of day in combination with human modification or variation in the availability of and need for cover during different seasons and times of day (e.g., selection for agriculture may increase during summer/fall when crops provide cover to avoid human activity in the presence of high human modification). In addition, regressions were sometimes quadratic and changed direction after a threshold of human modification, indicating coyotes may have more complex functional responses that completely changes response patterns when human modification becomes extremely prevalent in their home range (e.g., > 50% modified). Overall, coyote functional responses to human modification were more nuanced and temporally-dependent than those of bobcats.
Variation of response to anthropogenic change
Taken altogether, these results support the idea that bobcats and coyotes respond differently to human modification in the scale and complexity of their space use behaviors, including home range size, habitat selection, and functional responses. Bobcats exhibited a broad-scale response to human modification. When faced with human modification, bobcats expanded their home ranges and functionally responded in their selection in a predictable manner with little temporal variation and complexity in their habitat selection overall. These results corroborate previous work that shows that bobcats avoid humans (e.g., [71]) and rely on corridors across a development gradient [51, 66]. In contrast, human modification did not affect coyote home range size, but it did cause coyotes to have more temporally-dependent habitat selection behaviors and varied and complex functional responses in their habitat selection, which often changed temporally in intensity or direction. Compared to bobcats, coyotes were able to fine-tune their spatial behavior by avoiding undesirable features on the landscape (e.g., exurban areas) on a finer scale within their home range instead of expanding their range. While it might be unanticipated that a species adapted to coexistence with humans would avoid human modification, this avoidance of human-associated areas [22] is a part of their adjustment strategy. Coyote temporal adjustments have been documented, including changing habitat preferences on a daily scale to avoid risk [65, 74] and on a seasonal scale to exploit seasonal resources [90]. The overall complexity of coyote response to human modification illustrates how a species’ response to novel environments can occur on multiple scales. These responses highlight differences in behavioral plasticity capacity between the two species.
Focusing on multiple aspects of space-use by investigating home range size and habitat selection behaviors including spatial, temporal, and individual variation allowed us to reveal the complexity and differences in the responses of two carnivores to anthropogenic disturbance that moves beyond comparing fine-scale and broad-scale habitat selection. While investigating functional responses in resource selection is common [21, 26], investigating temporal variation in these functional responses is rarely done, yet considering this aspect is critical in understanding the degree of nuance in spatial behavior. However, using these characteristics allowed us to categorize species based on home range size, habitat selection, and functional response behaviors and highlight how a species is responding to anthropogenic change by combining investigations of functional responses in home range size and resource selection with selection behaviors on spatial and temporal scales.
Finding how species or populations adjust their space use behaviors to respond to features in their environment can have important conservation and management implications. As human modification continues, understanding the full extent of its effects on wildlife population dynamics and fitness [89] as a result of individual- and population-level responses is increasingly crucial. Species that are less plastic are more likely to be disadvantaged in high-disturbance environments, while behavioral flexibility leads to increased success and tolerance of anthropogenic environments [45, 46]. For example, bobcat populations in North America only recently began recovering after record lows in the 1900’s [75], while coyote populations have both increased in number and range across North America with anthropogenic land changes and extirpation of large predators [38, 43], illustrating the implications of plasticity and tolerance to human modification. However, while a nuanced response to human modification can provide benefits in exploiting anthropogenic habitat, there are also risks associated with this behavior. Forty-two percent of the coyotes tracked in this study (n = 13) were killed (hunted or trapped) within one year after being collared. While coyote abundance in this population appeared to remain stable despite these mortalities, there remains a risk to individuals living with humans.
Conclusions
The variation in complexity in spatiotemporal response described here could be used as another metric to predict how species will react to future changes, and potentially as to how to best manage them. Rettie and Messier [72] proposed the “hierarchy of limiting factors” hypothesis, stating that species will display space-use response at a broader scale to address their most limiting factors. Similar to this idea, species on the broad-scale end of the spectrum appear to respond to human development by displaying broad spatial response, indicating that habitat itself might be their biggest limiting factor [72]. As such, managing species like bobcats should focus on habitat manipulation to mitigate blanket responses in home range size and habitat selection. Species on the fine-scale end of the spectrum, like coyotes, may respond more to factors impacting the type of interactions with humans, such as harvest management, because they can be more flexible in habitat use and risk avoidance on a temporal scale. In such, our results highlight the importance of investigating spatial, temporal and individual responses to elucidate how other species might be impacted by human activities and how to best mitigate these activities.
Availability of data and materials
Data will be archived upon acceptance in a FigShare repository.
References
Atwood TC, Weeks HP, Gehring TM. Spatial ecology of coyotes along a suburban-to-rural gradient. J Wild Manag. 2004;68(4):1000–9.
Basille M, Fortin D, Dussault C, Ouellet J-P, Courtois R. Ecologically based definition of seasons clarifies predator–prey interactions. Ecography. 2013;36(2):220–9.
Bastille-Rousseau G, Potts JR, Schaefer JA, Lewis MA, Ellington EH, Rayl ND, Mahoney SP, Murray DL. Unveiling trade-offs in resource selection of migratory caribou using a mechanistic movement model of availability. Ecography. 2015;38(10):1049–59.
Bastille-Rousseau G, Schlichting PE, Keiter DA, Smith JB, Kilgo JC, Wittemyer G, Vercauteren KC, Beasley JC, Pepin KM. Multi-level movement response of invasive wild pigs (Sus scrofa) to removal. Pest Manag Sci. 2021;77(1):85–95.
Bastille-Rousseau G, Wittemyer G. Leveraging multidimensional heterogeneity in resource selection to define movement tactics of animals. Ecol Lett. 2019;22(9):1417–27.
Beltrán JF, Tewes ME. Immobilization of ocelots and bobcats with ketamine hydrochloride and xylazine hydrochloride. J Wildl Dis. 1995;31(1):43–8.
Beyer HL, Ung R, Murray DL, Fortin M-J. Functional responses, seasonal variation and thresholds in behavioural responses of moose to road density. J Appl Ecol. 2013;50(2):286–94.
Breck SW, Poessel SA, Mahoney P, Young JK. The intrepid urban coyote: a comparison of bold and exploratory behavior in coyotes from urban and rural environments. Sci Rep. 2019;9(1):2104.
Burnham KP, Anderson DR. Model Selection and multimodel inference: a practical information-theoretic approach. 2nd ed. Berlin: Springer; 2002.
Burt WH. Territoriality and home range concepts as applied to mammals. J Mammal. 1943;24(3):346–52.
Butler Z, Gamba D, Morris S, Zahratka J, Lehmer EM. Novel use of a rapidly reversible field anesthetic in small game mammals using yellow-bellied marmots (Marmota flaviventris) as a model organism. Bios. 2017;88(4):135–43.
Calabrese JM, Fleming CH, Gurarie E. ctmm: an R package for analyzing animal relocation data as a continuous-time stochastic process. Methods Ecol Evol. 2016;7(9):1124–32.
Červinka J, Drahníková L, Kreisinger J, Šálek M. Effect of habitat characteristics on mesocarnivore occurrence in urban environment in the Central Europe. Urban Ecosyst. 2014;17(4):893–909.
Fleming CH, Fagan WF, Mueller T, Olson KA, Leimgruber P, Calabrese JM. Rigorous home range estimation with movement data: a new autocorrelated kernel density estimator. Ecol. 2015;96(5):1182–8.
Flores-Morales M, Vázquez J, Bautista A, Rodríguez-Martínez L, Monroy-Vilchis O. Response of two sympatric carnivores to human disturbances of their habitat: the bobcat and coyote. Mammal Res. 2019;64(1):53–62.
Franckowiak GA, Perdicas M, Smith GA. Spatial ecology of coyotes in the urbanizing landscape of the Cuyahoga Valley, Ohio. PLoS ONE. 2020;14(12):e0227028.
Frey S, Volpe JP, Heim NA, Paczkowski J, Fisher JT. Move to nocturnality not a universal trend in carnivore species on disturbed landscapes. Oikos. 2020;129(8):1128–40.
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS. The influence of human disturbance on wildlife nocturnality. Science. 2018;360(6394):1232–5.
Gehrt SD, Anchor C, White LA. Home range and landscape use of coyotes in a metropolitan landscape: Conflict or coexistence? J Mammal. 2009;90(5):1045–57.
Gese EM, Rongstad OJ, Mytton WR. Home range and habitat use of coyotes in Southeastern Colorado. J Wild Manag. 1988;52(4):640–6.
Godvik IMR, Loe LE, Vik JO, Veiberg V, Langvatn R, Mysterud A. Temporal scales, trade-offs, and functional responses in red deer habitat selection. Ecol. 2009;90(3):699–710.
Gosselink TE, Van Deelen TR, Warner RE, Joselyn MG. Temporal habitat partitioning and spatial use of coyotes and red foxes in east-central Illinois. J Wild Manag. 2003;67(1):90–103.
Grinder MI, Krausman PR. Home range, habitat use, and nocturnal activity of coyotes in an urban environment. J Wild Manag. 2001;65(4):887–98.
Hartigan JA, Wong MA. Algorithm AS 136: a k-means clustering algorithm. J R Stat. 1979;28(1):100–8.
Hebblewhite M, Merrill E. Modelling wildlife–human relationships for social species with mixed-effects resource selection models. J Appl Ecol. 2008;45(3):834–44.
Herfindal I, Tremblay J-P, Hansen BB, Solberg EJ, Heim M, Sæther B-E. Scale dependency and functional response in moose habitat selection. Ecography. 2009;32(5):849–59.
Hertel AG, Niemelä PT, Dingemanse NJ, Mueller T. A guide for studying among-individual behavioral variation from movement data in the wild. Mov Ecol. 2020;8(1):1–18.
Holling CS. The functional response of predators to prey density and its role in mimicry and population regulation. Mem Ent Soc. 1965;97(S45):5–60.
Illinois State Geological Survey Prairie Research Institute. Streams and Shorelines. 2015. Accessed 5 October 2021. https://clearinghouse.isgs.illinois.edu/data/hydrology/streams-and-shorelines.
Illinois Technology Transfer Center. 2020 Statewide Highway. 2020. Accessed 5 October 2021. http://apps.dot.illinois.gov/gist2/.
Jędrzejewski W, Vivas I, Abarca M, Lampo M, Morales LG, Gamarra G, Schmidt K, Hoogesteijn R, Carreño R, Puerto MF, Viloria ÁL, Breitenmoser C, Breitenmoser U. Effect of sex, age, and reproductive status on daily activity levels and activity patterns in jaguars (Panthera onca). Mammal Res. 2021;66(4):531–9.
Johnson CJ, Boyce MS, Case RL, Cluff HD, Gau RJ, Gunn A, Mulders R. Cumulative effects of human developments on arctic wildlife. Wildl Monogr. 2005;160(1):1–36.
Johnson CJ, Parker KL, Heard DC, Gillingham MP. Movement parameters of ungulates and scale-specific responses to the environment. J Anim Ecol. 2002;71(2):225–35.
Johnson DH. The comparison of usage and availability measurements for evaluating resource preference. Ecol. 1980;61(1):65–71.
Kaiser MI, Müller C. What is an animal personality? Biol Philos. 2021;36(1):1.
Kennedy CM, Oakleaf JR, Theobald DM, Baruch-Mordo S, Kiesecker J. Managing the middle: a shift in conservation priorities based on the global human modification gradient. Glob Change Biol. 2019;25(3):811–26.
Knopff AA, Knopff KH, Boyce MS, St-Clair CC. Flexible habitat selection by cougars in response to anthropogenic development. Biol Conserv. 2014;178:136–45.
Laliberte AS, Ripple WJ. Range contractions of North American carnivores and ungulates. Bioscience. 2004;54(2):123–38.
Laundré JW, Keller BL. Home-range size of coyotes: a critical review. J Wild Manag. 1984;48(1):127–39.
Leblond M, Dussault C, Ouellet J-P. Avoidance of roads by large herbivores and its relation to disturbance intensity. J Zool. 2013;289(1):32–40.
Lehman CP, Rumble MA, Rota CT, Bird BJ, Fogarty DT, Millspaugh JJ. Elk resource selection at parturition sites, Black Hills. South Dakota J Wild Manag. 2016;80(3):465–78.
Lesmeister DB, Nielsen CK, Schauber EM, Hellgren EC. Spatial and temporal structure of a mesocarnivore guild in midwestern North America. Wildl Monogr. 2015;191(1):1–61.
Linnell JDC, Strand O. Interference interactions, co-existence and conservation of mammalian carnivores. Divers Distrib. 2000;6(4):169–76.
Litvaitis JA, Harrison DJ. Bobcat–coyote niche relationships during a period of coyote population increase. Can J Zool. 1989;67(5):1180–8.
Lovell C, Li S, Turner J, Carbone C. The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: the results of an intensive camera trap study. Ecol Evol. 2022;12(3):e8746.
Lowry H, Lill A, Wong BBM. Behavioural responses of wildlife to urban environments. Biol Rev. 2013;88(3):537–49.
MacQueen J. Some methods for classification and analysis of multivariate observations. In: Proceedings of the fifth Berkeley symposium on mathematical statististics and probability. 1967; 281–297.
Manly B, McDonald L, Thomas DL, McDonald TL, Erickson WP. Resource selection by animals: statistical design and analysis for field studies. 2nd ed. Berlin: Springer; 2002.
MapLogs. Sunrise Sunset Times of Illinois, USA. 2018. Accessed 5 October 2021. https://sunrise.maplogs.com/illinois_usa.968.html.
Mason G, Burn CC, Dallaire JA, Kroshko J, McDonald Kinkaid H, Jeschke JM. Plastic animals in cages: behavioural flexibility and responses to captivity. Anim Behav. 2013;85(5):1113–26.
Mayer AE, McGreevy TJ Jr, Brown C, Ganoe LS, Gerber BD. Transient persistence of bobcat (Lynx rufus) occurrence throughout a human-dominated landscape. Popul Ecol. 2022;64(4):323–35.
Mayor SJ, Schneider DC, Schaefer JA, Mahoney SP. Habitat selection at multiple scales. Ecoscience. 2009;16(2):238–47.
Moreau G, Fortin D, Couturier S, Duchesne T. Multi-level functional responses for wildlife conservation: the case of threatened caribou in managed boreal forests. J Appl Ecol. 2012;49(3):611–20.
Muhly TB, Semeniuk C, Massolo A, Hickman L, Musiani M. Human activity helps prey win the predator-prey space race. PLoS ONE. 2011;6(3):e17050.
Murtaugh PA. Simplicity and complexity in ecological data analysis. Ecol. 2007;88(1):56–62.
Mysterud A, Ims RA. Functional responses in habitat use: availablity influences relative use in trade-off situations. Ecol. 1998;79(4):1435–41.
National Oceanic and Atmospheric Administration (NOAA). National Weather Service: The Seasons (Equinoxes and Solstices) Page. 2018. Accessed 5 October 2021. https://www.weather.gov/dvn/Climate_Astronomical_Seasons.
National Oceanic and Atmospheric Administration (NOAA). National Centers for Environmental Information: Climate at a Glance. 2021. Accessed 21 September 2021. https://www.ncdc.noaa.gov/cag/county/mapping.
Newediuk L, Prokopenko CM, Wal EV. Individual differences in habitat selection mediate landscape level predictions of a functional response. Oecologia. 2022;198(1):99–110.
Nielsen C, Woolf A. Bobcat habitat use relative to human dwellings in southern Illinois. In: Proceedings of a symposium on current bobcat research and implications for management. The wildlife society 2000 conference. 2001; 40–44.
Nielsen CK, Bottom CR, Tebo RG, Greenspan E. Habitat overlap among bobcats (Lynx rufus), coyotes (Canis latrans), and Wild Turkeys (Meleagris gallopavo) in an agricultural landscape. Can J Zool. 2017;96(5):486–96.
Nilsen EB, John DCL, Andersen R. Individual access to preferred habitat affects fitness components in female roe deer Capreolus capreolus. J Anim Ecol. 2004;73(1):44–50.
O’Donnell K, delBarco-Trillo J. Changes in the home range sizes of terrestrial vertebrates in response to urban disturbance: a meta-analysis. J Urban Ecol. 2020;6(1):1–8.
Ordeñana MA, Crooks KR, Boydston EE, Fisher RN, Lyren LM, Siudyla S, Haas CD, Harris S, Hathaway SA, Turschak GM, Miles AK, Van Vuren DH. Effects of urbanization on carnivore species distribution and richness. J Mammal. 2010;91(6):1322–31.
Petroelje TR, Kautz TM, Beyer DE, Belant JL. Interference competition between wolves and coyotes during variable prey abundance. Ecol Evol. 2021;11(3):1413–31.
Popescu VD, Kenyon M, Brown RK, Dyck MA, Prange S, Peterman WE, Dennison C. Habitat connectivity and resource selection in an expanding bobcat (Lynx rufus) population. PeerJ. 2021;9:e12460.
Randa LA, Yunger JA. Carnivore occurrence along an urban-rural gradient: a landscape-level analysis. J Mammal. 2006;87(6):1154–64.
Réale D, Dingemanse NJ, Kazem AJN, Wright J. Evolutionary and ecological approaches to the study of personality. Philos Trans R Soc. 2010;365(1560):3937–46.
Réale D, Reader SM, Sol D, McDougall PT, Dingemanse NJ. Integrating animal temperament within ecology and evolution. Biol Rev. 2007;82(2):291–318.
Reed GC, Litvaitis JA, Ellingwood M, Tate P, Broman DJA, Sirén APK, Carroll RP. Describing habitat suitability of bobcats (Lynx rufus) using several sources of information obtained at multiple spatial scales. Mamm Biol. 2017;82(1):17–26.
Reilly CM, Suraci JP, Smith JA, Wang Y, Wilmers CC. Mesopredators retain their fear of humans across a development gradient. Behav Ecol. 2022;33(2):428–35.
Rettie WJ, Messier F. Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography. 2000;23(4):466–78.
Riley SPD, Sauvajot RM, Fuller TK, York EC, Kamradt DA, Bromley C, Wayne RK. Effects of urbanization and habitat fragmentation on bobcats and coyotes in southern California. Conserv Biol. 2003;17(2):566–76.
Rivera K, Fidino M, Farris ZJ, Magle SB, Murphy A, Gerber BD. Rethinking habitat occupancy modeling and the role of diel activity in an anthropogenic world. Am Nat. 2022;200(4):556–70.
Roberts NM, Crimmins SM. Bobcat population status and management in North America: evidence of large-scale population increase. J Fish Wildl Manag. 2010;1(2):169–74.
Roemer GW, Gompper ME, Van Valkenburgh B. The ecological role of the mammalian mesocarnivore. Bioscience. 2009;59(2):165–73.
Schielzeth H. Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol. 2010;1(2):103–13.
Schmidt K. Behavioural and spatial adaptation of the Eurasian lynx to a decline in prey availability. Acta Theriol. 2008;53(1):1–16.
Shamoon H, Maor R, Saltz D, Dayan T. Increased mammal nocturnality in agricultural landscapes results in fragmentation due to cascading effects. Biol Conserv. 2018;226:32–41.
Skinner DL, Todd AW. Evaluating efficiency of footholding devices for coyote capture. Wildl Soc Bull. 1990;18(2):166–75.
Snell-Rood EC. An overview of the evolutionary causes and consequences of behavioural plasticity. Anim Behav. 2013;85(5):1004–11.
Spalinger DE, Hobbs NT. Mechanisms of foraging in mammalian herbivores: new models of functional response. Am Nat. 1992;140(2):325–48.
Stamps JA, Groothuis TG. Developmental perspectives on personality: implications for ecological and evolutionary studies of individual differences. Philos Trans R Soc. 2010;365(1560):4029–41.
Streicher JP, Ramesh T, Downs CT. An African urban mesocarnivore: navigating the urban matrix of Durban, South Africa. Glob Ecol Conserv. 2021;26:e01482.
Thornton DH, Sunquist ME, Main MB. Ecological separation within newly sympatric populations of coyotes and bobcats in South-Central Florida. J Mammal. 2004;85(5):973–82.
Tigas LA, Van Vuren DH, Sauvajot RM. Behavioral responses of bobcats and coyotes to habitat fragmentation and corridors in an urban environment. Biol Conserv. 2002;108(3):299–306.
Tucker SA, Clark WR, Gosselink TE. Space use and habitat selection by bobcats in the fragmented landscape of south-central Iowa. J Wild Manag. 2008;72(5):1114–24.
United States Geological Survey (USGS). National Land Cover Database (NLCD) 2019 Land Cover Products. 2021. Accessed 5 December 2021. https://www.sciencebase.gov/catalog/item/604a4fb1d34eb120311b0039.
Webber QMR, Laforge MP, Bonar M, Robitaille AL, Hart C, Zabihi-Seissan S, Vander WE. The ecology of individual differences empirically applied to space-use and movement tactics. Am Nat. 2020;196(1):E1–15.
Webster SC, Beasley JC, Hinton JW, Chamberlain MJ. Resident and transient coyotes exhibit differential patterns of movement behavior across heterogeneous landscapes in the southeastern United States. Ecol Evol. 2022;12(3):e8725.
Worton BJ. Kernel methods for estimating the utilization distribution in home-range studies. Ecol. 1989;70(1):164–8.
Yan M, Ye K. Determining the number of clusters using the weighted gap statistic. Biometrics. 2007;63(4):1031–7.
Acknowledgements
Funding for this research was provided through Federal-aid Wildlife Restoration Act grants W87R. We thank the staffs at IDNR, U.S. Army Corps of Engineers, Touch of Nature, and Giant City State Park for their use of land, housing, and resources during fieldwork. We thank Brooks Pitman, Michele Lovara, Lane Jeakle, Sujay Singh, Blake Hoffman, Chris Hickling, and Cody Wertman for their work during bobcat and coyote captures. We also thank Clay Nielsen, Seth Riley, and an anonymous reviewer for comments on an earlier version of this paper and Michael Egan, Storm Crews, and other members of the SIU Cooperative Wildlife Research Lab for valuable input throughout the execution of this project.
Funding
This work was supported through Federal-aid Wildlife Restoration Act Grant W87R42-46 via Illinois Department of Natural Resources.
Author information
Authors and Affiliations
Contributions
NTG, MWE, DJS, PES, and GBR developed the idea and structure of the manuscript. MWE, DJS, PES, and GBR secured funding for animal tracking. NTG and GBR led the writing of the manuscript with input from all co-authors.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
Animal capture and handling was done in accordance with a pre-approved animal ethics protocol (SIU Animal Care and Use Protocol 19-002 and 21-028).
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gorman, N.T., Eichholz, M.W., Skinner, D.J. et al. Carnivore space use behaviors reveal variation in responses to human land modification. Mov Ecol 12, 51 (2024). https://doi.org/10.1186/s40462-024-00493-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-024-00493-7